Белки: история исследования, химический состав, свойства, биологические функции. Какие элементы входят в состав белков и какими свойствами они обладают? Что входит в состав белков
Химический состав белков.
3.1. Пептидная связь
Белки представляют собой нерегулярные полимеры, построенные из остатков -аминокислот, общую формулу которых в водном растворе при значениях pH близких к нейтральным можно записать как NH 3 + CHRCOO – . Остатки аминокислот в белках соединены между собой амидной связью между -амино- и -карбоксильными группами. Пептидная связь между двумя -аминокислотными остатками обычно называется пептидной связью , а полимеры, построенные из остатков -аминокислот, соединенных пептидными связями, называют полипептидами. Белок как биологически значимая структура может представлять собой как один полипептид, так и несколько полипептидов, образующих в результате нековалентных взаимодействий единый комплекс.
3.2. Элементный состав белков
Изучая химический состав белков, необходимо выяснить, во-первых, из каких химических элементов они состоят, во-вторых, - строение их мономеров. Для ответа на первый вопрос определяют количественный и качественный состав химических элементов белка. Химический анализ показал наличие во всех белках углерода (50-55%), кислорода (21-23%), азота (15-17%), водорода (6-7%), серы (0,3-2,5%). В составе отдельных белков обнаружены также фосфор, йод, железо, медь и некоторые другие макро- и микроэлементы, в различных, часто очень малых количествах.
Содержание основных химических элементов в белках может различаться, за исключением азота, концентрация которого характеризуется наибольшим постоянством и в среднем составляет 16%. Кроме того, содержание азота в других органических веществах мало. В соответствии с этим было предложено определять количество белка по входящему в его состав азоту. Зная, что 1г азота содержится в 6,25 г белка, найденное количество азота умножают коэффициент 6,25 и получают количество белка.
Для определения химической природы мономеров белка необходимо решить две задачи: разделить белок на мономеры и выяснить их химический состав. Расщепление белка на его составные части достигается с помощью гидролиза – длительного кипячения белка с сильными минеральными кислотами (кислотный гидролиз) или основаниями (щелочной гидролиз) . Наиболее часто применяется кипячение при 110 С с HCl в течение 24 ч. На следующем этапе разделяют вещества, входящие в состав гидролизата. Для этой цели применяют различные методы, чаще всего – хроматографию (подробнее – глава “Методы исследования…”). Главным частью разделенных гидролизатов оказываются аминокислоты.
3.3. Аминокислоты
В настоящее время в различных объектах живой природы обнаружено до 200 различных аминокислот. В организме человека их, например, около 60. Однако в состав белков входят только 20 аминокислот, называемых иногда природными.
Аминокислоты – это органические кислоты, у которых атом водорода -углеродного атома замещен на аминогруппу – NH 2 . Следовательно, по химической природе это -аминокислоты с общей формулой:
H – C – NH 2
Из этой формулы видно, что в состав всех аминокислот входят следующие общие группировки: – CH 2 , – NH 2 , – COOH. Боковые же цепи (радикалы – R ) аминокислот различаются. Как видно из Приложения I химическая природа радикалов разнообразна: от атома водорода до циклических соединений. Именно радикалы определяют структурные и функциональные особенности аминокислот.
Все аминокислоты, кроме простейшей аминоуксусной к-ты глицина (NH 3 + CH 2 COO ) имеют хиральный атом C и могут существовать в виде двух энантиомеров (оптических изомеров):
COO – COO –
NH 3 + R R NH 3 +
L -изомер D -изомер
В состав всех изученных в настоящее время белков входят только аминокислоты L-ряда, у которых, если рассматривать хиральный атом со стороны атома H, группы NH 3 + , COO и радикал R расположены по часовой стрелке. Необходимость при построении биологически значимой полимерной молекулы строить ее из строго определенного энантиомера очевидна – из рацемической смеси двух энантиомеров получилась бы невообразимо сложная смесь диастереоизомеров. Вопрос, почему жизнь на Земле основана на белках, построеных именно из L-, а не D--аминокислот, до сих пор остается интригующей загадкой. Следует отметить, что D-аминокислоты достаточно широко распространены в живой природе и, более того, входят в состав биологически значимых олигопептидов.
Из двадцати основных -аминокислот строятся белки, однако остальные, достаточно разнообразные аминокислоты образуются из этих 20 аминокислотных остатков уже в составе белковой молекулы. Среди таких превращений следует в первую очередь отметить образование дисульфидных мостиков при окислении двух остатков цистеина в составе уже сформированных пептидных цепей. В результате образуется из двух остатков цистеина остаток диаминодикарбоновой кислоты цистина (см. Приложение I). При этом возникает сшивка либо внутри одной полипептидной цепи, либо между двумя различными цепями. В качестве небольшого белка, имеющего две полипептидные цепи, соединенный дисульфидными мостиками, а также сшивки внутри одной из полипептидных цепей:
GIVEQCCA SVCSLY QLENYCN
FVNQHLC GSHLVEALYLVC GERGFFYTPKA
Важным примером модификации аминокислотных остатков является превращение остатков пролина в остатки гидроксипролина :
N – CH – CO – N – CH – CO –
CH 2 CH 2 CH 2 CH 2
CH 2 CHOH
Это превращение происходит, причем в значительном масштабе, при образовании важного белкового компонента соединительной ткани – коллагена .
Еще одним весьма важным видом модификации белков является фосфорилирование гидроксогрупп остатков серина, треонина и тирозина, например:
– NH – CH – CO – – NH – CH – CO –
CH 2 OH CH 2 OPO 3 2 –
Аминокислоты в водном растворе находятся в ионизированном состоянии за счет диссоциации амино- и карбоксильных групп, входящих в состав радикалов. Другими словами, они являются амфотерными соединениями и могут существовать либо как кислоты (доноры протонов), либо как основания (акцепторы доноров).
Все аминокислоты в зависимости от структуры разделены на несколько групп:
Ациклические . Моноаминомонокарбоновые аминокислоты имеют в своем составе одну аминную и одну карбоксильную группы, в водном растворе они нейтральны. Некоторые из них имеют общие структурные особенности, что позволяет рассматривать их вместе:
Глицин и аланин. Глицин (гликокол или аминоуксусная к-та) является оптически неактивным – это единственная аминокислота, не имеющая энатиомеров. Глицин участвует в образовании нуклеиновых и желчных к-т, гема, необходим для обезвреживания в печени токсичных продуктов. Аланин используется организмом в различных процессах обмена углеводов и энергии. Его изомер -аланин является составной частью витамина пантотеновой к-ты, коэнзима А (КоА), экстрактивных веществ мышц.
Серин и треонин. Они относятся к группе гидрооксикислот, т.к. имеют гидроксильную группу. Серин входит в состав различных ферментов, основного белка молока – казеина, а также в состав многих липопротеинов. Треонин участвует в биосинтезе белка, являясь незаменимой аминокислотой.
Цистеин и метионин. Аминокислоты, имеющие в составе атом серы. Значение цистеина определяется наличием в ее составе сульфгидрильной (– SH) группы, которая придает ему способность легко окисляться и защищать организм о веществ с высокой окислительной способностью (при лучевом поражении, отравлении фосфором). Метионин характеризуется наличием легко подвижной метильной группы, использующейся для синтеза важных соединений в организме (холина, креатина, тимина, адреналина и др.)
Валин, лейцин и изолейцин. Представляют собой разветвленные аминокислоты, которые активно участвуют в обмене веществ и не синтезируются в организме.
Моноаминодикарбоновые аминокислоты имеют одну аминную и две карбоксильные группы и в водном растворе дают кислую реакцию. К ним относятся аспарагиновая и глутаминовая к-ты, аспарагин и глутамин. Они входят в состав тормозных медиаторов нервной системы.
Диаминомонокарбоновые аминокислоты в водном растворе имеют щелочную реакцию за сет наличия двух аминных групп. Относящийся к ним лизин необходим для синтеза гистонов а также в ряд ферментов. Аргинин участвует в синтезе мочевины,креатина.
Циклические . Эти аминокислоты имеют в своем составе ароматическое или гетероциклическое ядро и, как правило, не синтезируется в организме человека и должны поступать с пищей. Они активно участвуют в разнообразных обменных процессах. Так
фенил-аланин служит основным источником синтеза тирозина – предшественника ряда биологически важных веществ: гормонов (тироксина, адреналина), некоторых пигментов. Триптофан помимо участия в синтезе белка, служит компонентом витамина PP, серотонина, триптамина, ряда пигментов. Гистидин необходим для синтеза белков, является предшественником гистамина, влияющего на кровяное давление и секрецию желудочного сока.
Свойства
Белки являются высокомолекулярными соединениями. Это полимеры, состоящие из сотен и тысяч аминокислотных остатков – мономеров.
Белки имеют высокую молекулярную массу, некоторые растворимы в воде, способны к набуханию, характеризуются оптической активностью, подвижностью в электрическом поле и некоторыми другими свойствами.
Белки активно вступают в химические реакции. Это свойство связано с тем, что аминокислоты, входящие в состав белков, содержат разные функциональные группы, способные реагировать с другими веществами. Важно, что такие взаимодействия происходят и внутри белковой молекулы, в результате чего образуется пептидная, водородная дисульфидная и другие виды связей. К радикалам аминокислот, а Соответственно и молекулярная масса белков находится в пределах 10 000 – 1 000 000. Так, в составе рибонуклеазы (фермента, расщепляющего РНК) содержится 124 аминокислотных остатка и ее молекулярная масса составляет примерно 14 000. Миоглобин (белок мышц), состоящий из 153 аминокислотных остатков, имеет молекулярную массу 17 000, а гемоглобин – 64 500 (574 аминокислотных остатка). Молекулярные массы других белков более высокие: -глобулин (образует антитела) состоит из 1250 аминокислот и имеет молекулярную массу около 150 000, а молекулярная масса фермента глутаматдегидрогеназы превышает 1 000 000.
Определение молекулярной массы проводится различными методами: осмометрическим, гельфильтрационным, оптическим и др. однако наиболее точным является метод седиментации, предложенный Т. Сведбергом. Он основан на том, что при ультрацентрифугировании ускорением до 900 000 g скорость осаждения белков зависит от их молекулярной массы.
Важнейшим свойством белков является их способность проявлять как кислые так и основные, то есть выступать в роли амфотерных электролитов. Это обеспечивается за счет различных диссоциирующих группировок, входящих в состав радикалов аминокислот. Например, кислотные свойства белку придают карбоксильные группы аспарагиновой глутаминовой аминокислот, а щелочные – радикалы аргинина, лизина и гистидина. Чем больше дикарбоновых аминокислот содержится в белке, тем сильнее проявляются его кислотные свойства и наоборот.
Эти же группировки имеют и электрические заряды, формирующие общий заряд белковой молекулы. В белках, где преобладают аспарагиновая и глутаминовая аминокислоты, заряд белка будет отрицательным, избыток основных аминокислот придает положительный заряд белковой молекуле. Вследствие этого в электрическом поле белки будут передвигаться к катоду или аноду в зависимости от величины их общего заряда. Так, в щелочной среде (рН 7 – 14) белок отдает протон и заряжается отрицательно, тогда как в кислой среде (рН 1 – 7) подавляется диссоциация кислотных групп и белок становится катионом.
Таким образом, фактором, определяющим поведение белка как катиона или аниона, является реакция среды, которая определяется концентрацией водородных ионов и выражается величиной рН. Однако при определенных значениях рН число положительных и отрицательных зарядов уравнивается и молекула становится электронейтральной, т.е. она не будет перемещаться в электрическом поле. Такое значение рН среды определяется как изоэлектрическая точка белков. При этом белок находится в наименее устойчивом состоянии и при незначительных изменениях рН в кислую или щелочную сторону легко выпадает в осадок. Для большинства природных белков изоэлектрическая точка находится в слабокислой среде (рН 4,8 – 5,4), что свидетельствует о преобладании в их составе дикарбоновых аминокислот.
Свойство амфотерности лежит в основе буферных свойств белков и их участии в регуляции рН крови. Величина рН крови человека отличается постоянством и находится в пределах 7,36 – 7,4 , несмотря на различные вещества кислого или основного характера, регулярно поступающие с пищей или образующиеся в обменных процессах – следовательно существуют специальные механизмы регуляции кислотно-щелочного равновесия внутренней среды организма. К таким системам относится рассматриваемая в гл. “ Классификация” гемоглобиновая буферная система (стр.28). Изменение рН крови более чем на 0,07 свидетельствует о развитии патологического процесса. Сдвиг рН в кислую сторону называется ацидозом, а в щелочную – алкалозом.
Важное значение для организма имеет способность белков адсорбироватьь на своей поверхности некоторые вещества и ионы (гормоны, витамины, железо, медь), которые либо плохо растворимы в воде, либо являются токсичными (билирубин, свободные жирные кислоты). Белки транспортируют их по крови к местам дальнейших превращений или обезвреживания.
Водные растворы белков имеют свои особенности. Во-первых, белки обладают большим сродством к воде, т.е. они гидрофильны. Это значит, что молекулы белка, как заряженные частицы, притягивают к себе диполи воды, которые располагаются вокруг белковой молекулы и образуют водную или гидратную оболочку. Эта оболочка предохраняет молекулы белка от склеивания и выпадения в осадок. Величина гидратной оболочки зависит от структуры белка. Например, альбумины более легко связываются с молекулами воды и имеют относительно большую водную оболочку, тогда как глобулины, фибриноген присоединяют воду хуже, и гидратная оболочка и них меньше. Таким образом, устойчивость водного раствора белка определяется двумя факторами: наличием заряда белковой молекулы и находящейся вокруг нее водной оболочки. При удалении этих факторов белок выпадает в осадок. Данный процесс может быть обратимым и необратимым.
Обратимое осаждение белков (высаливание) предполагает выпадение белка в осадок под действием определенных веществ, после удаления которых он вновь возвращается в свое исходное (нативное) состояние. Для высаливания белков используют соли щелочных и щелочноземельных металлов (наиболее часто в практике используют сульфат натрия и аммония). Эти соли удаляют водную оболочку (вызывают обезвоживание) и снимают заряд. Между величиной водной оболочки белковых молекул и концентрацией солей существует прямая зависимость: чем меньше гидратная оболочка, тем меньше требуется солей. Так, глобулины, имеющие крупные и тяжелые молекулы и небольшую водную оболочку, выпадают в осадок при неполном насыщении раствора солями, а альбумины как более мелкие молекулы, окруженные большой водной оболочкой, – при полном насыщении.
Нативная молекула белка
Денатурированная молекула белка. Черточки обозначают связи в молекуле нативного белка, разрывающиеся при денатурации


еобратимое осаждение связано с глубокими внутримолекулярными изменениями структуры белка, что приводит в потере ими нативных свойств (растворимости, биологической активности и др.). Такой белок называется денатурированным, а процесс денатурацией . Денатурация белков происходит в желудке, где имеется сильнокислая среда (рН 0,5 – 1,5), и это способствует расщеплению белков протеолитическими ферментами. Денатурация белков положена в основу лечения отравления тяжелыми металлами, когда больному вводят per os (“через рот”) молоко или сырые яйца с тем, чтобы металлы денатурируя белки молока или яиц.
Адсорбировались на их поверхности и не действовали на белки слизистой оболочки желудка и кишечника, а также не всасывались в кровь.
Размер белковых молекул лежит в пределах 1 мкм до 1 нм и, следовательно, они являются коллоидными частицами, которые в воде образуют коллоидные растворы. Эти растворы характеризуются высокой вязкостью, способностью рассеивать лучи видимого света, не проходят сквозь полупроницаемые мембраны.
Вязкость раствора зависит от молекулярной массы и концентрации растворенного вещества. Чем выше молекулярная масса, тем раствор более вязкий. Белки как высокомолекулярные соединения образуют вязкие растворы. Например, раствор яичного белка в воде.
Вода

оллоидные частицы не проходят через полупроницаемые мембраны (целлофан, коллоидную пленку), так как их поры меньше коллоидных частиц. Непроницаемыми для белка являются все биологические мембраны. Это свойство белковых растворов широко используется в медицине и химии для очистки белковых препаратов от посторонних примесей. Такой процесс разделения называется диализом. Явление диализа лежит в основе действия аппарата “искусственная почка”, который широко используется в медицине для лечения острой почечной недостаточности.
