Lämmastiku metabolismi lõpp-produktid. Kusihape veres: normid ja kõrvalekalded, miks see suureneb, dieet selle vähendamiseks Lämmastiku ainevahetuse lõpp-produkt on
Riiklik õppeasutus
Chita osariigi meditsiiniakadeemia
L.P. Nikitina, A.Ts. Gomboeva, N.S. Kuznetsova
Biokeemia lämmastiku metabolism normaalsetes ja patoloogilistes tingimustes
Toimetanud prof. B.S. Khyshyktueva
L.P. Nikitina
A.Ts. Gomboeva
N.S. Kuznetsova
See juhend on mõeldud meditsiiniülikoolide üliõpilastele. See esitab üsna ülevaatlikus ja arusaadavas keeles teavet erinevate lämmastikku sisaldavate ühendite kohta, eelkõige aminohapete, nukleotiidide ja nende biopolümeeride – valkude, nukleiinhapete kohta.
|
Lühendite loetelu. . . . . . . . . . . . . . . . . . . . . . . | |||
|
. . . . . . . . . . . . . . . . . . . . . . . | |||
|
Sissejuhatus. | |||
|
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . | |||
|
. . . . . . . . . . . . . . . . . . . . . . | |||
|
1. peatükk. Lämmastikku sisaldavate ühendite rollide klassifikatsioon ja üldistus. . | |||
|
Peatükk 2. Aminohapete ainevahetus. . . . . . . . . . | |||
|
. . . . . . . . . . . . . . . . . . . . . . . . | |||
|
2.1. | |||
|
Polüpeptiidide katabolismi hüdrolüütiline staadium. . . . . . . . . . | |||
|
2.2. | |||
|
Aminohapete saatus rakus. . . . . . . | |||
|
. . . . . . . . . . . . . . . . . . . . . . . | |||
|
2.2.1. | |||
|
Mittespetsiifiliste teisenduste valikud. . . . . . . . . . . . . . . | |||
|
2.2.1.1. | |||
|
Dekarboksüülimisreaktsioonid. . . . . . | |||
|
. . . . . . . . . . . . . . . . . . . | |||
|
3.2.2. | |||
|
Dinukleotiidide tähendus. . . . . . . . . . . | |||
|
. . . . . . . . . . . . . . . . . . . . | |||
|
3.2.3. | |||
|
Polünukleotiidid. . . . . . . . . . . . . . . . . . . | |||
|
. . . . . . . . . . . . . . . . . . . | |||
|
3.2.3.1. | |||
|
RNA tüübid. . . . . . . . . . . . . . . . . . . . . . . . . | |||
|
. . . . . . . . . . . . . . . . | |||
|
3.2.3.2. | |||
|
DNA variandid. . . . . . . . . . . . . . . . . . . . . | |||
|
. . . . . . . . . . . . . . . . | |||
|
3.2.3.3. | |||
|
Komplekssete nukleotiidide füüsikalis-keemilised ja bioloogilised omadused | |||
|
3.3. | |||
|
Nukleiinstruktuuride vahetuse kataboolne faas. . . . . . . . . . | |||
|
3.3.1. | |||
|
Nukleoproteiinide lagunemine seedetraktis ja kudedes. . . . . . . . . . . . . . . . . | |||
|
3.3.2. | |||
|
Nukleosiidide transformatsioonide spetsiifilised rajad. . . . . . . . . | |||
|
3.3.2.1. | |||
|
Puriinide ainevahetuse lõpp-produkt on kusihape. . . | |||
|
3.3.2.2. | |||
|
Pürimidiinitsüklite hävitamise skeem. . . . . . . . . . . . . . | |||
|
3.4. | |||
|
Mononukleotiidide sünteesi teed. . . . . . | |||
|
. . . . . . . . . . . . . . . . . . . . . . | |||
|
3.4.1. | |||
|
Geneetikaterminite sõnastik. . . . . . . . . . . . | |||
. . . . . . . . . . . . . . . . . . . . . . . .
Lühendite loetelu
ADP - adenosiintrifosfaat
ALT – alaniinaminotransferaas
AMP – adenosiinmonofosfaat
AO – antioksüdant
ARZ – radikaalivastane kaitse
AST – aspartaataminotransferaas
ATP - adenosiintrifosfaat
HFA – kõrgem rasvhape
GABA – gamma-aminovõihape
HGPRT – hüpoksantiinguaniinfosforibosüültransferaas
SKT – guanosiindifosfaat
GMP – guanosiinmonofosfaat
GNG – glükoneogenees
GP – glütseroolfosfatiid(id)
HSE – hormoonitundlik element
hRNA – heterogeenne tuuma ribonukleiinhape
DHAP – dihüdroksüatsetoonfosfaat
DOPA – di(hüdr)oksüfenüülalaniin
Seedetrakt - seedetrakt
IMP – inosiinmonofosfaat
iRNA – sõnumitooja ribonukleiinhape
CoA – atsüülimise koensüüm
NAD + –
NAD + P –fosfaat
NTP – nukleosiidtrifosfaat
OA – oksaloatsetaat
OMP – oritidiinmonofosfaat
PVA – püroviinamarihape
PPP – pentoosfosfaadi rada
RNDP – ribonukleosiiddifosfaat
RMNP – ribonukleosiidmonofosfaat
RNA – ribonukleiinhape
rRNA – ribosomaalne ribonukleiinhape
RNTP – ribonukleosiidtrifosfaat
STH – somatotroopne hormoon
THFA – tetrahüdrofoolhape
TDP – tiamiindifosfaat
TMP – tümidiinmonofosfaat
tRNA – transpordi ribonukleiinhape
UMP – uridiinmonofosfaat
UTP – uridiintrifosfaat
FAD – flaviinadeniini dinukleotiid
FAPS – fosfoadenosiinfosfosulfaat
FMN – flaviini mononukleotiid
PRPP – fosforibosüülpürofosfaat
c-AMP – tsükliline adenosiinmonofosfaat
CDP – tsütidiindifosfaat
CMP – tsütidiinmonofosfaat
TCA tsükkel – trikarboksüülhappe tsükkel
ETC – elektronide transpordiahel
H – histoon
SAM – S-adenosüülmetioniin
Sissejuhatus
Lämmastikuaatomite kaasamine, mis on võimelised prootoneid vastu võtma tänu üksikule elektronpaarile, s.o. omavad aluse omadusi, viib täidetavate funktsioonide kvalitatiivse muutuseni. Keha ei ole võimeline kasutama aminohappeid sisaldavaid molekule energiaallikatena, need teenivad muid eesmärke.
Täiskasvanud kahepaiksete eritussüsteem on esindatud paari neerupaariga - mesonephros, mis asuvad lülisamba sakraalse osa külgedel, kuid erinevalt kaladest ei ole neil linditaolist kuju, vaid need on ovaalsed ja väga kompaktsed. Kusejuht on Wolffi kanal (meestel toimib see ka vas deferensina), mis suubub kloaaki. Kõrgematel maapealsetel vormidel avaneb ulatuslik põis kloaaki, kuhu uriin siseneb kloaagist ja hoitakse ajutiselt.
