Hemijska formula primarne strukture proteina. Fizičko-hemijska svojstva proteina
>> Hemija: Proteini
Proteini, ili proteinske supstance, su visokomolekularni (molekularna težina varira od 5-10 hiljada do 1 milion ili više) prirodni polimeri, čiji su molekuli izgrađeni od ostataka aminokiselina povezanih amidnom (peptidnom) vezom.
Proteini se nazivaju i proteini (od grčkog "protos" - prvi, važan). Broj aminokiselinskih ostataka u proteinskom molekulu uvelike varira i ponekad doseže nekoliko hiljada. Svaki protein ima svoju inherentnu sekvencu aminokiselinskih ostataka.
Proteini obavljaju različite biološke funkcije: katalitičke (enzimi), regulatorne (hormoni), strukturne (kolagen, fibroin), motoričke (miozin), transportne (hemoglobin, mioglobin), zaštitne (imunoglobulini, interferon), rezervne (kazein, albumin, gliadin) i drugi. Među proteinima postoje antibiotici i tvari koje imaju toksično djelovanje.
Proteini su osnova biomembrana, najvažnijeg dijela ćelije i ćelijskih komponenti. Oni igraju ključnu ulogu u životu ćelije, čineći, takoreći, materijalnu osnovu njene hemijske aktivnosti.
Izuzetno svojstvo proteina je samoorganizacija strukture, odnosno njegova sposobnost da spontano stvori specifičnu prostornu strukturu svojstvenu samo datom proteinu. U suštini, sve aktivnosti tijela (razvoj, kretanje, obavljanje raznih funkcija i još mnogo toga) povezane su s proteinskim supstancama (Sl. 36). Nemoguće je zamisliti život bez proteina.
Proteini su najvažnija komponenta ljudske i životinjske hrane, dobavljač aminokiselina koje su im potrebne.
Struktura
U prostornoj strukturi proteina od velike je važnosti priroda radikala (ostataka) R- u molekulima aminokiselina. Nepolarni radikali aminokiselina obično se nalaze unutar proteinske makromolekule i uzrokuju hidrofobne (vidi dolje) interakcije; polarni radikali koji sadrže ionogene (jonske) grupe obično se nalaze na površini proteinske makromolekule i karakteriziraju elektrostatičke (jonske) interakcije. Polarni nejonski radikali (na primjer, koji sadrže alkoholne OH grupe, amidne grupe) mogu se nalaziti i na površini i unutar proteinske molekule. Učestvuju u stvaranju vodoničnih veza.
U proteinskim molekulima, a-amino kiseline su međusobno povezane peptidnim (-CO-NH-) vezama:
Ovako konstruisani polipeptidni lanci ili pojedinačni delovi unutar polipeptidnog lanca mogu u nekim slučajevima biti dodatno međusobno povezani disulfidnim (-S-S-) vezama, ili, kako ih često nazivaju, disulfidnim mostovima.
Važnu ulogu u stvaranju strukture proteina igraju jonske (solne) i vodonične veze, kao i hidrofobna interakcija - posebna vrsta kontakta između hidrofobnih komponenti proteinskih molekula u vodenom mediju. Sve ove veze imaju različitu snagu i omogućavaju formiranje složene, velike proteinske molekule.
Uprkos razlici u strukturi i funkcijama proteinskih supstanci, njihov elementarni sastav neznatno varira (u % suhe mase): ugljenik - 51-53; kiseonik - 21,5-23,5; azot - 16,8-18,4; vodonik - 6,5-7,3; sumpor - 0,3-2,5. Neki proteini sadrže male količine fosfora, selena i drugih elemenata.
Redoslijed povezivanja aminokiselinskih ostataka u polipeptidnom lancu naziva se primarna struktura proteina (slika 37).
Molekul proteina može se sastojati od jednog ili više polipeptidnih lanaca, od kojih svaki sadrži različit broj aminokiselinskih ostataka. S obzirom na broj njihovih mogućih kombinacija, može se reći da je raznolikost proteina gotovo neograničena, ali ne postoje svi u prirodi. Ukupan broj različitih vrsta proteina u svim vrstama živih organizama je 10 10 -10 12 . Za proteine čija je struktura izuzetno složena, pored primarne, razlikuju se i viši nivoi strukturne organizacije: sekundarne, tercijarne, a ponekad i kvartarne strukture (tabela 9). Većina proteina ima sekundarnu strukturu, iako ne uvijek kroz cijeli polipeptidni lanac. Polipeptidni lanci sa određenom sekundarnom strukturom mogu biti različito raspoređeni u prostoru.
Ovaj prostorni raspored naziva se tercijarna struktura (Sl. 39)
U formiranju tercijarne strukture, pored vodoničnih veza, važnu ulogu imaju ionske i hidrofobne interakcije. Po prirodi "pakiranja" proteinske molekule razlikuju se globularni ili sferični i fibrilarni ili filamentozni proteini.
Za globularne proteine, a-helikalna struktura je karakterističnija, spirale su zakrivljene, "presavijene". Makromolekula ima sferni oblik. Otapaju se u vodi i fiziološkim rastvorima i formiraju koloidne sisteme. Većina proteina životinja, biljaka i mikroorganizama su globularni proteini. 
Za fibrilarne proteine karakterističnija je filamentozna struktura. Uglavnom se ne otapaju u vodi. Fibrilarni proteini obično obavljaju funkcije formiranja strukture. Njihova svojstva (snaga, sposobnost rastezanja) zavise od načina na koji su polipeptidni lanci pakovani. Primjer fibrilarnih proteina su proteini mišićnog tkiva (miozin), keratin (rožnato tkivo). U nekim slučajevima, pojedinačne proteinske podjedinice formiraju složene ansamble uz pomoć vodikovih veza, elektrostatičkih i drugih interakcija. U tom slučaju se formira kvartarna struktura proteina.
Ipak, treba još jednom napomenuti da primarna struktura igra izuzetnu ulogu u organizaciji viših proteinskih struktura. 
Klasifikacija
Postoji nekoliko klasifikacija proteina. Zasnovani su na različitim karakteristikama:
Stepen težine (jednostavno i složeno);
Oblik molekula (globularni i fibrilarni proteini);
Rastvorljivost u pojedinačnim rastvaračima (topljivi u vodi, rastvorljivi u razblaženim slanim rastvorima - albumini, rastvorljivi u alkoholu - prolamini, rastvorljivi u razblaženim alkalijama i kiselinama - glutelini);
Izvršena funkcija (na primjer, skladišteni proteini, skeletni, itd.).
Svojstva
Proteini su amfoterni elektroliti. Pri određenoj pH vrijednosti medija (naziva se izoelektrična tačka), broj pozitivnih i negativnih naboja u proteinskoj molekuli je isti. Ovo je jedno od glavnih svojstava proteina. Proteini su u ovom trenutku električno neutralni, a njihova topljivost u vodi je najniža. Sposobnost proteina da smanje rastvorljivost kada njihove molekule postanu električno neutralne koristi se za njihovo izolovanje iz rastvora, na primer, u tehnologiji dobijanja proteinskih proizvoda.
Hidratacija
Proces hidratacije podrazumijeva vezivanje vode bjelančevinama, dok oni ispoljavaju hidrofilna svojstva: bubre, povećavaju se njihova masa i volumen. Oticanje proteina je praćeno njegovim djelomičnim otapanjem. Hidrofilnost pojedinih proteina zavisi od njihove strukture. Hidrofilne amidne (-CO-NH-, peptidna veza), aminske (NH2) i karboksilne (COOH) grupe prisutne u sastavu i smještene na površini proteinske makromolekule privlače molekule vode, striktno ih orijentirajući na površini molekule . Hidracijska (vodena) ljuska koja okružuje proteinske globule sprečava agregaciju i sedimentaciju i, posljedično, doprinosi stabilnosti proteinskih otopina. Na izoelektričnoj tački, proteini imaju najmanju sposobnost da vežu vodu, hidrataciona ljuska oko proteinskih molekula je uništena, pa se kombinuju i formiraju velike agregate. Do agregacije proteinskih molekula dolazi i kada se dehidriraju nekim organskim rastvaračima, kao što je etil alkohol. To dovodi do taloženja proteina. Kada se pH medijuma promeni, proteinska makromolekula postaje naelektrisana, a njen kapacitet hidratacije se menja.
Sa ograničenim bubrenjem, koncentrirani proteinski rastvori formiraju složene sisteme zvane žele. Želei nisu tečni, elastični, imaju plastičnost, određenu mehaničku čvrstoću i mogu zadržati svoj oblik. Globularni proteini mogu se potpuno hidratizirati otapanjem u vodi (na primjer, mliječni proteini), formirajući otopine niske koncentracije. Hidrofilna svojstva proteina, odnosno njihova sposobnost bubrenja, formiranja želea, stabilizacije suspenzija, emulzija i pjena, od velikog su značaja u biologiji i prehrambenoj industriji. Vrlo pokretljiv žele, izgrađen uglavnom od proteinskih molekula, je citoplazma - polutečni sadržaj ćelije. Visoko hidratizirani žele - sirovi gluten izolovan iz pšeničnog tijesta, sadrži do 65% vode. Različita hidrofilnost proteina glutena jedan je od znakova koji karakterišu kvalitet pšeničnog zrna i brašna dobijenog od njega (tzv. jaka i slaba pšenica). Hidrofilnost proteina zrna i brašna igra važnu ulogu u skladištenju i preradi žitarica, u pečenju. Tijesto koje se dobiva u pekarskoj industriji je protein nabubren u vodi, koncentrovani žele koji sadrži zrna škroba.
Denaturacija proteina
U toku denaturacije, pod uticajem spoljašnjih faktora (temperatura, mehaničko delovanje, dejstvo hemijskih agenasa i niz drugih faktora), dolazi do promene sekundarne, tercijarne i kvartarne strukture proteinske makromolekule, tj. prostorna struktura. Primarna struktura, a samim tim i hemijski sastav proteina se ne mijenja. Fizička svojstva se mijenjaju: topljivost se smanjuje, sposobnost hidratacije, gubi se biološka aktivnost. Oblik proteinske makromolekule se mijenja, dolazi do agregacije. Istovremeno se povećava aktivnost nekih hemijskih grupa, olakšava se dejstvo proteolitičkih enzima na proteine, a samim tim i lakše se hidrolizira.
U prehrambenoj tehnologiji od posebnog je praktičnog značaja termička denaturacija proteina, čiji stepen zavisi od temperature, trajanja zagrevanja i vlažnosti. Ovo se mora imati na umu kada se razvijaju načini toplinske obrade prehrambenih sirovina, poluproizvoda, a ponekad i gotovih proizvoda. Procesi termičke denaturacije imaju posebnu ulogu u blanširanju biljnih sirovina, sušenju žitarica, pečenju kruha i dobivanju tjestenine. Denaturacija proteina može biti uzrokovana i mehaničkim djelovanjem (pritisak, trljanje, tresenje, ultrazvuk). Konačno, djelovanjem hemijskih reagensa (kiseline, lužine, alkohol, aceton) dolazi do denaturacije proteina. Sve ove tehnike se široko koriste u hrani i biotehnologiji.
Pjenjenje
Proces pjene se podrazumijeva kao sposobnost proteina da formiraju visokokoncentrirane tečno-gasne sisteme, nazvane pjene. Stabilnost pjene, u kojoj je protein sredstvo za napuhavanje, ne ovisi samo o njenoj prirodi i koncentraciji, već i o temperaturi. Proteini kao sredstva za pjenjenje imaju široku primjenu u konditorskoj industriji (marshmallow, marshmallow, soufflé). Struktura pjene ima kruh, a to utječe na njegova svojstva okusa.
Molekuli proteina pod uticajem brojnih faktora mogu biti uništeni ili stupiti u interakciju sa drugim supstancama i formirati nove proizvode. Za prehrambenu industriju mogu se razlikovati dva veoma važna procesa: 1) hidroliza proteina pod dejstvom enzima i 2) interakcija amino grupa proteina ili aminokiselina sa karbonilnim grupama redukujućih šećera. Pod uticajem proteaza - enzima koji kataliziraju hidrolitičko cijepanje proteina, potonji se razlažu na jednostavnije produkte (poli- i dipeptide) i na kraju na aminokiseline. Brzina hidrolize proteina zavisi od njegovog sastava, molekularne strukture, aktivnosti enzima i uslova.
Hidroliza proteina
Reakcija hidrolize sa stvaranjem aminokiselina općenito se može zapisati na sljedeći način: 
Sagorijevanje
4. Koje se reakcije mogu koristiti za prepoznavanje proteina?
5. Kakvu ulogu imaju proteini u životu organizama?
6. Prisjetite se iz opšteg kursa biologije koji proteini određuju imunološka svojstva organizama.
7. Recite nam o AIDS-u i prevenciji ove strašne bolesti.
8. Kako prepoznati proizvod od prirodne vune i umjetnih vlakana?
9. Napišite jednačinu reakcije za hidrolizu proteina sa općom formulom (-NH-CH-CO-) n.
l
R
Kakav je značaj ovog procesa u biologiji i kako se koristi u industriji?
10. Napišite jednadžbe reakcija koje se mogu koristiti za sljedeće prijelaze: etan -> etil alkohol -> octeni aldehid -> octena kiselina -> hloroctena kiselina -> aminooctena kiselina -> polipeptid.
slučajevi iz hemije, zadaci i rješenja, napomene sa lekcija
PROTEINI (proteini), klasa složenih spojeva koji sadrže dušik, najkarakterističnije i najvažnije (uz nukleinske kiseline) komponente žive tvari. Proteini obavljaju mnoge i različite funkcije. Većina proteina su enzimi koji kataliziraju kemijske reakcije. Mnogi hormoni koji regulišu fiziološke procese su takođe proteini. Strukturni proteini kao što su kolagen i keratin su glavne komponente koštanog tkiva, kose i noktiju. Kontraktilni proteini mišića imaju sposobnost da mijenjaju svoju dužinu koristeći hemijsku energiju za obavljanje mehaničkog rada. Proteini su antitijela koja vežu i neutraliziraju toksične tvari. Neki proteini koji mogu odgovoriti na vanjske utjecaje (svjetlo, miris) služe kao receptori u osjetilnim organima koji percipiraju iritaciju. Mnogi proteini koji se nalaze unutar ćelije i na ćelijskoj membrani obavljaju regulatorne funkcije.U prvoj polovini 19. vijeka mnogi hemičari, a među njima prvenstveno J. von Liebig, postepeno su došli do zaključka da su proteini posebna klasa azotnih jedinjenja. Naziv "proteini" (od grč.
protos prvi) predložio je 1840. holandski hemičar G. Mulder. FIZIČKA SVOJSTVA Proteini su bijeli u čvrstom stanju, ali bezbojni u otopini, osim ako ne nose neku hromofornu (obojenu) grupu, kao što je hemoglobin. Rastvorljivost različitih proteina u vodi uvelike varira. Takođe varira sa pH i koncentracijom soli u rastvoru, tako da se mogu izabrati uslovi pod kojima će jedan protein selektivno taložiti u prisustvu drugih proteina. Ova metoda "salanja" se široko koristi za izolaciju i pročišćavanje proteina. Pročišćeni protein se često taloži iz otopine u obliku kristala.U poređenju sa drugim jedinjenjima, molekularna težina proteina je veoma velika - od nekoliko hiljada do mnogo miliona daltona. Stoga, tokom ultracentrifugiranja, proteini se talože, i to različitim brzinama. Zbog prisutnosti pozitivno i negativno nabijenih grupa u proteinskim molekulima, one se kreću različitim brzinama u električnom polju. Ovo je osnova elektroforeze, metode koja se koristi za izolaciju pojedinačnih proteina iz složenih mješavina. Pročišćavanje proteina se takođe vrši hromatografijom.
