Чем является комплекс гольджи в растительных клетках. ЭПС и комплекс Гольджи
Клетка — цельная система
Живая клетка — уникальная совершенная мельчайшая единица организма, она устроена так, чтобы максимально эффективно использовать кислород и питательные вещества, выполняя свои функции. Жизненно важными для клетки органеллами являются ядро, рибосомы, митохондрии, эндоплазматический ретикулум, аппарат Гольджи. Вот о последнем и поговорим подробнее.
Что это такое
Эта мембранная органелла является комплексом структур, которые выводят из клетки синтезированные в ней вещества. Чаще всего она располагается вблизи от наружной клеточной мембраны.
Аппарат Гольджи: строение
Он состоит из образованных мембранами “мешочков”, называемых цистернами. Последние имеют вытянутую форму, слегка сплющены посередине и расширены по краям. Также в комплексе имеются круглые пузырьки Гольджи - мелкие мембранные структуры. Цистерны “сложены” стопочками, которые называются диктиосомы. Аппарат Гольджи содержит различные типы “мешочков”, весь комплекс делят на некоторые части по степени удаленности от ядра. Различают их три: цис-отдел (ближе к ядру), срединный, и транс-отдел - самый дальний от ядра. Они характеризуются отличающимся составом ферментов, а следовательно, и выполняемой работой. В строении диктиосом есть одна особенность: они полярны, то есть ближайший к ядру отдел только принимает пузырьки, идущие от эндоплазматического ретикулума. Часть “стопки”, обращенная к мембране клетки, только формирует и отдает их.

Аппарат Гольджи: функции
Основными выполняемыми задачами являются сортировка белков, липидов, слизистых секретов и их выведение. Также через него проходят выделяемые клеткой небелковые вещества, углеводные компоненты наружной мембраны. При этом аппарат Гольджи вовсе не является индифферентным посредником, который просто “передает” вещества, в нем идут процессы активизации и модификации (“созревания”):
- Сортировка веществ, транспорт белков. Распределение белковых веществ происходит на три потока: для мембраны самой клетки, экспортные, лизосомальные ферменты. В первый поток помимо белков включаются и жиры. Интересный факт, что любые экспортные вещества переносятся внутри пузырьков. А вот предназначенные для мембраны клетки белки встраиваются в мембрану транспортного пузырька и перемещаются таким образом.
- Выделение всех продуктов, произведенных в клетке. Аппарат Гольджи “упаковывает” всю продукцию, как белковую, так и иной природы, в секреторные пузырьки. Все вещества выделяются наружу путем сложного взаимодействия последних с клеточной мембраной.
- Синтез полисахаридов (гликозаминогликанов и компонентов гликокаликса клеточной стенки).
- Сульфатирование, гликозилирование жиров и белков, частичный протеолиз последних (необходимый для перевода их из неактивной формы в активную), — это всё процессы “созревания” белков, нужные для их будущей полноценной работы.

В заключение
Рассмотрев то, как устроен и работает комплекс Гольджи, убеждаемся, что он является важнейшей и неотъемлемой частью любой клетки (особенно секреторных). Клетка, не продуцирующая веществ на экспорт, также не может обойтись без этой органеллы, поскольку от нее зависит “укомплектованность” клеточной мембраны и другие важные внутренние процессы жизнедеятельности.
Комплекс Гольджи был обнаружен Камилло Гольджи в 1898 году. Данная структура присутствует в цитоплазме практически всех эукариотических (составляющих высшие организмы) клеток, в особенности, секреторных клеток у животных.
Комплекс Гольджи. Строение.
Структура представлена стопкой уплощенных мешочков мембран. Они называются цистерны. Эта стопка мешочков связана с системой Гольджи). С одного конца стопок мешочков постоянно происходит формирование новых цистерн слиянием пузырьков, которые отпочковываются от эндоплазматического ретикулума (сети полостей). С другого же конца стопки с внутренней стороны цистерны завершают созревание и распадаются вновь на пузырьки. Так происходит постепенное перемещение цистерн в сопке к внутренней стороне от стороны наружной.
В цистернах структуры происходит созревание белков, предназначенных для секреции, трансмембранных белков белков лизосом и прочих. Созревающие вещества перемещаются последовательно по цистернам органеллы. В них происходит окончательное свертывание белков и их модификации - фосфорилирование и гликозилирование.
Характеризуются наличием ряда отдельных диктиосом (стопок). В часто присутствует несколько стопок соединенных трубками или одна большая стопка.
Содержит четыре главных отдела: транс-Гольджи сеть, цис-Гольджи, транс-Гольджи и медиал-Гольджи. К структуре присоединяют также и промежуточный компартмент (обособленную область). Он представлен скоплением мембранных пузырьков в промежутке между реиткулумом и цис-Гольджи.
Весь аппарат является органеллой весьма полиморфной (многообразной). Даже на различных этапах развития одной клетки комплекс Гольджи может по-разному выглядеть.