Kui põis üle ajab, tühjendab see oma sisu samasse kloaaki ja sealt väljub uriin. Kahepaiksete neerud eemaldavad verest jääkaineid ja säilitavad vee-soola tasakaalu (tasakaalu). Nefronite arv neerus sõltub sellest, kui tihedalt on loom veega seotud. Valdavalt vees elavatel kahepaiksetel on mõlemas neerus umbes 400–500 nefronit ja sabata kahepaiksetel umbes 2000. Seda seletatakse asjaoluga, et vee-elustikud kahepaiksed eritavad osa oma ainevahetusproduktidest lõpuste kaudu
ja kehakatted ümbritsevasse vette.
Kahepaiksete lämmastiku metabolismi lõppsaadus on uurea.
Liigne vesi eemaldatakse neerude kaudu, mis siseneb looma kehasse läbi naha, soolad aga reabsorbeeritakse (reabsorbeeritakse) uriinist, nii et suurem osa ioonidest - kuni 99% - naaseb verre.
Paljude munandist välja ulatuvate õhukeste seemnetorukeste kaudu läbivad sigimisproduktid neeru eesmise osa ja sisenevad Wolffi kanalisse, mis kahepaiksetel (nagu ka kõhrelistel kaladel) ühendab kusejuha ja vasdeferensi funktsioonid. Wolffi kanalid tühjenevad kloaaki, kuid veidi enne neist moodustavad nad väikese pikenduse - seemnepõiekese, millesse sperma ajutiselt koguneb. Nagu munandid ja rasvakehad, kahanevad ka seemnepõiekesed väljaspool sigimisperioodi.
Kahepaiksetel puuduvad isaste sigimissüsteemis oma sigimiskanalid, samuti puuduvad enamikul liikidel kopulatsiooniorganid.
Naiste reproduktiivsüsteemi moodustavad kaks soolestiku küljes rippuvat munasarja, mille kohal asuvad rasvkehad (375). Munasarjade suurus varieerub oluliselt olenevalt aastaajast, suureneb oluliselt pesitsushooajal. Kevadel on munasarjad eriti suured, läbi õhukese seina on näha suured munakollaserikkad munad.
Küpsed munad väljuvad munasarjast follikulaarse membraani purunemise kaudu ja satuvad kehaõõnde, kust nad seejärel munajuha lehtrisse sisenevad. Emastel kahepaiksetel on munajuha paaris Mülleri kanal, mis avaneb ühest otsast (lehter) kehaõõnde ja teisest küljest kloaaki. Pesitsusajal pikenevad munajuhad tugevasti ja nende seinad paksenevad.
Paljudele kahepaiksetele on iseloomulik paaritumiskäitumine, millega sageli kaasnevad häälesignaalid (mõnede konnade isased võivad teha äärmiselt valju häält). See on vajalik seksuaalpartnerite reproduktiivproduktide samaaegse vabanemise stimuleerimiseks. Väetamine võib olla sisemine või välimine.
Kahepaiksetele on iseloomulik areng koos moondega, kus munast väljub vastne, mis on oma struktuurilt palju lähedasem kaladele kui täiskasvanud kahepaiksetele. Sellel on iseloomulik kalataoline kuju, nii et see liigub keha pikisuunalisi painutusi kasutades. Hingamisorganid toimivad esmalt väliste lõpustena, mis on naha väljakasvud, hiljem tungivad läbi lõpused, avades sisemised lõpused ja seejärel vähenevad välised lõpused. Algstaadiumis jäsemed puuduvad. Sabaga kahepaiksetel toimivad välislõpused kogu vastsete perioodi, kuid sisemised ei arene.
Kahepaiksete vastsete arengu käigus rekonstrueeritakse tema sisesüsteemid: hingamis-, vereringe-, eritus- ja seedesüsteemid..
Järk-järgult arenevad jäsemed. Metamorfoos lõpeb täiskasvanud isendi miniatuurse koopia moodustumisega sabata loomadel, saba väheneb.
Ambystosid iseloomustab neoteenia, st nad paljunevad vastsetega, mida peeti pikka aega ekslikult iseseisvaks liigiks, mistõttu on neil oma nimi - aksolotl. Selline vastne on suurem kui täiskasvanud. Veel üks huvitav kahepaiksete rühm on proteasid, kes elavad püsivalt vees ja säilitavad kogu elu välised lõpused, st säilitavad vastsele omased omadused.
Räägime puriini aluste metabolismi iseärasustest. Enamiku inimeste jaoks ei tähenda see midagi. Kuid kui olete tuttav sõnadega "podagra", urolitiaas, insuliiniresistentsus, II tüüpi diabeet, siis peate teadma puriinide metabolismi olemust. Näib: mis on operatsioonil sellega pistmist? Ja hoolimata asjaolust, et paljud spetsialistid diagnoosivad "podagra" liigesevalu ja kõrge kusihappesisaldusega. Tegelikkuses on kõik palju keerulisem. Näiteks võib normaalse kusihappesisalduse korral tekkida podagra artriit ja vastupidi: kõrge kusihappesisaldus võib mõnel juhul tekkida tervel inimesel.
Inimkeha koosneb peamiselt neljast keemilisest elemendist, mis moodustavad 89% koostisest: C-süsinik (50%), O-hapnik (20%), H-vesinik (10%) ja N-lämmastik (8,5%). ). Edasi tulevad terve rida makroelemente: kaltsium, fosfor, kaalium, väävel, naatrium, kloor jne.. Siis mikroelemendid, mille kogus on küll väga väike, aga elutähtsad: mangaan, raud, jood jne.
Elusorganism on dünaamiline süsteem. Lihtsamalt öeldes: ained sisenevad sinna pidevalt (muutudes keha osaks) ja eemaldatakse sealt. Organismi peamine lämmastikuallikas on valgud. Seedetraktis toiduga kaasas olev valk laguneb aminohapeteks, mis seejärel kaasatakse ainevahetusse. Noh, kuidas lämmastikku sisaldavad ained organismist eemaldatakse?
Loomadel on evolutsiooni käigus välja kujunenud teatud lämmastiku metabolismi tunnused.
Lisaks on nende tunnuste määramisel võtmetähtsusega: elamistingimused ja juurdepääs veele.
Loomad jagunevad kolme rühma, millel on erinevused lämmastiku metabolismis:
Ammoniolüütiline. Lämmastiku metabolismi lõpp-produkt on ammoniaak NH3. See hõlmab enamikku veeselgrootuid ja kalu.