HEMIJSKA SVOJSTVA Struktura. Proteini su polimeri, tj. molekule izgrađene poput lanaca od ponavljajućih monomernih jedinica, ili podjedinica, čiju ulogu igraju a -amino kiseline. Opća formula aminokiselina gdje je R atom vodika ili neku organsku grupu.Molekul proteina (polipeptidni lanac) može se sastojati od samo relativno malog broja aminokiselina ili nekoliko hiljada monomernih jedinica. Povezivanje aminokiselina u lancu je moguće jer svaka od njih ima dvije različite hemijske grupe: amino grupu sa osnovnim svojstvima,
NH2 i kiselu karboksilnu grupu, COOH. Obje ove grupe su povezane sa a - atom ugljenika. Karboksilna grupa jedne amino kiseline može formirati amidnu (peptidnu) vezu sa amino grupom druge amino kiseline: Nakon što su dvije aminokiseline povezane na ovaj način, lanac se može produžiti dodavanjem treće drugoj aminokiselini, i tako dalje. Kao što se može vidjeti iz gornje jednadžbe, kada se formira peptidna veza, oslobađa se molekul vode. U prisustvu kiselina, lužina ili proteolitičkih enzima, reakcija se odvija u suprotnom smjeru: polipeptidni lanac se cijepa na aminokiseline uz dodatak vode. Ova reakcija se naziva hidroliza. Hidroliza se odvija spontano, a energija je potrebna za spajanje aminokiselina u polipeptidni lanac.
Nakon što su dvije aminokiseline povezane na ovaj način, lanac se može produžiti dodavanjem treće drugoj aminokiselini, i tako dalje. Kao što se može vidjeti iz gornje jednadžbe, kada se formira peptidna veza, oslobađa se molekul vode. U prisustvu kiselina, lužina ili proteolitičkih enzima, reakcija se odvija u suprotnom smjeru: polipeptidni lanac se cijepa na aminokiseline uz dodatak vode. Ova reakcija se naziva hidroliza. Hidroliza se odvija spontano, a energija je potrebna za spajanje aminokiselina u polipeptidni lanac. Karboksilna grupa i amidna grupa (ili slična imidna grupa u slučaju aminokiselina prolina) prisutne su u svim aminokiselinama, ali razlike između aminokiselina su određene prirodom te grupe, ili "bočnog lanca", što je gore naznačeno slovom
R . Ulogu bočnog lanca može imati jedan atom vodika, kao u aminokiselini glicin, ili neka glomazna grupa, kao u histidinu i triptofanu. Neki bočni lanci su hemijski inertni, dok su drugi visoko reaktivni.Mnogo hiljada različitih aminokiselina se može sintetizirati, i mnogo različitih aminokiselina se javlja u prirodi, ali samo 20 vrsta aminokiselina se koristi za sintezu proteina: alanin, arginin, asparagin, asparaginska kiselina, valin, histidin, glicin, glutamin, glutamin kiselina, izoleucin, leucin, lizin, metionin, prolin, serin, tirozin, treonin, triptofan, fenilalanin i cistein (u proteinima, cistein može biti prisutan kao dimer
cistin). Istina, u nekim proteinima postoje i druge aminokiseline pored dvadesetak koje se redovno pojavljuju, ali one nastaju kao rezultat modifikacije bilo koje od dvadeset navedenih nakon što je ona uključena u protein.optička aktivnost. Sve aminokiseline, osim glicina, a Atom ugljika ima četiri različite grupe vezane. Što se tiče geometrije, četiri različite grupe se mogu pričvrstiti na dva načina, pa su prema tome moguće dvije konfiguracije, odnosno dva izomera, koji su međusobno povezani kao objekt na njegovu sliku u ogledalu, tj. kao leva ruka desno. Jedna konfiguracija se zove lijeva ili ljevoruka ( L ), a drugi desni, ili desnorotirajući ( D ), budući da se dva takva izomera razlikuju u smjeru rotacije ravnine polarizirane svjetlosti. Nalazi se samo u proteinima L -aminokiseline (izuzetak je glicin; može se predstaviti samo u jednom obliku, pošto su dve od četiri njegove grupe iste), i sve imaju optičku aktivnost (pošto postoji samo jedan izomer). D -aminokiseline su rijetke u prirodi; nalaze se u nekim antibioticima i ćelijskom zidu bakterija.Redoslijed aminokiselina. Aminokiseline u polipeptidnom lancu nisu raspoređene nasumično, već u određenom fiksnom redoslijedu i upravo taj redoslijed određuje funkcije i svojstva proteina. Promjenom redoslijeda od 20 vrsta aminokiselina, možete dobiti ogroman broj različitih proteina, kao što možete sastaviti mnogo različitih tekstova od slova abecede.U prošlosti je određivanje sekvence aminokiselina proteina često trajalo nekoliko godina. Direktno određivanje je još uvijek prilično naporan zadatak, iako su stvoreni uređaji koji omogućavaju da se izvrši automatski. Obično je lakše odrediti nukleotidnu sekvencu odgovarajućeg gena i iz njega izvući sekvencu aminokiselina proteina. Do danas su aminokiselinske sekvence mnogih stotina proteina već određene. Funkcije dekodiranih proteina su obično poznate, a to pomaže da se zamisle moguće funkcije sličnih proteina nastalih, na primjer, u malignim neoplazmama.
Kompleksni proteini. Proteini koji se sastoje samo od aminokiselina nazivaju se jednostavnim. Međutim, često je atom metala ili neko hemijsko jedinjenje koje nije aminokiselina vezano za polipeptidni lanac. Takvi proteini se nazivaju kompleksni. Primjer je hemoglobin: sadrži željezni porfirin, koji mu daje crvenu boju i omogućava mu da djeluje kao prijenosnik kisika.Nazivi najsloženijih proteina sadrže indikaciju prirode vezanih grupa: šećeri su prisutni u glikoproteinima, masti u lipoproteinima. Ako katalitička aktivnost enzima ovisi o spojenoj grupi, onda se to naziva prostetičkom grupom. Često neki vitamin igra ulogu protetske grupe ili je njen dio. Vitamin A, na primjer, vezan za jedan od proteina retine, određuje njegovu osjetljivost na svjetlost.
Tercijarna struktura. Ono što je važno nije toliko aminokiselinska sekvenca proteina (primarna struktura), već način na koji je položen u svemiru. Po cijeloj dužini polipeptidnog lanca ioni vodika formiraju pravilne vodikove veze, koje mu daju oblik spirale ili sloja (sekundarna struktura). Iz kombinacije takvih spirala i slojeva nastaje kompaktni oblik sljedećeg reda - tercijarna struktura proteina. Oko veza koje drže monomerne karike lanca moguće su rotacije pod malim uglovima. Stoga, sa čisto geometrijske tačke gledišta, broj mogućih konfiguracija za bilo koji polipeptidni lanac je beskonačno velik. U stvarnosti, svaki protein normalno postoji u samo jednoj konfiguraciji, određenoj njegovom aminokiselinskom sekvencom. Ova struktura nije kruta, ona je, takoreći, « diše” fluktuira oko određene prosječne konfiguracije. Lanac je presavijen u konfiguraciju u kojoj je slobodna energija (sposobnost obavljanja rada) minimalna, kao što je oslobođena opruga komprimirana samo do stanja koje odgovara minimumu slobodne energije. Često je jedan dio lanca čvrsto vezan za drugi disulfid ( SS) veze između dva cisteinska ostatka. To je dijelom razlog zašto cistein među aminokiselinama igra posebno važnu ulogu.Složenost strukture proteina je toliko velika da još nije moguće izračunati tercijarnu strukturu proteina, čak i ako je poznata njegova aminokiselinska sekvenca. Ali ako je moguće dobiti proteinske kristale, onda se njegova tercijarna struktura može odrediti difrakcijom X zraka.
U strukturnim, kontraktilnim i nekim drugim proteinima, lanci su izduženi i nekoliko blago presavijenih lanaca koji leže jedan pored drugog formiraju fibrile; fibrile se zauzvrat savijaju u veće formacije vlakana. Međutim, većina proteina u rastvoru je globularna: lanci su umotani u kuglu, poput pređe u klupko. Slobodna energija sa ovom konfiguracijom je minimalna, budući da su hidrofobne ("vodoodbojne") aminokiseline skrivene unutar globule, a hidrofilne ("privlače vodu") aminokiseline su na njenoj površini.
Mnogi proteini su kompleksi nekoliko polipeptidnih lanaca. Ova struktura se naziva kvartarna struktura proteina. Molekul hemoglobina, na primjer, sastoji se od četiri podjedinice, od kojih je svaka globularni protein.
Strukturni proteini zbog svoje linearne konfiguracije formiraju vlakna u kojima je vlačna čvrstoća vrlo visoka, dok globularna konfiguracija omogućava proteinima da uđu u specifične interakcije sa drugim spojevima. Na površini globule, uz pravilno polaganje lanaca, pojavljuju se šupljine određenog oblika, u kojima se nalaze reaktivne kemijske grupe. Ako je dati protein enzim, onda drugi, obično manji, molekul neke supstance ulazi u takvu šupljinu, baš kao što ključ ulazi u bravu; u ovom slučaju se konfiguracija elektronskog oblaka molekula mijenja pod utjecajem kemijskih grupa koje se nalaze u šupljini, a to ga prisiljava da reagira na određeni način. Na taj način enzim katalizuje reakciju. Molekuli antitijela također imaju šupljine u koje se vežu različite strane tvari i na taj način postaju bezopasne. Model "ključ i brava", koji objašnjava interakciju proteina sa drugim spojevima, omogućava razumijevanje specifičnosti enzima i antitijela, tj. njihova sposobnost da reaguju samo sa određenim jedinjenjima.
Proteini u različitim vrstama organizama. Proteini koji obavljaju istu funkciju u različitim biljnim i životinjskim vrstama i stoga nose isto ime također imaju sličnu konfiguraciju. Oni se, međutim, donekle razlikuju u svojoj sekvenci aminokiselina. Kako se vrste razlikuju od zajedničkog pretka, neke aminokiseline na određenim pozicijama zamjenjuju se mutacijama s drugim. Štetne mutacije koje uzrokuju nasljedne bolesti prirodnom selekcijom odbacuju se, ali se korisne ili barem neutralne mogu sačuvati. Što su dvije biološke vrste bliže jedna drugoj, to je manje razlika u njihovim proteinima.Neki proteini se mijenjaju relativno brzo, drugi su prilično konzervativni. Potonji uključuju, na primjer, citokrom With respiratorni enzim koji se nalazi u većini živih organizama. Kod ljudi i čimpanzi, njegove aminokiselinske sekvence su identične, au citokromu With pšenice, pokazalo se da je samo 38% aminokiselina drugačije. Čak i poredeći ljude i bakterije, sličnost citokroma With(razlike utječu na 65% aminokiselina ovdje) se još uvijek mogu vidjeti, iako je zajednički predak bakterija i ljudi živio na Zemlji prije otprilike dvije milijarde godina. Danas se poređenje aminokiselinskih sekvenci često koristi za izgradnju filogenetskog (genealoškog) stabla koje odražava evolucijske odnose između različitih organizama.
Denaturacija. Sintetizirani proteinski molekul, savijajući se, poprima vlastitu konfiguraciju. Ova se konfiguracija, međutim, može uništiti zagrijavanjem, promjenom pH, djelovanjem organskih rastvarača, pa čak i jednostavnim miješanjem otopine dok se na njenoj površini ne pojave mjehurići. Protein izmijenjen na ovaj način naziva se denaturiranim; gubi svoju biološku aktivnost i obično postaje nerastvorljiv. Dobro poznati primjeri denaturiranih proteina su kuhana jaja ili šlag. Mali proteini, koji sadrže samo stotinjak aminokiselina, sposobni su da se renaturiraju, tj. ponovo nabavite originalnu konfiguraciju. Ali većina proteina se jednostavno transformiše u masu zapletenih polipeptidnih lanaca i ne obnavlja svoju prethodnu konfiguraciju.Jedna od glavnih poteškoća u izolaciji aktivnih proteina je njihova ekstremna osjetljivost na denaturaciju. Ovo svojstvo proteina nalazi korisnu primjenu u konzerviranju prehrambenih proizvoda: visoka temperatura nepovratno denaturira enzime mikroorganizama i mikroorganizmi umiru.
SINTEZA PROTEINA Za sintezu proteina, živi organizam mora imati sistem enzima sposobnih da vežu jednu aminokiselinu za drugu. Potreban je i izvor informacija koji bi odredio koje aminokiseline treba povezati. Budući da u tijelu postoje hiljade vrsta proteina, a svaki od njih se sastoji u prosjeku od nekoliko stotina aminokiselina, potrebne informacije moraju biti zaista ogromne. Pohranjuje se (slično kao što se zapis čuva na magnetnoj traci) u molekulima nukleinske kiseline koji čine gene. Cm . takođe NASLJEDNOST; NUKLEINSKE KISELINE.Aktivacija enzima. Polipeptidni lanac sintetiziran iz aminokiselina nije uvijek protein u svom konačnom obliku. Mnogi enzimi se prvo sintetiziraju kao neaktivni prekursori i postaju aktivni tek nakon što drugi enzim ukloni nekoliko aminokiselina s jednog kraja lanca. Neki od probavnih enzima, kao što je tripsin, sintetiziraju se u ovom neaktivnom obliku; ovi enzimi se aktiviraju u probavnom traktu kao rezultat uklanjanja terminalnog fragmenta lanca. Hormon insulin, čija se molekula u svom aktivnom obliku sastoji od dva kratka lanca, sintetiše se u obliku jednog lanca, tzv. proinsulin. Zatim se srednji dio ovog lanca uklanja, a preostali fragmenti se vežu jedni za druge, formirajući aktivni molekul hormona. Složeni proteini nastaju tek nakon što je određena hemijska grupa vezana za protein, a za to vezivanje često je potreban i enzim.Metabolička cirkulacija. Nakon hranjenja životinje aminokiselinama označenim radioaktivnim izotopima ugljika, dušika ili vodika, oznaka se brzo ugrađuje u njene proteine. Ako označene aminokiseline prestanu da ulaze u tijelo, tada se količina oznaka u proteinima počinje smanjivati. Ovi eksperimenti pokazuju da se dobijeni proteini ne pohranjuju u tijelu do kraja života. Svi su, uz nekoliko izuzetaka, u dinamičnom stanju, neprestano se razlažu do aminokiselina, a zatim ponovo sintetiziraju.Neki proteini se razgrađuju kada ćelije umru i budu uništene. To se stalno događa, na primjer, s crvenim krvnim zrncima i epitelnim stanicama koje oblažu unutrašnju površinu crijeva. Pored toga, razgradnja i resinteza proteina se dešavaju iu živim ćelijama. Začudo, manje se zna o razgradnji proteina nego o njihovoj sintezi. Ono što je, međutim, jasno je da su proteolitički enzimi uključeni u razgradnju, slično onima koji razlažu proteine u aminokiseline u probavnom traktu.