Отличается аппарат и асимметрией. Располагающиеся ближе к клеточному ядру цистерны (цис-Гольджи) содержат в себе самые незрелые белки. К цистернам этим присоединяются непрерывно мембранные пузырьки - везикулы. В разных цистернах содержатся различные резидентные ферменты (каталитические), что говорит о том, что в них с созревающими белками происходят последовательно разные процессы.
Комплекс Гольджи. Функции.
К задачам структуры относятся химическая модификация и транспорт веществ, поступающих в него. Белки, проникающие в аппарат из эндоплазматического ретикулума, являются для ферментов исходным субстратом. Пройдя концентрирование и модификацию, ферменты в пузырьках транспортируются к назначенному участку. Например, это может быть область формирования новой почки. При участии цитоплазматических микротрубочек процесс переноса осуществляется наиболее активно.
Комплекс Гольджи выполняет также задачу по присоединению углеводных групп к белкам и последующему использованию этих белков при построении мембраны лизосом и клеток.
У отдельных водорослей в структуре аппарата осуществляется синтез целлюлозных волокон.
Функции комплекса Гольджи достаточно разнообразны. Среди них следует выделить:
- Сортировку, выведение, накопление секреторных продуктов.
- Накопление липидных молекул и формирование липопротеидов.
- Завершение модификации белков (посттрансляционной), а именно гликозирования, сульфатирования и прочего.
- Формирование лизосом.
- Участие в образовании акросомы.
- Полисахаридный синтез для формирования восков, гликопротеидов, слизей, камеди, веществ матрикса в растений (пектинов, гемицеллюлозы и прочих).
- Образование сократимых вакуолей у простейших.
- Образование клеточной пластинки в растительных клетках после деления ядра.
18. Комплекс Гольджи, его строение и функции. Лизосомы. Их строение и функции. Типы лизосом.
Комплекс Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям, и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько, соединённых трубками, стопок.
1. Накапливает и выводит органические вещества, синтезируемые в эндоплазматической сети
2. Образует лизосомы
3. Формирование углеводных компонентов гликокаликса - в основном, гликолипидов.
Лизосомы представляют собой неотъемлемую часть состава клетки. Они являются разновидностью везикул. Эти клеточные помощники, являясь частью вакуома, покрыты оболочкой из мембраны и наполнены гидролитическими ферментами. Важность существования лизосом внутри клетки обеспечена секреторной функцией, которая необходима в процессе фагоцитоза и аутофагоцитоза.
Выполняют пищеварительную функцию - переваривают пищевые частицы и удаляют отмершие органоиды.
Первичные лизосомы - это мелкие мембранные пузырьки, которые имеют деаметр около ста нм, заполненные гомогенным мелкодисперсным содержимым, являющим собой набор гидролитических ферментов. В лизосомах есть около сорока ферментов.
Вторичные лизосомы образуются при слиянии первичных лизосом с эндоцитозными либо с пиноцитозными вакуолями. Если сказать иначе, то вторичные лизосомы - это внутриклеточные пищеварительные вакуоли, ферменты которых поставляются первичными лизосомами, а материал для переваривания - эндоцитозной (пиноцитозной) вакуолью.
19. Эпс, ее разновидности, роль в процессах синтеза веществ.
Эндоплазматическая сеть в разных клетках может быть представлена в форме уплощенных цистерн, канальцев или отдельных везикул. Стенка этих образований состоит из билипидной мембраны и включенных в нее некоторых белков и отграничивает внутреннюю среду эндоплазматической сети от гиалоплазмы.
Различают две разновидности эндоплазматической сети:
зернистая (гранулярная или шероховатая);
незернистая или гладкая.
На наружной поверхности мембран зернистой эндоплазматической сети содержатся прикрепленные рибосомы. В цитоплазме могут быть обе разновидности эндоплазматической сети, но обычно преобладает одна форма, что и обуславливает функциональную специфичность клетки. Следует помнить, что названные две разновидности являются не самостоятельными формами эндоплазматической сети, так как можно проследить переход зернистой эндоплазматической сети в гладкую и наоборот.
Функции зернистой эндоплазматической сети:
синтез белков, предназначенных для выведения из клетки ("на экспорт");
отделение (сегрегация) синтезированного продукта от гиалоплазмы;
конденсация и модификация синтезированного белка;
транспорт синтезированных продуктов в цистерны пластинчатого комплекса или непосредственно из клетки;
синтез билипидных мембран.
Гладкая эндоплазматическая сеть представлена цистернами, более широкими каналами и отдельными везикулами, на внешней поверхности которых отсутствуют рибосомы.
Функции гладкой эндоплазматической сети:
участие в синтезе гликогена;
синтез липидов;
дезинтоксикационная функция - нейтрализация токсических веществ, посредством соединения их с другими веществами.
Пластинчатый комплекс Гольджи (сетчатый аппарат) представлен скоплением уплощенных цистерн и небольших везикул, ограниченных билипидной мембраной. Пластинчатый комплекс подразделяется на субъединицы - диктиосомы. Каждая диктиосома представляет собой стопку уплощенных цистерн, по периферии которых локализуются мелкие пузырьки. При этом, в каждой уплощенной цистерне периферическая часть несколько расширена, а центральная сужена.