Диализ (белые крупные кружки – молекулы белка, черные – молекулы хлористого натрия)
Минеральные вещества молока
В золе молока содержатся такие минеральные вещества, как кальций, фосфор, магний, калий, натрий, хлор, сера, кремний. Количество отдельных элементов в молоке определяется в основном генетическими факторами. Кормление и другие факторы окружающей среды оказывают на их содержание лишь незначительное влияние. Количество минеральных веществ в молоке остается постоянным даже тогда, когда в рационах отдельных элементов мало. При недостаточном поступлении минеральных веществ с кормом мобилизуются резервы организма и таким образом концентрация их в молоке поддерживается на определенном уровне. При значительном недостатке одного или нескольких элементов содержание минеральных веществ в расчете на единицу объема молока остается более или менее постоянным. Однако молочная продуктивность, а затем и общее количество минеральных веществ в молоке снижается.
|
Минеральные вещества |
Содержится,г |
Минеральные вещества |
|
Общее количество микроэлементов в молоке составляет менее 0,15%.Содержание микроэлементов в молоке находится в тесной зависимости от наличия их в кормах.
Структурно-механические
свойства масла.
По
Ребиндеру существуют два основных типа
структур.
Первый тип – коагуляционная структура – это пространственные сетки, возникающие путем беспорядочного сцепления мельчайших частиц дисперсной фазы или микромолекул через тонкие расслойки данной среды.
Второй тип – это кристаллизационно-конденсационная структура , образующаяся в результате непосредственного срастания кристалликов с образованием при этом поликристаллического твердого тела.
Жировые основы маргарина относятся к коагуляционному типу структур. Консистенция и пластические свойства жировых основ маргарина в основном определяются соотношением твердой и жидкой фаз в том или ином пищевом жире. Это соотношение твердой и жидкой фаз характерно для каких-то определенных условий кристаллизации (температура, время, перемешивание). При этом важное значение имеет состав непрерывной среды и дисперсной фазы и характер размещения дисперсной фазы в непрерывной жидкой среде.
Для некоторых видов пищевого жира при определенной температуре и условиях кристаллизации количество твердой дисперсной фазы может выйти за предел оптимального соотношения фаз, и тогда на поверхности кристаллов образуются столь тонкие пленки непрерывной жидкой среды, что они не могут мешать массовому хаотическому сращиванию кристаллов друг с другом. В этом случае мы всегда будем иметь наибольшую твердость жировой основы, крошливую консистенцию и наихудшие пластические свойства.
Если при комнатной температуре пленки жидкой непрерывной среды являются оптимальными по толщине, т.е. такими, которые не создают условий для сращивания кристаллов при хранении, при механическом или термическом воздействии на систему, то в этом идеальном случае мы всегда будем получать упрочненные коагуляционные структуры, которые и определяют наилучшие пластические свойства жировых основ.
Чтобы получать упрочненные коагуляционные структуры, обладающие наилучшими пластическими свойствами, за рубежом часто вводят в рецептуру жировой основы два вида саломаса с температурой плавления 32°С и 42°С. При этом вводится довольно значительное количество жидких растительных масел. Указанное, с одной стороны, создает в жировой основе наилучшие соотношения твердой и жидкой фаз, обеспечивая консистенцию, сходную со сливочным маслом, а с другой стороны, создает условия для постоянства консистенции маргарина в довольно большом интервале температур. Наряду с этим, введение в жировую основу высокоплавких саломасов находится в противоречии с требованиями физиологов к составу пищевых жиров.
Прежде всего, следует отметить, что только наличие высокоэффективных эмульгаторов-стабилизаторов позволило создать современную технологию в производстве маргарина и обеспечить выработку пищевого жирового продукта высокого качества. Поверхностно-активные добавки обеспечивают получение тонкодисперсной эмульсии в прочную связь частиц дисперсной фазы с непрерывной средой (твердым при комнатной температуре жиром). Основной вопрос в производстве маргарина – это влияние поверхностно-активных добавок на структурно-механические свойства маргарина, и в частности на способность к солюбилизации.
Адсорбционный слой эмульгатора повышает устойчивость эмульсии, в особенности в тех случаях, когда этот слой структурируется, образуя пленку поверхностного геля с сильно повышенной вязкостью и прочностью.
Эти свойства имеют особое значение для производства маргарина, поскольку готовый продукт представляет собой эмульсию мельчайших частиц жидкой фазы, равномерно размещенных в непрерывной среде твердой фазы при комнатной температуре.
С проблемой прочности эмульсий тесно связан вопрос о типе образующихся с данным эмульгатором эмульсий. Существует возможность образования двух типов. Значение соотношения объемов фаз для определенного типа образующейся эмульсии объясняется тем, что коалесценция и расслоение эмульсии данного типа происходят тем интенсивнее, чем меньше объем дисперсионной среды и чем больше – дисперсной фазы. Если эмульгатор обеспечивает устойчивую эмульсию только одного типа, то соотношение объемов перестает иметь решающее значение в определении типа эмульсии. Инверсия зависит не только от соотношения объемов фаз, но и от концентрации и химической природы эмульгатора.
Эмульгаторы должны обладать следующими свойствами:
Уменьшать поверхностное натяжение;
-
достаточно быстро адсорбироваться на
поверхности раздела фаз, препятствуя
слиянию капель;
- иметь специфическую
молекулярную структуру с полярными и
неполярными группами;
- влиять на
вязкость эмульсии.
Эффективность действия эмульгатора является специфическим свойством, зависящим от его природы, типа эмульгируемых веществ, температуры, рН среды, концентрации, времени эмульгирования и т.д.
Эффективность действия и природа эмульгатора определяют тип эмульсии.
Гидрофильные эмульгаторы, лучше растворимые в воде, чем в углеводородах, способствуют образованию эмульсий типа масло – вода, а гидрофобные, лучше растворимые в углеводородах, – эмульсий типа вода – масло. Соотношение размеров полярной и неполярной частей молекул эмульгатора характеризуется специальным показателем – гидрофильно-липофильный баланс. Если ГЛБ эмульгатора составляет 3-6, образуется эмульсия вода – масло, при значении ГЛБ 8-13 образуется преимущественно эмульсия типа масло – вода.
Маргарин представляет собой переохлажденную эмульсию типа вода в масле. При этом не исключена возможность образования эмульсии смешанного типа с преобладанием эмульсии вода – масло.
Основные функции эмульгаторов:
Создание устойчивой высокодисперсной
эмульсии;
- стабилизация и предотвращение
отделения влаги и жира в готовом продукте;
- обеспечение стабильности при
хранении;
- обеспечение антиразбрызгивающей
способности при жарке;
- обеспечение
пластичности;
- обеспечение создания
устойчивой формы кристаллической
решетки в процессе структурообразования;
- обеспечение заданных функциональных
свойств готового продукта в зависимости
от области использования маргарина.
В Украине на протяжении многих лет использовались эмульгаторы, производимые в России, и собственного производства, вырабатываемые на полупромышленных производствах. К ним относятся эмульгаторы:
Т-1 – продукт глицеролиза говяжьего
жира или саломаса;
- Т-2 – продукт
полимеризации глицерина, этерифицированный
стеариновой кислотой;
- Т-Ф – смесь
эмульгатора Т-1 и пищевого фосфатидного
концентрата в соотношении 2:1;
- ПМД
– пищевые монодиглицериды;
- КЭ –
комбинированный эмульгатор – смесь
ПМД и фосфатидного концентрата в
соотношении 3:1.
Широкая гамма эмульгаторов Нижегородского завода – различные виды дистиллированных моноглицеридов. В настоящее время в Нижнем Новгороде освоено производство серии новых эмульгаторов на основе лецитина. Это лецитины стандартные, лецитины фракционированные – фосфадитилхолин и фосфадитилсерин, а также гидролизованные лецитины.
В последние годы в Украине преимущественно используются эмульгаторы различных модификаций серии Dimodan, Palsgaard (на некоторых предприятиях Квест).
В разные периоды преимущество в спросе на эти два вида эмульгаторов переходило от одного к другому. Можно сказать, что здесь имеет место конкуренция качество – цена.
В зависимости от жирности маргарина и сферы его применения используют эмульгаторы Dimodan PVP (Dimodan HP), Dimodan ОТ (Dimodan S-T PEL/B), Dimodan СР. Для маргаринов жирностью ниже 40%, которые в настоящее время пользуются спросом у населения, используют дополнительно (кроме Dimodan ОТ, или Dimodan СР., или Dimodan LS) эфиры полиглицерина и рицинолевой кислоты – Grinsted PGPR90.
При производстве низкожирных маргаринов, особенно с содержанием жира 25% и ниже, используют стабилизирующие системы – гидроколлоиды (альгинаты, пектины и др.).
Следует отметить, что фирмы-производители дают рекомендации по применению различных видов эмульгаторов и стабилизирующих систем в зависимости от назначения маргаринов. Соблюдение этих рекомендаций позволяет получить продукцию высокого качества
Мышечные белки
Мясо птицы содержит примерно 20-23% белков. Мышечные белки по их растворимости можно разделить на три группы: миофибриллярные, саркоплазматические и белки стромы.
Миофибриллярные , или солерастворимые белки нерастворимы в воде, но большинство растворяется в растворах поваренной соли концентрацией более 1%. Эта группа состоит примерно из 20 отдельных белков, входящих в состав миофибрилл сократительной мышцы. Миофибриллярные белки могут быть разделены на три группы в зависимости от выполняемой функции: сократительные, которые ответственны за мышечные сокращения, регуляторные, участвующие в управлении процессом сокращения, и цитоскелетные, скрепляющие миофибриллы и способствующие сохранению их структурной целостности.
Сократительные белки миозин и актин оказывают большое влияние на функциональность мышечного белка. Поскольку в окоченевшей мышце актин и миозин находятся в виде актомиозинового комплекса, изменяется функциональность миозина как в эмульгированных, так и в формованных продуктах из мяса птицы. Свойства продуктов зависят также от общего соотношения актина и миозина и соотношения миозина и актина в свободном состоянии. Саркоплазматические белки и белки стромы, в свою очередь, влияют на функциональные свойства миофибриллярных белков.
Саркоплазматические белки растворимы в воде или в растворах с малой ионной силой (
Белки стромы , часто называемые белками соединительной ткани, служат каркасом, поддерживающим структуру мышцы. Основным белком стромы является коллаген. Эластин и ретикулин составляют небольшую часть стромы. Все эти белки нерастворимы в воде и солевых растворах. Нежность мяса, как правило, уменьшается с увеличением возраста животных благодаря образованию поперечных связей и другим изменениям коллагена.
Кровь и её фракции
Цельную кровь применяют как основное сырьё для производства колбас, зельцев, консервов и других продуктов питания, а также в качестве аддитива, придающего традиционный цвет изделиям при использовании в них белковых препаратов (0,6-1,0%); с этой же целью применяют препарат гемоглобина или смесь форменных элементов после гидратации в воде (1:1).
По сравнению с другими видами белоксодержащего сырья цельная кровь используется недостаточной широко вследствие наличия специфических цвета и вкуса, модифицирующих органолептические характеристики готовых изделий. В настоящее время ведутся исследования по осветлению крови, однако по ряду причин предложенные способы не нашли практического применения в промышленности. Функционально-технологические свойства крови и её фракций (плазмы, сыворотки) в первую очередь зависят от их белкового состава. Цельная кровь содержит около 150 протеинов с различными физико-химическими свойствами, преобладающими из которых являются белки форменных элементов, альбумины, глобулины и фибриноген. В связи с этим на базе цельной крови целесообразно готовить эмульсии, предназначенные для введения в рецептуры мясопродуктов и обеспечивающие повышение стабильности мясных систем, пищевой ценности и выхода, улучшение органолептических показателей и структурно-механических свойств.
В качестве белкового препарата наиболее целесообразно применять соевый изолят либо казеинат натрия.
Уровень введения эмульсий, приготовленных на основе цельной крови, в мясные системы может составлять до 30-40% к массе основного сырья.
Белки плазмы крови обладают уникальным комплексом ФТС. Альбумины легко взаимодействуют с другими белками, могут быть связаны с липидами и углеводами, имеют высокую водосвязывающую и пенообразующую способность.
Глобулины - хорошие эмульгаторы.
Фибриноген - имеет выраженную гелеобразующую способность, переходя в фибрин под воздействием ряда факторов (сдвиг рН к изоточке, введение ионов Са++ в плазму) и образуя пространственный каркас.
смесей Эти свойства фибриногена можно использовать при получении многокомпонентных белоксодержащих, включающих ПК, гелеподобных текстуратов, в процессе вторичного структурообразования мясных эмульсий при производстве вареных колбасных изделий.
Все белки плазмы характеризуются хорошей растворимостью, и как следствие - высокой водосвязывающей и эмульгирующей способностью, способны образовывать гели при нагревании. Введение поваренной соли оказывает отрицательное влияние на стабильность эмульсий на базе плазмы крови при рН 7,0. Важнейшим свойством плазмы является её способность к образованию гелей при тепловой обработке, причем их прочность и уровень водосвязывающей способности зависит от концентрации белков в системе, величины рН, присутствия солей, температуры и продолжительности нагрева.
Введение в плазму неплазменных белков (яичный альбумин, соевый изолят, казеинат натрия) существенно увеличивает как прочность гелей, так и их водо- и жиропоглощающую способность после термообработки.
В зависимости от состояния плазмы крови и условий первичной обработки, состав и функционально-технологические свойства её и, соответственно, область использования могут изменяться.
Систематизация имеющихся в настоящее время данных по переработке ПК позволяет оценить современные подходы к реализации биологического и функционально-технологического потенциала белкового компонента ПК при производстве пищевых продуктов.
Схема дает представление о состоянии, способах обработки, составе и свойствах белковых препаратов, получаемых на основе ПК, определяет области их практического использования, причем полифункциональность целевого назначения ПК отражена в формируемых при том или ином способе обработки ФТС.
Необходимо отметить, что уровень отдельных показателей ФТС, приведенных в Таблице 13 и служащих для расшифровки условных обозначений, принятых в схеме, является относительным в связи с тем, что фактическая величина каждой характеристики решающим образом зависит от концентрации белка, значения рН в системе, температуры среды, ионной силы и ряда других факторов.
Анализ классификационной схемы показывает, что одним из путей технологического использования плазмы крови является её применение в жидком стабилизированном виде (а также после охлаждения и замораживания) с относительно невысоким содержанием белка и сохраненными нативными ФТС.
В этом случае белки ПК характеризуются высоким уровнем ВСС и эмульгирования, что обусловлено наличием в ней водорастворимых белков, способных образовывать гели при нагреве. Совокупность этих свойств позволяет широко использовать плазму не только как компонент, балансирующий общий химический состав готовых изделий, но и как функциональную добавку при производстве эмульгированных мясопродуктов с высоким конечным влагосодержанием: вареных колбас, сосисок, сарделек, рубленых полуфабрикатов, фаршевых консервов, ветчинных изделий. Наиболее рациональным является введение в рецептуры 10% плазмы взамен 3% говядины или 2% свинины; введение 20% ПК вместо воды при куттеровании обеспечивает улучшение органолептических, структурно-механических показателей и повышение выхода готовой продукции на 0,3-0,5%. Прекрасный эффект дает применение плазмы крови в качестве среды для гидратации белковых препаратов (3-4 частей ПК на 1 часть белкового препарата).
Незаменима ПК при изготовлении белково-жировых эмульсий, связующих, многокомпонентных белковых систем с заданным составом и функционально-технологическими свойствами, структурированных белковых препаратов.
Концентрирование ПК методами сушки, ультрафильтрации и криоконцентрирования, позволяя существенно повысить содержание белка, приводит к некоторой модификации ФТС препарата.
Особенно существенное влияние на степень изменения ФТС оказывает сушка плазмы, в то время как сухой концентрат ПК, подвергнутый ультрафильтрации, имеет весьма высокие функциональные свойства.
Полученные данными методами концентраты успешно применяют при производстве мясопродуктов наряду с жидкой ПК.
Американские специалисты считают, что плазмой" крови крупного рогатого скота, благодаря её ФТС, можно успешно заменять яичный белок.
Денатурационно-коагуляционное осаждение, обеспечивая совмещение процессов термотропного структурирования, флокуляции (осаждения) и концентрирования белков ПК, дает возможность получать препараты с относительно высокой концентрацией белка и неординарными ФТС, что позволяет использовать их в рецептурах полукопченых, копченозапеченых, ливерных колбас, паштетных консервов и полуфабрикатов, имеющих ограниченное конечное влагосодержание и высокую жиропоглотительную способность. К этой группе препаратов относят: "осажденный белок плазмы", "белковые плазменные преципитаты", ливексы, "плазменный сыр", гранулированную ПК.
Применение данных видов препаратов плазмы крови в практике мясного производства весьма ограничено.