Poluživot različitih proteina je različit od nekoliko sati do više mjeseci. Jedini izuzetak je molekul kolagena. Jednom formirane, ostaju stabilne i ne obnavljaju se ili zamjenjuju. Vremenom se, međutim, mijenjaju neka njihova svojstva, a posebno elastičnost, a kako se ne obnavljaju, rezultat toga su određene promjene vezane za starenje, na primjer, pojava bora na koži.
sintetički proteini. Hemičari su odavno naučili kako da polimeriziraju aminokiseline, ali se aminokiseline nasumično kombinuju, tako da proizvodi takve polimerizacije malo liče na prirodne. Istina, moguće je kombinirati aminokiseline određenim redoslijedom, što omogućava dobivanje nekih biološki aktivnih proteina, posebno inzulina. Proces je prilično kompliciran i na taj način se mogu dobiti samo oni proteini čiji molekuli sadrže oko stotinu aminokiselina. Umjesto toga, poželjno je sintetizirati ili izolirati nukleotidnu sekvencu gena koja odgovara željenoj aminokiselinskoj sekvenci, a zatim uvesti ovaj gen u bakteriju, koja će replikacijom proizvesti veliku količinu željenog proizvoda. Ova metoda, međutim, ima i svoje nedostatke. Cm . Vidi također GENETIČKI INŽENJERING. PROTEINI I ISHRANA Kada se proteini u tijelu razgrade na aminokiseline, ove aminokiseline se mogu ponovo koristiti za sintezu proteina. Istovremeno, same aminokiseline su podložne raspadanju, tako da se ne koriste u potpunosti. Takođe je jasno da tokom rasta, trudnoće i zarastanja rana, sinteza proteina mora biti veća od razgradnje. Tijelo kontinuirano gubi neke proteine; to su proteini kose, noktiju i površinskog sloja kože. Stoga, za sintezu proteina, svaki organizam mora dobiti aminokiseline iz hrane. Zelene biljke se sintetiziraju iz CO 2 , voda i amonijak ili nitrati su svih 20 aminokiselina koje se nalaze u proteinima. Mnoge bakterije su također sposobne sintetizirati aminokiseline u prisustvu šećera (ili nekog ekvivalenta) i fiksiranog dušika, ali šećer u konačnici isporučuju zelene biljke. Kod životinja je sposobnost sinteze aminokiselina ograničena; dobijaju aminokiseline jedući zelene biljke ili druge životinje. U probavnom traktu apsorbirani proteini se razlažu na aminokiseline, potonje se apsorbiraju i od njih se izgrađuju proteini karakteristični za dati organizam. Nijedan od apsorbiranih proteina nije ugrađen u strukture tijela kao takve. Jedini izuzetak je da kod mnogih sisara dio majčinih antitijela može proći netaknut kroz placentu u fetalnu cirkulaciju, a kroz majčino mlijeko (posebno kod preživara) se prenijeti na novorođenče odmah nakon rođenja.Potreba za proteinima. Jasno je da za održavanje života tijelo mora primiti određenu količinu proteina iz hrane. Međutim, veličina ove potrebe ovisi o brojnim faktorima. Tijelu je potrebna hrana i kao izvor energije (kalorije) i kao materijal za izgradnju svojih struktura. Na prvom mjestu je potreba za energijom. To znači da kada u prehrani ima malo ugljikohidrata i masti, dijetetski proteini se ne koriste za sintezu vlastitih proteina, već kao izvor kalorija. Uz produženo gladovanje, čak se i vaši vlastiti proteini troše za zadovoljavanje energetskih potreba. Ako u ishrani ima dovoljno ugljikohidrata, onda se unos proteina može smanjiti.balans azota. U prosjeku cca. 16% ukupne mase proteina je dušik. Kada se aminokiseline koje čine proteine razgrađuju, dušik koji se nalazi u njima izlučuje se iz tijela mokraćom i (u manjoj mjeri) fecesom u obliku različitih azotnih spojeva. Stoga je prikladno koristiti takav pokazatelj kao što je ravnoteža dušika za procjenu kvalitete proteinske prehrane, tj. razlika (u gramima) između količine dušika unesenog u tijelo i količine azota izlučenog dnevno. Uz normalnu ishranu odrasle osobe, ove količine su jednake. U rastućem organizmu količina izlučenog azota je manja od količine dolaznog, tj. bilans je pozitivan. Sa nedostatkom proteina u ishrani, balans je negativan. Ako u ishrani ima dovoljno kalorija, a proteini u njoj potpuno nedostaju, tijelo štedi proteine. Istovremeno, metabolizam proteina se usporava, a ponovno korištenje aminokiselina u sintezi proteina odvija se što je moguće efikasnije. Međutim, gubici su neizbježni, a dušični spojevi se i dalje izlučuju urinom, a dijelom i izmetom. Količina azota koji se dnevno izluči iz organizma tokom proteinskog gladovanja može poslužiti kao mjera dnevnog nedostatka proteina. Prirodno je pretpostaviti da je unošenjem u ishranu količine proteina koja je ekvivalentna ovom nedostatku, moguće vratiti ravnotežu dušika. Međutim, nije. Nakon što primi ovu količinu proteina, tijelo počinje manje efikasno da koristi aminokiseline, pa je potreban dodatni protein da bi se uspostavila ravnoteža dušika.Ako količina proteina u prehrani premašuje ono što je potrebno za održavanje ravnoteže dušika, čini se da od toga nema štete. Višak aminokiselina jednostavno se koristi kao izvor energije. Posebno upečatljiv primjer su Eskimi, koji konzumiraju malo ugljikohidrata i oko deset puta više proteina nego što je potrebno za održavanje ravnoteže dušika. U većini slučajeva, međutim, korištenje proteina kao izvora energije nije korisno, jer možete dobiti mnogo više kalorija iz određene količine ugljikohidrata nego iz iste količine proteina. U siromašnim zemljama stanovništvo prima potrebne kalorije iz ugljikohidrata i konzumira minimalnu količinu proteina.
Ako tijelo dobije potreban broj kalorija u obliku neproteinskih proizvoda, tada je minimalna količina proteina koja održava ravnotežu dušika cca. 30 g dnevno. Otprilike isto toliko proteina sadrži četiri kriške hljeba ili 0,5 litara mlijeka. Nešto veća količina se obično smatra optimalnom; preporučuje se od 50 do 70 g.
Esencijalne aminokiseline. Do sada se protein smatrao kao cjelina. U međuvremenu, da bi se odvijala sinteza proteina, u tijelu moraju biti prisutne sve potrebne aminokiseline. Neke od aminokiselina koje tijelo same životinje može sintetizirati. Nazivaju se zamjenjivim jer ne moraju biti prisutni u ishrani, važno je samo da je općenito dovoljan unos proteina kao izvora dušika; tada, uz nedostatak neesencijalnih aminokiselina, tijelo ih može sintetizirati na račun onih koje su prisutne u višku. Preostale "esencijalne" aminokiseline se ne mogu sintetizirati i moraju se unijeti hranom. Esencijalni za ljude su valin, leucin, izoleucin, treonin, metionin, fenilalanin, triptofan, histidin, lizin i arginin. (Iako se arginin može sintetizirati u tijelu, smatra se esencijalnom aminokiselinom jer novorođenčad i djeca u rastu proizvode ga nedovoljne količine. S druge strane, za osobu zrele dobi unos neke od ovih aminokiselina iz hrane može postati opciono.)Ova lista esencijalnih aminokiselina je približno ista kod drugih kralježnjaka, pa čak i kod insekata. Nutritivna vrijednost proteina se obično određuje hranjenjem pacova koji rastu i praćenjem povećanja tjelesne težine životinja.
Nutritivna vrijednost proteina. Nutritivnu vrijednost proteina određuje esencijalna aminokiselina koja je najnedostatnija. Ilustrirajmo to primjerom. Proteini našeg tijela sadrže u prosjeku cca. 2% triptofana (po težini). Recimo da ishrana uključuje 10 g proteina koji sadrže 1% triptofana i da u njoj ima dovoljno drugih esencijalnih aminokiselina. U našem slučaju, 10 g ovog defektnog proteina je u suštini ekvivalentno 5 g potpunog; preostalih 5 g može poslužiti samo kao izvor energije. Imajte na umu da, budući da se aminokiseline praktički ne pohranjuju u tijelu, a da bi se odvijala sinteza proteina, sve aminokiseline moraju biti prisutne istovremeno, efekat unosa esencijalnih aminokiselina može se otkriti samo ako sve uđu u telo u isto vreme.. Prosječan sastav većine životinjskih bjelančevina je blizak prosječnom sastavu bjelančevina ljudskog tijela, pa je malo vjerovatno da ćemo se suočiti s nedostatkom aminokiselina ako je naša ishrana bogata namirnicama poput mesa, jaja, mlijeka i sira. Međutim, postoje proteini, poput želatine (proizvod denaturacije kolagena), koji sadrže vrlo malo esencijalnih aminokiselina. Biljni proteini, iako su u tom smislu bolji od želatine, takođe su siromašni esencijalnim aminokiselinama; posebno malo u njima lizina i triptofana. Ipak, čisto vegetarijanska ishrana nije nimalo štetna, osim ako ne konzumira nešto veću količinu biljnih proteina, dovoljnu da obezbedi organizam esencijalnim aminokiselinama. Najviše proteina nalazi se u biljkama u sjemenkama, posebno u sjemenu pšenice i raznih mahunarki. Mladi izdanci, kao što su špargle, takođe su bogati proteinima.Sintetički proteini u ishrani. Dodavanjem malih količina sintetičkih esencijalnih aminokiselina ili proteina bogatih njima nepotpunim proteinima, kao što su proteini kukuruza, moguće je značajno povećati nutritivnu vrijednost ovih potonjih, tj. čime se povećava količina konzumiranih proteina. Druga mogućnost je uzgoj bakterija ili kvasaca na naftnim ugljovodonicima uz dodatak nitrata ili amonijaka kao izvora dušika. Mikrobni proteini dobijeni na ovaj način mogu poslužiti kao hrana za živinu ili stoku, ili ih ljudi mogu direktno konzumirati. Treća, široko korištena, metoda koristi fiziologiju preživara. Kod preživača se u početnom dijelu želuca javlja tzv. U buragu žive posebni oblici bakterija i protozoa koji defektne biljne proteine pretvaraju u potpunije mikrobne proteine, a ovi se, nakon probave i apsorpcije, pretvaraju u životinjske proteine. Urea, jeftino sintetičko jedinjenje koje sadrži dušik, može se dodati u stočnu hranu. Mikroorganizmi koji žive u buragu koriste dušik uree za pretvaranje ugljikohidrata (kojih ima mnogo više u hrani) u proteine. Otprilike trećina ukupnog dušika u stočnoj hrani može doći u obliku uree, što u suštini znači, u određenoj mjeri, hemijsku sintezu proteina. U SAD-u ova metoda igra važnu ulogu kao jedan od načina dobivanja proteina.LITERATURA Murray R, Grenner D, Meyes P, Rodwell W. ljudska biohemija, tt. 12. M., 1993Alberts B., Bray D., Lewis J. et al. Molekularna biologija ćelije, tt. 13. M., 1994
Među organskim materijama vjeverice, ili proteini, su najbrojniji, najraznovrsniji i od najveće važnosti biopolimeri. Oni računaju 50 - 80% suva težina ćelije.
Molekuli proteina su veliki, zbog čega se nazivaju makromolekule. Osim ugljika, kisika, vodika i dušika, proteini mogu sadržavati sumpor, fosfor i željezo. Proteini se međusobno razlikuju po broju (od sto do nekoliko hiljada), sastavu i redoslijedu monomera. Proteinski monomeri su aminokiseline (slika 1)
Beskrajna raznolikost proteina nastaje različitim kombinacijama svega 20 amino kiseline. Svaka aminokiselina ima svoje ime, posebnu strukturu i svojstva. Njihova opšta formula može se predstaviti na sledeći način:

Molekul aminokiseline sastoji se od dva identična dijela za sve aminokiseline, od kojih je jedna amino grupa ( -NH2) s osnovnim svojstvima, drugi s karboksilnom grupom ( -COOH) sa kiselim svojstvima. Dio molekule koji se naziva radikal ( R), različite aminokiseline imaju različite strukture. Prisutnost baznih i kiselih grupa u jednom molekulu aminokiseline određuje njihovu visoku reaktivnost. preko ovih grupa, aminokiseline su povezane da formiraju protein. U tom slučaju se pojavljuje molekul vode, a oslobođeni elektroni formiraju peptidnu vezu. Zato se proteini nazivaju polipeptidi.
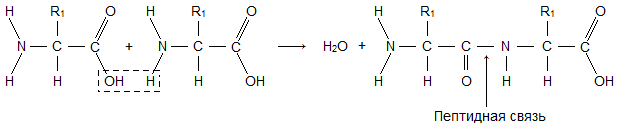
Molekuli proteina mogu imati različite prostorne konfiguracije, a u njihovoj strukturi razlikuju se četiri nivoa strukturne organizacije.
Redoslijed aminokiselina u polipeptidnom lancu je primarna struktura vjeverica. Jedinstven je za svaki protein i određuje njegov oblik, svojstva i funkcije.
Većina proteina ima spiralni oblik kao rezultat formiranja vodikovih veza između njih -CO- I -NH- grupe različitih aminokiselinskih ostataka polipeptidnog lanca. Vodikove veze su slabe, ali u kombinaciji daju prilično jaku strukturu. Ova spirala je sekundarna struktura vjeverica.
Tercijarna struktura- trodimenzionalno prostorno "pakovanje" polipeptidnog lanca. Kao rezultat toga, pojavljuje se bizarna, ali specifična konfiguracija za svaki protein - globule. Snagu tercijarne strukture osiguravaju različite veze koje nastaju između radikala aminokiselina.
Kvartarna struktura nije tipično za sve proteine. Nastaje kao rezultat kombinacije nekoliko makromolekula tercijarne strukture u složeni kompleks. Na primjer, hemoglobin ljudske krvi je kompleks od četiri proteinske makromolekule.
Ova složenost strukture proteinskih molekula povezana je s različitim funkcijama svojstvenim ovim biopolimerima.
Povreda prirodne strukture proteina se naziva denaturacija. Može nastati pod uticajem temperature, hemikalija, energije zračenja i drugih faktora. Sa slabim udarom raspada se samo kvartarna struktura, kod jačeg tercijarna, a zatim sekundarna, a protein ostaje u obliku polipeptidnog lanca.
Ovaj proces je djelomično reverzibilan: ako primarna struktura nije poremećena, tada denaturirani protein može obnoviti svoju strukturu. Iz toga slijedi da su sve karakteristike strukture proteinske makromolekule određene njegovom primarnom strukturom.
Osim jednostavnih proteina, koji se sastoje samo od aminokiselina, takođe postoje kompleksnih proteina
Vjevericesu organska jedinjenja visoke molekularne težine izgrađena od 20 aminokiselinskih ostataka. Po svojoj strukturi spadaju u polimere. Njihovi molekuli su u obliku dugih lanaca koji se sastoje od ponavljajućih molekula - monomera. Da bi se formirala molekula polimera, svaki od monomera mora imati najmanje dvije reaktivne veze s drugim monomerima.
Protein je po strukturi sličan polimernom najlonu: oba polimera su lanac monomera. Ali postoji značajna razlika između njih. Najlon se sastoji od dvije vrste monomera, dok se protein sastoji od 20 različitih monomera zvanih aminokiseline. U zavisnosti od redosleda smenjivanja monomera, formira se mnogo različitih vrsta proteina.