Asi on selles, et ammoniaak on mürgine aine. Ja selle eemaldamiseks on vaja palju, palju vedelikku. Õnneks on see vees hästi lahustuv. Evolutsiooni käigus maale pääsedes tekkis vajadus muuta ainevahetust. Nii nad ilmusid:
Ureolüütiline. Need loomad arendasid välja niinimetatud "uurea tsükli". Ammoniaak ühineb CO2-ga (süsinikdioksiid). Moodustub lõpptoode - uurea. Karbamiid ei ole nii mürgine aine ja selle eemaldamiseks kulub märgatavalt vähem vedelikku. Muide, sina ja mina kuulume sellesse rühma. Kusihapet tekib ka ainevahetuse käigus palju väiksemates kogustes, kuid laguneb vähetoksiliseks ja hästi lahustuvaks allantoiiniks. Aga... Välja arvatud inimesed ja ahvid. See on väga oluline ja me tuleme selle juurde tagasi.
Urikoteel. Urolüütilise ainevahetusega kahepaiksete esivanemad pidid kohanema kuivade piirkondadega. Need on roomajad ja dinosauruste otsesed esivanemad - linnud. Nende lõpp-produkt on kusihape. See lahustub vees väga halvasti ja ei vaja selle kehast eemaldamiseks palju vett. Samade lindude väljaheidetes on kusihappe kogus väga suur. . Samuti halveneb auto värvkate – ole ettevaatlik ja pese see kohe puhtaks.
See on klassikaline kuusnurkne maksasagar. Üldiselt näeb maks mikroskoobi all välja selline. Sarnaselt Moskva linnaga on ainult Kremli asemel kesksoon. Ja meid huvitavad üksteisega tihedalt külgnevad "majad". Need on hepatotsüüdid - maksa võtmerakud.
Slaavi sõna maks tuleneb sõnast "ahi". Tõepoolest, elundi temperatuur on kehatemperatuurist kraadi võrra kõrgem. Selle põhjuseks on väga aktiivne metabolism hepatotsüütides. Rakud on tõeliselt ainulaadsed, neis toimub umbes 2 tuhat keemilist reaktsiooni.
Maks on peamine organ, mis toodab kusihapet. 95% eritunud lämmastikust on kusihappe süntees kui maksas toimuvate keemiliste reaktsioonide lõpp-produkt. Ja ainult 5% on toiduga väljastpoolt tulevate puriini aluste oksüdatsioon. Seetõttu ei ole hüperurikeemia toitumise korrigeerimine ravi võti.
Kusihappe metabolismi diagramm
 Kust tulevad puriinid?
Kust tulevad puriinid?
1. Puriinid, mis pärinevad toidust
. Nagu juba märgitud, on see väike summa - umbes 5%. Need puriinid, mis sisalduvad toidus (enim muidugi maksas ja neerudes, punases lihas).
2. Puriini aluste süntees keha enda poolt
. Suurem osa sellest sünteesitakse maksa hepatotsüütides. Väga oluline punkt, tuleme selle juurde tagasi. Ja ka seda, mis pistmist on fruktoosiga, mida soovitavad diabeetikud ja mille imendumiseks ei ole vaja insuliini.
3. Puriini alused, mis tekivad organismis kudede lagunemise tõttu: onkoloogiliste protsesside käigus, psoriaas
. Miks võib kusihappe tase sportlastel tõusta? See on kolmas viis. Raske füüsiline aktiivsus põhjustab kudede lagunemise ja sünteesi protsesside suurenemist. Kui tegite eelmisel päeval rasket füüsilist tööd ja läksite hommikul testima, võib teie kusihappe tase olla keskmisest kõrgem.  Saame tuttavaks: adeniin ja guaniin. Need on puriini alused. Koos tümiini ja tsütosiiniga moodustavad nad DNA heeliksi. Arstitudengitele ei meeldi biokeemiakursustel tuupimine :). Nagu teate, koosneb DNA kahest ahelast. Adeniinile vastandub alati tümiin ja guaniinile vastandub tsütosiin. Kaks DNA ahelat kleepuvad kokku nagu tõmbluku kaks poolt. Nende ainete hulk suureneb koos aktiivse kudede lagunemisega, nagu juhtub näiteks vähiprotsesside ajal
Saame tuttavaks: adeniin ja guaniin. Need on puriini alused. Koos tümiini ja tsütosiiniga moodustavad nad DNA heeliksi. Arstitudengitele ei meeldi biokeemiakursustel tuupimine :). Nagu teate, koosneb DNA kahest ahelast. Adeniinile vastandub alati tümiin ja guaniinile vastandub tsütosiin. Kaks DNA ahelat kleepuvad kokku nagu tõmbluku kaks poolt. Nende ainete hulk suureneb koos aktiivse kudede lagunemisega, nagu juhtub näiteks vähiprotsesside ajal
Järjestikuste keemiliste reaktsioonide käigus muudetakse puriinid kusihappeks.

Kusihappe metabolism inimestel ja primaatidel
Plaanisin teha diagrammi võimalikult lihtsalt arusaadavaks. Las õpetavad 2. kursuse arstitudengid :). Kuid ta jättis ensüümide nimed. Kõige olulisem punkt on ksantiinoksüdaasi ensüüm
. See on selle aktiivsus, mis ravi ajal väheneb allopurinool(täpsemalt tõhusus, kuna allopurinool konkureerib sellega retseptori pärast), mis vähendab kusihappe sünteesi.
Harva esineb kaasasündinud haigus, millega kaasneb ksantiinoksüdaasi sünteesi geneetiline häire, mille puhul kusihappe tase väheneb. Sel juhul kogunevad ksantiin ja hüpoksantiin. Ksantinuuria. Tundub hea, kusihapet vähem. Selgus aga, et kusihape pole mitte ainult kahjulik, vaid ka kasulik...
Vestlust kusihappe ohtude ja eeliste üle tuleks alustada kaugelt. Siis, 17 miljonit aastat tagasi, miotseeni ajastul, kogesid meie esivanemad mutatsiooni geenis, mis toodab ensüümi urikaasi. Ja me saime puriinide metabolismi "alla lõigatud" versiooni.
 Teistel imetajatel muudab urikaas kusihappe allantoiiniks, mis lahustub ja eritub organismist kergesti. Ja neil loomadel pole kunagi podagra. Võib eeldada, et sellel mutatsioonil pole mingit mõtet. Kuid evolutsioon ei välistanud seda geeni: mutatsioon osutus vajalikuks.
Teistel imetajatel muudab urikaas kusihappe allantoiiniks, mis lahustub ja eritub organismist kergesti. Ja neil loomadel pole kunagi podagra. Võib eeldada, et sellel mutatsioonil pole mingit mõtet. Kuid evolutsioon ei välistanud seda geeni: mutatsioon osutus vajalikuks.
Kaasaegsed uuringud on näidanud, et kusihape on maksas fruktoosi lagunemise kõrvalsaadus ja kusihappe soolade kogunemine aitab kaasa fruktoosi tõhusale muundamisele rasvaks. Seega fikseeriti säästlikkuse geen meie esivanemate genoomis. Siis oli geeni vaja näljaperioodi reservide loomiseks. On tõestatud, et urikaasi lõplik inaktiveerimine langes kokku Maa kliima globaalse jahenemisega. Külmaperioodiks oli vaja võimalikult palju nahaalusest rasvavarust “ära süüa” ja viljades sisalduv fruktoos rasvavarudeks muuta. Praegu viiakse läbi mitmeid katseid ensüümi urikaasi viimisega maksarakkudesse. Võimalik, et tulevikus ilmuvad urikaasi ensüümi baasil ravimid podagra raviks. Seega on meie kalduvus rasvuda meie geenides. Kahjuks neile paljudele meestele ja naistele, kes kannatavad ülekaalulisuse all. Kuid probleem pole ainult geneetikas. Kaasaegse inimese toitumise olemus on muutunud.