Структурирование плазмы крови путем рекальцинирования существенно расширяет возможности её технологического использования. Перевод ПК и многокомпонентных систем на её основе в гель-форму позволяет получать структурные матрицы, имитирующие природные биообъекты по внешнему виду, составу и свойствам, создает предпосылки к регулированию ФТС, обеспечивает вовлечение в процесс производства низкосортного сырья, дает возможность с новых позиций подойти к решению вопроса разработки новых видов пищевых продуктов. Особенно эффективно комплексное использование ПК и белковых препаратов (соевые изоляты, казеинат натрия и т. п.).Структурированные формы ПК применяют при производстве вареных колбас, рубленых полуфабрикатов, ветчины в оболочке, полукопченых и ливерных колбас, паштетов, фаршевых консервов, текстурированных наполнителей рецептур, аналогов мясопродуктов.
СОЗРЕВАНИЕ МЯСА
Вопрос «созревания мяса» до сего времени не получил окончательного освещения. Из наблюдений практиков известно, что после прекращения жизни животного в мясе происходят физико-химические изменения, характеризующиеся окоченением, затем расслаблением (размягчением) мышечных волокон. В результате мясо приобретает некоторый аромат и лучше поддается кулинарной обработке. Пищевые достоинства его повышаются. Эти изменения в мягких тканях туши получили название «созревание» («вызревание») или «ферментация мяса».
Для объяснения процесса созревания мяса заслуживает большого внимания учение Мейергофа, Эмбдена, Палладина и Абдергальдена о динамике и обмене углеводов в мышцах при жизни животного.
Мейергоф
показал, что содержащийся в мышце
гликоген расходуется на образование
молочной кислоты при сокращении мышцы.
Во время расслабления
(отдыха) мышцы,
благодаря поступлению кислорода, из
молочной кислоты снова синтезируется
гликоген
Люндсград
показал, что креатинофосфорная кислота
находится в мышечных клетках и при
сокращении их расщепляется на креатин
и фосфорную кислоту (по
Палладину),
которая соединяется с гексозой (глюкозой).
Аденозинофосфорная кислота, содержащаяся
в мышцах, также расщепляется с образованием
аденозина и фосфорной кислоты, которая
дри соединении с гексозой (глюкозой)
способствует образованию молочной
кислоты (Эмбден и Цимммерман).
Мясо только
что убитого животного (парное мясо)-
плотной консистенции, без выраженного
приятного специфического запаха, при
варке дает мутноватый неароматный
бульон и не обладает высокими вкусовыми
качествами. Более того, в первые часы
после убоя животного мясо окоченевает
и становится жестким.
Спустя 24-72 ч
после убоя животного (в зависимости от
температуры среды, аэрации и других
факторов) мясо приобретает новые
качественные показатели: исчезает его
жесткость, оно приобретает сочность и
специфический приятный запах, на
поверхности туши образуется плотная
пленка (корочка подсыхания), при варке
дает прозрачный ароматный бульон,
становится нежным и т. д.
Происходящие
в мясе процессы и изменения, в результате
которых оно приобретает желательные
качественные показатели, принято
называть созреванием мяса.
Созревание мяса представляет собой совокупность сложных биохимических процессов в мышечной ткани и изменений физико-коллоидной структуры белка, протекающих под действием его собственных ферментов.
Процессы, происходящие в мышечной ткани после убоя животного, можно условно подразделить на три следующие фазы: послеубойное окоченение, созревание и автолиз.
Послеубойное
окоченение в туше развивается в первые
часы после убоя животного При этом мышцы
становятся упругими и слегка укорачиваются
Это значительно увеличивает их жесткость
и сопротивление на разрезе.
Способность
такого мяса к набуханию очень низкая.
При температуре 15-20"С полное окоченение
происходит через 3-5 ч после убоя
животного, а при температуре 0-2°С-через
18-20 ч.
Процесс
послеубойного окоченения сопровождается
некоторым повышением температуры в
туше в результате выделения тепла,
которое образуется от протекающих в
тканях химических реакций. Окоченение
мышечной ткани, наблюдающееся в первые
часы и сутки после убоя животных,
обусловлено образованием из белков
актина и миозина нерастворимого
актомиозинового комплекса. Предпосылкой
его образования являются отсутствие
аденозинтрифосфорной кислоты (АТФ),
кислая среда мяса и накопление в нем
молочной кислоты. Биохимические изменения
в мясе создают эти предпосылки.
Уменьшение
и полное исчезновение АТФ связано с ее
распадом в результате ферментативного
действия миозина Распад АТФ до
аденозиндифосфорной (АДФ, аденозинмонофосфорной
(АМФ) и фосфорной кислот сам по себе
приводит к появлению кислой среды в
мясе. Более того, уже в этой фазе начинается
распад мышечного гликогена, что приводит
к накоплению молочной кислоты, так же
способствующей образованию в нем кислой
среды.
Кислая среда, которая является закономерным явлением распада АТФ и началом необратимого процесса гликолиза (распада мышечного гликогена), усиливает мышечное окоченение. Замечено, что мышцы животных, погибших при явлениях судорог, окоченевают быстрее. Окоченение без накопления молочной кислоты характеризуется слабым мышечным напряжением и быстрым разрешением процесса.
Однако уже
задолго до завершения фазы окоченения
в мясе развиваются процессы, связанные
с фазами его собственного созревания
и аутолиза.
Ведущими для них являются
два процесса - интенсивный распад
мышечного гликогена, приводящий к
резкому сдвигу величины рН мяса в кислую
сторону, а также некоторые изменения
химического состава и физико-коллоидной
структуры белков.
В связи с тем что мышцы мяса кислорода не получают и окислительные процессы в них заторможены, в мясе накапливаются избытки молочной и фосфорной кислоты. Так, например, при мышечном утомлении организма (при его жизни) достигается максимум 0,25% молочной кислоты, а при посмертном окоченении ее накопляется до 0,82%. Активная реакция среды (рН) при этом изменяется от 7,26 до 6,02. От накопления молочной кислоты наступает быстрое сокращение (окоченение) мускулатуры, сопровождающееся коагуляцией белка (Саксль). При этом актомиозин теряет свою растворимость, белки стабилизируются, а кальций выпадает из коллоидов белка и переходит в раствор (мясной сок). Вследствие избыточного содержания молочной кислоты вначале наступает набухание коллоидоанизотропного вещества (темного диска) мышечных волокон (оно сопровождается укорочением- окоченением мышц); затем по мере увеличения концентрации молочной кислоты и коагуляции белка происходит размягчение этого вещества. Свернувшиеся белки теряют свои коллоидные свойства, становятся неспособными связывать (удерживать) воду и в известной степени лишаются своей дисперсной среды (воды): вместо первоначального разбухания наступает сморщивание (съеживание) коллоидов клеток, и мышцы становятся мягкими (разрешение окоченения).
В результате накопления молочной, фосфорной и других кислот в мясе увеличивается концентрация водородных ионов, вследствие чего к концу суток рН снижается до 5,8-5,7 (и даже ниже).
В кислой среде при распаде АТФ, АДФ, АМФ и фосфорной кислоты происходит частичное накопление неорганического фосфора. Резко кислая среда и наличие неорганического фосфора считается причиной диссоциации актомиозинового комплекса на актин и миозин. Распад этого комплекса снимает явления окоченения и жесткости мяса. Следовательно, фазу окоченения от других фаз обособить нельзя и ее необходимо считать одним из этапов процесса созревания мяса.
Схему биохимических изменений в процессе созревания мяса можно представить следующим образом.
Кислая среда сама по себе действует бактериостатически и даже бактерицидно, а поэтому при сдвиге рН в кислую сторону в мясе создаются неблагоприятные условия для развития микроорганизмов.
Наконец,
кислая среда приводит к некоторым
изменениям химического состава и
физико-коллоидной структуры белков.
Она изменяет проницаемость мышечных
оболочек и степень дисперсности белков.
Кислоты вступают во взаимодействие с
протеинатами кальция и кальций отщепляют
от белков.
Переход кальция в экстракт
ведет к уменьшению дисперсности белков,
в результате чего теряется часть гидратно
связанной воды. Поэтому из созревшего
мяса центрифугированием можно частично
отделить мясной сок.
Высвободившаяся гидратносвязанная вода, воздействие про-теолитических ферментов и кислая среда создают условия разрыхления сарколеммы мышечных волокон, и в первую очередь разрыхления и набухания коллагена. Это в значительной степени способствует изменению консистенции мяса и более выраженной его сочности. Очевидно, с набуханием коллагена, а затем частичной отдачей влаги с поверхности туши в окружающую среду следует связывать образование на ее поверхности корочки подсыхания.
Фаза собственного созревания во многом определяет интенсивность течения физико-коллоидных процессов и микроструктурных изменений мышечных волокон, которые бывают в фазе автолиза. Автолиз при созревании мяса понижают в широком смысле слова и связывают его не только с распадом белков, но и с процессом распада любых составных частей клеток. В связи с этим процессы, происходящие в фазе собственного созревания, невозможно отделить или обособить от таковых при автолизе. Тем не менее в результате комплекса причин (действие протеолитических ферментов, резко кислая среда, продукты автолитического распада небелковых веществ и др.) происходит автолитический распад мышечных волокон на отдельные сегменты.
Созревание
мяса совершается в течение 24-72 часов
при температуре +4°.
Однако не всегда
удастся выдерживать мясо при +4°. Иногда
приходится хранить его в обычных условиях
(не в остывочных) при температуре +6-8°
и выше; при повышенной температуре
процессы посмертного окоченения и
разрешения мышц протекают быстрее.
Скорость созревания мяса зависит также
от вида и состояния здоровья убитого
животного, его упитанности и возраста;
но эти вопросы требуют дальнейшего
наблюдения и изучения.
При созревании
мяса происходит расщепление некоторых
нуклеидов
(азотистых экстрактивных
веществ). Образуются летучие вещества,
эфиры и альдегиды, придающие аромат
мясу. Появляются адениловая и инозиновая
кислоты, аденин, ксантин, гипоксантин,
от которых и зависят вкусовые качества
мяса. Меняется реакция среды мяса в
сторону кислотности (рН 6,2-
5,8). Это
способствует набуханию коллоидов
протоплазмы, благодаря чему мясо
приобретает мягкость, нежность и хорошо
поддается кулинарной обработке.
Мясо
такого качества получается через 1-3
суток его хранения при температуре от
4 до 12° (в зависимости от возможностей
предприятий).
На первом этапе этого процесса обнаруживается сегментация в отдельных мышечных волокнах при сохранении эндомизия волокон. При этом в сегментах сохраняется структура ядер, поперечная и продольная исчерченность.
На втором
этапе сегментации подвергаются
большинство мышечных волокон.
Как и
на первом этапе, эндомизий волокон, а в
сегментах структура ядер, поперечная
и продольная исчерченность продолжают
сохраняться. Наконец, на третьем этапе
(фаза глубокого автолиза) обнаруживается
распад сегментов на миофибриллы, а
миофибрилл на саркомеры.
Саркомеры при микроскопии срезов, сделанных из такого Мяса, просматриваются в виде зернистой массы, заключенной в эндомизий.
Морфологические и микроструктурные изменения в тканях также являются причиной размягчения и разрыхления мяса в процессе его созревания, благодаря чему пищеварительные соки более свободно проникают к саркоплазме, что улучшает ее переваримость. Необходимо отметить, что соединительнотканные белки при созревании мяса почти не подвергаются протеолитическим процессам. Поэтому при равных условиях созревания нежность различных отрубов мяса одного и того же животного, а также одинаковых отрубов различных животных оказывается неодинаковой; нежность мяса, содержащего много соединительной ткани, невелика, а мясо молодых животных нежнее, чем старых.
В результате комплекса автолитических превращений различных компонентов мяса при его созревании образуются и накапливаются вещества, обусловливающие аромат и вкус созревшего мяса. Определенный вкус и аромат придают созревшему мясу азотсодержащие экстрактивные вещества - гипоксантин, креатин и креатинин, образующиеся при распаде АТФ, а также накапливающиеся свободные аминокислоты (глутаминовая кислота, аргинин, треонин, фенилаланин и др.). В образовании букета вкуса и аромата, по- видимому, участвуют пировиноградная и молочная кислоты.
И. А. Смородинцев высказывал предположение, что вкус и аромат зависят от накопления в созревшем мясе легкорастворимых и летучих веществ типа эфиров, альдегидов и кетонов. В дальнейшем в ряде исследований показано, что ароматические свойства созревшего мяса улучшаются по мере накопления в нем общего количества летучих редуцирующих веществ. В настоящее время при помощи газовой хроматографии и масс-спектрометрического анализа установлено, что к соединениям, обусловливающим запах вареного мяса, относятся ацетальдегид, ацетон, мртилэтилкетон, метанол, метилмеркаптан, диметилсульфид, этилмеркаптан и др.
При повышении температуры (до 30 °С), а также при длительной выдержке мяса (свыше 20-26 суток) в условиях низких плюсовых температур ферментативный процесс созревания заходит так глубоко, что в мясе заметно увеличивается количество продуктов распада белков в виде малых пептидов и свободных аминокислот. На этой стадии мясо приобретает коричневую окраску, в нем увеличивается количество аминного и аммиачного азота, происходит заметный гидролитический распад жиров, что резко снижает его товарные и пищевые качества.
Биохимические
процессы, происходящие при созревании
в мясе больных животных, отличаются от
биохимических процессов в мясе здоровых
животных.
При лихорадке и переутомлении
энергетический процесс в организме
повышен.
Окислительные процессы в
тканях усилены. Изменение углеводного
обмена при болезнях и переутомлении
характеризуется быстрой убылью гликогена
в мускулатуре. Поэтому почти при всяком
патологическом процессе в организме
животного содержание гликогена в мышцах
сокращается. Поскольку гликогена в мясе
больных животных меньше, чем в мясе
здоровых, то и количество продуктов
распада гликогена (глюкозы, молочной
кислоты и др.) в мясе больных животных
незначительное.
Кроме того, при тяжело протекающих заболеваниях еще при жизни животного в его мускулатуре накапливаются промежуточные и конечные продукты белкового метаболизма. В этих случаях уже в первые часы после убоя животного в мясе обнаруживается повышенное количество аминного и аммиачного азота.
Незначительное накопление кислот и повышенное содержание полипептидов, аминокислот и аммиака являются причиной меньшего снижения показателя концентрации водородных ионов при созревании мяса больных животных. Этот фактор влияет на активность ферментов мяса. В большинстве случаев концентрация водородных ионов, устанавливающаяся в результате созревания мяса больных животных, более благоприятна для действия пептидаз и протеаз.
В итоге накопление в мясе больных животных экстрактивных азотистых веществ и отсутствие резкого сдвига величины рН в кислую сторону считаются условиями, благоприятными для развития микроорганизмов.
Изменения, происходящие в мясе больных животных, по-иному влияют и на характер физико-коллоидной структуры мяса. Меньшая кислотность вызывает незначительное выпадение солей кальция, что, в свою очередь, является причиной меньшего изменения степени дисперсности белков и других изменений, характерных для них при нормальном созревании мяса. Сравнительно высокий показатель рН, накопление продуктов распада белков и благоприятные условия для развития микроорганизмов предопределяют меньшую стойкость мяса больных животных при хранении. Перечисленные признаки свойственны мясу каждого тяжелобольного животного; они являются причиной известной однотипности в изменении физико-химических показателей мяса, полученного от животных, убитых с течением патологического процесса, независимо от природы заболевания. Это положение не отрицает, специфических изменений в составе мяса при отдельных заболеваниях, но дает основание говорить об общих закономерностях созревания мяса при патологии в животном организме.
Соединениями, характеризующимися большой молекулярной массой. В состав всех известных белков ... постоянной свою форму и химический состав , несмотря на непрерывное их...
Химический состав и физические свойства спермы
Доклад >> Медицина, здоровьеХимический состав и физические свойства спермы Сперма – смесь... , А), макро и микроэлементы. Химический состав спермы: 1)вода- 75% 2) сухое вещество- 25%: -белки - 85% -Липиды...
Аминокислоты — структурные компоненты белков.Белки, или протеины (греч. protos — первостепенный), — это биологические гетерополимеры, мономерами которых являются аминокислоты.
Аминокислоты представляют собой низкомолекулярные органические соединения, содержащие карбоксильную (-СООН) и аминную (-NH 2) группы, которые связаны с одним и тем же атомом углерода. К атому углерода присоединяется боковая цепь — какой-либо радикал, придающий каждой аминокислоте определенные свойства. Общая формула аминокислот имеет вид:

У большей части аминокислот имеется одна карбоксильная группа и одна аминогруппа; эти аминокислоты называютсянейтральными. Существуют, однако, и основные аминокислоты — с более чем одной аминогруппой, а также кислые аминокислоты — с более чем одной карбоксильной группой.