Opća formula za aminokiseline koje čine protein je:
Ova formula pokazuje da su četiri različite grupe vezane za centralni atom ugljika. Tri od njih - atom vodonika H, alkalna amino grupa H N i karboksilna grupa COOH - su iste za sve aminokiseline. Po sastavu i strukturi četvrta grupa ozna
R aminokiseline se razlikuju jedna od druge. U najjednostavnijim slučajevima, u molekuli glicerola - takva grupa je atom vodika, u molekuli alanina - CH, itd.Hemijska veza (- CO -
NH -), povezivanje amino grupe jedne aminokiseline sa karboksilnom grupom druge u proteinskim molekulima, naziva se peptidnu vezu(vidi sl.7.5).
Svi aktivni organizmi, bilo da su biljke, životinje, bakterije ili virusi, sadrže proteine izgrađene od istih aminokiselina. Stoga svaka vrsta hrane sadrži iste aminokiseline koje su dio proteina organizama koji hranu konzumiraju.
Definicija "proteini su polimeri izgrađeni od 20 različitih aminokiselina" sadrži nepotpunu karakterizaciju proteina. U laboratorijskim uslovima nije teško dobiti peptidne veze u rastvoru aminokiselina i tako formirati dugačke molekularne lance. Međutim, u takvim lancima raspored aminokiselina bit će haotičan, a rezultirajući molekuli će se razlikovati jedni od drugih. Istovremeno, u svakom od prirodnih proteina raspored pojedinih vrsta aminokiselina je uvijek isti. A to znači da se tokom sinteze proteina u živom sistemu koriste informacije u skladu s kojima se formira dobro definirana sekvenca aminokiselina za svaki protein.
Redoslijed aminokiselina u proteinu određuje njegovu prostornu strukturu. Većina proteina djeluje kao katalizator. U njihovoj prostornoj strukturi postoje aktivni centri u obliku udubljenja dobro definisanog oblika. U takve centre ulaze molekule, čiju transformaciju katalizira ovaj protein. Protein, koji u ovom slučaju djeluje kao enzim, može katalizirati reakciju samo ako se oblik transformirajuće molekule i aktivnog centra podudaraju. Ovo određuje visoku selektivnost proteina-enzima.
Aktivni centar enzima može nastati kao rezultat presavijanja dijelova proteinskog lanca koji su veoma udaljeni jedan od drugog. Stoga, zamjena jedne aminokiseline drugom, čak i na maloj udaljenosti od aktivnog mjesta, može ili utjecati na selektivnost enzima ili potpuno uništiti mjesto. Stvaranjem različitih sekvenci aminokiselina, možete dobiti širok izbor aktivnih centara. Ovo je jedna od najvažnijih karakteristika proteina koji djeluju kao enzimi.
Sadržaj članka
PROTEINI (član 1)- klasa bioloških polimera prisutnih u svakom živom organizmu. Uz sudjelovanje proteina odvijaju se glavni procesi koji osiguravaju vitalnu aktivnost tijela: disanje, probava, kontrakcija mišića, prijenos nervnih impulsa. Koštano tkivo, koža, kosa, formacije rogova živih bića sastoje se od proteina. Za većinu sisara, rast i razvoj organizma nastaje zahvaljujući proizvodima koji sadrže proteine kao komponentu hrane. Uloga proteina u organizmu i, shodno tome, njihova struktura je vrlo raznolika.
Sastav proteina.
Svi proteini su polimeri, čiji su lanci sastavljeni od fragmenata aminokiselina. Aminokiseline su organska jedinjenja koja u svom sastavu (u skladu sa nazivom) sadrže NH2 amino grupu i organsku kiselinu, tj. karboksil, COOH grupa. Od čitavog niza postojećih aminokiselina (teoretski, broj mogućih aminokiselina je neograničen), samo one koje imaju samo jedan atom ugljika između amino grupe i karboksilne grupe učestvuju u formiranju proteina. Generalno, aminokiseline uključene u formiranje proteina mogu se predstaviti formulom: H 2 N–CH(R)–COOH. R grupa vezana za atom ugljika (ona između amino i karboksilne grupe) određuje razliku između aminokiselina koje čine proteine. Ova grupa se može sastojati samo od atoma ugljika i vodika, ali češće sadrži, osim C i H, različite funkcionalne (sposobne za daljnje transformacije) grupe, na primjer HO-, H 2 N- itd. opcija kada je R \u003d H.
Organizmi živih bića sadrže više od 100 različitih aminokiselina, međutim, ne koriste se sve u izgradnji proteina, već samo 20, takozvanih „osnovnih“. U tabeli. 1 prikazana su njihova imena (većina imena se razvila istorijski), strukturna formula, kao i široko korištena skraćenica. Sve strukturne formule su raspoređene u tabeli tako da je glavni fragment aminokiseline na desnoj strani.
| Ime | Struktura | Oznaka |
| GLYCINE | GLI | |
| ALANIN | ALA | |
| VALIN | SHAFT | |
| LEUCINE | LEI | |
| IZOLEUCIN | ILE | |
| SERIN | SER | |
| THREONINE | TRE | |
| CYSTEINE | CIS | |
| METIONINE | MET | |
| LYSINE | LIZ | |
| ARGININ | ARG | |
| ASPARAGIC ACID | ACH | |
| ŠPARAŽIN | ACH | |
| GLUTAMIC ACID | GLU | |
| GLUTAMIN | GLN | |
| fenilalanin | fen | |
| TIROZIN | TIR | |
| triptofan | TRI | |
| HISTIDIN | GIS | |
| PROLINE | PRO | |
| U međunarodnoj praksi prihvaćeno je skraćeno označavanje navedenih aminokiselina pomoću latiničnih troslovnih ili jednoslovnih skraćenica, na primjer, glicin - Gly ili G, alanin - Ala ili A. | ||
Od ovih dvadeset aminokiselina (Tabela 1), samo prolin sadrži NH grupu (umesto NH 2) pored COOH karboksilne grupe, pošto je deo cikličkog fragmenta.
Osam aminokiselina (valin, leucin, izoleucin, treonin, metionin, lizin, fenilalanin i triptofan), koje se nalaze u tabeli na sivoj pozadini, nazivaju se esencijalnim, jer ih tijelo mora stalno primati s proteinskom hranom za normalan rast i razvoj.
Kao rezultat uzastopnog povezivanja aminokiselina nastaje proteinski molekul, dok karboksilna grupa jedne kiseline stupa u interakciju s amino grupom susjednog molekula, kao rezultat toga nastaje –CO–NH– peptidna veza i voda oslobađa se molekul. Na sl. 1 prikazuje serijsku vezu alanina, valina i glicina.
Rice. 1 SERIJSKI POVEZIVANJE AMINOKISELINA tokom formiranja proteinske molekule. Put od terminalne amino grupe H 2 N do terminalne karboksilne grupe COOH odabran je kao glavni pravac polimernog lanca.
Za kompaktan opis strukture proteinske molekule koriste se skraćenice za aminokiseline (tablica 1, treći stupac) uključene u formiranje polimernog lanca. Fragment molekula prikazan na sl. 1 se piše na sljedeći način: H 2 N-ALA-VAL-GLY-COOH.
Molekuli proteina sadrže od 50 do 1500 aminokiselinskih ostataka (kraći lanci se nazivaju polipeptidi). Individualnost proteina određena je skupom aminokiselina koje čine polimerni lanac i, ne manje važno, redoslijedom njihove izmjene duž lanca. Na primjer, molekula inzulina sastoji se od 51 aminokiselinskog ostatka (to je jedan od najkraćih lanaca proteina) i sastoji se od dva međusobno povezana paralelna lanca nejednake dužine. Slijed fragmenata aminokiselina prikazan je na sl. 2.
Rice. 2 INSULIN MOLECULE, izgrađen od 51 aminokiselinskog ostatka, fragmenti istih aminokiselina označeni su odgovarajućom bojom pozadine. Cisteinski aminokiselinski ostaci (skraćena oznaka CIS) sadržani u lancu formiraju disulfidne mostove -S-S-, koji povezuju dvije polimerne molekule, ili formiraju skakače unutar jednog lanca.
Molekule aminokiseline cisteina (Tablica 1) sadrže reaktivne sulfhidridne grupe -SH, koje međusobno djeluju, formirajući disulfidne mostove -S-S-. Uloga cisteina u svijetu proteina je posebna, uz njegovo učešće nastaju poprečne veze između polimernih proteinskih molekula.
Povezivanje aminokiselina u polimerni lanac događa se u živom organizmu pod kontrolom nukleinskih kiselina, upravo one osiguravaju strogi redoslijed sklapanja i reguliraju fiksnu dužinu polimerne molekule ( cm. NUKLEINSKE KISELINE).
Struktura proteina.
Sastav proteinske molekule, predstavljen u obliku naizmjeničnih aminokiselinskih ostataka (slika 2), naziva se primarna struktura proteina. Vodikove veze nastaju između imino grupa HN prisutnih u polimernom lancu i karbonilnih grupa CO ( cm. VODIKOVA VEZA), kao rezultat toga, proteinski molekul poprima određeni prostorni oblik, koji se naziva sekundarna struktura. Najčešća su dva tipa sekundarne strukture u proteinima.
Prva opcija, nazvana α-heliks, implementirana je korištenjem vodoničnih veza unutar jedne polimerne molekule. Geometrijski parametri molekule, određeni dužinama veza i uglovima veze, takvi su da je moguće formiranje vodoničnih veza za H-N i C=O grupe, između kojih se nalaze dva peptidna fragmenta H-N-C=O (slika 3) .
Sastav polipeptidnog lanca prikazan na sl. 3 se piše u skraćenom obliku kako slijedi:
H 2 N-ALA VAL-ALA-LEY-ALA-ALA-ALA-ALA-VAL-ALA-ALA-ALA-COOH.
Kao rezultat kontrakcije vodikovih veza, molekula poprima oblik spirale - tzv. α-helix, prikazana je kao zakrivljena spiralna vrpca koja prolazi kroz atome koji formiraju polimerni lanac (slika 4)
Rice. 4 3D MODEL PROTEINSKOG MOLEKULA u obliku α-heliksa. Vodikove veze su prikazane kao zelene isprekidane linije. Cilindrični oblik spirale je vidljiv pod određenim kutom rotacije (atomi vodika nisu prikazani na slici). Boja pojedinačnih atoma data je u skladu s međunarodnim pravilima, koja preporučuju crnu za atome ugljika, plavu za dušik, crvenu za kisik i žutu za sumpor (bijela boja se preporučuje za atome vodika koji nisu prikazani na slici, u ovom slučaju cijela struktura prikazana na tamnoj pozadini).
Druga varijanta sekundarne strukture, nazvana β-struktura, takođe se formira uz učešće vodoničnih veza, razlika je u tome što H-N i C=O grupe dva ili više polimernih lanaca lociranih paralelno. Budući da polipeptidni lanac ima smjer (slika 1), moguće su varijante kada je smjer lanaca isti (paralelna β-struktura, slika 5), ili su suprotni (antiparalelna β-struktura, slika 6) .
Polimerni lanci različitog sastava mogu učestvovati u formiranju β-strukture, dok organske grupe koje uokviruju polimerni lanac (Ph, CH 2 OH i dr.) u većini slučajeva imaju sporednu ulogu, međusobni raspored H-N i C =O grupa je odlučujuća. Budući da su H-N i C=O grupe usmjerene u različitim smjerovima u odnosu na polimerni lanac (gore i dolje na slici), postaje moguće da tri ili više lanaca međusobno djeluju istovremeno.
Sastav prvog polipeptidnog lanca na Sl. 5:
H 2 N-LEI-ALA-FEN-GLI-ALA-ALA-COOH
Sastav drugog i trećeg lanca:
H 2 N-GLY-ALA-SER-GLY-TRE-ALA-COOH
Sastav polipeptidnih lanaca prikazanih na sl. 6, isto kao na sl. 5, razlika je u tome što drugi lanac ima suprotan (u poređenju sa slikom 5) smjer.
Moguće je formirati β-strukturu unutar jedne molekule, kada se ispostavi da je fragment lanca u određenom dijelu rotiran za 180°, u ovom slučaju dvije grane jedne molekule imaju suprotan smjer, kao rezultat, antiparalela Formira se β-struktura (slika 7).
Struktura prikazana na sl. 7 na ravnoj slici, prikazanoj na sl. 8 u obliku trodimenzionalnog modela. Dijelovi β-strukture obično se pojednostavljeno označavaju ravnom valovitom vrpcom koja prolazi kroz atome koji formiraju polimerni lanac.
U strukturi mnogih proteina izmjenjuju se dijelovi α-heliksa i β-strukture nalik vrpci, kao i pojedinačni polipeptidni lanci. Njihov međusobni raspored i izmjena u polimernom lancu naziva se tercijarna struktura proteina.
Metode za opisivanje strukture proteina prikazane su u nastavku koristeći biljni protein krambin kao primjer. Strukturne formule proteina, koje često sadrže i do stotine fragmenata aminokiselina, složene su, glomazne i teško razumljive, pa se ponekad koriste pojednostavljene strukturne formule - bez simbola hemijskih elemenata (slika 9, opcija A), ali istovremeno vremena zadržavaju boju valentnih poteza u skladu sa međunarodnim pravilima (slika 4). U ovom slučaju, formula nije prikazana u ravnoj, već u prostornoj slici, koja odgovara stvarnoj strukturi molekula. Ova metoda omogućava, na primjer, razlikovanje disulfidnih mostova (slično onima koji se nalaze u inzulinu, slika 2), fenilnih grupa u bočnom okviru lanca, itd. Slika molekula u obliku trodimenzionalnog modela (loptice povezane šipkama) je nešto jasnije (slika 9, opcija B). Međutim, obje metode ne dozvoljavaju prikazivanje tercijarne strukture, pa je američka biofizičarka Jane Richardson predložila da se α-strukture prikažu kao spiralno uvijene vrpce (vidi sliku 4), β-strukture kao ravne valovite vrpce (slika 8) i spojne to su pojedinačni lanci - u obliku tankih snopova, svaka vrsta strukture ima svoju boju. Ova metoda prikazivanja tercijarne strukture proteina se danas široko koristi (slika 9, varijanta B). Ponekad su, radi većeg informativnog sadržaja, tercijarna struktura i pojednostavljena strukturna formula prikazane zajedno (slika 9, varijanta D). Postoje i modifikacije metode koju je predložio Richardson: α-heliksi su prikazani kao cilindri, a β-strukture su u obliku ravnih strelica koje pokazuju smjer lanca (Sl. 9, opcija E). Manje uobičajena je metoda u kojoj se cijeli molekul prikazuje kao snop, gdje se nejednake strukture razlikuju različitim bojama, a disulfidni mostovi prikazani kao žuti mostovi (Sl. 9, varijanta E).
Opcija B je najpogodnija za percepciju, kada se pri oslikavanju tercijarne strukture ne navode strukturne karakteristike proteina (fragmenti aminokiselina, njihov redoslijed izmjene, vodikove veze), dok se pretpostavlja da svi proteini sadrže "detalje" uzeto iz standardnog skupa od dvadeset aminokiselina (Tabela 1). Glavni zadatak u prikazu tercijarne strukture je prikazati prostorni raspored i izmjenu sekundarnih struktura.
Rice. 9 RAZLIČITE VERZIJE SLIKE STRUKTURE CRUMBIN PROTEINA.
A je strukturna formula u prostornoj slici.
B - struktura u obliku trodimenzionalnog modela.
B je tercijarna struktura molekula.
G - kombinacija opcija A i B.
E - pojednostavljena slika tercijarne strukture.
E - tercijarna struktura sa disulfidnim mostovima.