В 1898 г. итальянский ученый К. Гольджи, используя свойства связывания тяжелых металлов (осмия и серебра) с клеточными структурами, выявил в нервных клетках сетчатые образования, которые он назвал “внутренним сетчатым аппаратом” (рис. 174). Дальнейшее усовершенствование метода окраски металлами (импрегнации) дало возможность убедиться, что сетчатые структуры (аппарат Гольджи) встречаются во всех клетках любых эукариотных организмов. Обычно элементы аппарата Гольджи расположены около ядра, вблизи клеточного центра (центриоли). Участки аппарата Гольджи, четко выявляемые методом импрегнации, имели в некоторых клетках вид сложных сетей, где ячейки были связаны друг с другом или представлялись в виде отдельных темных участков, лежащих независимо друг от друга (диктиосомы), имеющих вид палочек, зерен, вогнутых дисков и т.д. (рис. 175). Между сетчатой и диффузной формой аппарата Гольджи нет принципиального различия, так как часто в одних и тех же клетках наблюдается смена форм этого органоида. Элементы аппарата Гольджи часто связаны с вакуолями, что особенно характерно для секретирующих клеток.
Было обнаружено, что морфология АГ меняется в зависимости от стадий клеточной секреции, что послужило основанием Д.Н. Насонову (1924) выдвинуть гипотезу о том, что АГ является органоидом, обеспечивающим сепарацию и накопление веществ в самых различных клетках.
Долгое время в растительных клетках не удавалось обнаружить элементов аппарата Гольджи обычными методами микротехники. Однако с появлением метода электронной микроскопии элементы АГ были обнаружены во всех растительных клетках, где они расположены по периферии клетки.
Тонкое строение аппарата Гольджи
В электронном микроскопе видно, что аппарат Гольджи представлен мембранными структурами, собранными вместе в небольшой зоне (рис. 176, 177). Отдельная зона скопления этих мембран является диктиосомой (рис. 178). В диктиосоме плотно друг к другу (на расстоянии 20-25 нм) расположены в виде стопки плоские мембранные мешки, или цистерны, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм и переменную толщину; в центре ее мембраны могут быть сближены (25 нм), а на периферии иметь расширения, ампулы, ширина которых непостоянна. Количество таких мешков в стопке обычно не превышает 5-10. У некоторых одноклеточных их число может достигать 20 штук. Кроме плотно расположенных плоских цистерн в зоне АГ наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках зоны АГ; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн. Принято различать в зоне диктиосомы проксимальный или формирующийся, цис-участок, и дистальный или зрелый, транс-участок (рис. 178). Между ними располагается средний или промежуточный участок АГ.
Во время деления клеток сетчатые формы АГ распадаются до диктиосом, которые пассивно и случайно распределяются по дочерним клеткам. При росте клеток общее количество диктиосом увеличивается.
В секретирующих клетках обычно АГ поляризован: его проксимальная часть обращена к цитоплазме и ядру, а дистальная - к поверхности клетки. В проксимальном участке к стопкам сближенных цистерн примыкает зона мелких гладких пузырьков и коротких мембранных цистерн. В образцах препаративно выделенных зон АГ при негативном контрастировании видно, что к проксимальной части диктиосомы примыкает сетевидная или губкообразная система мембранных полостей. Считается, что эта система может представлять собой зону перехода элементов ЭР в зону аппарата Гольджи (рис. 179).
В средней части диктиосомы периферия каждой цистерны также сопровождается массой мелких вакуолей около 50 нм в диаметре.
В дистальном или транс-участке диктиосом к последней мембранной плоской цистерне примыкает участок, состоящий из трубчатых элементов и массой мелких вакуолей, часто имеющих фибриллярную опушенность по поверхности со стороны цитоплазмы - это опушенные или окаймленные пузырьки такого же типа, как и окаймленные пузырьки при пиноцитозе. Это - так называемая транс-сеть аппарата Гольджи (TGN), где происходит разделение и сортировка секретируемых продуктов. Еще дистальнее располагается группа более крупных вакуолей - это уже продукт слияния мелких вакуолей и образования секреторных вакуолей.
При изучении толстых срезов клеток в мегавольтный электронный микроскоп было найдено, что в клетках отдельные диктосомы могут быть связаны друг с другом системой вакуолей и цистерн. Так что образуется рыхлая трехмерная сеть, выявляемая в световом микроскопе. В случае диффузной формы АГ каждый отдельный его участок представлен диктиосомой. У клеток растений преобладает диффузный тип организации АГ, обычно в среднем на клетку приходится около 20 диктиосом. В клетках животных часто с зоной мембран аппарата Гольджи ассоциированы центриоли; между радиально отходящих от них пучков микротрубочек лежат группы стопок мембран и вакуолей, которые концентрически окружают клеточный центр. Эта связь, вероятно, отражает участие микротрубочек в движении вакуолей.