Известно около 200 аминокислот, встречающихся в живых организмах, однако только 20 из них входят в состав белков. Это так называемые основные, или белокобразующие (протеиногенные), аминокислоты.
В зависимости от вида радикала основные аминокислоты делят на три группы: 1) неполярные (аланин, метионин, валин, про-лин, лейцин, изолейцин, триптофан, фенилаланин); 2) полярные незаряженные (аспарагин, глутамин, серии, глицин, тирозин, треонин, цистеин); 3) полярные заряженные (аргинин, гистидин, лизин — положительно; аспарагиновая и глутаминовая кислоты — отрицательно).
Боковые цепи аминокислот (радикал) могут быть гидрофобными или гидрофильными, что придает белкам соответствующие свойства, которые проявляются при образовании вторичной, третичной и четвертичной структур белка.
У растений все необходимые аминокислоты синтезируются из первичных продуктов фотосинтеза. Человек и животные не способны синтезировать ряд протеиногенных аминокислот и должны получать их в готовом виде вместе с пищей. Такие аминокислоты называются незаменимыми. К ним относятся лизин, валин, лейцин, изолейцин, треонин, фенилаланин, триптофан, метионин; атакже аргинин и гистидин — незаменимые для детей,
В растворе аминокислоты могут выступать в роли как кислот, так и оснований, т. е. они являются амфотерными соединениями. Карбоксильная группа -СООН способна отдавать протон, функционируя как кислота, а аминная - NH2 — принимать протон, проявляя таким образом свойства основания.
Пептиды. Аминогруппа одной аминокислоты способна вступать в реакцию с карбоксильной группой другой аминокислоты.
Образующаяся при этом молекула представляет собой дипептид, а связь -CO-NH- называется пептидной связью:

На одном конце молекулы дипептида находится свободная аминогруппа, а на другом — свободная карбоксильная группа. Благодаря этому дипептид может присоединять к себе другие аминокислоты, образуя олигопептиды. Если таким образом соединяется много аминокислот (более десяти), то получается полипептид.
Пептиды играют важную роль в организме. Многие олиго- и полипептиды являются гормонами, антибиотиками, токсинами.
К олигопептидам относятся окситоцин, вазопрессин, тиреотропин, а также брадикинин (пептид боли) и некоторые опиаты («естественные наркотики» человека), выполняющие функцию обезболивания. Принятие наркотиков разрушает опиатную систему организма, поэтому наркоман без дозы наркотиков испытывает сильную боль — «ломку», которая в норме снимается опиатами. К олигопептидам относятся и некоторые антибиотики (например, грамицидин S).
Многие гормоны (инсулин, адренокортикотропный гормон и др,), антибиотики (например, грамицидин А), токсины (например, дифтерийный токсин) являются полипептидами.
Белки представляют собой полипептиды, в молекулу которых входит от пятидесяти до нескольких тысяч аминокислот с относительной молекулярной массой свыше 10 000.
Структура белков. Каждому белку в определенной среде свойственна особая пространственная структура. При характеристике пространственной (трехмерной) структуры выделяют четыре уровня организации молекул белков (рис. 1,1).
|
лиэ—глу—тре—ала—ала—ала—лиз—фен—глу—арг—глн—гиc—мет—асп—сер— |
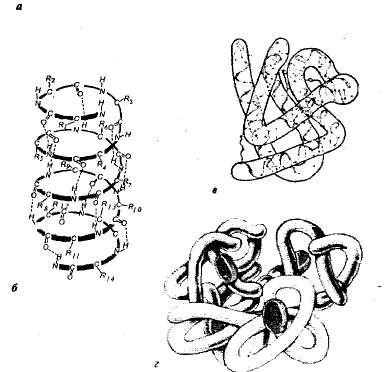
Рис. 1.1. Уровни структурной организации белка: а — первичная структура — аминокислотная последовательность белкарибонуклеазы (124 аминокислотных звена); б — вторичная структура — пояипептидная цепь закручена в виде спирали; в — третичная структура белка миоглобина; г — четвертичная структура гемоглобина.
Первичная структура — последовательность аминокислот в полипептидной цепи. Такая структура специфична для каждого белка и определяется генетической информацией, т. е. зависит от последовательности нуклеотидов в участке молекулы ДНК, кодирующем данный белок. От первичной структуры зависят все свойства и функции белков. Замена одной-единственной аминокислоты в составе молекул белка или нарушение порядка в их расположении обычно влечет за собой изменение функции белка.
Учитывая, что в состав белков входит 20 видов аминокислот, число вариантов их комбинаций в полипептидной цепи поистине безгранично, что обеспечивает огромное количество видов белков в живых клетках. Например, в организме человека обнаружено более 10 тыс. различных белков, и все они построены из одних и тех же 20 основных аминокислот.
В живых клетках молекулы белков или отдельные их участки представляют собой не вытянутую цепь, а скручены в спираль, напоминающую растянутую пружину (это так называемая а-спираль), или сложены в складчатый слой (р-слой). Такие а-спирали и р-слои являются вторичной структурой. Она возникает в результате образования водородных связей внутри одной полипептидной цепи (спиральная конфигурация) или между двумя полипептидными цепями (складчатые слои).
Полностью a-спиральную конфигурацию имеет белок кератин. Это структурный белок волос, ногтей, когтей, клюва, перьев и рогов; он входит в состав наружного слоя кожи позвоночных.
У большинства белков спиральные и неспиральные участки полипептидной цепи складываются в трехмерное образование шаровидной формы — глобулу (характерна для глобулярных белков). Глобула определенной конфигурации является третичной структурой белка. Такая структура стабилизируется ионными, водородными, ковалентными дисульфидными связями (образуются между атомами серы, входящими в состав цистеи-на, цистина и мегионина), а также гидрофобными взаимодействиями. Наиболее важными в возникновении третичной структуры являются гидрофобные взаимодействия; белок при этом свертывается таким образом, что его гидрофобные боковые цепи скрыты внутри молекулы, т. е. защищены от соприкосновения с водой, а гидрофильные боковые цепи, наоборот, выставлены наружу.
Многие белки с особо сложным строением состоят из нескольких полипептидных цепей (субъединиц), образуя четвертичную структуру белковой молекулы. Такая структура имеется, например, у глобулярного белка гемоглобина. Его молекула состоит из четырех отдельных полипептидных субъединиц (протомеров), находящихся в третичной структуре, и небелковой части — гема.
Только в такой структуре гемоглобин способен выполнять свою транспортную функцию.
Под влиянием различных химических и физических факторов (обработка спиртом, ацетоном, кислотами, щелочами, высокой температурой, облучением, высоким давлением и т. д.) происходит изменение вторичной, третичной и четвертичной структур белка вследствие разрыва водородных и ионных связей. Процесс нарушения нативной (естественной) структуры белка называетсяденатурацией. При этом наблюдается уменьшение растворимости белка, изменение формы и размеров молекул, потеря ферментативной активности и т. д. Процесс денатурации может быть полным или частичным. В некоторых случаях переход к нормальным условиям среды сопровождается самопроизвольным восстановлением естественной структуры белка. Такой процесс называется ренатурацией.
Простые и сложные белки. По химическому составу выделяют белки простые и сложные. К простьм относятся белки, состоящие только из аминокислот, а к сложный — белки, содержащие белковую часть и небелковую (простетическую); простетическую группу могут образовывать ионы металлов, остаток фосфорной кислоты, углеводы, липиды и др. Простыми белками являются сывороточный альбумин крови, фибрин, некоторые ферменты (трипсин) и др. К сложным белкам относятся все протеолипиды и гликопротеины; сложными белками являются, например, иммуноглобулины (антитела), гемоглобин, большинство ферментов и т. д.
Функции белков.
- Структурная. Белки входят в состав клеточных мембран и матрикса органелл клетки. Стенки кровеносных сосудов, хрящи, сухожилия, волосы, ногти, когти у высших животных состоят преимущественно из белков.
- Каталитическая (ферментативная). Белки-ферменты катализируют протекание всех химических реакций в организме. Они обеспечивают расщепление питательных веществ в пищеварительном тракте, фиксацию углерода при фотосинтезе и т. д.
- Транспортная. Некоторые белки способны присоединять и переносить различные вещества. Альбумины крови транспортируют жирные кислоты, глобулины -— ионы металлов и гормоны, гемоглобин — кислород и углекислый газ. Молекулы белков, входящие в состав плазматической мембраны, принимают участие в транспортировке веществ в клетку.
- Защитная. Ее выполняют иммуноглобулины (антитела) крови, обеспечивающие иммунную защиту организма. Фибриноген и тромбин участвуют в свертывании крови и предотвращают кровотечение.
- Сократительная. Благодаря скольжению относительно друг друга актиновых и миозиновых протофибрилл происходит сокращение мышц, а также немышечные внутриклеточные сокращения. Движение ресничек и жгутиков связано со скольжением относительно друг друга микротрубочек, имеющих белковую природу.
- Регуляторная. Многие гормоны являются олигопептидами или бедками (например, инсулин, глюкагон [антагонист инсулина], адренокортикотропный гормон и др.).
- Рецепторная. Некоторые белки, встроенные в клеточную мембрану, способны изменять свою структуру под воздействием внешней среды. Так происходит прием сигналов извне и передача информации в клетку. Примером может служить фито-хром —- светочувствительный белок, регулирующий фотопериодическую реакцию растений, и опсин — составная часть родопсина, пигмента, находящегося в клетках сетчатки глаза.
- Энергетическая. Белки могут служить источником энергии в клетке (после их гидролиза). Обычно белки расходуются на энергетические нужды в крайних случаях, когда исчерпаны запасы углеводов и жиров.
Ферменты (энзимы). Это специфические белки, которые присутствуют во всех живых организмах и играют роль биологических катализаторов.
Химические реакции в живой клетке протекают при определенной температуре, нормальном давлении и соответствующей кислотности среды. В таких условиях реакции синтеза или распада веществ протекали бы в клетке очень медленно, если бы они не подвергались воздействиям ферментов. Ферменты ускоряют реакцию без изменения ее общего результата за счет сниженияэнергии активации, т. е. при их присутствии требуется значительно меньше энергии для придания реакционной способности молекулам, которые вступают в реакцию, или реакция идет по другому пути с меньшим энергетическим барьером.
Все процессы в живом организме прямо или косвенно осуществляются с участием ферментов. Например, под их действием составные компоненты пищи (белки, углеводы, липиды и др.) расщепляются до более простых соединений, а из них уже затем синтезируются новые, свойственные данному виду макромолекулы. Поэтому нарушения образования и активности ферментов нередко ведут к возникновению тяжелых болезней.
По пространственной организации ферменты состоят из нескольких пол и пептидных цепей и обычно обладают четвертичной структурой. Кроме того, ферменты могут включать и небелковые структуры. Белковая часть носит название апофермент, а небелковая — кофактор (если это катионы или анионы неорганических веществ, например, Zn 2- Мп 2+ и т. д.) или кофермент (коэнзим) (если это низкомолекулярное органическое вещество).
Предшественниками или составными частями Многих кофер-ментов являются витамины. Так, пантотеновая кислота — составная часть коэнзима А, никотиновая кислота (витамин РР) — предшественник НАД и НАДФ и т. д.
Ферментативный катализ подчиняется тем же законам, что и неферментативный катализ в химической промышленности, однако в отличие от него характеризуется необычайно высокой степенью специфичности (фермент катализирует только одну реакцию или действует только на один тип связи). Этим обеспечивается тонкая регуляция всех жизненно важных процессов (дыхание, пищеварение, фотосинтез и др.), протекающих в клетке и организме. Например, фермент уреаза катализирует расщепление лишь одного вещества — мочевины (H 2 N-CO-NH 2 + Н 2 О —> —» 2NH 3 + СО 2), не оказывая каталитического действия на структурно-родственные соединения.
Для понимания механизма действия ферментов, обладающих высокой специфичностью, очень важна теория активного центра. Согласно ей, в молекуле каждого фермента имеется одни участок или более, в которых происходит катализ за счет тесного (во многих точках) контакта между молекулами фермента и специфического вещества (субстрата). Активным центром выступает или функциональная группа (например, ОН-группа серина), или отдельная аминокислота. Обычно же для каталитического действия необходимо сочетание нескольких (в среднем от 3 до 12) расположенных в определенном порядке аминокислотных остатков. Активный центр также формируется связанными с ферментом ионами металлов, витаминами и другими соединениями небелковой природы — коферментами, или кофакторами. Причем форма и химическое строение активного центра таковы, что с ним могут связываться только определенные субстраты в силу их идеального соответствия (взаимодополняемости или. комплементарности) друг другу. Роль остальных аминокислотных остатков в крупной молекуле фермента состоит в том, чтобы обеспечить его молекуле соответствующую глобулярную форму, которая нужна для эффективной работы активного центра. Кроме того, вокруг крупной молекулы фермента возникает сильное электрическое поле. В таком поле становится возможной ориентация молекул субстрата и приобретение ими асимметричной формы. Это приводит к ослаблению химических связей, и катализируемая реакция происходит с меньшей начальной затратой энергии, а следовательно, с намного большей скоростью. Например, одна молекула фермента каталазы может расщепить за 1 мин более 5 млн. молекул пероксида водорода (Н 2 0 2), который возникает при окислении в организме различных соединений.
У некоторых ферментов в присутствии субстрата конфигурация активного центра претерпевает изменения, т. е. фермент ориентирует свои функциональные группы таким образом, чтобы обеспечить наибольшую каталитическую активность.
На заключительном этапе химической реакции фермент-субстратный комплекс разъединяется с образованием конечных продуктов и свободного фермента. Освободившийся при этом активный центр может принимать новые молекулы субстрата.
Скорость ферментативных реакций зависит от многих факторов: природы и концентрации фермента и субстрата, температуры, давления, кислотности среды, наличия ингибиторов и т. д. Например, при температурах, близких к нулю, скорость биохимических реакций замедляется до минимума. Это свойство широко используется в различных отраслях народного хозяйства, особен-но в сельском хозяйстве и медицине. В частности, консервация различных органов (почек, сердца, селезенки, печени) перед их пересадкой больному происходит при охлаждении с целью снижения интенсивности биохимических реакций и продления времени жизни органов. Быстрое замораживание пищевых продуктов предотвращает рост и размножение микроорганизмов (бактерий, грибов и др.), атакже инактивирует их пищеварительные ферменты, так что они оказываются уже не в состоянии вызвать разложение пищевых продуктов.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов "Пособие по биологии для поступающих в ВУЗы"
Белки — высокомолекулярные органические соединения, состоящие из остатков α-аминокислот.
В состав белков входят углерод, водород, азот, кислород, сера. Часть белков образует комплексы с другими молекулами, содержащими фосфор, железо, цинк и медь.
Белки обладают большой молекулярной массой: яичный альбумин — 36 000, гемоглобин — 152 000, миозин — 500 000. Для сравнения: молекулярная масса спирта — 46, уксусной кислоты — 60, бензола — 78.
Аминокислотный состав белков
Белки — непериодические полимеры, мономерами которых являются α-аминокислоты . Обычно в качестве мономеров белков называют 20 видов α-аминокислот, хотя в клетках и тканях их обнаружено свыше 170.
В зависимости от того, могут ли аминокислоты синтезироваться в организме человека и других животных, различают: заменимые аминокислоты — могут синтезироваться; незаменимые аминокислоты — не могут синтезироваться. Незаменимые аминокислоты должны поступать в организм вместе с пищей. Растения синтезируют все виды аминокислот.
В зависимости от аминокислотного состава, белки бывают: полноценными — содержат весь набор аминокислот; неполноценными — какие-то аминокислоты в их составе отсутствуют. Если белки состоят только из аминокислот, их называют простыми . Если белки содержат помимо аминокислот еще и неаминокислотный компонент (простетическую группу), их называют сложными . Простетическая группа может быть представлена металлами (металлопротеины), углеводами (гликопротеины), липидами (липопротеины), нуклеиновыми кислотами (нуклеопротеины).
Все аминокислоты содержат : 1) карбоксильную группу (-СООН), 2) аминогруппу (-NH 2), 3) радикал или R-группу (остальная часть молекулы). Строение радикала у разных видов аминокислот — различное. В зависимости от количества аминогрупп и карбоксильных групп, входящих в состав аминокислот, различают: нейтральные аминокислоты , имеющие одну карбоксильную группу и одну аминогруппу; основные аминокислоты , имеющие более одной аминогруппы; кислые аминокислоты , имеющие более одной карбоксильной группы.