Najprikladnija za percepciju je trodimenzionalna tercijarna struktura (opcija B), oslobođena detalja strukturne formule.
Molekul proteina koji ima tercijarnu strukturu u pravilu poprima određenu konfiguraciju, koja nastaje polarnim (elektrostatičkim) interakcijama i vodikovim vezama. Kao rezultat toga, molekula poprima oblik kompaktne zavojnice - globularnih proteina (globule, lat. loptasta), ili filamentozno - fibrilarni proteini (fibra, lat. vlakna).
Primjer globularne strukture je protein albumin, protein kokošjeg jajeta pripada klasi albumina. Polimerni lanac albumina sastavljen je uglavnom od alanina, asparaginske kiseline, glicina i cisteina, naizmjenično određenim redoslijedom. Tercijarna struktura sadrži α-helike povezane pojedinačnim lancima (slika 10).
Rice. 10 GLOBULARNA STRUKTURA ALBUMINA
Primjer fibrilarne strukture je protein fibroina. Sadrže veliku količinu ostataka glicina, alanina i serina (svaki drugi aminokiselinski ostatak je glicin); cisteinski ostaci koji sadrže sulfhidridne grupe su odsutni. Fibroin, glavna komponenta prirodne svile i paučine, sadrži β-strukture povezane pojedinačnim lancima (slika 11).
Rice. jedanaest FIBRILARNI PROTEIN FIBROIN
Mogućnost formiranja tercijarne strukture određenog tipa inherentna je primarnoj strukturi proteina, tj. unaprijed određen redoslijedom alternacije aminokiselinskih ostataka. Iz određenih skupova takvih ostataka pretežno nastaju α-heliksi (takvih skupova ima dosta), drugi skup dovodi do pojave β-struktura, pojedinačni lanci se odlikuju svojim sastavom.
Neki proteinski molekuli, zadržavajući tercijarnu strukturu, u stanju su da se kombinuju u velike supramolekularne agregate, dok se drže zajedno polarnim interakcijama, kao i vodoničnim vezama. Takve formacije se nazivaju kvarterna struktura proteina. Na primjer, protein feritin, koji se uglavnom sastoji od leucina, glutaminske kiseline, asparaginske kiseline i histidina (fericin sadrži svih 20 aminokiselinskih ostataka u različitim količinama) formira tercijarnu strukturu od četiri paralelno položene α-heliksa. Kada se molekuli spoje u jedan ansambl (slika 12), formira se kvartarna struktura koja može uključivati do 24 molekula feritina.
Fig.12 FORMIRANJE KVARTERNE STRUKTURE GLOBULARNOG PROTEINA FERITINA
Drugi primjer supramolekularnih formacija je struktura kolagena. To je fibrilarni protein čiji su lanci izgrađeni uglavnom od glicina naizmjenično s prolinom i lizinom. Struktura sadrži pojedinačne lance, trostruke α-heliksa, koji se izmjenjuju sa β-strukturama nalik vrpci složenim u paralelne snopove (slika 13).
Fig.13 SUPRAMOLEKULARNA STRUKTURA KOLAGENOG FIBRILARNOG PROTEINA
Hemijska svojstva proteina.
Pod djelovanjem organskih otapala, otpadnih proizvoda nekih bakterija (fermentacija mliječne kiseline) ili povećanjem temperature, sekundarne i tercijarne strukture se uništavaju bez oštećenja njegove primarne strukture, zbog čega protein gubi topljivost i gubi biološku aktivnost. proces se naziva denaturacija, odnosno gubitak prirodnih svojstava, na primjer, zgrušavanje kiselog mlijeka, zgrušanog proteina kuhanog kokošjeg jajeta. Na povišenim temperaturama, proteini živih organizama (posebno mikroorganizama) brzo denaturiraju. Takvi proteini ne mogu sudjelovati u biološkim procesima, zbog čega mikroorganizmi umiru, pa se kuhano (ili pasterizirano) mlijeko može duže čuvati.
Peptidne veze H-N-C=O, formirajući polimerni lanac proteinske molekule, hidroliziraju se u prisustvu kiselina ili alkalija, a polimerni lanac se raskida, što, u konačnici, može dovesti do originalnih aminokiselina. Peptidne veze uključene u α-heliksa ili β-strukture su otpornije na hidrolizu i različite hemijske napade (u poređenju sa istim vezama u pojedinačnim lancima). Delikatnije rastavljanje proteinske molekule na sastavne aminokiseline vrši se u bezvodnom mediju koristeći hidrazin H 2 N–NH 2, dok svi fragmenti aminokiselina, osim posljednjeg, tvore tzv. hidrazide karboksilne kiseline koji sadrže fragment C (O)–HN–NH 2 (Sl. 14).
Rice. 14. CIJEPANJE POLIPEPTIDA
Takva analiza može pružiti informacije o sastavu aminokiselina proteina, ali je važnije znati njihov slijed u proteinskom molekulu. Jedna od metoda koja se široko koristi u tu svrhu je djelovanje fenilizotiocijanata (FITC) na polipeptidni lanac, koji se u alkalnom mediju veže za polipeptid (sa kraja koji sadrži amino grupu), a kada se reakcija medija promijeni u kiseli, odvaja se od lanca, uzimajući sa sobom fragment jedne aminokiseline (slika 15).
Rice. 15 SEKVENCIJALNO cijepanje polipeptida
Za takvu analizu razvijene su mnoge posebne metode, uključujući one koje počinju da „rastavljaju“ proteinski molekul na njegove sastavne komponente, počevši od karboksilnog kraja.
Unakrsni disulfidni mostovi S-S (nastali interakcijom cisteinskih ostataka, sl. 2 i 9) se cijepaju, pretvarajući ih u HS-grupe djelovanjem različitih redukcijskih sredstava. Djelovanje oksidirajućih sredstava (kiseonik ili vodikov peroksid) opet dovodi do stvaranja disulfidnih mostova (Sl. 16).
Rice. 16. Cepanje disulfidnih mostova
Za stvaranje dodatnih poprečnih veza u proteinima koristi se reaktivnost amino i karboksilnih grupa. Za različite interakcije pristupačnije su amino grupe koje se nalaze u bočnom okviru lanca – fragmenti lizina, asparagina, lizina, prolina (tabela 1). Kada takve amino grupe stupe u interakciju sa formaldehidom, dolazi do procesa kondenzacije i pojavljuju se poprečni mostovi –NH–CH2–NH– (slika 17).
Rice. 17 STVARANJE DODATNIH TRANSVERZALNIH MOSTOVA IZMEĐU PROTEINSKIH MOLEKULA.
Terminalne karboksilne grupe proteina mogu da reaguju sa kompleksnim jedinjenjima nekih polivalentnih metala (češće se koriste jedinjenja hroma), a takođe se javljaju i poprečne veze. Oba procesa se koriste u štavljenju kože.
Uloga proteina u organizmu.
Uloga proteina u organizmu je raznolika.
Enzimi(fermentacija lat. - fermentacija), njihov drugi naziv je enzimi (en zumh greek. - u kvascu) - to su proteini sa katalitičkom aktivnošću, sposobni su da povećaju brzinu biohemijskih procesa hiljadama puta. Pod djelovanjem enzima sastavni dijelovi hrane: bjelančevine, masti i ugljikohidrati razgrađuju se na jednostavnija jedinjenja iz kojih se potom sintetiziraju novi makromolekuli neophodni za određeni tip tijela. Enzimi također učestvuju u mnogim biohemijskim procesima sinteze, na primjer, u sintezi proteina (neki proteini pomažu u sintezi drugih). Cm. ENZIMI
Enzimi nisu samo visoko efikasni katalizatori, već su i selektivni (usmjeravaju reakciju striktno u datom smjeru). U njihovom prisustvu, reakcija se odvija sa skoro 100% prinosom bez stvaranja nusproizvoda, a istovremeno su uslovi protoka blagi: normalan atmosferski pritisak i temperatura živog organizma. Poređenja radi, sinteza amonijaka iz vodonika i dušika u prisustvu aktiviranog željeznog katalizatora vrši se na 400-500°C i pritisku od 30 MPa, prinos amonijaka je 15-25% po ciklusu. Enzimi se smatraju nenadmašnim katalizatorima.
Intenzivno proučavanje enzima počelo je sredinom 19. vijeka, do sada je proučavano više od 2.000 različitih enzima; ovo je najraznovrsnija klasa proteina.
Nazivi enzima su sljedeći: naziv reagensa s kojim enzim stupa u interakciju, ili naziv katalizirane reakcije, dodaje se završetkom -aza, na primjer, arginaza razgrađuje arginin (tabela 1), dekarboksilaza katalizira dekarboksilaciju, tj. eliminacija CO 2 iz karboksilne grupe:
– COOH → – CH + CO 2
Često, da bi se preciznije ukazala na ulogu enzima, i predmet i vrsta reakcije su naznačeni u njegovom nazivu, na primjer, alkohol dehidrogenaza je enzim koji dehidrogenira alkohole.
Za neke davno otkrivene enzime sačuvano je istorijsko ime (bez završetka -aza), na primjer pepsin (pepsis, grčki. varenje) i tripsin (thrypsis grčki. tečenje), ovi enzimi razgrađuju proteine.
Za sistematizaciju, enzimi se kombinuju u velike klase, klasifikacija se zasniva na tipu reakcije, klase se imenuju prema opštem principu - naziv reakcije i završetak - aza. Neke od ovih klasa su navedene u nastavku.
Oksidoreduktaza su enzimi koji katalizuju redoks reakcije. Dehidrogenaze uključene u ovu klasu vrše prijenos protona, na primjer, alkohol dehidrogenaza (ADH) oksidira alkohole u aldehide, naknadnu oksidaciju aldehida u karboksilne kiseline kataliziraju aldehid dehidrogenaze (ALDH). Oba procesa se dešavaju u organizmu tokom prerade etanola u sirćetnu kiselinu (slika 18).
Rice. 18 DVOSTEPENA OKSIDACIJA ETANOLA na sirćetnu kiselinu
Nije etanol taj koji ima narkotično djelovanje, već intermedijarni produkt acetaldehid, što je aktivnost enzima ALDH niža, to sporije prolazi druga faza - oksidacija acetaldehida u octenu kiselinu, a opojno djelovanje od uzimanja duže i jače etanola. Analiza je pokazala da više od 80% predstavnika žute rase ima relativno nisku aktivnost ALDH, a samim tim i izrazito veću toleranciju na alkohol. Razlog za ovu urođenu smanjenu aktivnost ALDH je taj što je dio ostataka glutaminske kiseline u „oslabljenoj“ molekuli ALDH zamijenjen fragmentima lizina (Tabela 1).
Transferaze- enzimi koji kataliziraju prijenos funkcionalnih grupa, na primjer, transiminaza katalizuje prijenos amino grupe.
Hidrolaze su enzimi koji katalizuju hidrolizu. Prethodno spomenuti tripsin i pepsin hidroliziraju peptidne veze, a lipaze cijepaju estersku vezu u mastima:
–RC(O)OR 1 + H 2 O → –RC(O)OH + HOR 1
Liase- enzimi koji katalizuju reakcije koje se odvijaju na nehidrolitički način, kao rezultat takvih reakcija se prekidaju C-C, C-O, C-N veze i stvaraju nove veze. Enzim dekarboksilaza pripada ovoj klasi
Izomeraze- enzimi koji kataliziraju izomerizaciju, na primjer, konverziju maleinske kiseline u fumarnu kiselinu (slika 19), ovo je primjer cis-trans izomerizacije (vidi IZOMERIJA).
Rice. 19. IZOMERIZACIJA MALEINSKE KISELINE u fumarnu kiselinu u prisustvu enzima.
U radu enzima uvažava se opći princip prema kojem uvijek postoji strukturna korespondencija između enzima i reagensa ubrzane reakcije. Prema figurativnom izrazu jednog od osnivača doktrine enzima, E. Fishera, reagens se približava enzimu poput ključa od brave. U tom smislu, svaki enzim katalizira određenu kemijsku reakciju ili grupu reakcija istog tipa. Ponekad enzim može djelovati na jedno jedinjenje, kao što je ureaza (uron grčki. - urin) katalizira samo hidrolizu ureje:
(H 2 N) 2 C \u003d O + H 2 O \u003d CO 2 + 2NH 3
Najfiniju selektivnost pokazuju enzimi koji razlikuju optički aktivne antipode - lijevo i desnoruke izomere. L-arginaza djeluje samo na levorotirajući arginin i ne utiče na desnorotirajući izomer. L-laktat dehidrogenaza djeluje samo na levorotirajuće estere mliječne kiseline, tzv. laktate (lactis lat. mlijeko), dok D-laktat dehidrogenaza razgrađuje samo D-laktate.
Većina enzima ne djeluje na jedan, već na grupu srodnih spojeva, na primjer, tripsin "radije" cijepa peptidne veze koje formiraju lizin i arginin (tablica 1.)
Katalitička svojstva nekih enzima, kao što su hidrolaze, određena su isključivo strukturom same proteinske molekule, druga klasa enzima - oksidoreduktaze (na primjer, alkohol dehidrogenaza) mogu biti aktivna samo u prisustvu neproteinskih molekula povezanih sa njih - vitamini koji aktiviraju Mg, Ca, Zn, Mn i fragmenti nukleinskih kiselina (slika 20).
Rice. 20 MOLEKULA ALKOHODA DEHIDROGENAZE
Transportni proteini vezuju i transportuju različite molekule ili ione kroz ćelijske membrane (unutar i izvan ćelije), kao i od jednog organa do drugog.
Na primjer, hemoglobin veže kisik dok krv prolazi kroz pluća i isporučuje ga u različita tkiva u tijelu, gdje se kisik oslobađa i zatim koristi za oksidaciju komponenti hrane, ovaj proces služi kao izvor energije (ponekad koriste izraz "sagorijevanje" hranu u organizmu).
Pored proteinskog dela, hemoglobin sadrži kompleksno jedinjenje gvožđa sa cikličkom molekulom porfirina (porfiros grčki. - ljubičasta), koja određuje crvenu boju krvi. Upravo ovaj kompleks (slika 21, lijevo) igra ulogu nosača kiseonika. U hemoglobinu, kompleks gvožđa porfirin nalazi se unutar proteinske molekule i zadržava se polarnim interakcijama, kao i koordinacionom vezom sa azotom u histidinu (tabela 1), koji je deo proteina. Molekul O2, koji se prenosi hemoglobinom, vezan je koordinacionom vezom za atom gvožđa sa strane suprotne onoj za koju je vezan histidin (slika 21, desno).
Rice. 21 STRUKTURA KOMPLEKSA GVOŽĐA
Struktura kompleksa prikazana je desno u obliku trodimenzionalnog modela. Kompleks se drži u proteinskoj molekuli koordinacionom vezom (isprekidana plava linija) između atoma Fe i atoma N u histidinu, koji je dio proteina. Molekul O 2, koji se prenosi hemoglobinom, koordiniran je (crvena tačkasta linija) na atom Fe iz suprotne zemlje planarnog kompleksa.
Hemoglobin je jedan od najviše proučavanih proteina, sastoji se od a-heliksa povezanih pojedinačnim lancima i sadrži četiri kompleksa gvožđa. Dakle, hemoglobin je poput voluminoznog paketa za prijenos četiri molekula kisika odjednom. Oblik hemoglobina odgovara globularnim proteinima (slika 22).