Секреторная функция аппарата Гольджи
Мембранные элементы АГ участвуют в сегрегации и накоплении продуктов, синтезированных в ЭР, участвуют в их химических перестройках, созревании: это, главным образом перестройка олигосахаридных компонентов гликопротеинов в составе водорастворимых секретов или в составе мембран (рис. 180).
В цистернах АГ происходит синтез полисахаридов, их взаимосвязь с белками, приводящая к образованию мукопротеидов. Но главное, с помощью элементов аппарата Гольджи происходит процесс выведения готовых секретов за пределы клетки. Кроме того, АГ является источником клеточных лизосом.
Участие АГ в процессах выведения секреторных продуктов было очень хорошо изучено на примере экзокринных клеток поджелудочной железы. Для этих клеток характерно наличие большого числа секреторных гранул (зимогеновых гранул), которые представляют собой мембранные пузырьки, заполненные белковым содержимым. В составе белков зимогеновых гранул входят разнообразные ферменты: протеазы, липазы, карбогидразы, нуклеазы. При секреции содержимое этих зимогеновых гранул выбрасывается из клеток в просвет железы, а затем перетекает в полость кишечника. Так как основным продуктом, выводимым клетками поджелудочной железы, является белок, то исследовали последовательность включения радиоактивных аминокислот в различные участки клетки (рис. 181). Для этого животным вводили меченную тритием аминокислоту (3 Н-лейцин) и с помощью электронно-микроскопической радиоавтографии следили во времени за локализацией метки. Оказалось, что через короткий промежуток времени (3-5 мин) метка локализовалась только в базальных участках клеток, в участка, богатых гранулярным ЭР. Так как метка включалась в белковую цепь во время синтеза белка, то было ясно, что ни в зоне АГ, ни в самих зимогеновых гранулах синтез белка не происходит, а он синтезируется исключительно в эргастоплазме на рибосомах. Несколько позднее (через 20-40 мин) метка кроме эргастоплазмы была обнаружена в зоне вакуолей АГ. Следовательно, после синтеза в эргастоплазме белок был транспортирован в зону АГ. Еще позднее (через 60 мин) метка обнаруживалась уже и в зоне зимогеновых гранул. В дальнейшем метку можно было видеть в просвете ацинусов этой железы. Таким образом, стало ясно, что АГ является промежуточным звеном между собственно синтезом секретируемого белка и выведением его из клетки. Также подробно процессы синтеза и выведения белков были изучены на других клетках (молочная железа, бокаловидные клетки кишечника, щитовидная железа и др.), и были исследованы морфологические особенности этого процесса. Синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭР, по которым он транспортируется к зоне мембран АГ. Здесь от гладких участков ЭР отщепляются мелкие вакуоли, содержащие синтезированный белок, которые поступают в зону вакуолей в проксимальной части диктиосомы. В этом месте вакуоли могут сливаться друг с другом и с плоскими цис-цистернами диктиосомы. Таким образом происходит перенесение белкового продукта уже внутри полостей цистерн АГ.
По мере модификации белков в цистернах аппарата Гольджи, они с помощью мелких вакуолей переносятся от цистерн к цистерне в дистальную часть диктиосомы, пока не достигают трубчатой мембранной сети в транс-участке диктиосомы. В этом участке происходит отщепление мелких пузырьков, содержащих уже зрелый продукт. Цитоплазматическая поверхность таких пузырьков бывает сходна с поверхностью окаймленных пузырьков, которые наблюдаются при рецепторном пиноцитозе. Отделившиеся мелкие пузырьки сливаются друг с другом, образуя секреторные вакуоли. После этого секреторные вакуоли начинают двигаться к поверхности клетки, соприкасаются с плазматической мембраной, с которой сливаются их мембраны, и, таким образом, содержимое этих вакуолей оказывается за пределами клетки. Морфологически этот процесс экструзии (выбрасывания) напоминает пиноцитоз, только с обратной последовательностью стадий. Он носит название экзоцитоз .
Такое описание событий является только общей схемой участия аппарата Гольджи в секреторных процессах. дело усложняется тем, что одна и та же клетка может участвовать в синтезе многих выделяемых белков, может их друг от друга изолировать и направлять к клеточной поверхности или же в состав лизосом. В аппарате Гольджи происходит не просто “перекачка” продуктов из одной полости в другую, но и постепенно идет их “созревание”, модификация белков, которая заканчивается “сортировкой” продуктов, направляющихся или к лизосомам, или к плазматической мембране, или к секреторным вакуолям.
Модификация белков в аппарате Гольджи
В цис-зону аппарата Гольджи синтезированные в ЭР белки попадают после первичного гликозилирования и редукции там же нескольких сахаридных остатков. В конечном итоге все белки там имеют одинаковые олигосахаридные цепи, состоящие из двух молекул N-ацетилглюкозамина, шести молекул маннозы (рис. 182). В цис-цистернах начинается вторичная модификация олигосахаридных цепей и их сортировка на два класса. В результате олигосахариды на гидролитических ферментах, предназначенных для лизосом (богатые маннозой олгосахариды), фосфорилируются, а олигосахариды других белков, направляемых в секреторные гранулы, или к плазматической мембране, подвергаются сложным превращениям, теряя ряд сахаров и присоединяя галактозу, N-ацетилглюкозамин и сиаловые кислоты.