Kusihappe kahjude ja eeliste kohta, samuti hüperurikeemia toitumise kohta
On teada, et kusihappe pidev tase võib märkimisväärselt suurendada mitmete haiguste riski. Siiski on tõestatud, et kusihappe taseme perioodiline tõus võib avaldada kasulikku mõju. Ajalooliselt oli juurdepääs lihatoitudele (peamine puriinide allikas) ebaregulaarne. Põhitoit: erinevad juured, puuviljad. Noh, kui primitiivne jahimees toob saaki, siis on see puhkus. Seetõttu oli lihatoodete perioodiline söömine tavaline eluviis. Saak on olemas – sööme isu täis. Saaki pole – sööme taimset toitu. Nüüdseks on kindlaks tehtud, et kusihappe taseme lühiajaline perioodiline tõus avaldab soodsat mõju närvisüsteemi arengule ja talitlusele. Võib-olla sellepärast hakkas aju arenema?
Kuidas see kusihape organismist eritub?
Kaks võimalust: neerud ja maks
Peamine tee on eritumine neerude kaudu - 75%
25 protsenti eritub sapi kaudu maksa kaudu. Kusihape, mis siseneb soolestiku luumenisse, hävib (tänu meie bakteritele soolestikus).
Kusihape siseneb neerudesse naatriumsoola kujul. Atsidoosiga (uriini hapestumine) võivad neeruvaagnasse tekkida mikroliidid. Sama "liiv" ja "kivid". Muide, alkohol vähendab oluliselt uraadi eritumist uriiniga. Sellepärast põhjustab see podagra rünnakut.
Niisiis, milliseid järeldusi tuleks teha kusihappe vähendamise meetodid?
1. Proovige muuta 1-2 päeva nädalas puhtalt taimetoiduks
2. Suurim kogus puriine leidub loomset päritolu kudedes. Veelgi enam, aktiivse ainevahetusega loomarakkudes: maks, neerud - kõige rohkem.
3. Sa peaksid sööma vähem rasvaseid toite, kuna liigne küllastunud rasv pärsib organismi võimet töödelda kusihapet.
4. Söö vähem fruktoosi. Kusihape on fruktoosi metabolismi produkt. Varem soovitati diabeediga patsientidel glükoosi asendada fruktoosiga. Tõepoolest, fruktoos ei vaja selle imendumiseks insuliini osalemist. Kuid fruktoosi on veelgi raskem seedida. Tähelepanu: suhkrus on sahharoosi molekuliks disahhariid - glükoos + fruktoos. Seega sööme vähem suhkrut.
5. Väldi alkoholi, eriti õlle joomist. Vein väikestes kogustes ei mõjuta kusihappe taset.
6. Väga intensiivne treening tõstab kusihappe taset.
7. Sa pead jooma palju vett. See võimaldab teil kusihapet tõhusalt eemaldada.
Kui teil on kõrge kusihappesisaldus
Noh, esiteks, õnneks pole see alati patoloogia: lühiajaline tõus võib olla normi variant.
Kui probleem on endiselt olemas, peate välja selgitama, millisel tasemel on rikkumine (see kõige esimene skeem): puriinide sünteesi häired (sama metaboolne sündroom), toitumisfaktor (sööme palju liha, joome õlu), neerufunktsiooni kahjustus (uriini erituvate hapete kahjustus) või kaasuvad haigused, millega kaasneb kudede hävimine.
Edu teile ja pädevatele arstidele.
Kui leiate tekstist kirjavea, andke mulle sellest teada. Valige tekstiosa ja klõpsake Ctrl+Enter.
Lämmastikuühendite hulka kuuluvad valgud, peptiidid, aminohapped ja nende derivaadid, nukleiinhapped, nukleotiidid ja nende derivaadid, aga ka suhkrute lämmastikuderivaadid. Suurema osa fikseeritud lämmastikust moodustavad valgud, seega identifitseeritakse lämmastiku metabolism sageli valkude metabolismiga. Kuigi valkude lagunemisel tekkinud aminohappeid saab taaskasutada valkude biosünteesiks (keskmiselt 5 korda), on normaalseks eluks vajalik pidev aminohapetega varustamine toidus. Valgutoodete ebapiisava tarbimise tagajärgi illustreerib kwashiorkor, laste haigus, mis on tingitud valgupuudusest, kui süsivesikute ja lipiidide vajadus on normaalselt rahuldatud (joonis 8-1).
Riis. 8-1. Kwashiorkori nõiaring
Üldtunnustatud seisukoht on, et täiskasvanu minimaalne päevane valguvajadus on umbes 100 g. See vajadus suureneb intensiivse kasvu, haigusest taastumise, raseduse ja imetamise ajal. Lämmastiku tasakaal (s.o tarbitud ja väljutatava lämmastiku suhe) on organismi seisundi ja eelkõige kasvuprotsesside oluline näitaja.
Valkude eluiga organismis varieerub kümnetest minutitest mitme kuuni, keskmiselt 3 nädalat. Seda perioodi võivad vähendada katabolismi stimuleerivad tegurid, nagu glükokortikoidid või põletikueelsed tsütokiinid.
VALGUDE SEEDIMINE
Mao pepsiin. Peamine mao proteinaasi pepsiin (aspartaatproteinaaside perekond, mis põhineb kahe asparagiinhappejäägi olemasolul aktiivses keskuses) moodustub maoõõne happelise keskkonna mõjul kahest prekursorist - pepsinogeenidest I ja II (või A). ja C), mida eritavad mao peamised rakud. Toidu tarbimine stimuleerib pepsinogeenide vabanemist sekretoorsetest graanulitest koos sünteesi suurenemisega de novo. Stimuleeriv toime erinevate närvi- ja humoraalsete tegurite pepsinogeenide sekretsioonile realiseerub eelkõige cAMP (sekretiin, vasoaktiivne soolepeptiid (VIP), katehhoolamiinid) või intratsellulaarse Ca 2+ (koletsüstokiniin - CCK, gastriin, gastriini vabastav) taseme tõstmisega. peptiid - GRP, bombesiin, atsetüülkoliin). Somatostatiin pärsib sekretsiooni. Glükokortikoidide farmakoloogiliste annuste teadaolev haavandiline toime võib osaliselt olla tingitud nende stimuleerivast toimest pepsinogeeni C geeni ekspressioonile, mille regulatoorses piirkonnas leitakse funktsionaalne GKSE. Pepsinogeeni ekspressiooni normaalseks tasemeks on vaja glükokortikoidide baastasemeid.