Аминокислоты являются амфотерными соединениями , так как в растворе они могут выступать как в роли кислот, так и оснований. В водных растворах аминокислоты существуют в разных ионных формах.
Пептидная связь
Пептиды — органические вещества, состоящие из остатков аминокислот, соединенных пептидной связью.
Образование пептидов происходит в результате реакции конденсации аминокислот. При взаимодействии аминогруппы одной аминокислоты с карбоксильной группой другой между ними возникает ковалентная азот-углеродная связь, которую и называют пептидной . В зависимости от количества аминокислотных остатков, входящих в состав пептида, различают дипептиды, трипептиды, тетрапептиды и т.д. Образование пептидной связи может повторяться многократно. Это приводит к образованию полипептидов . На одном конце пептида находится свободная аминогруппа (его называют N-концом), а на другом — свободная карбоксильная группа (его называют С-концом).

Пространственная организация белковых молекул
Выполнение белками определенных специфических функций зависит от пространственной конфигурации их молекул, кроме того, клетке энергетически невыгодно держать белки в развернутой форме, в виде цепочки, поэтому полипептидные цепи подвергаются укладке, приобретая определенную трехмерную структуру, или конформацию. Выделяют 4 уровня пространственной организации белков .
Первичная структура белка — последовательность расположения аминокислотных остатков в полипептидной цепи, составляющей молекулу белка. Связь между аминокислотами — пептидная.

Если молекула белка состоит всего из 10 аминокислотных остатков, то число теоретически возможных вариантов белковых молекул, отличающихся порядком чередования аминокислот, — 10 20 . Имея 20 аминокислот, можно составить из них еще большее количество разнообразных комбинаций. В организме человека обнаружено порядка десяти тысяч различных белков, которые отличаются как друг от друга, так и от белков других организмов.
Именно первичная структура белковой молекулы определяет свойства молекул белка и ее пространственную конфигурацию. Замена всего лишь одной аминокислоты на другую в полипептидной цепочке приводит к изменению свойств и функций белка. Например, замена в β-субъединице гемоглобина шестой глутаминовой аминокислоты на валин приводит к тому, что молекула гемоглобина в целом не может выполнять свою основную функцию — транспорт кислорода; в таких случаях у человека развивается заболевание — серповидноклеточная анемия.
Вторичная структура — упорядоченное свертывание полипептидной цепи в спираль (имеет вид растянутой пружины). Витки спирали укрепляются водородными связями, возникающими между карбоксильными группами и аминогруппами. Практически все СО- и NН-группы принимают участие в образовании водородных связей. Они слабее пептидных, но, повторяясь многократно, придают данной конфигурации устойчивость и жесткость. На уровне вторичной структуры существуют белки: фиброин (шелк, паутина), кератин (волосы, ногти), коллаген (сухожилия).



Третичная структура — укладка полипептидных цепей в глобулы, возникающая в результате возникновения химических связей (водородных, ионных, дисульфидных) и установления гидрофобных взаимодействий между радикалами аминокислотных остатков. Основную роль в образовании третичной структуры играют гидрофильно-гидрофобные взаимодействия. В водных растворах гидрофобные радикалы стремятся спрятаться от воды, группируясь внутри глобулы, в то время как гидрофильные радикалы в результате гидратации (взаимодействия с диполями воды) стремятся оказаться на поверхности молекулы. У некоторых белков третичная структура стабилизируется дисульфидными ковалентными связями, возникающими между атомами серы двух остатков цистеина. На уровне третичной структуры существуют ферменты, антитела, некоторые гормоны.

Четвертичная структура характерна для сложных белков, молекулы которых образованы двумя и более глобулами. Субъединицы удерживаются в молекуле благодаря ионным, гидрофобным и электростатическим взаимодействиям. Иногда при образовании четвертичной структуры между субъединицами возникают дисульфидные связи. Наиболее изученным белком, имеющим четвертичную структуру, является гемоглобин . Он образован двумя α-субъединицами (141 аминокислотный остаток) и двумя β-субъединицами (146 аминокислотных остатков). С каждой субъединицей связана молекула гема, содержащая железо.
Если по каким-либо причинам пространственная конформация белков отклоняется от нормальной, белок не может выполнять свои функции. Например, причиной «коровьего бешенства» (губкообразной энцефалопатии) является аномальная конформация прионов — поверхностных белков нервных клеток.
Свойства белков
Аминокислотный состав, структура белковой молекулы определяют его свойства . Белки сочетают в себе основные и кислотные свойства, определяемые радикалами аминокислот: чем больше кислых аминокислот в белке, тем ярче выражены его кислотные свойства. Способность отдавать и присоединять Н + определяют буферные свойства белков ; один из самых мощных буферов — гемоглобин в эритроцитах, поддерживающий рН крови на постоянном уровне. Есть белки растворимые (фибриноген), есть нерастворимые, выполняющие механические функции (фиброин, кератин, коллаген). Есть белки активные в химическом отношении (ферменты), есть химически неактивные, устойчивые к воздействию различных условий внешней среды и крайне неустойчивые.
Внешние факторы (нагревание, ультрафиолетовое излучение, тяжелые металлы и их соли, изменения рН, радиация, обезвоживание)
могут вызывать нарушение структурной организации молекулы белка. Процесс утраты трехмерной конформации, присущей данной молекуле белка, называют денатурацией . Причиной денатурации является разрыв связей, стабилизирующих определенную структуру белка. Первоначально рвутся наиболее слабые связи, а при ужесточении условий и более сильные. Поэтому сначала утрачивается четвертичная, затем третичная и вторичная структуры. Изменение пространственной конфигурации приводит к изменению свойств белка и, как следствие, делает невозможным выполнение белком свойственных ему биологических функций. Если денатурация не сопровождается разрушением первичной структуры, то она может быть обратимой , в этом случае происходит самовосстановление свойственной белку конформации. Такой денатурации подвергаются, например, рецепторные белки мембраны. Процесс восстановления структуры белка после денатурации называется ренатурацией . Если восстановление пространственной конфигурации белка невозможно, то денатурация называется необратимой .
Функции белков
| Функция | Примеры и пояснения |
|---|---|
| Строительная | Белки участвуют в образовании клеточных и внеклеточных структур: входят в состав клеточных мембран (липопротеины, гликопротеины), волос (кератин), сухожилий (коллаген) и т.д. |
| Транспортная | Белок крови гемоглобин присоединяет кислород и транспортирует его от легких ко всем тканям и органам, а от них в легкие переносит углекислый газ; в состав клеточных мембран входят особые белки, которые обеспечивают активный и строго избирательный перенос некоторых веществ и ионов из клетки во внешнюю среду и обратно. |
| Регуляторная | Гормоны белковой природы принимают участие в регуляции процессов обмена веществ. Например, гормон инсулин регулирует уровень глюкозы в крови, способствует синтезу гликогена, увеличивает образование жиров из углеводов. |
| Защитная | В ответ на проникновение в организм чужеродных белков или микроорганизмов (антигенов) образуются особые белки — антитела, способные связывать и обезвреживать их. Фибрин, образующийся из фибриногена, способствует остановке кровотечений. |
| Двигательная | Сократительные белки актин и миозин обеспечивают сокращение мышц у многоклеточных животных. |
| Сигнальная | В поверхностную мембрану клетки встроены молекулы белков, способных изменять свою третичную структуру в ответ на действие факторов внешней среды, таким образом осуществляя прием сигналов из внешней среды и передачу команд в клетку. |
| Запасающая | В организме животных белки, как правило, не запасаются, исключение: альбумин яиц, казеин молока. Но благодаря белкам в организме могут откладываться про запас некоторые вещества, например, при распаде гемоглобина железо не выводится из организма, а сохраняется, образуя комплекс с белком ферритином. |
| Энергетическая | При распаде 1 г белка до конечных продуктов выделяется 17,6 кДж. Сначала белки распадаются до аминокислот, а затем до конечных продуктов — воды, углекислого газа и аммиака. Однако в качестве источника энергии белки используются только тогда, когда другие источники (углеводы и жиры) израсходованы. |
| Каталитическая | Одна из важнейших функций белков. Обеспечивается белками — ферментами, которые ускоряют биохимические реакции, происходящие в клетках. Например, рибулезобифосфаткарбоксилаза катализирует фиксацию СО 2 при фотосинтезе. |
Ферменты
Ферменты , или энзимы , — особый класс белков, являющихся биологическими катализаторами. Благодаря ферментам биохимические реакции протекают с огромной скоростью. Скорость ферментативных реакций в десятки тысяч раз (а иногда и в миллионы) выше скорости реакций, идущих с участием неорганических катализаторов. Вещество, на которое оказывает свое действие фермент, называют субстратом .
Ферменты — глобулярные белки, по особенностям строения ферменты можно разделить на две группы: простые и сложные. Простые ферменты являются простыми белками, т.е. состоят только из аминокислот. Сложные ферменты являются сложными белками, т.е. в их состав помимо белковой части входит группа небелковой природы — кофактор . У некоторых ферментов в качестве кофакторов выступают витамины. В молекуле фермента выделяют особую часть, называемую активным центром. Активный центр — небольшой участок фермента (от трех до двенадцати аминокислотных остатков), где и происходит связывание субстрата или субстратов с образованием фермент-субстратного комплекса. По завершении реакции фермент-субстратный комплекс распадается на фермент и продукт (продукты) реакции. Некоторые ферменты имеют (кроме активного) аллостерические центры — участки, к которым присоединяются регуляторы скорости работы фермента (аллостерические ферменты ).

Для реакций ферментативного катализа характерны: 1) высокая эффективность, 2) строгая избирательность и направленность действия, 3) субстратная специфичность, 4) тонкая и точная регуляция. Субстратную и реакционную специфичность реакций ферментативного катализа объясняют гипотезы Э. Фишера (1890 г.) и Д. Кошланда (1959 г.).
Э. Фишер (гипотеза «ключ-замок») предположил, что пространственные конфигурации активного центра фермента и субстрата должны точно соответствовать друг другу. Субстрат сравнивается с «ключом», фермент — с «замком».
Д. Кошланд (гипотеза «рука-перчатка») предположил, что пространственное соответствие структуры субстрата и активного центра фермента создается лишь в момент их взаимодействия друг с другом. Эту гипотезу еще называют гипотезой индуцированного соответствия .
Скорость ферментативных реакций зависит от: 1) температуры, 2) концентрации фермента, 3) концентрации субстрата, 4) рН. Следует подчеркнуть, что поскольку ферменты являются белками, то их активность наиболее высока при физиологически нормальных условиях.
Большинство ферментов может работать только при температуре от 0 до 40 °С. В этих пределах скорость реакции повышается примерно в 2 раза при повышении температуры на каждые 10 °С. При температуре выше 40 °С белок подвергается денатурации и активность фермента падает. При температуре, близкой к точке замерзания, ферменты инактивируются.
При увеличении количества субстрата скорость ферментативной реакции растет до тех пор, пока количество молекул субстрата не станет равным количеству молекул фермента. При дальнейшем увеличении количества субстрата скорость увеличиваться не будет, так как происходит насыщение активных центров фермента. Увеличение концентрации фермента приводит к усилению каталитической активности, так как в единицу времени преобразованиям подвергается большее количество молекул субстрата.

Для каждого фермента существует оптимальное значение рН, при котором он проявляет максимальную активность (пепсин — 2,0, амилаза слюны — 6,8, липаза поджелудочной железы — 9,0). При более высоких или низких значениях рН активность фермента снижается. При резких сдвигах рН фермент денатурирует.
Скорость работы аллостерических ферментов регулируется веществами, присоединяющимися к аллостерическим центрам. Если эти вещества ускоряют реакцию, они называются активаторами , если тормозят — ингибиторами .
Классификация ферментов
По типу катализируемых химических превращений ферменты разделены на 6 классов:
- оксиредуктазы (перенос атомов водорода, кислорода или электронов от одного вещества к другому — дегидрогеназа),
- трансферазы (перенос метильной, ацильной, фосфатной или аминогруппы от одного вещества к другому — трансаминаза),
- гидролазы (реакции гидролиза, при которых из субстрата образуются два продукта — амилаза, липаза),
- лиазы (негидролитическое присоединение к субстрату или отщепление от него группы атомов, при этом могут разрываться связи С-С, С-N, С-О, С-S — декарбоксилаза),
- изомеразы (внутримолекулярная перестройка — изомераза),
- лигазы (соединение двух молекул в результате образования связей С-С, С-N, С-О, С-S — синтетаза).
Классы в свою очередь подразделены на подклассы и подподклассы. В действующей международной классификации каждый фермент имеет определенный шифр, состоящий из четырех чисел, разделенных точками. Первое число — класс, второе — подкласс, третье — подподкласс, четвертое — порядковый номер фермента в данном подподклассе, например, шифр аргиназы — 3.5.3.1.
Перейти к лекции №2 «Строение и функции углеводов и липидов»
Перейти к лекции №4 «Строение и функции нуклеиновых кислот АТФ»
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru
ГЛАВА 1. ВВЕДЕНИЕ
Довольно банальными стали сейчас сообщения о революции в биологии. Бесспорным считается и то, что эти революционные изменения были связаны с формированием на стыке биологии и химии комплекса наук, среди которых центральное положение занимали и занимают молекулярная биология и биоорганическая химия.
“Молекулярная биология наука, ставящая своей целью познание природы явлений жизнедеятельности путем изучения биологических объектов и систем на уровне, приближающемся к молекулярному… характерные проявления жизни… обусловлены структурой, свойствами и взаимодействием молекул биологически важных веществ, в первую очередь белков и нуклеиновых кислот ”
“Биоорганическая химия - наука, изучающая вещества, лежащие в основе процессов жизнедеятельности…основные объекты биоорганической химии биополимеры (белки и пептиды, нуклеиновые кислоты и нуклеотиды, липиды, полисахариды и т.д.).
Из этого сопоставления становится очевидным, сколь важно для развития современной биологии изучение белков.
биология белок биохимия
ГЛАВА 2. ИСТОРИЯ ИССЛЕДОВАНИЯ БЕЛКА
2.1 Начальные этапы в химии белка
Белок попал в число объектов химических исследований 250 лет тому назад. В 1728 году итальянский ученый Якопо Бартоломео Беккари получил из пшеничной муки первый препарат белкового вещества - клейковины. Он подверг клейковину сухой перегонке и убедился, что продукты такой перегонки были щелочными. Это было первое доказательство единства природы веществ растительного и животного царств. Он опубликовал результаты своей работы в 1745 году, и это была первая статья о белке.
В XVIII - начале XIX веков неоднократно описывали белковые вещества растительного и животного происхождения. Особенностью таких описаний было сближение этих веществ и сопоставление их с веществами неорганическими.
Важно отметить, что в это время, еще до появления элементного анализа, сложилось представление о том, что белки из различных источников - это группа близких по общим свойствам индивидуальных веществ.
В 1810 году Ж. Гей-Люссак и Л. Тенар впервые определили элементный состав белковых веществ. В 1833 году Ж. Гей-Люссак доказал, что в белках обязательно присутствует азот, а вскоре было показано, что содержание азота в различных белках приблизительно одинаково. В это же время английский химик Д. Дальтон попытался изобразить первые формулы белковых веществ. Он представлял их довольно просто устроенными веществами, но чтобы подчеркнуть их индивидуальное различие при одинаковом составе, он прибег к изображению молекул, которые бы сейчас назвали изомерными. Однако понятия изомерии во времена Дальтона еще не было.
Формулы белков Д. Дальтона
Были выведены первые эмпирические формулы белков и выдвинуты первые гипотезы относительно закономерностей их состава. Так, Н.Либеркюн считал, что альбумин описывается формулой C 72 H 112 N 18 SO 22 , а А.Данилевский полагал, что молекула этого белка по крайней мере на порядок больше: C 726 H 1171 N 194 S 3 O 214 .