При этом возникает специальный комплекс олигосахаридов. Такие превращения олигосахаридов осуществляются с помощью ферментов - гликозилтрансфераз, входящих в состав мембран цистерн аппарата Гольджи. Так как каждая зона в диктиосомах имеет свой набор ферментов гликозилирования, то гликопротеиды как бы по эстафете переносятся из одного мембранного отсека (“этажа” в стопке цистерн диктиосомы) в другой и в каждом подвергаются специфическому воздействию ферментов. Так в цис-участке происходит фосфорилирование манноз в лизосомных ферментах и образуется особая маннозо-6-группировка, характерная для всех гидролитических ферментов, которые потом попадут в лизосомы.
В средней части диктиосом протекает вторичное гликозилирование секреторных белков: дополнительное удаление маннозы и присоединение N-ацетилглюкозамина. В транс-участке к олигосахаридной цепи присоединяются галактоза и сиаловые кислоты (рис. 183).
Эти данные были получены совершенно разными методами. С помощью дифференциального центрифугирования удалось получить раздельные более тяжелые (цис-) компоненты аппарата Гольджи и более легкие (транс-) и определить в них наличие гликозидаз и их продуктов. С другой стороны, используя моноклональные антитела к различным ферментам с помощью электронной микроскопии удалось их локализовать прямо на срезах клеток.
В ряде специализированных клеток в аппарате Гольджи происходит синтез собственно полисахаридов.
В аппарате Гольджи растительных клеток происходит синтез полисахаридов матрикса клеточной стенки (гемицеллюлозы, пектины). Кроме того, диктиосомы растительных клеток участвуют в синтезе и выделении слизей и муцинов, в состав которых входят также полисахариды. Синтез же основного каркасного полисахарида растительных клеточных стенок, целлюлозы, происходит как уже говорилось, на поверхности плазматической мембраны.
В аппарате Гольджи клеток животных происходит синтез длинных неразветвленных полисахаридных цепей глюкозаиногликанов. Один из них, гиалуроновая кислота, входящая в состав внеклеточного матрикса соединительной ткани, содержит несколько тысяч повторяющихся дисахаридных блоков. Многие глюкозаиногликаны ковалентно связаны с белками и образуют протеогликаны (мукопротеины). Такие полисахаридные цепи модифицируются в аппарате Гольджи и связываются с белками, которые в виде протеогликанов секретируются клетками. В аппарате Гольджи происходит также сульфатирование глюкозаиногликанов и некоторых белков.
Сортировка белков в аппарате Гольджи
Итак, через аппарат Гольджи проходит по крайней мере, три потока синтезированных клеткой нецитозольных белков: поток гидролитических ферментов в компартмент лизосом, поток выделяемых белков, которые накапливаются в секреторных вакуолях, и выделяются из клетки только по получении специальных сигналов, поток постоянно выделяемых секреторных белков. Следовательно, должен быть какой-то специальный механизм пространственного разделения этих разных белков и их путей следования.
В цис- и средних зонах диктиосом все эти белки идут вместе без разделения, они только раздельно модифицируются в зависимости от их олигосахаридных маркеров.
Собственно разделение белков, их сортировка, происходит в транс-участке аппарата Гольджи. Этот процесс не до конца расшифрован, но на примере сортировки лизосомных ферментов можно понять принцип отбора определенных белковых молекул (рис. 184).
Известно, что только белки-предшественники лизосомных гидролаз имеют специфическую олигосахаридную, а именно маннозную группу. В цис-цистернах эти группировки фосфорилируются и дальше вместе с другими белками переносятся от цистерны к цистерне, через среднюю зону в транс-участок. Мембраны транс-сети аппарата Гольджи содержат трансмембранный белок - рецептор (манноза-6-фосфатный рецептор или М-6-Ф-рецептор), который узнает фосфорилированные маннозные группировки олигосахаридной цепи лизосомных ферментов и связывается с ними. Это связывание происходит при нейтральных значениях рН внутри цистерн транс-сети. На мембранах эти М-6-Ф-рецепторные белки образуют кластеры, группы, которые концентрируются в зонах образования мелких пузырьков, покрытых клатрином. В транс-сети аппарата Гольджи происходит их отделение, отпочковывание и дальнейший перенос к эндосомам. Следовательно М-6-Ф-рецепторы, являясь трансмембранными белками, связываясь с лизосомными гидролазами, отделяют их, отсортировывают, от других белков (например, секреторных, нелизосомных) и концентрируют их в окаймленных пузырьках. Оторвавшись от транс-сети эти пузырьки быстро теряют шубу, сливаются с эндосомами, перенося свои лизосомные ферменты, связанные с мембранными рецепторами, в эту вакуоль. Как уже говорилось, внутри эндосом из-за активности протонного переносчика происходит закисление среды. Начиная с рН 6 лизосомные ферменты диссоциируют от М-6-Ф-рецепторов, активируются и начинают работать в полости эндолизосомы. Участки же мембран вместе с М-6-Ф-рецепторами возвращаются путем рециклизации мембранных пузырьков обратно в транс-сеть аппарата Гольджи.