Pankrease proteinaasid. Maost soolde siseneva pepsiini seedimise peptiidprodukte hüdrolüüsivad edasi mitmed proteinaasid, mis moodustuvad kaksteistsõrmiksoole valendikus kõhunäärmest tulevatest inaktiivsetest prekursoritest (zymogeenidest). Aktiveerimiskaskaadi käivitab enteropeptidaas (või enterokinaas), mida ekspresseeritakse enterotsüütides ja kaksteistsõrmiksoole pokaalrakkudes. See ensüüm eemaldab inhibeerivad fragmendid trüpsinogeeni polüpeptiididest (3 isovormi). Saadud trüpsiin (seriini proteinaaside perekond) katalüüsib veelgi kümotrüpsinogeeni (seriini proteinaaside perekond) ja prokarboksüpeptidaasi (2 isovormi) autoaktivatsiooni ja aktivatsiooni. Sümogeenide sünteesi ja sekretsiooni kõhunäärmes stimuleerivad adaptiivselt valgurikkad toidud, arvatavasti
eriti koletsüstokiniini osalusel. Samuti on näidatud atsetüülkoliini, insuliini, sekretiini, bombesiini stimuleerivat toimet sümogeenide sekretsioonile. Arvatakse, et pankreatiidi tekke üheks põhjuseks võib olla zymogeenide rakusisene (st enneaegne) aktiveerimine.
OLIGOPEPTIIDIDE JA AMINOHAPETE TRANSPORT
Oligopeptiidid. Põhiosa valkude seedimise saadustest maos ja sooltes moodustavad di- ja tripeptiidid. Need oligopeptiidid imenduvad peensoole limaskesta rakkudesse, kus need hüdrolüüsitakse peptidaaside toimel aminohapeteks. Oligopeptiidide transport läbi soolerakkude apikaalse membraani on energiast sõltuv ja selle määrab H+-peptiidi kaastransporter PepT1.
See transporter on glükosüülitud valk, mis sisaldab 12 transmembraanset domeeni. Seda iseloomustab madal substraadi spetsiifilisus (võimeline transportima 400 erinevat dipeptiidi, 8000 tripeptiidi, mitmeid ravimeid, mis imiteerivad di- ja tripeptiidide struktuuri, näiteks β-laktaamantibiootikumid) ja madal afiinsus substraatide suhtes. Transpordienergiat annab Na + ,K + -ATPaasi töö, mis paikneb rakkude basolateraalsel pinnal. Na+ kontsentratsiooni langus rakkudes on liikumapanev jõud rakkude apikaalsel pinnal paikneva Na + /H + soojusvaheti toimimiseks, mis eemaldab rakkudest prootonid soolestiku luumenisse. Seejärel suunatakse need prootonid koos oligopeptiididega rakkudesse tagasi H+-peptiidi kaastransporteri PepT1 kaudu (joonis 8-2).
H + -peptiidi kotransporteri PepT1 aktiivsust reguleerivad substraadid transkriptsiooni tasemel, kuid andmed selle regulatsiooni suuna kohta on vastuolulised. Kuigi dipeptiidide stimuleerivat toimet PepT1 tasemele saab reprodutseerida in vitro soolestiku rakukultuuri kohta on võimalik, et in vivo substraadi toimet võib vahendada selle stimuleeriv toime seedetrakti hormoonidele, nagu glükagoonitaoline peptiid.
 Insuliin stimuleerib PepT1 aktiivsust, kuid see toimub translatsioonijärgsel tasemel, suurendades PepT1 sisestamist plasmamembraani. Leptiin, mis võib jõuda soolerakkudesse nii süsteemse vereringe kui ka luumeni kaudu Riis. 8-2.
Insuliin stimuleerib PepT1 aktiivsust, kuid see toimub translatsioonijärgsel tasemel, suurendades PepT1 sisestamist plasmamembraani. Leptiin, mis võib jõuda soolerakkudesse nii süsteemse vereringe kui ka luumeni kaudu Riis. 8-2.
Oligopeptiidide transport soole limaskesta rakkudesse
soolest pärast selle sekretsiooni mao poolt, suurendab PepT1 ekspressiooni transkriptsiooni tasemel ja vähendab kilpnäärmehormoone.
Neerud, kopsud, aju ja mitmed teised elundid ja koed ekspresseerivad PepT1-ga seotud oligopeptiidi transporterit PepT2. Selle transporteri tööpõhimõte on sarnane PepT1 omaga.
PepT2 võib kasutada substraatidena di-, tri- ja tetrapeptiide, eelistatavalt dipeptiide. Selle afiinsus substraatide suhtes on oluliselt kõrgem kui PepT1-l. Neerudes paikneb PepT2 neerutuubulite rakkude apikaalsel pinnal, kus see vahendab oligopeptiidide reabsorptsiooni uriinist. Välja arvatud kilpnäärmehormoonide ja epidermise kasvufaktori pärssiv toime PepT2 ekspressioonile, ei ole selle transporteri hormonaalset regulatsiooni praktiliselt uuritud. Rakkude plasmamembraani lipiidne kaksikkiht on aminohapetele mitteläbilaskev. Aminohapete imendumiseks soolestikus, reabsorptsiooniks uriinist, tagasihaardeks sünapsis, vereringesse vabanemiseks ja tarbijarakkudesse sisenemiseks kasutatakse laia valikut aminohapete transportereid, mis jagunevad kahte põhirühma: sõltumatud ja Na + sõltuv. Aminohapete transporteritel on teatud substraatide rühmade suhtes väga kõrge spetsiifilisus.
Biogeensed amiinid, kilpnäärmehormoonid ja mitmed ravimid võivad samuti olla aminohapete transportijate substraadid. Märkimisväärne osa Na+-sõltuvatest transporteritest teostab aminohapete energiast sõltuvat transporti läbi plasmamembraani, samas kui Na+-sõltumatud transporterid tagavad substraatide hõlbustatud difusiooni. Esimese rühma transporterite näide on prootoniga seotud aminohapete transporter PAT1, mida ekspresseeritakse peamiselt soole limaskesta rakkude apikaalsel pinnal. Selle transporteri tööpõhimõte on sarnane ülalkirjeldatud oligopeptiidi transporteri PepT1 omaga. PAT1 aktiivsust saab reguleerida eelkõige Na + /H + -vaheti või seda reguleerivate valkude fosforüülimise teel. SN1 transporter kannab ka aminohappeid Na+/H+ vahetuse tõttu, kuid selle vahetuse viib läbi transporter ise ja see toimub vastupidises suunas, kui on kirjeldatud PAT1 transporteri puhul, ühendatud Na+/H+ vahetusega (joonis 8). -3). SN1 transporter tagab eelkõige glutamiini sisenemise periportaalsetesse hepatotsüütidesse karbamiidi sünteesiks ja vastupidi, peritsentraalsete hepatotsüütide kaudu glutamiini ekspordi vereringesse ja edasi neerudesse. Neerude proksimaalsetes tuubulites suureneb selle transporteri ekspressioon adaptiivselt kroonilise atsidoosi ajal (funktsioon on prootonite eritumise ammoniagenees, vt allpool) ja seda tõusu soodustavad glükokortikoidid. Aminohapete Na + -sõltuva transpordi teist varianti võib täheldada ATA1 transporteri puhul, mis teostab aminohapete ja Na + ühistransporti. Sel juhul toimivad prootonid transporteri allosteeriliste regulaatoritena (vt joonis 8-3).