Немецкий химик Ю. Либих в 1841 году предположил, что белки животного происхождения имеют аналоги среди растительных белков: усвоение белка легумина в организме животного, по Либиху, вело к накоплению аналогичного белка - казеина. Одной из самых распространенных теорий доструктурной органической химии была теория радикалов - неизменных компонентов родственных веществ. В 1836 году голландец Г. Мульдер высказал предположение о том, что все белки содержат один и тот же радикал, который он назвал протеином (от греческого слова “первенствую”, “занимаю первое место”). Протеин, по Мульдеру, имел состав Pr = C 40 H 62 N 10 O 12 . В 1838 году Г. Мульдер опубликовал формулы белков, построенные на основании теории протеина. Это были т.н. дуалистические формулы, где радикал протеина служил положительной группировкой, а атомы серы или фосфора - отрицательной. Вместе они образовывали электронейтральную молекулу: белок сыворотки крови Pr 10 S 2 P, фибрин Pr 10 SP. Однако аналитическая проверка данных Г. Мульдера, проведенная русским химиком Лясковским, а также Ю. Либихом, показала, что “белковых радикалов” не существует.
В 1833 году немецкий ученый Ф. Розе открыл биуретовую реакцию на белки - одну из основных цветных реакций на белковые вещества и их производные в настоящее время (подробнее о цветных реакциях на стр.53). Был сделан также вывод о том, что это самая чувствительная реакция на белок, поэтому она в то время привлекла наибольшее внимание химиков.
В середине XIX века были разработаны многочисленные методы экстракции белков, очистки и выделения их в растворах нейтральных солей. В 1847 году К. Рейхерт открыл способность белков образовывать кристаллы. В 1836 году Т. Шванн открыл пепсин - фермент, расщепляющий белки. В 1856 году Л. Корвизар открыл еще один подобный фермент - трипсин. Изучая действие этих ферментов на белки, биохимики пытались разгадать тайну пищеварения. Однако наибольшее внимание внимание привлекли вещества, получающиеся в результате действия на белки протелитических фермнтов (протеаз, к ним относятся вышеприведенные ферменты): одни из них были фрагментами исходных молекул белка (их назвали пептонами ), другие же не подвергались дальнейшему расщеплению протеазами и относились к известному еще с начала века классу соединений - аминокислот (первое аминокислотное производное - амид аспарагин был открыт в 1806 году, а первая аминокислота - цистин в 1810). Аминокислоты в составе белков впервые обнаружил в 1820 году французский химик А. Браконно. Он применил кислотный гидролиз белка и в гидролизате обнаружил сладковатое вещество, названное им глицином. В 1839 году было доказано существование в составе белков лейцина, а в 1849 году Ф. Бопп выделил из белка еще одну аминокислоту - тирозин (полный список дат открытий аминокислот в белках см. Приложение II).
К концу 80-х гг. XIX века из белковых гидролизатов было выделено уже 19 аминокислот и стало медленно укрепляться мнение, что сведения о продуктах гидролиза белков несут важную информацию о строении белковой молекулы. Тем не менее, аминокислоты считались обязательным, но неглавным компонентом белка.
В связи с открытиями аминокислот в составе белков французский ученый П. Шютценберже в 70-х гг. XIX века предложил т. н. уреидную теорию строения белка. Согласно ей молекула белка состояла из центрального ядра, роль которого выполняла молекула тирозина, и присоединенных к нему (с замещением 4 атомов водорода) слож ных группировок, названных Шютценберже лейцинами . Однако гипотеза было очень слабо подкреплена экспериментально, и дальнейшие исследования показали несостоятельность.
2.2 Теория “углеазотных комплексов” А.Я. Данилевского
Оригинальную теорию о строении белка высказал в 80-х гг. XIX века русский биохимик А. Я. Данилевский. Первым из химиков он обратил внимание на возможный полимерный характер строения белковых молекул. В начале 70-х гг. он писал А.М. Бутлерову, что “частицы альбумина есть смешанный полимерид”, что для определения белка он не находит “термина более подходящего, чем слово полимер в широком смысле”. Изучая биуретовую реакцию он предположил, что эта реакция связана со структурой перемежающихся атомов углерода и азота - N - C - N - C - N - , которые входят в т.н. углеазо т ный комплекс R" - NH - CO - NH - CO - R”. На основе данной формулы Данилевский полагал, что в молекуле белка содержится 40 таких углеазотных комплексов. Отдельные углеазотноаминокислотные комплекс, по Данилевскому, выглядели так:
По Данилевскому углеазотные комплексы могли соединяться эфирной или амидной связью с образованием высокомолекулярной структуры.
2.3 Теория “киринов” А. Косселя
Немецкий физиолог и биохимик А. Коссель, изучая протамины и гистоны, относительно просто устроенные белки, он установил, что при их гидролизе образуется большое количество аргинина. Кроме того он открыл в составе гидролизата неизвестную тогда аминокислоту - гистидин. На основании этого Коссель предположил, что эти белковые вещества можно рассматривать как некие простейшие модели более сложных белков, построенных, по его мнению, согласно следующему принципу: аргинин и гистидин составляют центральное ядро (“протаминовое ядро”), которое окружено комплексами из других аминокислот.
Теория Косселя представляла собой наиболее совершенный пример развития гипотезы о фрагментарном строении белков (впервые предложенной, как было сказано выше, Г.Мульдером). Этой гипотезой воспользовался немецкий химик М. Зигфрид в начале XX века. Он полагал, что белки построены из комплексов аминокислот (аргинин+лизин+глутаминовая к-та), названных им киринами (от греческого “кириос” основной). Однако эта гипотеза была высказана в 1903 году, когда Э. Фишер активно разрабатывал свою пептидную теорию , давшую ключ к тайне строения белков.
2.4 Пептидная теория Э. Фишера
Немецкий химик Эмиль Фишер, уже прославившийся на весь мир исследованиями пуриновых соединений (алкалоидов группы кофеина) и расшифровкой структуры сахаров, создал пептидную теорию, во многом подтвердившуюся практически и получившую всеобщее признание еще при его жизни, за что он был удостоен второй в истории химии Нобелевской премии (первую получил Я.Г. Вант-Гофф).
Немаловажно, что Фишер построил план исследования, резко отличающийся от того, что предпринималось раньше, однако учитывающий все известные на тот момент факты. Прежде всего он принял, как наиболее вероятную гипотезу о том, что белки построены из аминокислот, соединенных амидной связью:
Такой тип связи Фишер назвал (по аналогии с пептонами) пептидной . Он предположил, что белки представляют собой полимеры аминокислот, соединенных пептидной связью . Идея о полимерном характере строения белков как известно высказывалась еще Данилевским и Хертом, но они считали, что “мономеры” представляют собой очень сложные образования - пептоны или “углеазотные комплексы”.
Доказывая пептидный тип соединения аминокислотных остатков. Э. Фишер исходил из следующих наблюдений. Во-первых, и при гидролизе белков, и при их ферментативном разложении образовывались различные аминокислоты. Другие соединения было чрезвычайно трудно описать а еще труднее получить. Кроме того Фишеру было известно, что у белков не наблюдается преобладания ни кислотных, ни основных свойств, значит, рассуждал он, амино- и карбоксильные группы в составе аминокислот в белковых молекулах замыкаются и как бы маскируют друг друга (амфотерность белков, как сказали бы сейчас).
Решение проблемы строения белка Фишер разделил, сведя ее к следующим положениям:
Качественное и количественное определение продуктов полного гидролиза белков.
Установление строения этих конечных продуктов.
Синтез полимеров аминокислот с соединениями амидного (пептидного) типа.
Сравнение полученных таким образом соединений с природными белками.
Из этого плана видно, что Фишер применил впервые новый методологический подход - синтез модельных соединений, как способ доказательства по аналогии.
2.5 Разработка методов синтеза аминокислот
Для того чтобы перейти к синтезу производных аминокислот, соединенных пептидной связью, Фишер провел большую работу по изучению строения и синтезу аминокислот.
До Фишера общим методом синтеза аминокислот был циангидринный синтез А. Штреккера:
По реакции Штреккера удалось синтезировать аланин, серин и некоторые другие аминокислоты, а по ее модификации (реакции Зелинского-Стадникова) как -аминокислоты, так и их N-замещенные.
Однако сам Фишер стремился разработать методы синтеза всех известных тогда аминокислот. Он считал метод Штреккера недостаточно универсальным. Поэтому Э. Фишеру пришлось искать общий метод синтеза аминокислот в том числе аминокислот со сложными боковыми радикалами.
Он предложил аминировать бромзамещенные в -положении карбоновые кислоты. Для получения бромпроизводных он использовал, как например, в синтезе лейцина, арилированную или алкилированную малоновую кислоту:
Но создать абсолютно универсальный метод Э. Фишеру не удалось. Были разработаны и более надежные реакции. Например, ученик Фишера Г. Лейкс предложил следующую модификацию для получения серина:
Фишер также доказал, что белки состоят из остатков оптически активных аминокислот (см. стр.11). Это заставило его разработать новую номенклатуру оптически активных соединений, методы разделения и синтеза оптических изомеров аминокислот. Фишер также пришел к выводу, что в белках содержатся остатки L-форм оптически активных аминокислот, и он доказал это, впервые использовав принцип диастереоизомерии. Этот принцип заключался в следующем: к N-ацилпроизводному рацемической аминокислоты добавляли оптически активный алкалоид (бруцин, стрихнин, цинхонин, хинидин, хинин). В результате этого образовывались две стереоизомерные формы солей, обладающие различной растворимостью. После разделения этих диастереоизомеров алкалоид регенерировали и ацильную группу удаляли путем гидролиза.
Фишер сумел разработать метод полного определения аминокислот в продуктах гидролиза белков: он переводил хлоргидраты эфиров аминокислот обработкой концентрированной щелочью на холоду в свободные эфиры, которые заметно не омылялись. Затем смесь этих эфиров подвергал фракционной перегонке и из полученных фракций выделял отдельные аминокислоты путем дробной кристаллизации.
Новый метод анализа не только окончательно подтвердил, что белки состоят из аминокислотных остатков, но позволил уточнить и пополнить список встречающихся в белках аминокислот. Но все же количественные анализы не могли дать ответа на основной вопрос: каковы принципы строения молекулы белка. И Э.Фишер сформулировал одну из основных задач в изучении строения и свойств белка: разработка экспериментальные м е тоды синтеза соединений, основными компонентами которых были бы аминокисл о ты, соединенные пептидной связью.
Таким образом Фишер поставил нетривиальную задачу - синтезировать новый класс соединений с целью установления принципов их строения.
Задачу эту Фишер решил, и химики получили убедительные доказательства, что белки представляют собой полимеры аминокислот, соединенных пептидной связью:
CO - CHR" - NH - CO - CHR"" - NH - CO CHR""" - NH -
Это положение подтверждалось биохимическими доказательствами. Попутно выяснилось, что протеазы гидролизуют не все связи между аминокислотами с одинаковой скоростью. На их способность расщеплять пептидную связь влияли оптическая конфигурация аминокислот, заместители по азоту аминогруппы, длина цепи пептида, а также набор входящих в него остатков.
Главным доказательством пептидной теории стал синтез модельных пептидов и сопоставление их с пептонами гидролизата белков. Результаты показали, что из белковых гидролизатов выделяются пептиды, идентичные синтезированным.
В процессе выполнения этих исследований Э.Фишер и его ученик Э.Абдергальд- ен впервые разработали метод определения аминокислотной последовательности в белка. Сущность его заключалась в установлении природы аминокислотного остатка полипептида, имеющего свободную аминогруппу (N-концевую аминокислоту). Для этого они предложили блокировать в пептиде аминоконец -нафталин-сулфониловой группой, которая не отщепляется при гидролизе. Выделяя затем из гидролизата аминокислоту, меченую такой группой, можно было определить, какая из аминокислот была N-концевой.
После исследований Э.Фишера стало ясно, что белки представляют собой полипептиды. Это было важное достижение, в том числе и для задач синтеза белков: стало ясно, что именно нужно синтезировать. Только после этих работ проблема синтеза белка приобрела определенную направленность и необходимую строгость.
Говоря о работе Фишера в целом, следует отметить, что сам подход к исследованию был типичен скорее для наступающего XX века - он оперировал широким набором теоретических положений и методических приемов; его синтезы все менее и менее походили на искусство, основанное на интуиции, чем на точном знании, и приближались к созданию серий точных, почти технологических приемов.
2. 6 Кризис пептидной теории
В связи с применением новых физических и физико-химических методов исследований в начале 20-х гг. XX в. появились сомнения в том, что молекула белка представляет длинную полипептидную цепь. К гипотезе о возможности компактной укладки пептидных цепочек относились со скептицизмом. Все это потребовало пересмотра пептидной теории Э.Фишера.
В 20-30-е гг. распространение получила дикетопиперазиновая теория. Согласно ей, центральная роль в построении структуры белка играют дикетопиперазивные кольца, образующиеся при циклизации двух аминокислотных остатков. Также предполагалось, что эти структуры составляют центральное ядро молекулы, к которому присоединены короткие пептиды или аминокислоты (“наполнители” циклического скелета основной структуры). Наиболее убедительные схемы участия дикетопиперазинов в построении структуры белка были представлены Н.Д.Зелинским и учениками Э.Фишера.
Однако попытки синтеза модельных соединений, содержащих дикетопиперазины мало, что дали для химии белка впоследствии восторжествовала пептидная теория, однако эти работы оказали стимулирующее влияние на химию пиперазинов в целом.
После пептидной и дикетопиперазивной теорий продолжались попытки доказать существование только пептидных структур в молекуле белка. При этом стремились представить себе не только тип молекулы, но и общие ее очертания.
Оригинальную гипотезу высказал советский химик Д.Л.Талмуд. Он предположил, что пептидные цепи в составе белковых молекул свернуты в большие кольца, что в свою очередь стало шагом к созданию им представления о белковой глобуле.
Одновременно появились данные, свидетельствующие о различном наборе аминокислот в различных белка. Но закономерности, которым подчиняется последовательность аминокислот в структуре белка, были не ясны.
Первыми ответ на этот вопрос пытались дать М.Бергман и К.Ниман в разработанной ими гипотезе “перемежающихся частот”. Согласно ей последовательность аминокислотных остатков в белковой молекуле подчинялась числовым закономерностям, основы которых были выведены из принципов строения белковой молекулы фиброина шелка. Но этот выбор был неудачным, т.к. этот белок фибриллярный, строение же глобулярных белков подчиняется совсем другим закономерностям.
По М.Бергману и К.Ниману, каждая аминокислота встречается в полипептидной цепи через определенной интервал или, как говорил М.Бергман, обладает определенной “периодичностью”.эта периодичность определяется природой аминокислотных остатков.
Молекулу фиброина шелка они представляли себе следующим образом:
GlyAlaGlyTyr GlyAlaGlyArg GlyAlaGlyx GlyAlaGlyx
(GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyx) 12
GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyArg
(GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyx) 13
Гипотеза Бергмана-Нимана оказала значительное влияние на развитие химии аминокислот большое количество работ было посвящено ее проверке.
В заключение этой главы следует отметить, что к середине XX в. было накоплено достаточно доказательств справедливости пептидной теории, основные ее положения были дополнены и уточнены. Поэтому центр исследований белков в XX в. лежал уже области исследования и поиска методов синтеза белка искусственным путем. Эта задача была успешно решена, были разработаны надежные методы определения первичной структуры белка - последовательности аминокислот в пептидной цепи, разработаны методы химического (абиогенного) синтеза нерегулярных полипептидов (подробнее эти методы рассматриваются в гл.8, стр.36), в том числе методы автоматического синтеза полипептидов. Это позволило уже в 1962 г. крупнейшему английскому химику Ф.Сенгеру расшифровать структуру и синтезировать искусственным путем гормон инсулин, что ознаменовало новую эру в синтезе полипептидов функциональных белков.
ГЛАВА 3. ХИМИЧЕСКИЙ СОСТАВ БЕЛКОВ
3.1 Пептидная связь
Белки представляют собой нерегулярные полимеры, построенные из остатков -аминокислот, общую формулу которых в водном растворе при значениях pH близких к нейтральным можно записать как NH 3 + CHRCOO - . Остатки аминокислот в белках соединены между собой амидной связью между -амино- и -карбоксильными группами. Пептидная связь между двумя -аминокислотными остатками обычно называется пептидной связью , а полимеры, построенные из остатков -аминокислот, соединенных пептидными связями, называют полипептидами. Белок как биологически значимая структура может представлять собой как один полипептид, так и несколько полипептидов, образующих в результате нековалентных взаимодействий единый комплекс.
3.2 Элементный состав белков
Изучая химический состав белков, необходимо выяснить, во-первых, из каких химических элементов они состоят, во-вторых, - строение их мономеров. Для ответа на первый вопрос определяют количественный и качественный состав химических элементов белка. Химический анализ показал наличие во всех белках углерода (50-55%), кислорода (21-23%), азота (15-17%), водорода (6-7%), серы (0,3-2,5%). В составе отдельных белков обнаружены также фосфор, йод, железо, медь и некоторые другие макро- и микроэлементы, в различных, часто очень малых количествах.