Вероятнее всего, что та часть белков, которая накапливается в секреторных вакуолях и выводится из клетки после поступления сигнала (например нервного или гормонального) проходит такую же процедуру отбора, сортировки на рецепторах транс-цистерн аппарата Гольджи. Эти секреторные белки попадают сначала в мелкие вакуоли тоже одетые клатрином, которые затем сливаются друг с другом. В секреторных вакуолях часто происходит агрегация накопленных белков в виде плотных секреторных гранул. Это приводит к повышению концентрации белка в этих вакуолях примерно в 200 раз, по сравнению с его концентрацией в аппарате Гольджи. Затем эти белки по мере накопления в секреторных вакуолях выбрасываются из клетки путем экзоцитоза, поле получения клеткой соответствующего сигнала.
От аппарата Гольджи исходит и третий поток вакуолей, связанный с постоянной, конститутивной секрецией. Так фибробласты выделяют большое количество гликопротеидов и муцинов, входящих в основное вещество соединительной ткани. Многие клетки постоянно выделяют белки, способствующие связыванию их с субстратами, постоянно идет поток мембранных пузырьков к поверхности клетки, несущие элементы гликокаликса и мембранных гликопротеидов. Этот поток выделяемых клеткой компонентов не подлежит сортировке в рецепторной транс-системе аппарата Гольджи. Первичные вакуоли этого потока также отщепляются от мембран и относятся по своей структуре к окаймленным вакуолям, содержащим клатрин (рис. 185).
Заканчивая рассмотрение строения и работы такой сложной мембранной органеллы, как аппарат Гольджи, необходимо подчеркнуть, что несмотря на кажущуюся морфологическую однородность его компонентов, вакуоли и цистерны, на самом деле, это не просто скопище пузырьков, а стройная, динамичная сложно организованная, поляризованная система.
В АГ происходит не только транспорт везикул от ЕР к плазматической мембране. Существует ретроградный перенос везикул. Так от вторичных лизосом отщепляются вакуоли и возвращаются вместе с рецепторными белками в транс-АГ зону. Кроме того существует поток вакуолей от транс-зоны к цис-зоне АГ, а так же от цис-зоны к эндоплазматическому ретикулуму. В этих случаях вакуоли одеты белками COP I-комплекса. Считается, что таким путем возвращаются различные ферменты вторичного гликозилирования и рецепторные белки в составе мембран.
Эти особенности поведения транспортных везикул дали основу гипотезе о существовании двух типов транспорта компонентов АГ (рис. 186).
По одному из них, наиболее старому, в АГ существуют стабильные мембранные компоненты, к которым от ЭР эстафетно переносятся вещества с помощью транспортных вакуолей. По альтернативной модели АГ является динамическим производным ЭР: отщепившиеся от ЭР мембранные вакуоли сливаются друг с другом в новую цис-цистерну, которая затем продвигается через всю зону АГ и в конце распадается на транспортные везикулы. По этой модели ретроградные COP I везикулы возвращают постоянные белки АГ в более молодые цистерны. Таким образом предполагается, что переходная зона ЭР представляет собой “родильный дом” для аппарата Гольджи.
А- Гранулярной цитоплазматической сети.
Б- Микропузырьков.
В- Микрофиламентов.
Г- Цистерн.
Д- Вакуолей.
Ответ: Б,Г,Д.
16. Указать, какие функции выполняет комплекс Гольджи:
А- Синтез белка.
Б- Образование комплексных химических соединений (гликопротеидов, липопротеидов).
В- Образование первичных лизосом.
Г- Участие в выведении из клетки секреторного продукта.
Д- Образование гиалоплазмы.
Ответ: Б,В,Г.
Какие структурные элементы клетки наиболее активно участвуют в экзоцитозе?
А- Цитолемма.
Б- Цитоскелет.
В- Митохондрии.
Г- Рибосомы.
Ответ: А,Б.
18 . Что определяет специфичность синтезируемого белка?
А- Информационная РНК.
Б- Рибосомная РНК.
Г- Мембраны цитоплазматической сети.
Ответ: А,В
19 . Какие структурные элементы активно участвуют в выполнении
Фагоцитарной функции?
А- Кариолемма.
Б- Эндоплазматическая сеть.
В- Цитолемма.
Г- Лизосомы.
Д- Микрофиламенты.
Ответ: В,Г,Д.
20 .Какие структурные компоненты клетки обусловливают базофилию цитоплазмы?
А- Рибосомы.
Б- Агранулярная эндоплазматическая сеть.