Lihtsustatud difusiooniga aminohapete transpordisüsteemi näide on L-süsteem. Selle rühma transporterid on üles ehitatud kahest subühikust: kergest ja raskest, mis on seotud disulfiidsidemega. Kerge alaühik, nagu LAT1, läbib plasmamembraani 12 korda. Raske glükosüülitud subühik, nagu 4F2hc, sisaldab ainult ühte transmembraanset domeeni (joonis 8-4). See transporterite rühm tagab eelkõige rakus ja rakuvälises keskkonnas paiknevate aminohapete vahelise vahetuse. Selle transpordisüsteemi tähtsus näib olevat aminohapete transmembraanse liikumise tagamine, mis on Na + -sõltuvate transporterite jaoks halvad substraadid. Seega paiknes dimeerne transporter rbAT/ neerude proksimaalsete tuubulite rakkude apikaalsel pinnal.
 Riis. 8-3. Na+-sõltuvate aminohapete transporterite (a.a.) variandid: a- SN1 transporter teostab Na+ ja aminohapete ühistranspordi prootoni vastu (prooton on vajalik SN1 ümberorienteerumiseks membraanis). SN1 vahendab glutamiini importi periportaalsetesse hepatotsüütidesse vereringest ja sünteesitud glutamiini eksporti peritsentraalsetest hepatotsüütidest;
Riis. 8-3. Na+-sõltuvate aminohapete transporterite (a.a.) variandid: a- SN1 transporter teostab Na+ ja aminohapete ühistranspordi prootoni vastu (prooton on vajalik SN1 ümberorienteerumiseks membraanis). SN1 vahendab glutamiini importi periportaalsetesse hepatotsüütidesse vereringest ja sünteesitud glutamiini eksporti peritsentraalsetest hepatotsüütidest;
b- elektrogeenne transporter ATA1 toimib sarnaselt SN1-ga, kuid ei edasta prootonit
 Riis. 8-4. Aminohapete transportijad:
Riis. 8-4. Aminohapete transportijad:
A- dimeerne aminohapete transporter b 0,+ on üles ehitatud raskest ahelast rbAT (hele toon) ja kergest ahelast b 0,+ +AT (tume toon), mis on seotud disulfiidsidemega. Lokaliseeritud neerude, peensoole ja aju rakkude apikaalsel pinnal;
b- b 0,+ transporter teostab Na+-sõltumatut neutraalsete ja kahealuseliste aminohapete vahetust ning tagab koos seotud transporteriga LAT2-4F2hc koostöös Na+-sõltuvate transporteritega tsüstiini, arginiini, lüsiini, ornitiini reabsorptsiooni . Transporteri puudulikkusega kaasneb tsüstinuuria
b 0,.+ AT tagab tsüstiini reabsorptsiooni primaarsest uriinist, mis seejärel muundatakse rakkudes tsüsteiiniks, mis seejärel eritub verre läbi raku basolateraalse membraani läbi teise dimeerse transporteri - 4F2/LAT2. Selle reabsorptsioonisüsteemi energeetilise töö tagab Na+-sõltuv aminohapete transport, mis vahetatakse vastavalt tsüstiini ja tsüsteiini vastu. Näidatud on glükokortikoidide stimuleerivat toimet seda tüüpi transporterite ekspressioonile neerudes.
AMINOHAPETE AINEVAHETUS
Aminohapped on ühelt poolt valkude ja teiste lämmastikku sisaldavate ühendite biosünteesi ehitusmaterjal, teisalt aga energiaallikas. Sõltuvalt sellest, kas teatud aminohapet saab kehas sünteesida, eristatakse mitteolulisi ja asendamatuid aminohappeid.
Valkude biosünteesi jaoks on vajalik tingimus aminohapete vahelise tasakaalu säilitamine rakus, mis vastab nende sisaldusele valkudes. See tasakaal säilib suuresti tänu aminohapete omavahelisele muundamisele. Interkonversioonid hõlmavad kahte tüüpi reaktsioone: transamiinimine ja oksüdatiivne deamineerimine / redutseeriv amiinimine. Neid samu reaktsioone kasutatakse aminohapete lagundamiseks glükoneogeneesi ajal.
Pöörduvad transaminatsioonireaktsioonid, st. aminorühma ülekandumist ühelt substraadilt teisele katalüüsivad aminotransferaasid. Üks substraat on aminohape X, mille nime järgi nimetatakse vastavat aminotransferaasi või vastavat α-ketohapet X." Teine substraat on esindatud α-ketoglutaraadi/glutamaadi paariga. Sõltuvalt kontsentratsioonide suhtest substraatide puhul on reaktsioon suunatud kas aminohappe X või glutamaadi moodustumisele. Glutamaadi akumuleerumisel võib viimane muutuda α-ketohappe Y" aminorühma doonoriks koos aminohappe Y moodustumisega. reaktsioon, mida katalüüsib vastav aminotransferaas. Sarnaselt saab saavutada ka teiste aminohapete vahelise pariteedi.
Üleliigseid aminohappeid saab kõrvaldada glutamaadi oksüdatiivse deamineerimisega glutamaadi dehüdrogenaasi toimel. Redutseeriva amiinimise pöördreaktsioon võib vastupidi suurendada aminohapete kogumit rakus
(Joonis 8-5). Joonisel näidatud reaktsiooniahelat kasutatakse ka glükoneogeneesi protsessis: näiteks glükokortikoidide poolt stimuleeritud lihasvalgu lagunemisega stressi ajal kaasneb alaniini sisenemine verre, mis transporditakse püruvaadi kaudu maksa.
 Riis. 8-5. Aminohapete vastastikused muundumised.
Riis. 8-5. Aminohapete vastastikused muundumised.
Aminotransferaasid kannavad aminorühma α-ketoglutaraadiks. Saadud glutamaat läbib oksüdatiivse deaminatsiooni. Pöördreaktsioonid tagavad aminohapete sünteesi.
saadetakse glükoosi sünteesile. Glükokortikoidide poolt indutseeritud glükoneogeneesi soodustab ka nende hormoonide teadaolev võime stimuleerida mitmete aminotransferaaside geenide transkriptsiooni.
AMMONIAAGI VAHETUS
Glutamiin on oluline energiaallikas, eriti soolerakkude ja immuunsüsteemi jaoks, glükoneogeneesi eelkäija ja ammoniaagi transportija. See on veres domineeriv aminohape. Glutamiini metabolism toimub tsütosoolse ensüümi glutamiini süntaasi ja kahe mitokondriaalse glutaminaasi isoensüümi osalusel (joonis 8-6).