Содержание основных химических элементов в белках может различаться, за исключением азота, концентрация которого характеризуется наибольшим постоянством и в среднем составляет 16%. Кроме того, содержание азота в других органических веществах мало. В соответствии с этим было предложено определять количество белка по входящему в его состав азоту. Зная, что 1г азота содержится в 6,25 г белка, найденное количество азота умножают коэффициент 6,25 и получают количество белка.
Для определения химической природы мономеров белка необходимо решить две задачи: разделить белок на мономеры и выяснить их химический состав. Расщепление белка на его составные части достигается с помощью гидролиза - длительного кипячения белка с сильными минеральными кислотами (кислотный гидролиз) или основаниями (щелочной гидролиз) . Наиболее часто применяется кипячение при 110 С с HCl в течение 24 ч. На следующем этапе разделяют вещества, входящие в состав гидролизата. Для этой цели применяют различные методы, чаще всего - хроматографию (подробнее - глава “Методы исследования…”). Главным частью разделенных гидролизатов оказываются аминокислоты.
3.3. Аминокислоты
В настоящее время в различных объектах живой природы обнаружено до 200 различных аминокислот. В организме человека их, например, около 60. Однако в состав белков входят только 20 аминокислот, называемых иногда природными.
Аминокислоты - это органические кислоты, у которых атом водорода -углеродного атома замещен на аминогруппу - NH 2 . Следовательно, по химической природе это -аминокислоты с общей формулой:
Из этой формулы видно, что в состав всех аминокислот входят следующие общие группировки: - CH 2 , - NH 2 , - COOH. Боковые же цепи (радикалы - R ) аминокислот различаются. Как видно из Приложения I химическая природа радикалов разнообразна: от атома водорода до циклических соединений. Именно радикалы определяют структурные и функциональные особенности аминокислот.
Все аминокислоты, кроме простейшей аминоуксусной к-ты глицина (NH 3 + CH 2 COO) имеют хиральный атом C и могут существовать в виде двух энантиомеров (оптических изомеров):
В состав всех изученных в настоящее время белков входят только аминокислоты L-ряда, у которых, если рассматривать хиральный атом со стороны атома H, группы NH 3 + , COO и радикал R расположены по часовой стрелке. Необходимость при построении биологически значимой полимерной молекулы строить ее из строго определенного энантиомера очевидна - из рацемической смеси двух энантиомеров получилась бы невообразимо сложная смесь диастереоизомеров. Вопрос, почему жизнь на Земле основана на белках, построеных именно из L-, а не D--аминокислот, до сих пор остается интригующей загадкой. Следует отметить, что D-аминокислоты достаточно широко распространены в живой природе и, более того, входят в состав биологически значимых олигопептидов.
Из двадцати основных -аминокислот строятся белки, однако остальные, достаточно разнообразные аминокислоты образуются из этих 20 аминокислотных остатков уже в составе белковой молекулы. Среди таких превращений следует в первую очередь отметить образование дисульфидных мостиков при окислении двух остатков цистеина в составе уже сформированных пептидных цепей. В результате образуется из двух остатков цистеина остаток диаминодикарбоновой кислоты цистина (см. Приложение I). При этом возникает сшивка либо внутри одной полипептидной цепи, либо между двумя различными цепями. В качестве небольшого белка, имеющего две полипептидные цепи, соединенный дисульфидными мостиками, а также сшивки внутри одной из полипептидных цепей:
Важным примером модификации аминокислотных остатков является превращение остатков пролина в остатки гидроксипролина :
Это превращение происходит, причем в значительном масштабе, при образовании важного белкового компонента соединительной ткани - коллагена .
Еще одним весьма важным видом модификации белков является фосфорилирование гидроксогрупп остатков серина, треонина и тирозина, например:
Аминокислоты в водном растворе находятся в ионизированном состоянии за счет диссоциации амино- и карбоксильных групп, входящих в состав радикалов. Другими словами, они являются амфотерными соединениями и могут существовать либо как кислоты (доноры протонов), либо как основания (акцепторы доноров).
Все аминокислоты в зависимости от структуры разделены на несколько групп:
Ациклические . Моноаминомонокарбоновые аминокислоты имеют в своем составе одну аминную и одну карбоксильную группы, в водном растворе они нейтральны. Некоторые из них имеют общие структурные особенности, что позволяет рассматривать их вместе:
Глицин и аланин. Глицин (гликокол или аминоуксусная к-та) является оптически неактивным - это единственная аминокислота, не имеющая энатиомеров. Глицин участвует в образовании нуклеиновых и желчных к-т, гема, необходим для обезвреживания в печени токсичных продуктов. Аланин используется организмом в различных процессах обмена углеводов и энергии. Его изомер -аланин является составной частью витамина пантотеновой к-ты, коэнзима А (КоА), экстрактивных веществ мышц.
Серин и треонин. Они относятся к группе гидрооксикислот, т.к. имеют гидроксильную группу. Серин входит в состав различных ферментов, основного белка молока - казеина, а также в состав многих липопротеинов. Треонин участвует в биосинтезе белка, являясь незаменимой аминокислотой.
Цистеин и метионин. Аминокислоты, имеющие в составе атом серы. Значение цистеина определяется наличием в ее составе сульфгидрильной (- SH) группы, которая придает ему способность легко окисляться и защищать организм о веществ с высокой окислительной способностью (при лучевом поражении, отравлении фосфором). Метионин характеризуется наличием легко подвижной метильной группы, использующейся для синтеза важных соединений в организме (холина, креатина, тимина, адреналина и др.)
Валин, лейцин и изолейцин. Представляют собой разветвленные аминокислоты, которые активно участвуют в обмене веществ и не синтезируются в организме.
Моноаминодикарбоновые аминокислоты имеют одну аминную и две карбоксильные группы и в водном растворе дают кислую реакцию. К ним относятся аспарагиновая и глутаминовая к-ты, аспарагин и глутамин. Они входят в состав тормозных медиаторов нервной системы.
Диаминомонокарбоновые аминокислоты в водном растворе имеют щелочную реакцию за сет наличия двух аминных групп. Относящийся к ним лизин необходим для синтеза гистонов а также в ряд ферментов. Аргинин участвует в синтезе мочевины, креатина.
Циклические . Эти аминокислоты имеют в своем составе ароматическое или гетероциклическое ядро и, как правило, не синтезируется в организме человека и должны поступать с пищей. Они активно участвуют в разнообразных обменных процессах. Так фенил-аланин служит основным источником синтеза тирозина - предшественника ряда биологически важных веществ: гормонов (тироксина, адреналина), некоторых пигментов. Триптофан помимо участия в синтезе белка, служит компонентом витамина PP, серотонина, триптамина, ряда пигментов. Гистидин необходим для синтеза белков, является предшественником гистамина, влияющего на кровяное давление и секрецию желудочного сока.
ГЛАВА 4. СТРУКТУРА
При изучении состава белков было установлено, что все они построены по единому принципу и имеют четыре уровня организации: первичную, вторичную, третичную, а отдельные из них и четвертичную структуры.
4.1 Первичная структура
Представляет собой линейную цепь аминокислот, расположенных в определенной последовательности и соединенных между собой пептидными связями. Пептидная связь образуется за счет -карбоксильной группы одной аминокислоты и -аминной группы другой:
Пептидная связь вследствие p, -сопряжения -связи карбонильной группы и р-орбитали атома N, на котором находится не поделенная пара электронов, не может рассматриваться как одинарная и вращение вокруг нее практически отсутствует. По этой же причине хиральный атом C и карбонильный атом C k любого i-го аминокислотного остатка пептидной цепи и атомы N и С (i+1)-го остатка находятся в одной плоскости. В этой же плоскости находятся карбонильный атом О и амидный атом Н (однако накопленный при изучении структуры белков материал показывает, что это утверждение не совсем строго: атомы, связанные с пептидным атомом азота, находятся не в одной плоскости с ним, а образуют трехгранную пирамиду с углами между связями, очень близкими к 120. Поэтому между плоскостями, образованными атомами C i , C i k , O i и N i +1 , H i +1 , C i +1 , существует некоторый угол, отличающийся от 0. Но, как правило, он не превышает 1 и не играет особой роли). Поэтому геометрически полипептидную цепочку можно рассматривать как образованную такими плоскими фрагментами, содержащими каждый по шесть атомов. Взаимное расположение этих фрагментов, как и всякое взаимное расположение двух плоскостей, должно определятся двумя углами. В качестве таковых принято брать торсионные углы, характеризующие вращения вокруг -связей N C и C C k .
Геометрия любой молекулы определяется тремя группами геометрических характеристик ее химических связей - длинами связей, валентными углами и торсионными углами между связями, примыкающими к соседним атомам. Первые две группы в решающей мере определяются природой участвующих атомов и образующихся связей. Поэтому пространственная структура полимеров в основном определяется торсионными углами между звеньями полимерного остова молекул, т.е. конформацией полимерной цепи. То р сионный угол , т.е. угол поворота связи А-В вокруг связи В-С относительно связи С- D , определяется как угол между плоскостями, содержащими атомы А, В, С и атомы B , C , D .
В такой системе возможен случай, когда связи А-В и С-D расположены параллельно и находятся по одну сторону от связи В-С. Если рассматривать эту систему вдоль св я зи В-С, то связь А-В как бы заслоняет связь C - D , поэтому такая конформация наз ы вается заслоненной. Согласно рекомендациям международных союзов химии IUPAC (International Union of Pure and Applied Chemistry) и IUB (International Union of Biochemistry), угол между плоскостями ABC и BCD считается положительным, если для приведения конформации в заслоненное состояние путем поворота на угол не выше 180 ближнюю к наблюдателю связь нужно поворачивать по часовой стрелке. Если эту связь для получения заслоненной конформации нужно поворачивать против часовой стрелки, то угол считается отрицательным. Можно заметить, что это определение не зависит от того, какая из связей находится ближе к наблюдателю.
При этом, как видно из рисунка, ориентация фрагмента, содержащего атомы C i -1 и C i [(i-1)-й фрагмент], и фрагмента, содержащего атомы C i и C i +1 (i-й фрагмент), определяется торсионными углами, соответствующими вращению вокруг связи N i C i и связи C i C i k . Эти углы принято обозначать как и, в приведенном случае соответственно i и i . Их значениями для всех мономерных звеньев полипептидной цепи в основном определяется геометрия этой цепи. Никаких однозначных величин ни для значения каждого из этих углов, ни для их комбинаций не существует, хотя на те и на другие накладываются ограничения, определяемые как свойствами самих пептидных фрагментов, так и природой боковых радикалов, т.е. природой аминокислотных остатков.
К настоящему времени установлены последовательности аминокислот для нескольких тысяч различных белков. Запись структуры белков в виде развернутых структурных формул громоздка и не наглядна. Поэтому используется сокращенная форма записи - трехбуквенная или однобуквенная (молекула вазопрессина):
При записи аминокислотной последовательности в полипептидных или олигопептидных цепях с помощью сокращенной символики предполагается, если это особо не оговорено, что -аминогруппа находится слева, а -карбоксильная группа - справа. Соответствующие участки полипептидной цепи называют N-концом (аминным концом) и С-концом (карбоксильным концом), а аминокислотные остатки - соответственно N-концевым и С-концевым остатками.
4.2 Вторичная структура
Фрагменты пространственной структуры биополимер, имеющие периодическое строение полимерного остова, рассматривают как элементы вторичной структуры.
Если на протяжении некоторого участка цепи однотипные углы, о которых говорилось на стр.15, приблизительно одинаковы, то структура полипептидной цепи приобретает периодический характер. Существует два класса таких структур - спиральные и растянутые (плоские или складчатые).
Спиральной считается структура, у которой все однотипные атомы лежат на одной винтовой линии. При этом спираль считается правой, если при наблюдении вдоль оси спирали она удаляется от наблюдателя по часовой стрелке, и левой - если удаляется против часовой стрелки. Полипептидная цепь имеет спиральную конформацию, если все атомы C находятся на одной винтовой линии, все карбонильные атомы C k - на другой, все атомы N - на третьей, причем шаг спирали для всех трех групп атомов должен быть одинаков. Одинаковым должно быть и число атомов, приходящихся на один виток спирали, независимо от того, идет ли речь об атомах C k , C или N. Расстояние же до общей винтовой линии для каждого из этих трех типов атомов свое.
Главными элементами вторичной структуры белков являются -спирали и -складки.
Спиральные структуры белка. Для полипептидных цепей известно несколько различных типов спиралей. Среди них наиболее распространена правая -спираль. Идеальная -спираль имеет шаг 0,54 нм и число однотипных атомов на один виток спирали 3,6, что означает полную периодичность на пяти витках спирали через каждые 18 аминокислотных остатков. Значения торсионных углов для идеальной -спирали = - 57 = - 47 , а расстояния от атомов, образующих полипептидную цепь, до оси спирали составляет для N 0,15 нм, для C 0,23 нм, для C k 0,17 нм. Любая конформация существует при условии, что имеются факторы, стабилизирующие ее. В случае -спирали такими факторами являются водородные связи, образуемые каждым карбонильным атомом (i+4)-го фрагмента. Важным фактором стабилизации -спирали также является параллельная ориентация дипольных моментов пептидных связей.
Складчатые структуры белка. Одним из распространенных примеров складчатой периодической структуры белка являются т.н. -складки , состоящие из двух фрагментов, каждый из которых представлен полипептидом.
Складки также стабилизируются водородными связями между атомом водорода аминной группы одного фрагмента и атомом кислорода карбоксильной группы другого фрагмента. При этом фрагменты могут иметь как параллельную, так и антипараллельную ориентацию относительно друг друга.
Структура, образующаяся в результате таких взаимодействий, представляет собой гофрированную структуру. Это сказывается на значениях торсионных углов и. Если в плоской, полностью растянутой структуре они должны были бы составить 180, то в реальных -слоях они имеют значения = - 119 и = + 113. Для того чтобы два участка полипептидной цепи располагались в ориентации, благоприятствующей образованию -складок, между ними должен существовать участок, имеющий структуру, резко отличающийся от периодической.
4.2.1 Факторы, влияющие на образование вторичной структуры
Структура определенного участка полипептидной цепи существенно зависит от структуры молекулы в целом. Факторы, влияющие на формирование участков с определенной вторичной структурой, весьма многообразны и далеко не во всех случаях полностью выявлены. Известно, что ряд аминокислотных остатков предпочтительно встречается в -спиральных фрагментах, ряд других - в -складках, некоторые аминокислоты - преимущественно в участках, лишенных периодической структуры. Вторичная структура в значительной степени определяется первичной структурой. В некоторых случаях физический смысл такой зависимости может быть понят из стереохимического анализа пространственной структуры. Например, как видно из рисунка в -спирали сближены не только боковые радикалы соседних вдоль цепи аминокислотных остатков, но и некоторые пары остатков, находящихся на соседних витках спирали, в первую очередь каждый (i+1)-й остаток с (i+4)-м и с (i+5)-м. Поэтому в положениях (i+1) и (i+2), (i+1) и (i+4), (i+1) и (i+5) -спиралей редко одновременно встречается два объемных радикала, таких, например, как боковые радикалы тирозина, триптофана, изолейцина. Еще менее совместимо со структурой спирали одновременное наличие трех объемных остатков в положениях (i+1), (i+2) и (i+5) или (i+1), (i+4) и (i+5). Поэтому такие комбинации аминокислот в -спиральных фрагментах являются редким исключением.
4.3 Третичная структура
Под этим термином понимают полную укладку в простанстве всей полипептидной цепи, включая укладку боковых радикалов. Полное представление о третичной структуре дают координаты всех атомов белка. Благодаря огромным успехом рентгеноструктурного анализа такие данные, за исключением координат атомов водорода получены для значительного числа белков. Это огромные массивы информации, хранящиеся в специальных банках данных на машиночитаемых носителях, и их обработка немыслима без применения быстродействующих компьютеров. Полученные на компьютерах координаты атомов дают полную информацию о геометрии полипептидной цепи, в том числе значения торсионных углов, что позволяет выявить спиральную структуру, -складки или нерегулярные фрагменты. Примером такого исследовательского подхода может служить следующая пространственная модель структуры фермента фосфоглицераткиназы:
Общая схема строения фосфоглицераткиназы. Для наглядности -спиральные участки представлены в виде цилиндров, а -складки - в виде лент со стрелкой, указывающей направление цепи от N-конца к С-концу. Линии - нерегулярные участки, соединяющие структурированные фрагменты.