В- Лизосомы.
Г- Пероксисомы.
Д- Комплекс Гольджи.
Е- Гранулярная эндоплазматическая сеть.
Ответ: А,Е.
21 . Какие из перечисленных органелл имеют мембранное строение?
А- Клеточный центр.
Б- Митохондрии.
В- Комплекс Гольджи.
Г- Рибосомы.
Д- Цитоскелет.
Ответ: Б,В.
22 .Что общего между митохондриями и пероксисомами?
А- Относятся к органоидам мембранного строения.
Б- Имеют двойную мембрану.
Д- Это органеллы общего значения.
Ответ: А,В,Д.
Какие функции в клетке выполняют лизосомы?
А- Биосинтез белка
Б- Участие в фагоцитозе
В- Окислительное фосфорилирование
Г- Внутриклеточное пищеварение
Ответ: Б.Г.
Какова структурная организация лизосом?
А- Окружены мембраной.
В- Заполнены гидролитическими ферментами.
Г- Образуются в комплексе Гольджи.
Ответ: А,В,Г.
25. Гликокаликс:
А- Находится в гладкой эндоплазматической сети.
Б- Находится на наружной поверхности цитолеммы.
В- Образован углеводами.
Г- Участвует в клеточной адгезии и клеточном узнавании.
Д- Находится на внутренней поверхности цитолеммы.
Ответ: Б,В,Г.
26. Маркерные ферменты лизосом:
А- Кислая фосфатаза.
Б- АТФ-аза.
В- Гидролазы.
Г- Каталаза и оксидазы.
Ответ: А,В.
Каково значение ядра в жизнедеятельности клетки?
А- Хранение наследственной информации.
Б- Центр накопления энергии.
В- Центр управления внутриклеточным метаболизмом.
Г- Место образование лизосом.
Д- Воспроизведение и передача генетической информации дочерним клеткам.
Ответ: А,В,Д.
28. Что не относится к структурным компонентам ядра:
А- Кариолемма.
Б- Ядрышки.
В- Кариоплазма.
Г- Рибосомы.
Д- Хроматин, хромосомы.
Е- Пероксисомы.
Ответ: Г,Е.
Что транспортируется из ядра через ядерные поры в цитоплазму?
А- Фрагменты ДНК.
Б- Субъединицы рибосом.
В- Информационные РНК.
Г- Фрагменты эндоплазматической сети.
Ответ: Б,В.
Что такое ядерно-цитоплазматическое отношение и как оно меняется при повышении функциональной активности клетки?
А- Положение ядра в цитоплазме.
Б- Форма ядра.
В- Отношение размера ядра к размеру цитоплазмы.
Г- Снижено при повышенной функциональной активности клетки.
Ответ: В,Г.
Что верно для ядрышек?
А- Хорошо видны во время митоза.
Б- Состоят из гранулярного и фибриллярного компонентов.
В- Гранулы ядрышка - субъединицы рибосом.
Г- Нити ядрышка - рибонуклеопротеиды
Ответ: Б,В,Г.
Какие перечисленные признаки относятся к некрозу?
А- Это генетически запрограммированная гибель клетки
Б- В начале апоптоза синтез РНК и белка возрастает.
В- разрушаются мембраны
Г- ферменты лизосом выходят в цитоплазму
Д- Фрагментация цитоплазмы с образованием апоптических тел
Ответ: В,Г.
Верно все, кроме
1.Функция комплекса Гольджи (верно все, кроме):
А- сортировка белков по транспортным пузырькам
Б- гликозилирование белков
В- реутилизация мембран секреторных гранул после экзоцитоза
Г- упаковка секреторного продукта
Д- синтез стероидных гормонов
2. Микротрубочки обеспечивают (верно все, кроме):
А- организацию внутреннего пространства клетки
Б- поддержание формы клетки
В- поляризацию клетки при делении
Г- формируют сократительный аппарат
Д- организацию цитоскелета
Е- транспорт органелл
3. К специализированным структурам, построенным на основе цитоскелета, относятся (верно все, кроме):
А- реснички, жгутики
Б- базальная исчерченность
В- микроворсинки
4. Локализация ресничек (верно все, кроме):
А- эпителий слизистой оболочки воздухоносных путей
Б- эпителий проксимального отдела нефрона
В- эпителий слизистой репродуктивного тракта женщин
Г- эпителий слизистой семявыносящих путей
5. Локализация микроворсинок (верно все, кроме):
А- эпителий слизистой оболочки тонкой кишки
Б- эпителий слизистой оболочки трахеи
В- эпителий проксимального отдела нефрона
6. Базальная исчерченность (верно все, кроме):
А- обеспечивает транспорт веществ против градиента концентрации
Б- участок клетки, где идут высоко энергоемкие процессы
В- участок клетки, где происходит простая диффузия ионов
Г- где происходит реабсорбция элементов первичной мочи в проксимальном канальце нефрона
Д- участвует в концентрации слюнного секрета
7. Щеточная каемка (верно все, кроме):
А- располагается на апикальной поверхности клеток