Maksa glutaminaasi aktiivsus suureneb paastumise, diabeedi ja toidu kõrge valgusisalduse korral. Kõiki neid haigusseisundeid iseloomustab maksa sisenevate aminohapete suurenenud katabolism, mis on osaliselt suunatud glükoneogeneesi suurendamisele ja liigse lämmastiku eemaldamisele uurea tsükli kaudu.
 Ensüümide aktiivsuse suurenemine toimub eelkõige glükagooni mõjul Riis. 8-6.
Ensüümide aktiivsuse suurenemine toimub eelkõige glükagooni mõjul Riis. 8-6.
Glutamaadi ja glutamiini vastastikused muundumised
geeni transkriptsiooni indutseerimise tõttu cAMP-responsive element (CRE) ja glükokortikoidide kaudu GKCE kaudu (joonis 8-7).
Kroonilise metaboolse atsidoosi korral suureneb neerude glutaminaasi aktiivsus. Selle ensüümi põhiülesanne on prootonite sidumiseks ja uriiniga väljutamiseks ammoniaagi tootmine. Mõju realiseerub transkriptsioonijärgsel tasemel tänu ensüümi mRNA stabiliseerumisele. Mehhanism hõlmab mRNA 3"-mittetransleeritava piirkonna (mis on adenosiini ja uridiiniga rikastatud 8-aluseline kordus) pH-le reageeriva elemendi (pHRE) interaktsiooni zeta-kristalliiniga (vt joonis 8-7). .
Stressi korral suureneb glutamiini süntaasi aktiivsus lihastes ja kopsudes. Mõju realiseerub transkriptsiooni tasemel. Näiteks kopsudes suureneb ensüümi mRNA 10 korda. Glükokortikoidid mõjutavad ensüümi ekspressiooni kanoonilise GKSE kaudu GKSE 1. intronis ja GKSE 3. poolsaidis geeni kaugemas regulatoorses piirkonnas.
Kroonilise stressi korral suureneb glutamaatdehüdrogenaasi aktiivsus neerudes, muutes glutamaadi α-ketoglutaraadiks koos ammoniaagi vabanemisega. Reguleerimine toimub transkriptsioonijärgsel tasemel, stabiliseerides ensüümi mRNA tänu nelja 3"-mittetransleeritavas piirkonnas paikneva pHRE seondumisele zetakristalliiniga, st sarnaselt neerude glutaminaasi regulatsiooniga.
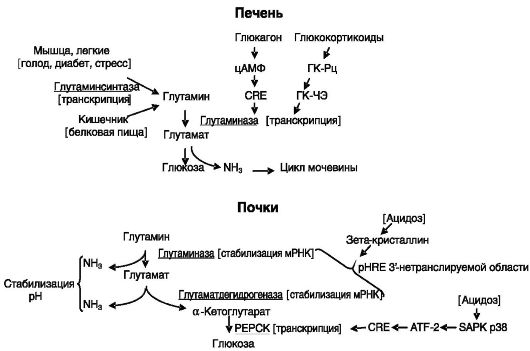 Atsidoosi käigus neerudes moodustunud α-ketoglutaraadi kasutamine toimub glükoneogeneesi ensüümide indutseerimise kaudu. Neist ühe, fosfoenoolpüruvaadi karboksükinaasi (PEPCK) indutseerimine toimub transkriptsiooni tasemel: sisemise vähenemine. Riis. 8-7.
Atsidoosi käigus neerudes moodustunud α-ketoglutaraadi kasutamine toimub glükoneogeneesi ensüümide indutseerimise kaudu. Neist ühe, fosfoenoolpüruvaadi karboksükinaasi (PEPCK) indutseerimine toimub transkriptsiooni tasemel: sisemise vähenemine. Riis. 8-7.
Raku pH-ga kaasneb stressiga aktiveeritud proteiinkinaasi p38 (SAPK p38) aktiveerimine, mis fosforüülib transkriptsioonifaktorit ATF-2, mis interakteerub PEPCK geeni cAMP-responsive elementiga (CRE), mis stimuleerib transkriptsiooni.
UREA tsükkel
Imetajate maksas toimuv uurea energiast sõltuv moodustumine on peamine viis aminohapetest ja muudest lämmastikku sisaldavatest ühenditest moodustunud ammoniaagi kasutamiseks. Karbamiidi süntees hõlmab karbamoüülfosfaadi moodustumist süsinikdioksiidist ja ammoniaagist, mis interakteerudes ornitiiniga tekitab tsitrulliini. Aspartaadi aminorühma osalusel vaheühendi argininosuktsiinhappe moodustumisel sünteesitakse arginiin, mille hüdrolüüsil tekib uurea ja algne ornitiin (joonis 8-8).
Karbamoüülfosfaadi sünteesi võivad katalüüsida kaks ensüümi: mitokondriaalne karbamoüülfosfaadi süntaas I, mis on iseloomulik maksale ja osaliselt sooltele, ja laialdaselt ekspresseeritud valk CAD (karbamoüülfosfaadi süntaas II), millel on karbamoüülfosfaadi süntaasi aktiivsus.
 Riis. 8-8. Karbamiidi tsükkel
Riis. 8-8. Karbamiidi tsükkel
bamoüülfosfaadi süntaas, aspartaattranskarbamülaas ja dihüdroorotaas, mis katalüüsib 3 kuuest pürimidiini biosünteesi reaktsioonist.
Karbamoüülrühma ülekandumist ornitiinile katalüüsib ornitiinkarbamoüültransferaas. Mitokondriaalset ensüümi ekspresseeritakse peamiselt maksas ja soole limaskestas. Ensüümipuudus on üks hüperammoneemia põhjusi, millega kaasnevad oksendamine, letargia, krambid ja mõnikord surm.
Tsitrulliini ja aspartaadi koostoimet katalüüsiva ensüümi argininosuktsinaadi süntaasi puudus põhjustab tsitrullineemiat, millega kaasnevad oksendamine ja vaimne alaareng. Arginiini moodustumist argininosuktsiinhappest katalüüsib argininosuktsinaatlüaas. Ensüümipuudus põhjustab arginiini suktsinitsisiduuriat, millega kaasneb vaimse ja füüsilise arengu mahajäämus, maksa suurenemine, nahakahjustus ja perioodiline teadvusekaotus.
Arginiinist karbamiidi moodustumist katalüüsib arginaas. Ensüümi esindavad kaks isosüümi (I ja II). Tsütosoolne arginaas I (homotrimeer) ekspresseerub peamiselt maksas, kus see vahendab uurea moodustumist. Maksa arginaasi puudulikkus põhjustab arginineemiat, millega kaasneb psühhomotoorse funktsiooni hiline areng, nelja jäseme spastiline halvatus.
tey.
Arginaas I, aga ka mitokondriaalse arginaas II (homoheksameer) ekspressioon teistes elundites ja kudedes võib pakkuda muid lämmastikuühendite metabolismi aspekte (pakkudes ornitiini glutamaadi, glutamiini, GABA, agmatiini, polüamiinide, kreatiini, proliini biosünteesiks , EI). Eelkõige võib arginaas konkureerida NO süntaasiga arginiini kui substraadi pärast ja osaleda seeläbi NO ja cGMP poolt kontrollitavate protsesside reguleerimises (näiteks erektsioon).