Rice. 22 GLOBULARNI OBLIK HEMOGLOBINA
Glavna "prednost" hemoglobina je što se dodavanje kiseonika i njegovo kasnije odvajanje tokom prenosa u različita tkiva i organe odvija brzo. Ugljen-monoksid, CO (ugljen-monoksid), još brže se vezuje za Fe u hemoglobinu, ali, za razliku od O 2 , formira kompleks koji se teško razgrađuje. Kao rezultat, takav hemoglobin nije u stanju da veže O 2, što dovodi (kada se udiše velike količine ugljičnog monoksida) do smrti tijela od gušenja.
Druga funkcija hemoglobina je prijenos izdahnutog CO 2, ali ne atoma željeza, već je H 2 N-grupe proteina uključen u proces privremenog vezivanja ugljičnog dioksida.
"Učinak" proteina ovisi o njihovoj strukturi, na primjer, zamjena jedinog aminokiselinskog ostatka glutaminske kiseline u polipeptidnom lancu hemoglobina ostatkom valina (rijetko uočena kongenitalna anomalija) dovodi do bolesti zvane anemija srpastih stanica.
Postoje i transportni proteini koji mogu vezati masti, glukozu, aminokiseline i prenositi ih unutar i izvan stanica.
Transportni proteini posebne vrste ne nose same supstance, već deluju kao „regulator transporta“, propuštajući određene supstance kroz membranu (vanjski zid ćelije). Takvi proteini se često nazivaju membranskim proteinima. Imaju oblik šupljeg cilindra i, budući da su ugrađeni u zid membrane, osiguravaju kretanje nekih polarnih molekula ili jona u ćeliju. Primjer membranskog proteina je porin (slika 23).
Rice. 23 PORIN PROTEIN
Proteini za hranu i skladištenje, kao što samo ime govori, služe kao izvori unutrašnje ishrane, češće za embrione biljaka i životinja, kao i u ranim fazama razvoja mladih organizama. Proteini u ishrani uključuju albumin (slika 10) - glavnu komponentu bjelanjka, kao i kazein - glavni protein mlijeka. Pod dejstvom enzima pepsina, kazein se zgrušava u želucu, što obezbeđuje njegovo zadržavanje u digestivnom traktu i efikasnu apsorpciju. Kazein sadrži fragmente svih aminokiselina potrebnih tijelu.
U feritinu (slika 12), koji se nalazi u tkivima životinja, pohranjeni su ioni željeza.
Mioglobin je takođe protein za skladištenje, koji po sastavu i strukturi podseća na hemoglobin. Mioglobin je koncentrisan uglavnom u mišićima, njegova glavna uloga je skladištenje kisika koji mu daje hemoglobin. Brzo se zasićuje kiseonikom (mnogo brže od hemoglobina), a zatim ga postepeno prenosi u različita tkiva.
Strukturni proteini obavljaju zaštitnu funkciju (koža) ili potporu – drže tijelo zajedno i daju mu snagu (hrskavica i tetive). Njihova glavna komponenta je fibrilarni protein kolagen (slika 11), najčešći protein životinjskog svijeta, u tijelu sisara čini skoro 30% ukupne mase proteina. Kolagen ima veliku vlačnu čvrstoću (poznata je čvrstoća kože), ali zbog niskog sadržaja poprečnih veza u kolagenu kože životinjske kože nisu baš pogodne u sirovom obliku za proizvodnju raznih proizvoda. Da bi se smanjilo oticanje kože u vodi, skupljanje tokom sušenja, kao i da bi se povećala čvrstoća u natopljenom stanju i povećala elastičnost u kolagenu, stvaraju se dodatne poprečne veze (slika 15a), radi se o tzv. proces štavljenja kože.
U živim organizmima molekule kolagena koje su nastale u procesu rasta i razvoja organizma ne ažuriraju se i ne zamjenjuju se novosintetiziranim. Starenjem tijela povećava se broj poprečnih veza u kolagenu, što dovodi do smanjenja njegove elastičnosti, a kako do obnove ne dolazi, pojavljuju se promjene vezane za starenje - povećanje krhkosti hrskavice i tetiva, pojava bora na koži.
Zglobni ligamenti sadrže elastin, strukturni protein koji se lako rasteže u dvije dimenzije. Najveću elastičnost ima protein resilin, koji se nalazi na mjestima spajanja krila na šarkama kod nekih insekata.
Formacije rogova - kosa, nokti, perje, sastoje se uglavnom od proteina keratina (slika 24). Njegova glavna razlika je primjetan sadržaj cisteinskih ostataka, koji formiraju disulfidne mostove, što kosi, kao i vunenim tkaninama daje visoku elastičnost (sposobnost vraćanja prvobitnog oblika nakon deformacije).
Rice. 24. FRAGMENT FIBRILARNOG PROTEINA KERATINA
Za nepovratnu promjenu oblika keratinskog predmeta, prvo morate uništiti disulfidne mostove uz pomoć redukcionog sredstva, dati mu novi oblik, a zatim ponovo stvoriti disulfidne mostove uz pomoć oksidacijskog sredstva (sl. . 16), ovako se radi, na primjer, trajna kosa.
S povećanjem sadržaja cisteinskih ostataka u keratinu i, shodno tome, povećanjem broja disulfidnih mostova, sposobnost deformacije nestaje, ali se istovremeno pojavljuje visoka čvrstoća (rogovi kopitara i oklopa kornjače sadrže do 18% cisteinskih fragmenata). Sisavci imaju do 30 različitih vrsta keratina.
Fibrilarni protein vezan za keratin, fibroin koji luče gusjenice svilene bube tokom uvijanja čahure, kao i pauci tokom tkanja mreže, sadrži samo β-strukture povezane jednostrukim lancima (Sl. 11). Za razliku od keratina, fibroin nema poprečne disulfidne mostove, ima vrlo jaku vlačnu čvrstoću (čvrstoća po jedinici poprečnog presjeka nekih uzoraka mreže veća je od čeličnih kablova). Zbog odsustva poprečnih veza, fibroin je neelastičan (poznato je da su vunene tkanine gotovo neizbrisive, a svilene tkanine se lako gužvaju).
regulatorni proteini.
Regulatorni proteini, koji se češće nazivaju hormonima, uključeni su u različite fiziološke procese. Na primjer, hormon insulin (slika 25) sastoji se od dva α-lanca povezana disulfidnim mostovima. Inzulin reguliše metaboličke procese koji uključuju glukozu, njegov nedostatak dovodi do dijabetesa.
Rice. 25 PROTEIN INSULIN
Hipofiza mozga sintetizira hormon koji regulira rast tijela. Postoje regulatorni proteini koji kontroliraju biosintezu različitih enzima u tijelu.
Kontraktilni i motorni proteini daju tijelu sposobnost kontrakcije, promjene oblika i kretanja, prije svega, riječ je o mišićima. 40% mase svih proteina sadržanih u mišićima je miozin (mys, myos, grčki. - mišić). Njegova molekula sadrži i fibrilarni i globularni dio (slika 26)
Rice. 26 MYOSIN MOLECULE
Takvi molekuli se spajaju u velike agregate koji sadrže 300-400 molekula.
Kada se koncentracija kalcijevih jona promijeni u prostoru koji okružuje mišićna vlakna, dolazi do reverzibilne promjene u konformaciji molekula - promjene oblika lanca zbog rotacije pojedinačnih fragmenata oko valentnih veza. To dovodi do kontrakcije i opuštanja mišića, signal za promjenu koncentracije kalcijevih jona dolazi iz nervnih završetaka u mišićnim vlaknima. Umjetna kontrakcija mišića može biti uzrokovana djelovanjem električnih impulsa, što dovodi do nagle promjene koncentracije kalcijevih jona, što je osnova za stimulaciju srčanog mišića da obnovi rad srca.
Zaštitni proteini vam omogućavaju da zaštitite tijelo od invazije napadačkih bakterija, virusa i od prodora stranih proteina (općeniti naziv stranih tijela je antigeni). Ulogu zaštitnih proteina obavljaju imunoglobulini (njihovo drugo ime je antitijela), oni prepoznaju antigene koji su prodrli u tijelo i čvrsto se vežu za njih. U tijelu sisara, uključujući i čovjeka, postoji pet klasa imunoglobulina: M, G, A, D i E, njihova struktura je, kao što naziv govori, globularna, osim toga, svi su građeni na sličan način. Molekularna organizacija antitela je prikazana u nastavku na primeru imunoglobulina klase G (slika 27). Molekul sadrži četiri polipeptidna lanca povezana sa tri S-S disulfidna mosta (na slici 27 oni su prikazani sa zadebljanim valentnim vezama i velikim S simbolima), osim toga, svaki polimerni lanac sadrži intralančane disulfidne mostove. Dva velika polimerna lanca (označena plavom bojom) sadrže 400-600 aminokiselinskih ostataka. Druga dva lanca (označena zelenom bojom) su skoro upola kraća i sadrže otprilike 220 aminokiselinskih ostataka. Sva četiri lanca su locirana na način da su terminalne H 2 N-grupe usmjerene u jednom smjeru.
Rice. 27 ŠEMATSKI CRTEŽ STRUKTURE IMUNOGLOBULINA
Nakon što tijelo dođe u kontakt sa stranim proteinom (antigenom), ćelije imunog sistema počinju proizvoditi imunoglobuline (antitijela), koji se akumuliraju u krvnom serumu. U prvoj fazi, glavni posao obavljaju sekcije lanca koje sadrže terminal H 2 N (na slici 27 odgovarajuće sekcije su označene svijetloplavom i svijetlozelenom bojom). Ovo su mjesta za hvatanje antigena. U procesu sinteze imunoglobulina, ova mjesta se formiraju na način da njihova struktura i konfiguracija što više odgovaraju strukturi antigena koji se približava (poput ključa brave, poput enzima, ali zadaci su u ovom slučaju drugačije). Tako se za svaki antigen stvara striktno individualno antitijelo kao imuni odgovor. Niti jedan poznati protein ne može tako "plastično" promijeniti svoju strukturu ovisno o vanjskim faktorima, pored imunoglobulina. Enzimi rješavaju problem strukturne usklađenosti sa reagensom na drugačiji način - uz pomoć gigantskog skupa raznih enzima za sve moguće slučajeve, a imunoglobulini svaki put obnavljaju "radni alat". Štaviše, zglobni region imunoglobulina (slika 27) obezbedjuje dva regiona hvatanja neku nezavisnu pokretljivost, kao rezultat toga, molekul imunoglobulina može odmah „pronaći“ dva najpogodnija regiona za hvatanje u antigenu kako bi se sigurno fiksirao to, ovo liči na postupke rakova.
Zatim se uključuje lanac uzastopnih reakcija imunološkog sustava tijela, povezuju imunoglobulini drugih klasa, kao rezultat toga, strani protein se deaktivira, a zatim se antigen (strani mikroorganizam ili toksin) uništava i uklanja.
Nakon kontakta sa antigenom, maksimalna koncentracija imunoglobulina se postiže (u zavisnosti od prirode antigena i individualnih karakteristika samog organizma) u roku od nekoliko sati (ponekad i nekoliko dana). Tijelo zadržava sjećanje na takav kontakt, a pri ponovnom napadu istim antigenom, imunoglobulini se mnogo brže akumuliraju u krvnom serumu iu većim količinama - javlja se stečeni imunitet.
Navedena klasifikacija proteina je u određenoj mjeri uslovna, na primjer, trombin protein, koji se spominje među zaštitnim proteinima, u suštini je enzim koji katalizira hidrolizu peptidnih veza, odnosno pripada klasi proteaza.
Zaštitni proteini se često nazivaju proteinima zmijskog otrova i toksičnim proteinima nekih biljaka, jer je njihov zadatak da štite tijelo od oštećenja.
Postoje proteini čije su funkcije toliko jedinstvene da ih je teško klasificirati. Na primjer, protein monelin, koji se nalazi u afričkoj biljci, vrlo je slatkog okusa i bio je predmet proučavanja kao netoksična supstanca koja se može koristiti umjesto šećera za sprječavanje gojaznosti. Krvna plazma nekih antarktičkih riba sadrži proteine sa svojstvima antifriza koji sprečavaju smrzavanje krvi ovih riba.
Umjetna sinteza proteina.
Kondenzacija aminokiselina koja vodi do polipeptidnog lanca je dobro proučen proces. Moguće je izvesti, na primjer, kondenzaciju bilo koje jedne aminokiseline ili mješavine kiselina i dobiti, respektivno, polimer koji sadrži iste jedinice, ili različite jedinice, koje se izmjenjuju nasumičnim redoslijedom. Takvi polimeri malo liče na prirodne polipeptide i nemaju biološku aktivnost. Glavni zadatak je povezati aminokiseline u strogo definiranom, unaprijed planiranom redoslijedu kako bi se reproducirao slijed aminokiselinskih ostataka u prirodnim proteinima. Američki naučnik Robert Merrifield predložio je originalnu metodu koja je omogućila rješavanje takvog problema. Suština metode je da je prva aminokiselina vezana za nerastvorljivi polimerni gel koji sadrži reaktivne grupe koje se mogu kombinovati sa –COOH – grupama aminokiseline. Kao takav polimerni supstrat uzet je umreženi polistiren sa unesenim hlorometil grupama. Da aminokiselina uzeta za reakciju ne reaguje sama sa sobom i kako ne bi spojila H 2 N-grupu sa supstratom, amino grupa ove kiseline je prethodno blokirana glomaznim supstituentom [(C 4 H 9) 3] 3 OS (O) -grupa. Nakon što se aminokiselina veže na polimerni nosač, blokirajuća grupa se uklanja i druga aminokiselina se uvodi u reakcionu smjesu, u kojoj je H2N grupa također prethodno blokirana. U takvom sistemu moguća je samo interakcija H 2 N-grupe prve aminokiseline i –COOH grupe druge kiseline, koja se odvija u prisustvu katalizatora (fosfonijumovih soli). Zatim se cijela šema ponavlja, uvodeći treću aminokiselinu (slika 28).
Rice. 28. ŠEMA SINTEZI POLIPEPTIDNIH LANCA
U posljednjem koraku, nastali polipeptidni lanci se odvajaju od polistirenske podloge. Sada je cijeli proces automatiziran, postoje automatski sintetizatori peptida koji rade prema opisanoj shemi. Mnogi peptidi koji se koriste u medicini i poljoprivredi sintetizirani su ovom metodom. Također je bilo moguće dobiti poboljšane analoge prirodnih peptida sa selektivnim i pojačanim djelovanjem. Neki mali proteini su sintetizirani, kao što je hormon inzulin i neki enzimi.
Postoje i metode sinteze proteina koje repliciraju prirodne procese: sintetiziraju fragmente nukleinskih kiselina konfiguriranih da proizvode određene proteine, zatim se ti fragmenti ubacuju u živi organizam (na primjer, u bakteriju), nakon čega tijelo počinje proizvoditi željeni protein. Na ovaj način sada se dobijaju značajne količine teško dostupnih proteina i peptida, kao i njihovih analoga.
Proteini kao izvori hrane.
Proteini se u živom organizmu neprestano razlažu na svoje izvorne aminokiseline (uz neizostavno učešće enzima), neke aminokiseline prelaze u druge, zatim se ponovo sintetišu proteini (također uz učešće enzima), tj. tijelo se stalno obnavlja. Neki proteini (kolagen kože, kose) se ne obnavljaju, tijelo ih kontinuirano gubi i umjesto toga sintetiše nove. Proteini kao izvori hrane obavljaju dvije glavne funkcije: opskrbljuju tijelo građevinskim materijalom za sintezu novih proteinskih molekula i, osim toga, opskrbljuju tijelo energijom (izvori kalorija).