Изображение полной структуры даже небольшой белковой молекулы на плоскости, будь то страница книги или экран дисплея мало информативно из-за чрезвычайно сложного строения объекта. Чтобы исследователь мог наглядно представлять простанственное строение молекул сложных веществ, используют методы трехмерной компьютерной графики, позволяющей выводить на дисплей отдельные части молекул и манипулировать с ними, в частности поворачивать их в нужных ракурсах.
Третичная структура формируется в результате нековалентных взаимодействий (электростатические, ионные, силы Ван-дер-Ваальса и др.) боковых радикалов, обрамляющих -спирали и -складки, и непериодических фрагментов полипептидной цепи. Среди связей, удерживающих третичную структуру следует отметить:
а) дисульфидный мостик (- S - S -)
б) сложноэфирный мостик (между карбоксильной группой и гидроксильной группой)
в) солевой мостик (между карбоксильной группой и аминогруппой)
г) водородные связи.
В соответствии с формой белковой молекулы, обусловленной третичной структурой, выделяют следующие группы белков:
Глобулярные белки. Пространственная структура этих белков в грубом приближении может быть представлена в виде шара или не слишком вытянутого эллипсоида - глоб у лы . Как правило, значительная часть полипептидной цепи таких белков формирует -спирали и -складки. Соотношение между ними может быть самым различным. Например, у миоглобина (подробнее о нем на стр.28) имеется 5 -спиральных сегментов и нет ни одной -складки. У иммуноглобулинов (подробнее на стр.42), наоборот, основными элементами вторичной структуры являются -складки, а -спирали вообще отсутствуют. В вышеприведенной структуре фосфоглицераткиназы и те и другие типы структур представлены примерно одинаково. В некоторых случаях, как это видно на примере фосфоглицераткиназы, отчетливо просматриваются две или более четко разделеннные в пространстве (но тем не менее, конечно, связанные пептидными мостиками) части - домены. Зачастую различные функциональные зоны белка разнесены по разным доменам.
Фибриллярные белки. Эти белки имеют вытянутую нитевидную форму, они выполняют в организме структурную функцию. В первичной структуре они имеют повторяющиеся участки и формируют достаточно однотипную для всей полипептидной цепи вторичнкю структуру. Так, белок -креатин (основной белковый компонент ногтей, волос, кожи) построен из протяженных -спиралей. Фиброин шелка состоит из периодически повторяющихся фрагментов Gly - Ala - Gly - Ser , образующими -складки. Существуют менее распростаненные элементы вторичной структуры, пример - полипептидные цепи коллагена, образующие левые спирали с параметрами, резко отличающимися от параметров -спиралей. В коллагеновых волокнах три спиральные полипептидные цепи скручены в единую правую суперспираль:
4.4 Четвертичная структура
В большинстве случаев для функционирования белков необходимо, чтобы несколько полимерных цепей были объединены в единый комплекс. Такой комплекс также рассматривается как белок, состоящий из нескольких субъединиц . Субъединичная структура часто фигурирует в научной литературе как четвертичная структура.
Белки, состоящие из нескольких субъединиц, широко распространены в природе. Классический пример - четвертичная структура гемоглобина (подробнее - стр.26). субъединицы принято обозначать греческими буквами. У гемоглобина имеется по две и субъединицы. Наличие нескольких субъединиц важно в функциональном отношении - это увеличивает степень насыщения кислородом. Четвертичную структуру гемоглобина обозначают как 2 2 .
Субъединичное строение свойственно многим ферментам, в первую очередь тем, которые выполняют сложные функции. Например, РНК-полимераза из E . coli имеет субъединичную структуру 2 ", т.е. построен из четырех разнотипных субъединиц, причем -субъединица продублирована. Этот белок выполняет сложные и разнообразные функции - инициирует ДНК, связывает субстраты - рибонуклеозидтрифосфаты, а также переносит нуклеотидные остатки на растущую полирибонуклеотидную цепь и некоторые другие функции.
Работа многих белков подвержена т.н. аллостерической регуляции - специальные соединения (эффекторы) “выключают” или “включают” работу активного центра фермента. Такие ферменты имеют специальные участки опознавания эффектора. И даже существуют специальные регуляторные субъединицы , в состав которых в том числе входят указанные участки. Классический пример - ферменты протеинкиназы, катализирующие перенос остатка фосфорной к-ты от молекулы АТФ на белки-субстраты.
ГЛАВА 5. СВОЙСТВА
Белки имеют высокую молекулярную массу, некоторые растворимы в воде, способны к набуханию, характеризуются оптической активностью, подвижностью в электрическом поле и некоторыми другими свойствами.
Белки активно вступают в химические реакции. Это свойство связано с тем, что аминокислоты, входящие в состав белков, содержат разные функциональные группы, способные реагировать с другими веществами. Важно, что такие взаимодействия происходят и внутри белковой молекулы, в результате чего образуется пептидная, водородная дисульфидная и другие виды связей. К радикалам аминокислот, а следовательно и белков, могут присоединяться различные соединения и ионы, что обеспечивает их транспорт по крови.
Белки являются высокомолекулярными соединениями. Это полимеры, состоящие из сотен и тысяч аминокислотных остатков - мономеров. Соответственно и молекулярная масса белков находится в пределах 10 000 - 1 000 000. Так, в составе рибонуклеазы (фермента, расщепляющего РНК) содержится 124 аминокислотных остатка и ее молекулярная масса составляет примерно 14 000. Миоглобин (белок мышц), состоящий из 153 аминокислотных остатков, имеет молекулярную массу 17 000, а гемоглобин - 64 500 (574 аминокислотных остатка). Молекулярные массы других белков более высокие: -глобулин (образует антитела) состоит из 1250 аминокислот и имеет молекулярную массу около 150 000, а молекулярная масса фермента глутаматдегидрогеназы превышает 1 000 000.
Определение молекулярной массы проводится различными методами: осмометрическим, гельфильтрационным, оптическим и др. однако наиболее точным является метод седиментации, предложенный Т. Сведбергом. Он основан на том, что при ультрацентрифугировании ускорением до 900 000 g скорость осаждения белков зависит от их молекулярной массы.
Важнейшим свойством белков является их способность проявлять как кислые так и основные, то есть выступать в роли амфотерных электролитов. Это обеспечивается за счет различных диссоциирующих группировок, входящих в состав радикалов аминокислот. Например, кислотные свойства белку придают карбоксильные группы аспарагиновой глутаминовой аминокислот, а щелочные - радикалы аргинина, лизина и гистидина. Чем больше дикарбоновых аминокислот содержится в белке, тем сильнее проявляются его кислотные свойства и наоборот.
Эти же группировки имеют и электрические заряды, формирующие общий заряд белковой молекулы. В белках, где преобладают аспарагиновая и глутаминовая аминокислоты, заряд белка будет отрицательным, избыток основных аминокислот придает положительный заряд белковой молекуле. Вследствие этого в электрическом поле белки будут передвигаться к катоду или аноду в зависимости от величины их общего заряда. Так, в щелочной среде (рН 7 - 14) белок отдает протон и заряжается отрицательно, тогда как в кислой среде (рН 1 - 7) подавляется диссоциация кислотных групп и белок становится катионом.
Таким образом, фактором, определяющим поведение белка как катиона или аниона, является реакция среды, которая определяется концентрацией водородных ионов и выражается величиной рН. Однако при определенных значениях рН число положительных и отрицательных зарядов уравнивается и молекула становится электронейтральной, т.е. она не будет перемещаться в электрическом поле. Такое значение рН среды определяется как изоэлектрическая точка белков. При этом белок находится в наименее устойчивом состоянии и при незначительных изменениях рН в кислую или щелочную сторону легко выпадает в осадок. Для большинства природных белков изоэлектрическая точка находится в слабокислой среде (рН 4,8 - 5,4), что свидетельствует о преобладании в их составе дикарбоновых аминокислот.
Свойство амфотерности лежит в основе буферных свойств белков и их участии в регуляции рН крови. Величина рН крови человека отличается постоянством и находится в пределах 7,36 - 7,4 , несмотря на различные вещества кислого или основного характера, регулярно поступающие с пищей или образующиеся в обменных процессах - следовательно существуют специальные механизмы регуляции кислотно-щелочного равновесия внутренней среды организма. К таким системам относится рассматриваемая в гл. “ Классификация” гемоглобиновая буферная система (стр.28). Изменение рН крови более чем на 0,07 свидетельствует о развитии патологического процесса. Сдвиг рН в кислую сторону называется ацидозом, а в щелочную - алкалозом.
Важное значение для организма имеет способность белков адсорбироватьь на своей поверхности некоторые вещества и ионы (гормоны, витамины, железо, медь), которые либо плохо растворимы в воде, либо являются токсичными (билирубин, свободные жирные кислоты). Белки транспортируют их по крови к местам дальнейших превращений или обезвреживания.
Водные растворы белков имеют свои особенности. Во-первых, белки обладают большим сродством к воде, т.е. они гидрофильны. Это значит, что молекулы белка, как заряженные частицы, притягивают к себе диполи воды, которые располагаются вокруг белковой молекулы и образуют водную или гидратную оболочку. Эта оболочка предохраняет молекулы белка от склеивания и выпадения в осадок. Величина гидратной оболочки зависит от структуры белка. Например, альбумины более легко связываются с молекулами воды и имеют относительно большую водную оболочку, тогда как глобулины, фибриноген присоединяют воду хуже, и гидратная оболочка и них меньше. Таким образом, устойчивость водного раствора белка определяется двумя факторами: наличием заряда белковой молекулы и находящейся вокруг нее водной оболочки. При удалении этих факторов белок выпадает в осадок. Данный процесс может быть обратимым и необратимым.
...Подобные документы
Белки (протеины) – высоко молекулярные, азотосодержащие природные органические вещества, молекулы которых построены из аминокислот. Строение белков. Классификация белков. Физико-химические свойства белков. Биологические функции белков. Фермент.
реферат , добавлен 15.05.2007
Основные особенности метаболических процессов. Обмен веществ и энергии. Общая характеристика, классификация, функции, химический состав и свойства белков, их биологическая роль в построении живой материи. Структурные и сложные белки. Способы их осаждения.
презентация , добавлен 24.04.2013
Физические и химические свойства, цветные реакции белков. Состав и строение, функции белков в клетке. Уровни структуры белков. Гидролиз белков, их транспортная и защитная роль. Белок как строительный материал клетки, его энергетическая ценность.
реферат , добавлен 18.06.2010
Физические, биологические и химические свойства белков. Синтез и анализ белков. Определение первичной, вторичной, третичной и четвертичной структуры белков. Денатурация, выделение и очистка белков. Использование белков в промышленности и медицине.
реферат , добавлен 10.06.2015
Белки - высокомолекулярные органические соединения, их аминокислотный состав. Определение свойств белков их составом и структурой белковой молекулы. Характеристика основных функций белков. Органоиды клетки и их функции. Клеточное дыхание и его строение.
контрольная работа , добавлен 24.06.2012
Понятие и структура белков, аминокислоты как их мономеры. Классификация и разновидности аминокислот, характер пептидной связи. Уровни организации белковой молекулы. Химические и физические свойства белков, методы их анализа и выполняемые функции.
презентация , добавлен 14.04.2014
Биологическая роль воды. Функции минеральных солей. Простые и сложные липиды. Уровни организации белков. Строительная, энергетическая, запасающая и регуляторная функции липидов. Структурная, каталитическая, двигательная, транспортная функции белков.
презентация , добавлен 21.05.2015
Аминокислотный состав белков в организмах, роль генетического кода. Комбинации из 20 стандартных аминокислот. Выделение белков в отдельный класс биологических молекул. Гидрофильные и гидрофобные белки. Принцип построения белков, уровень их организации.
творческая работа , добавлен 08.11.2009
Основные элементы и химический состав мышечной ткани. Виды белков саркоплазмы и миофибрилл, их содержание к общему количеству белков, молекулярная масса, распределение в структурных элементах мышцы. Их функции и роль организме. Строение молекулы миозина.
презентация , добавлен 14.12.2014
Белки как источники питания, их основные функции. Аминокислоты, участвующие в создании белков. Строение полипептидной цепи. Превращения белков в организме. Полноценные и неполноценные белки. Структура белка, химические свойства, качественные реакции.
Известно, что в основе живой материи лежат органические вещества – белки, жиры, углеводы и нуклеиновые кислоты. Но самое важное место среди этих веществ занимает белок.
Большинство известных науке веществ при нагревании переходят из твердого вещества в жидкое. Но есть вещества, которые, наоборот, при нагревании переходят в твердое состояние. Эти вещества объединил в отдельный класс французский химик Пьер Джозеф Маке в 1777 г. По аналогии с яичным белком, который сворачивается при нагревании, эти вещества были названы белками. Белки иначе называются протеинами. По-гречески протеин (протейос) означает «занимающий первое место». Это название белок получил в 1838 г., когда голландский биохимик Жерар Мюльдер написал, что жизнь на планете была бы невозможна без некоего вещества, которое является наиболее важным из всех известных науке веществ и которое обязательно присутствует абсолютно во всех растениях и животных. Это вещество Мюльдер назвал протеин.
Белок – это самое сложное вещество среди всех питательных веществ. В каждой клетке человеческого организма происходят химические реакции, в которых очень важную роль играет белок.
Из чего состоит белок

В состав белков входят: азот, кислород, водород, углерод. А вот другие питательные вещества азот не содержат.
Белок – это природный полимер. А полимеры – это вещества, молекулы которых содержат очень большое количество атомов. Ещё в XIX векерусский химик Александр Михайлович Бутлеров доказал, что если изменяется строение молекулы, то изменяются и свойства вещества. Основным строительным материалом белков являются аминокислоты. А в белках встречаются различные сочетания аминокислот. Следовательно, в природе существует большое разнообразие белков с различными свойствами. С помощью исследований обнаружили примерно 20 аминокислот, которые участвуют в создании белков.
Как происходит процесс образования молекулы белка

Аминокислоты присоединяются друг к другу последовательно. В результате этого процесса образуется цепочка, которая называется полипептид. Впоследствии полипептиды могут сворачиваться в спирали или принимать другую форму. Свойства белка зависят от состава аминокислот, от того, какое количество аминокислот участвует в синтезе, и в каком порядке эти аминокислоты присоединяются друг к другу. Например, в синтезе двух белков участвует одинаковое количество аминокислот, имеющих к тому же одинаковый состав. Но если эти аминокислоты будут располагаться в разной последовательности, то мы получим два абсолютно разных белка.
Если пептиды содержат не более 15 аминокислотных остатков, то они называются олигопептиды. А пептиды, содержащие до нескольких десятков тысяч или даже сотен тысяч аминокислотных остатков, называются белками. Молекула белка имеет компактную пространственную структуру. Эта структура может быть в виде волокон. Такие белки называются фибриллярными. Они являются строительными белками. Если молекула белка имеет структуру в виде шара, то белки называются глобулярными. К таким белкам относятся ферменты, антитела, некоторые гормоны.
В зависимости от того, какие аминокислоты входят в состав белков, белки бывают полноценные и неполноценные. В состав полноценных белков входит полный набор аминокислот. В неполноценных белках некоторые аминокислоты отсутствуют.
Белки также подразделяются на простые и сложные. Простые белки содержат только аминокислоты. В состав сложных белков кроме аминокислот входят ещё и металлы, углеводы, липиды, нуклеиновые кислоты.
Роль белков в организме человека

В организме человека белки выполняют различные функции.
1. Структурная . Белки входят в состав клеток всех тканей и органов.
2. Защитная . Белок интерферон синтезируется в организме для защиты от вирусов.
3. Двигательна я. Белок миозин участвует в процессе сокращения мышц.
4. Транспортная. Гемоглобин, являющийся белком, в составе эритроцитов участвует в переносе кислорода и углекислого газа.
5. Энергетическа я. В результате окисления молекул белков освобождается энергия, необходимая для жизнедеятельности организма.
6. Каталитическа я. Белки ферменты выступают в роли биологических катализаторов, увеличивающих скорость химических реакций в клетках.
7. Регуляторна я. Гормоны регулируют различные функции организма. Например, инсулин регулирует уровень сахара в крови.
В природе существует огромное количество белков, способных выполнять самые разнообразные функции. Но самая главная функция белков – поддержание жизни на Земле совместно с другими биомолекулами.
 Федор Успенский История Византийской империи
Федор Успенский История Византийской империи Энциклопедия сказочных героев: "Карлик нос"
Энциклопедия сказочных героев: "Карлик нос" Станкостроение в современной россии Среди российских предприятий станкостроения
Станкостроение в современной россии Среди российских предприятий станкостроения