Б- увеличивает площадь всасывательной поверхности
В- состоит из ресничек
Г- состоит из микроворсинок
Д- увеличивает транспортную поверхность в проксимальных канальцах нефрона
8. Органоиды общего назначения (верно все, кроме):
А- митохондрии
В- комплекс Гольджи
Г- реснички
Д- лизосомы
Е- пероксисомы
Ж- центриоли
З- элементы цитоскелета
9.Функция пероксисом (верно все, кроме):
А- окисление органического субстрата с образованием перекиси водорода
Б- синтез фермента – каталазы
В- утилизация перекиси водорода
10. Рибосомы (верно все, кроме):
А- при световой микроскопии об их наличии судят по выраженной базофилии цитоплазмы
Б- состоят из малой и большой субъединиц
В- образуются в гранулярной ЭПС
Г- состоят из рРНК и белков
Д- немембранного строения
11.Какие органоиды хорошо развиты в стероидпродуцирующих клетках (верно все, кроме):
А- гранулярная эндоплазматическая сеть
Б- агранулярная эндоплазматическая сеть
В- митохондрии с трубчатыми кристами
12.Трофические включения (верно все, кроме):
А- углеводные
Б- слизистые
В- белковые
Г- липидные
13.Ядерная оболочка (верно все, кроме):
А- состоит из одинарной мембраны
Б- состоит из двух мембран
В- снаружи на ней расположены рибосомы
Г- изнутри с ней связана ядерная пластинка
Д- пронизана порами
14. Структурные компоненты ядра (верно все, кроме):
А- нуклеоплазма
Б- нуклеолемма
В- микротрубочки
Г- хроматин
Д- ядрышки
15. Строение ядерной поры (верно все, кроме):
А- мембранный компонент
Б- хромосомный компонент
В- фибриллярный компонент
Г- гранулярный компонент
16. Ядрышко (верно все, кроме):
А- окружено мембраной
Б- не окружено мембраной
В- в его организации участвуют пять пар хромосом
Г- содержит гранулярный и фибриллярный компонент
17. Ядрышко (верно все, кроме):
А- количество зависит от метаболической активности клетки
Б- участвует в образовании субъединиц рибосом
В- в организации участвуют 13,14, 15, 21 и 22 хромосомы
Г- в организации участвуют 7, 8, 10, 11 и 23 хромосомы
Д- состоит из трех компонентов
18. Клеточный центр (верно все, кроме):
А- локализуется вблизи ядра
Б- является центром организации веретена деления
В- состоит из двух центриолей
Г- центриоли образованы 9 дуплетами микротрубочек
Д- центриоли дуплицируются в S периоде интерфазы
19. Митохондрии (верно все, кроме):
А- наличие крист
Б- способность делиться
20. Функции актиновых филаментов (верно все, кроме):
А- движение клетки
Б- изменение формы клетки
В- участие в экзо- и эндоцитозе
Г- обеспечивают движение ресничек
Д- входят в состав микроворсинок
21. Для ядрышка верно все, кроме:
А- Образуются в области ядрышковых организаторов (вторичных перетяжек хромосом)
Б- Гранулы ядрышек выходят в цитоплазму
В- Белки ядрышек синтезируются в цитоплазме
Г- Ядрышковая РНК образуется в цитоплазме
На соответствие
1. Сопоставьтепериоды интерфазы с процессами, происходящими в них:
1. Пресинтетический А- удвоение ДНК, увеличение синтеза РНК
2. Синтетический Б- синтез рРНК, иРНК, тубулинов
3. Постсинтетический В- рост клеток, подготовка их к синтезу ДНК
Ответ: 1-В; 2-А; 3-Б.
2 .Сопоставьтефазы митоза с процессами, происходящими в них:
1. Профаза А- образование экваториальной пластинки из хромосом
2. Метафаза Б- образование нуклеолеммы, деспирализация хромосом,
образование ядрышка, цитотомия
3. Анафаза В-спирализация хромосом, исчезновение ядрышка,
фрагментация нуклеолеммы
4. Телофаза Г- расхождение хроматид к противоположным полюсам
Ответ: 1-В; 2-А; 3-Г; 4-Б.
3. Изменение структуры ядра называется (сопоставьте):
1.кариолизис А- уменьшение размеров и уплотнение хроматина
2.кариорексис Б- фрагментация
3.кариопикноз В- растворение его компонентов
Ответ: 1-В, 2-Б, 3-А.
4. Характеристика компонентов препарата:
1.хромофобный А- окрашивается красителем суданом
2.хромофильный Б- не окрашивается красителем
3.суданофильный В- окрашивается красителем
 Корпускулярные и волновые свойства частиц
Корпускулярные и волновые свойства частиц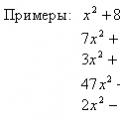 Уравнения онлайн X 5 0 решение
Уравнения онлайн X 5 0 решение Предельные монокарбоновые кислоты
Предельные монокарбоновые кислоты