Sisavci mesožderi (uključujući ljude) dobijaju potrebne proteine iz biljne i životinjske hrane. Nijedan od proteina dobijenih hranom nije integrisan u organizam u nepromenjenom obliku. U probavnom traktu svi apsorbirani proteini se razgrađuju do aminokiselina i od njih su već izgrađeni proteini neophodni za određeni organizam, dok se preostalih 12 može sintetizirati iz 8 esencijalnih kiselina (Tabela 1) u tijelu ako nisu snabdjevene hranom u dovoljnim količinama, ali esencijalne kiseline moraju se obavezno snabdjeti hranom. Atome sumpora u cisteinu tijelo dobiva esencijalnom aminokiselinom metioninom. Dio proteina se razgrađuje, oslobađajući energiju potrebnu za održavanje života, a dušik koji se nalazi u njima se izlučuje iz tijela urinom. Obično ljudsko tijelo gubi 25-30 g proteina dnevno, tako da proteinska hrana mora uvijek biti prisutna u pravoj količini. Minimalne dnevne potrebe za proteinima su 37 g za muškarce i 29 g za žene, ali je preporučeni unos skoro duplo veći. Prilikom procjene hrane važno je uzeti u obzir kvalitet proteina. U nedostatku ili malom sadržaju esencijalnih aminokiselina, protein se smatra malovrijednim, pa takve proteine treba unositi u većim količinama. Dakle, proteini mahunarki sadrže malo metionina, a proteini pšenice i kukuruza imaju malo lizina (obe aminokiseline su esencijalne). Životinjski proteini (osim kolagena) klasifikovani su kao potpuna hrana. Kompletan set svih esencijalnih kiselina sadrži mlečni kazein, kao i svježi sir i sir pripremljen od njega, pa vegetarijanska ishrana, ako je veoma stroga, tj. “bez mliječnih proizvoda” zahtijeva povećanu konzumaciju mahunarki, orašastih plodova i gljiva kako bi se tijelo opskrbilo esencijalnim aminokiselinama u pravoj količini.
Sintetičke aminokiseline i proteini se također koriste kao prehrambeni proizvodi, dodajući ih u hranu za životinje, koja sadrži esencijalne aminokiseline u malim količinama. Postoje bakterije koje mogu prerađivati i asimilirati ugljovodonike nafte, u ovom slučaju, za potpunu sintezu proteina, potrebno ih je hraniti spojevima koji sadrže dušik (amonijak ili nitrati). Ovako dobijeni proteini koriste se kao hrana za stoku i živinu. U hranu za životinje često se dodaje skup enzima, karbohidrata, koji kataliziraju hidrolizu ugljikohidratnih komponenti hrane koje se teško razgrađuju (stanični zidovi žitarica), zbog čega se biljna hrana potpunije apsorbira.
Mikhail Levitsky
PROTEINI (član 2)
(proteini), klasa složenih spojeva koji sadrže dušik, najkarakterističnije i najvažnije (uz nukleinske kiseline) komponente žive tvari. Proteini obavljaju mnoge i različite funkcije. Većina proteina su enzimi koji kataliziraju kemijske reakcije. Mnogi hormoni koji regulišu fiziološke procese su takođe proteini. Strukturni proteini kao što su kolagen i keratin su glavne komponente koštanog tkiva, kose i noktiju. Kontraktilni proteini mišića imaju sposobnost da mijenjaju svoju dužinu koristeći hemijsku energiju za obavljanje mehaničkog rada. Proteini su antitijela koja vežu i neutraliziraju toksične tvari. Neki proteini koji mogu odgovoriti na vanjske utjecaje (svjetlo, miris) služe kao receptori u osjetilnim organima koji percipiraju iritaciju. Mnogi proteini koji se nalaze unutar ćelije i na ćelijskoj membrani obavljaju regulatorne funkcije.
U prvoj polovini 19. vijeka mnogi hemičari, a među njima prvenstveno J. von Liebig, postepeno su došli do zaključka da su proteini posebna klasa azotnih jedinjenja. Naziv "proteini" (od grčkog protos - prvi) predložio je 1840. holandski hemičar G. Mulder.
FIZIČKA SVOJSTVA
Proteini su bijeli u čvrstom stanju, ali bezbojni u otopini, osim ako ne nose neku hromofornu (obojenu) grupu, kao što je hemoglobin. Rastvorljivost različitih proteina u vodi uvelike varira. Takođe varira sa pH i koncentracijom soli u rastvoru, tako da se mogu izabrati uslovi pod kojima će jedan protein selektivno taložiti u prisustvu drugih proteina. Ova metoda "salanja" se široko koristi za izolaciju i pročišćavanje proteina. Pročišćeni protein se često taloži iz otopine u obliku kristala.
U poređenju sa drugim jedinjenjima, molekularna težina proteina je veoma velika - od nekoliko hiljada do mnogo miliona daltona. Stoga, tokom ultracentrifugiranja, proteini se talože, i to različitim brzinama. Zbog prisutnosti pozitivno i negativno nabijenih grupa u proteinskim molekulima, one se kreću različitim brzinama u električnom polju. Ovo je osnova elektroforeze, metode koja se koristi za izolaciju pojedinačnih proteina iz složenih mješavina. Pročišćavanje proteina se takođe vrši hromatografijom.
HEMIJSKA SVOJSTVA
Struktura.
Proteini su polimeri, tj. molekule izgrađene poput lanaca od ponavljajućih monomernih jedinica, ili podjedinica, čiju ulogu igraju alfa-amino kiseline. Opća formula aminokiselina
gdje je R atom vodika ili neka organska grupa.
Molekul proteina (polipeptidni lanac) može se sastojati od samo relativno malog broja aminokiselina ili nekoliko hiljada monomernih jedinica. Povezivanje aminokiselina u lanac je moguće jer svaka od njih ima dvije različite hemijske grupe: amino grupu sa osnovnim svojstvima, NH2, i kiselu karboksilnu grupu, COOH. Obje ove grupe su vezane za atom ugljika. Karboksilna grupa jedne amino kiseline može formirati amidnu (peptidnu) vezu sa amino grupom druge amino kiseline:
Nakon što su dvije aminokiseline povezane na ovaj način, lanac se može produžiti dodavanjem treće drugoj aminokiselini, i tako dalje. Kao što se može vidjeti iz gornje jednadžbe, kada se formira peptidna veza, oslobađa se molekul vode. U prisustvu kiselina, lužina ili proteolitičkih enzima, reakcija se odvija u suprotnom smjeru: polipeptidni lanac se cijepa na aminokiseline uz dodatak vode. Ova reakcija se naziva hidroliza. Hidroliza se odvija spontano, a energija je potrebna za spajanje aminokiselina u polipeptidni lanac.
Karboksilna grupa i amidna grupa (ili njoj slična imidna grupa - u slučaju aminokiseline prolina) prisutne su u svim aminokiselinama, dok su razlike između aminokiselina određene prirodom te grupe, odnosno "strane lanac", koji je gore označen slovom R. Ulogu bočnog lanca može igrati jedan atom vodonika, poput aminokiseline glicin, i neke glomazne grupe, poput histidina i triptofana. Neki bočni lanci su hemijski inertni, dok su drugi visoko reaktivni.
Mnogo hiljada različitih aminokiselina se može sintetizirati, i mnogo različitih aminokiselina se javlja u prirodi, ali samo 20 vrsta aminokiselina se koristi za sintezu proteina: alanin, arginin, asparagin, asparaginska kiselina, valin, histidin, glicin, glutamin, glutamin kiselina, izoleucin, leucin, lizin, metionin, prolin, serin, tirozin, treonin, triptofan, fenilalanin i cistein (u proteinima cistein može biti prisutan kao dimer-cistin). Istina, u nekim proteinima postoje i druge aminokiseline pored dvadesetak koje se redovno pojavljuju, ali one nastaju kao rezultat modifikacije bilo koje od dvadeset navedenih nakon što je ona uključena u protein.
optička aktivnost.
Sve aminokiseline, sa izuzetkom glicina, imaju četiri različite grupe vezane za atom α-ugljika. Što se tiče geometrije, četiri različite grupe se mogu pričvrstiti na dva načina, pa su prema tome moguće dvije konfiguracije, odnosno dva izomera, koji su međusobno povezani kao objekt na njegovu sliku u ogledalu, tj. kao leva ruka desno. Jedna konfiguracija se naziva lijeva, ili ljevoruka (L), a druga desnoruka, ili desnoruka (D), jer se dva takva izomera razlikuju u smjeru rotacije ravni polarizirane svjetlosti. U proteinima se javljaju samo L-aminokiseline (izuzetak je glicin; može se predstaviti samo u jednom obliku, jer su dvije od njegove četiri grupe iste), i sve imaju optičku aktivnost (pošto postoji samo jedan izomer). D-amino kiseline su rijetke u prirodi; nalaze se u nekim antibioticima i ćelijskom zidu bakterija.
Redoslijed aminokiselina.
Aminokiseline u polipeptidnom lancu nisu raspoređene nasumično, već u određenom fiksnom redoslijedu i upravo taj redoslijed određuje funkcije i svojstva proteina. Promjenom redoslijeda od 20 vrsta aminokiselina, možete dobiti ogroman broj različitih proteina, kao što možete sastaviti mnogo različitih tekstova od slova abecede.
U prošlosti je određivanje sekvence aminokiselina proteina često trajalo nekoliko godina. Direktno određivanje je još uvijek prilično naporan zadatak, iako su stvoreni uređaji koji omogućavaju da se izvrši automatski. Obično je lakše odrediti nukleotidnu sekvencu odgovarajućeg gena i iz njega izvući sekvencu aminokiselina proteina. Do danas su aminokiselinske sekvence mnogih stotina proteina već određene. Funkcije dekodiranih proteina su obično poznate, a to pomaže da se zamisle moguće funkcije sličnih proteina nastalih, na primjer, u malignim neoplazmama.
Kompleksni proteini.
Proteini koji se sastoje samo od aminokiselina nazivaju se jednostavnim. Međutim, često je atom metala ili neko hemijsko jedinjenje koje nije aminokiselina vezano za polipeptidni lanac. Takvi proteini se nazivaju kompleksni. Primjer je hemoglobin: sadrži željezni porfirin, koji mu daje crvenu boju i omogućava mu da djeluje kao prijenosnik kisika.
Nazivi najsloženijih proteina sadrže indikaciju prirode vezanih grupa: šećeri su prisutni u glikoproteinima, masti u lipoproteinima. Ako katalitička aktivnost enzima ovisi o spojenoj grupi, onda se to naziva prostetičkom grupom. Često neki vitamin igra ulogu protetske grupe ili je njen dio. Vitamin A, na primjer, vezan za jedan od proteina retine, određuje njegovu osjetljivost na svjetlost.
Tercijarna struktura.
Ono što je važno nije toliko aminokiselinska sekvenca proteina (primarna struktura), već način na koji je položen u svemiru. Po cijeloj dužini polipeptidnog lanca ioni vodika formiraju pravilne vodikove veze, koje mu daju oblik spirale ili sloja (sekundarna struktura). Iz kombinacije takvih spirala i slojeva nastaje kompaktni oblik sljedećeg reda - tercijarna struktura proteina. Oko veza koje drže monomerne karike lanca moguće su rotacije pod malim uglovima. Stoga, sa čisto geometrijske tačke gledišta, broj mogućih konfiguracija za bilo koji polipeptidni lanac je beskonačno velik. U stvarnosti, svaki protein normalno postoji u samo jednoj konfiguraciji, određenoj njegovom aminokiselinskom sekvencom. Ova struktura nije kruta, čini se da "diše" - oscilira oko određene prosječne konfiguracije. Lanac je presavijen u konfiguraciju u kojoj je slobodna energija (sposobnost obavljanja rada) minimalna, kao što je oslobođena opruga komprimirana samo do stanja koje odgovara minimumu slobodne energije. Često je jedan dio lanca čvrsto povezan s drugim disulfidnim (–S–S–) vezama između dva cisteinska ostatka. To je dijelom razlog zašto cistein među aminokiselinama igra posebno važnu ulogu.
Složenost strukture proteina je toliko velika da još nije moguće izračunati tercijarnu strukturu proteina, čak i ako je poznata njegova aminokiselinska sekvenca. Ali ako je moguće dobiti proteinske kristale, onda se njegova tercijarna struktura može odrediti difrakcijom X zraka.
U strukturnim, kontraktilnim i nekim drugim proteinima, lanci su izduženi i nekoliko blago presavijenih lanaca koji leže jedan pored drugog formiraju fibrile; fibrile se zauzvrat savijaju u veće formacije - vlakna. Međutim, većina proteina u rastvoru je globularna: lanci su umotani u kuglu, poput pređe u klupko. Slobodna energija sa ovom konfiguracijom je minimalna, budući da su hidrofobne ("vodoodbojne") aminokiseline skrivene unutar globule, a hidrofilne ("privlače vodu") aminokiseline su na njenoj površini.
Mnogi proteini su kompleksi nekoliko polipeptidnih lanaca. Ova struktura se naziva kvartarna struktura proteina. Molekul hemoglobina, na primjer, sastoji se od četiri podjedinice, od kojih je svaka globularni protein.
Strukturni proteini zbog svoje linearne konfiguracije formiraju vlakna u kojima je vlačna čvrstoća vrlo visoka, dok globularna konfiguracija omogućava proteinima da uđu u specifične interakcije sa drugim spojevima. Na površini globule, uz pravilno polaganje lanaca, pojavljuju se šupljine određenog oblika, u kojima se nalaze reaktivne kemijske grupe. Ako je ovaj protein enzim, onda drugi, obično manji, molekul neke supstance ulazi u takvu šupljinu, baš kao što ključ ulazi u bravu; u ovom slučaju se konfiguracija elektronskog oblaka molekula mijenja pod utjecajem kemijskih grupa koje se nalaze u šupljini, a to ga prisiljava da reagira na određeni način. Na taj način enzim katalizuje reakciju. Molekuli antitijela također imaju šupljine u koje se vežu različite strane tvari i na taj način postaju bezopasne. Model "ključ i brava", koji objašnjava interakciju proteina sa drugim spojevima, omogućava razumijevanje specifičnosti enzima i antitijela, tj. njihova sposobnost da reaguju samo sa određenim jedinjenjima.
Proteini u različitim vrstama organizama.
Proteini koji obavljaju istu funkciju u različitim biljnim i životinjskim vrstama i stoga nose isto ime također imaju sličnu konfiguraciju. Oni se, međutim, donekle razlikuju u svojoj sekvenci aminokiselina. Kako se vrste razlikuju od zajedničkog pretka, neke aminokiseline na određenim pozicijama zamjenjuju se mutacijama s drugim. Štetne mutacije koje uzrokuju nasljedne bolesti prirodnom selekcijom odbacuju se, ali se korisne ili barem neutralne mogu sačuvati. Što su dvije biološke vrste bliže jedna drugoj, to je manje razlika u njihovim proteinima.
Neki proteini se mijenjaju relativno brzo, drugi su prilično konzervativni. Potonji uključuju, na primjer, citokrom c, respiratorni enzim koji se nalazi u većini živih organizama. Kod ljudi i čimpanzi, njegove aminokiselinske sekvence su identične, dok se u citokromu c pšenice pokazalo da je samo 38% aminokiselina drugačije. Čak i kada se porede ljudi i bakterije, sličnost citokroma sa (razlike ovde utiču na 65% aminokiselina) još uvek se može videti, iako je zajednički predak bakterija i ljudi živeo na Zemlji pre oko dve milijarde godina. Danas se poređenje aminokiselinskih sekvenci često koristi za izgradnju filogenetskog (genealoškog) stabla koje odražava evolucijske odnose između različitih organizama.
Denaturacija.
Sintetizirani proteinski molekul, savijajući se, poprima vlastitu konfiguraciju. Ova se konfiguracija, međutim, može uništiti zagrijavanjem, promjenom pH, djelovanjem organskih rastvarača, pa čak i jednostavnim miješanjem otopine dok se na njenoj površini ne pojave mjehurići. Protein izmijenjen na ovaj način naziva se denaturiranim; gubi svoju biološku aktivnost i obično postaje nerastvorljiv. Dobro poznati primjeri denaturiranih proteina su kuhana jaja ili šlag. Mali proteini, koji sadrže samo stotinjak aminokiselina, sposobni su da se renaturiraju, tj. ponovo nabavite originalnu konfiguraciju. Ali većina proteina se jednostavno transformiše u masu zapletenih polipeptidnih lanaca i ne obnavlja svoju prethodnu konfiguraciju.
Jedna od glavnih poteškoća u izolaciji aktivnih proteina je njihova ekstremna osjetljivost na denaturaciju. Ovo svojstvo proteina nalazi korisnu primjenu u konzerviranju prehrambenih proizvoda: visoka temperatura nepovratno denaturira enzime mikroorganizama i mikroorganizmi umiru.
SINTEZA PROTEINA
Za sintezu proteina, živi organizam mora imati sistem enzima sposobnih da vežu jednu aminokiselinu za drugu. Potreban je i izvor informacija koji bi odredio koje aminokiseline treba povezati. Budući da u tijelu postoje hiljade vrsta proteina, a svaki od njih se sastoji u prosjeku od nekoliko stotina aminokiselina, potrebne informacije moraju biti zaista ogromne. Pohranjuje se (slično kao što se zapis čuva na magnetnoj traci) u molekulima nukleinske kiseline koji čine gene.
Aktivacija enzima.
Polipeptidni lanac sintetiziran iz aminokiselina nije uvijek protein u svom konačnom obliku. Mnogi enzimi se prvo sintetiziraju kao neaktivni prekursori i postaju aktivni tek nakon što drugi enzim ukloni nekoliko aminokiselina s jednog kraja lanca. Neki od probavnih enzima, kao što je tripsin, sintetiziraju se u ovom neaktivnom obliku; ovi enzimi se aktiviraju u probavnom traktu kao rezultat uklanjanja terminalnog fragmenta lanca. Hormon insulin, čija se molekula u svom aktivnom obliku sastoji od dva kratka lanca, sintetiše se u obliku jednog lanca, tzv. proinsulin. Zatim se srednji dio ovog lanca uklanja, a preostali fragmenti se vežu jedni za druge, formirajući aktivni molekul hormona. Složeni proteini nastaju tek nakon što je određena hemijska grupa vezana za protein, a za to vezivanje često je potreban i enzim.
Metabolička cirkulacija.
Nakon hranjenja životinje aminokiselinama označenim radioaktivnim izotopima ugljika, dušika ili vodika, oznaka se brzo ugrađuje u njene proteine. Ako označene aminokiseline prestanu da ulaze u tijelo, tada se količina oznaka u proteinima počinje smanjivati. Ovi eksperimenti pokazuju da se dobijeni proteini ne pohranjuju u tijelu do kraja života. Svi su, uz nekoliko izuzetaka, u dinamičnom stanju, neprestano se razlažu do aminokiselina, a zatim ponovo sintetiziraju.
Neki proteini se razgrađuju kada ćelije umru i budu uništene. To se stalno događa, na primjer, s crvenim krvnim zrncima i epitelnim stanicama koje oblažu unutrašnju površinu crijeva. Pored toga, razgradnja i resinteza proteina se dešavaju iu živim ćelijama. Začudo, manje se zna o razgradnji proteina nego o njihovoj sintezi. Ono što je, međutim, jasno je da su proteolitički enzimi uključeni u razgradnju, slično onima koji razlažu proteine u aminokiseline u probavnom traktu.
Poluživot različitih proteina je različit - od nekoliko sati do više mjeseci. Jedini izuzetak su molekule kolagena. Jednom formirane, ostaju stabilne i ne obnavljaju se ili zamjenjuju. Vremenom se, međutim, mijenjaju neka njihova svojstva, a posebno elastičnost, a kako se ne obnavljaju, rezultat toga su određene promjene vezane za starenje, na primjer, pojava bora na koži.
sintetički proteini.
Hemičari su odavno naučili kako da polimeriziraju aminokiseline, ali se aminokiseline nasumično kombinuju, tako da proizvodi takve polimerizacije malo liče na prirodne. Istina, moguće je kombinirati aminokiseline određenim redoslijedom, što omogućava dobivanje nekih biološki aktivnih proteina, posebno inzulina. Proces je prilično kompliciran i na taj način se mogu dobiti samo oni proteini čiji molekuli sadrže oko stotinu aminokiselina. Umjesto toga, poželjno je sintetizirati ili izolirati nukleotidnu sekvencu gena koja odgovara željenoj aminokiselinskoj sekvenci, a zatim uvesti ovaj gen u bakteriju, koja će replikacijom proizvesti veliku količinu željenog proizvoda. Ova metoda, međutim, ima i svoje nedostatke.
PROTEINI I ISHRANA
Kada se proteini u tijelu razgrade na aminokiseline, ove aminokiseline se mogu ponovo koristiti za sintezu proteina. Istovremeno, same aminokiseline su podložne raspadanju, tako da se ne koriste u potpunosti. Takođe je jasno da tokom rasta, trudnoće i zarastanja rana, sinteza proteina mora biti veća od razgradnje. Tijelo kontinuirano gubi neke proteine; to su proteini kose, noktiju i površinskog sloja kože. Stoga, za sintezu proteina, svaki organizam mora dobiti aminokiseline iz hrane.
Izvori aminokiselina.
Zelene biljke sintetiziraju svih 20 aminokiselina koje se nalaze u proteinima iz CO2, vode i amonijaka ili nitrata. Mnoge bakterije su također sposobne sintetizirati aminokiseline u prisustvu šećera (ili nekog ekvivalenta) i fiksiranog dušika, ali šećer u konačnici isporučuju zelene biljke. Kod životinja je sposobnost sinteze aminokiselina ograničena; dobijaju aminokiseline jedući zelene biljke ili druge životinje. U probavnom traktu apsorbirani proteini se razlažu na aminokiseline, potonje se apsorbiraju i od njih se izgrađuju proteini karakteristični za dati organizam. Nijedan od apsorbiranih proteina nije ugrađen u strukture tijela kao takve. Jedini izuzetak je da kod mnogih sisara dio majčinih antitijela može proći netaknut kroz placentu u fetalnu cirkulaciju, a kroz majčino mlijeko (posebno kod preživara) se prenijeti na novorođenče odmah nakon rođenja.
Potreba za proteinima.
Jasno je da za održavanje života tijelo mora primiti određenu količinu proteina iz hrane. Međutim, veličina ove potrebe ovisi o brojnim faktorima. Tijelu je potrebna hrana i kao izvor energije (kalorije) i kao materijal za izgradnju svojih struktura. Na prvom mjestu je potreba za energijom. To znači da kada u prehrani ima malo ugljikohidrata i masti, dijetetski proteini se ne koriste za sintezu vlastitih proteina, već kao izvor kalorija. Uz produženo gladovanje, čak se i vaši vlastiti proteini troše za zadovoljavanje energetskih potreba. Ako u ishrani ima dovoljno ugljikohidrata, onda se unos proteina može smanjiti.
balans azota.
U prosjeku cca. 16% ukupne mase proteina je dušik. Kada se aminokiseline koje čine proteine razgrađuju, dušik koji se nalazi u njima izlučuje se iz tijela mokraćom i (u manjoj mjeri) fecesom u obliku različitih azotnih spojeva. Stoga je prikladno koristiti takav pokazatelj kao što je ravnoteža dušika za procjenu kvalitete proteinske prehrane, tj. razlika (u gramima) između količine dušika unesenog u tijelo i količine azota izlučenog dnevno. Uz normalnu ishranu odrasle osobe, ove količine su jednake. U rastućem organizmu količina izlučenog azota je manja od količine dolaznog, tj. bilans je pozitivan. Sa nedostatkom proteina u ishrani, balans je negativan. Ako u ishrani ima dovoljno kalorija, a proteini u njoj potpuno nedostaju, tijelo štedi proteine. Istovremeno, metabolizam proteina se usporava, a ponovno korištenje aminokiselina u sintezi proteina odvija se što je moguće efikasnije. Međutim, gubici su neizbježni, a dušični spojevi se i dalje izlučuju urinom, a dijelom i izmetom. Količina azota koji se dnevno izluči iz organizma tokom proteinskog gladovanja može poslužiti kao mjera dnevnog nedostatka proteina. Prirodno je pretpostaviti da je unošenjem u ishranu količine proteina koja je ekvivalentna ovom nedostatku, moguće vratiti ravnotežu dušika. Međutim, nije. Nakon što primi ovu količinu proteina, tijelo počinje manje efikasno da koristi aminokiseline, pa je potreban dodatni protein da bi se uspostavila ravnoteža dušika.
Ako količina proteina u prehrani premašuje ono što je potrebno za održavanje ravnoteže dušika, čini se da od toga nema štete. Višak aminokiselina jednostavno se koristi kao izvor energije. Posebno upečatljiv primjer su Eskimi, koji konzumiraju malo ugljikohidrata i oko deset puta više proteina nego što je potrebno za održavanje ravnoteže dušika. U većini slučajeva, međutim, korištenje proteina kao izvora energije nije korisno, jer možete dobiti mnogo više kalorija iz određene količine ugljikohidrata nego iz iste količine proteina. U siromašnim zemljama stanovništvo prima potrebne kalorije iz ugljikohidrata i konzumira minimalnu količinu proteina.
Ako tijelo dobije potreban broj kalorija u obliku neproteinskih proizvoda, tada je minimalna količina proteina koja održava ravnotežu dušika cca. 30 g dnevno. Otprilike isto toliko proteina sadrži četiri kriške hljeba ili 0,5 litara mlijeka. Nešto veća količina se obično smatra optimalnom; preporučuje se od 50 do 70 g.
Esencijalne aminokiseline.
Do sada se protein smatrao kao cjelina. U međuvremenu, da bi se odvijala sinteza proteina, u tijelu moraju biti prisutne sve potrebne aminokiseline. Neke od aminokiselina koje tijelo same životinje može sintetizirati. Nazivaju se zamjenjivima, jer ne moraju biti prisutni u ishrani – važno je samo da je općenito dovoljan unos proteina kao izvora dušika; tada, uz nedostatak neesencijalnih aminokiselina, tijelo ih može sintetizirati na račun onih koje su prisutne u višku. Preostale "esencijalne" aminokiseline se ne mogu sintetizirati i moraju se unijeti hranom. Esencijalni za ljude su valin, leucin, izoleucin, treonin, metionin, fenilalanin, triptofan, histidin, lizin i arginin. (Iako se arginin može sintetizirati u tijelu, smatra se esencijalnom aminokiselinom jer novorođenčad i djeca u rastu proizvode ga nedovoljne količine. S druge strane, za osobu zrele dobi unos neke od ovih aminokiselina iz hrane može postati opciono.)
Ova lista esencijalnih aminokiselina je približno ista kod drugih kralježnjaka, pa čak i kod insekata. Nutritivna vrijednost proteina se obično određuje hranjenjem pacova koji rastu i praćenjem povećanja tjelesne težine životinja.
Nutritivna vrijednost proteina.
Nutritivnu vrijednost proteina određuje esencijalna aminokiselina koja je najnedostatnija. Ilustrirajmo to primjerom. Proteini našeg tijela sadrže u prosjeku cca. 2% triptofana (po težini). Recimo da ishrana uključuje 10 g proteina koji sadrže 1% triptofana i da u njoj ima dovoljno drugih esencijalnih aminokiselina. U našem slučaju, 10 g ovog defektnog proteina je u suštini ekvivalentno 5 g potpunog; preostalih 5 g može poslužiti samo kao izvor energije. Imajte na umu da s obzirom da se aminokiseline praktički ne skladište u tijelu, a da bi se odvijala sinteza proteina, sve aminokiseline moraju biti prisutne istovremeno, efekat unosa esencijalnih aminokiselina može se otkriti samo ako sve one uđu u telo u isto vreme.
Prosječan sastav većine životinjskih bjelančevina je blizak prosječnom sastavu bjelančevina ljudskog tijela, pa je malo vjerovatno da ćemo se suočiti s nedostatkom aminokiselina ako je naša ishrana bogata namirnicama poput mesa, jaja, mlijeka i sira. Međutim, postoje proteini, poput želatine (proizvod denaturacije kolagena), koji sadrže vrlo malo esencijalnih aminokiselina. Biljni proteini, iako su u tom smislu bolji od želatine, takođe su siromašni esencijalnim aminokiselinama; posebno malo u njima lizina i triptofana. Ipak, čisto vegetarijanska ishrana nije nimalo štetna, osim ako ne konzumira nešto veću količinu biljnih proteina, dovoljnu da obezbedi organizam esencijalnim aminokiselinama. Najviše proteina nalazi se u biljkama u sjemenkama, posebno u sjemenu pšenice i raznih mahunarki. Mladi izdanci, kao što su špargle, takođe su bogati proteinima.
Sintetički proteini u ishrani.
Dodavanjem malih količina sintetičkih esencijalnih aminokiselina ili proteina bogatih njima nepotpunim proteinima, kao što su proteini kukuruza, moguće je značajno povećati nutritivnu vrijednost ovih potonjih, tj. čime se povećava količina konzumiranih proteina. Druga mogućnost je uzgoj bakterija ili kvasaca na naftnim ugljovodonicima uz dodatak nitrata ili amonijaka kao izvora dušika. Mikrobni proteini dobijeni na ovaj način mogu poslužiti kao hrana za živinu ili stoku, ili ih ljudi mogu direktno konzumirati. Treća, široko korištena, metoda koristi fiziologiju preživara. Kod preživača se u početnom dijelu želuca javlja tzv. Burag je naseljen posebnim oblicima bakterija i protozoa koji defektne biljne proteine pretvaraju u potpunije mikrobne proteine, a ovi se, nakon probave i apsorpcije, pretvaraju u životinjske proteine. Urea, jeftino sintetičko jedinjenje koje sadrži dušik, može se dodati u stočnu hranu. Mikroorganizmi koji žive u buragu koriste dušik uree za pretvaranje ugljikohidrata (kojih ima mnogo više u hrani) u proteine. Otprilike trećina ukupnog dušika u stočnoj hrani može doći u obliku uree, što u suštini znači, u određenoj mjeri, hemijsku sintezu proteina.
 Rune, glagoljica, ćirilica: šta su Ćirilo i Metodije zapravo izmislili Ko se smatra tvorcem staroslovenskog pisma
Rune, glagoljica, ćirilica: šta su Ćirilo i Metodije zapravo izmislili Ko se smatra tvorcem staroslovenskog pisma Kako se nositi sa umorom na poslu
Kako se nositi sa umorom na poslu Jeljcinisti koji su izgubili pamćenje: ko su Vlasovci Nacionalni sastav vojske Vlasovske roa
Jeljcinisti koji su izgubili pamćenje: ko su Vlasovci Nacionalni sastav vojske Vlasovske roa