 Karbamiidi tsükli ensüüme stimuleerivad kataboolsed hormoonid (glükokortikoidid, glükagoon) transkriptsiooni tasemel. Induktsioon võib olla otsene või kaudne transkriptsioonifaktori C/EBP biosünteesi stimuleerimise kaudu (joonis 8-9). Riis. 8-9.
Karbamiidi tsükli ensüüme stimuleerivad kataboolsed hormoonid (glükokortikoidid, glükagoon) transkriptsiooni tasemel. Induktsioon võib olla otsene või kaudne transkriptsioonifaktori C/EBP biosünteesi stimuleerimise kaudu (joonis 8-9). Riis. 8-9.
Kaugvõimendaja stimuleerib karbamoüülfosfaadi süntaasi geeni transkriptsiooni glükokortikoidide ja glükagooni (a) poolt. Arginaas-1 geeni tugevdaja stimuleerib glükagooni ja (kaudselt, C/EBP indutseerimise kaudu) glükokortikoidide (b):
C/EBP, CCAAT/võimendajat siduv valk; HNF3 – hepatotsüütide tuumafaktor 3; GRU - glükokortikoidide sensor Adibi S.A.
Soole oligopeptiidi transporteri (Pept-1) ekspressiooni reguleerimine tervises ja haigustes. Am J Physiol Gastrointest Liver Physiol. 2003;285(5):G779-788. Curthoys N.P., Gstraunthaler G.
Neerude suurenenud geeniekspressiooni mehhanism metaboolse atsidoosi ajal. Am J Physiol Renal Physiol. 2001;281(3):F381-390. Desvergne B., Michalik L., Wahli W.
Ainevahetuse transkriptsiooniline reguleerimine. Physiol Rev. 2006;86(2):465-514. Wagner C.A., Lang F., Broer S.
Heterodimeersete aminohapete transporterite funktsioon ja struktuur. Am J Physiol Cell Physiol. 2001; 281(4):C1077-1093. Wu G., Jaeger L.A., Bazer F.W., Rhoads J.M.
Arginiini puudulikkus enneaegsetel imikutel: biokeemilised mehhanismid ja toitumisalased tagajärjed. J Nutr Biochem. 2004;15(8):442-451.
Valgu lämmastiku eritumise vorm - ammoniaagi, uurea või kusihappe kujul - on tihedalt seotud värvuse elutingimuste ja vee kättesaadavusega (tabel 10.4). Ammoniaak on väga mürgine isegi väga väikestes kontsentratsioonides, seega peab see kiiresti minema
Enamikul veeselgrootutel on valkude ainevahetuse lõpp-produktiks ammoniaak. Tänu oma kergele lahustuvusele ja madalale molekulmassile hajub see ülikiiresti. Märkimisväärne osa sellest võib erituda läbi mis tahes veega kokkupuutuva pinna, mitte tingimata neerude kaudu. Luukaladel eritub suurem osa lämmastikust ammoniaagi kujul lõpuste kaudu. Karpkalal ja kuldkalal eritavad lõpused 6-10 korda rohkem lämmastikku kui neerud ja ainult 10% sellest moodustab uurea; ülejäänud 90% eritub ammoniaagina (Smith, 1929).
UREAM
Karbamiid lahustub vees kergesti ja on üsna madala toksilisusega. Karbamiidi sünteesi kõrgematel loomadel uuris kuulus biokeemik Hans Krebs – sama teadlane, kelle järgi nimetati oksüdatiivse energia metabolismi tsükkel (trikarboksüülhappe tsükkel ehk Krebsi tsükkel).
Karbamiidi sünteesil kondenseeruvad ammoniaak ja süsinikdioksiid fosfaadiga, moodustades karbamoüülfosfaadi, mida seejärel kasutatakse tsitrulliini sünteesimiseks ornitiinist, nagu on näidatud joonisel fig. 10.13. Pärast seda lisatakse asparagiinhappest veel üks ammoniaagi molekul ja see viib aminohappe arginiini moodustumiseni. Ensüümi arginaasi juuresolekul laguneb arginiin uureaks ja ornitiiniks. Ornitiinist sünteesitakse uus tsitrulliini molekul ja kogu tsükkel kordub; seetõttu nimetatakse kogu seda transformatsiooni teed ornitiini tsükkel uurea süntees. Arginaasi esinemine loomal näitab tema võimet toota karbamiidi ja sageli näitab, et uurea on tema peamine lämmastikueritus. Kuid see ei pruugi nii olla, kuna arginaas võib esineda isegi kogu tsükli puudumisel.
UREEMIA SELGroogsetes
Selgroogsed, kes eritavad peamiselt uureat ja mille sünteesiks on ornitiinitsükli ensüümid, on näidatud joonisel fig. 10.14. Luukalad eritavad teatud kogust karbamiidi ning elastsusharudel, kahepaiksetel ja imetajatel on see peamine lämmastikueritus. Elasmobranchides (haid ja raid), samuti krabisöövas konnas ja koelakantis Latimeeria uurea jääb kehasse ja mängib
omab olulist rolli eneseregulatsioonis ja on seetõttu väärtuslik ainevahetusprodukt. Elasmobranchide korral filtreeritakse uurea glomerulites, kuid selle tähtsuse tõttu osmoregulatsiooni jaoks ei tohiks see uriiniga kaduda; seetõttu tagastatakse see tuubulites toimuva aktiivse reabsorptsiooni tulemusena. Kahepaiksete puhul on olukord erinev.

Karbamiid filtreeritakse ja lisaks lisatakse märkimisväärne kogus seda uriinile aktiivse sekretsiooni teel tuubulites. Seega on karbamiidi aktiivne tubulaarne transport nii elastseharudel kui ka kahepaiksetel, kuid see kulgeb nendes rühmades eri suundades. Ilmselgelt ei ole pumpamismehhanismid siin metaboolselt identsed, kuna katsed mitmete üksteise lähedal asuvate karbamiidi derivaatidega annavad mõlemas loomarühmas erinevaid tulemusi (tabel 10.5). See on suurepärane näide sellest, kuidas sama füsioloogiline funktsioon toimub kahes rühmas sõltumatult, ilma et sama eesmärgi saavutamiseks (antud juhul aktiivse uurea transpordi) saavutamiseks kasutataks samu mehhanisme.
Krabi sööval konnal, kes säilitab osmoregulatsiooniks ka karbamiidi, toimub selle aine aktiivne reabsorptsioon tuubulites
ei leitud (Schmidt-Nielsen, Lee, 1962). Tema uriin moodustub aeglaselt ja neerutuubulid on uureale väga läbilaskvad. Seetõttu difundeerub uurea torukujulisest vedelikust välja

Riis. 10.14. Lämmastiku vabanemine selgroogsete fülogeneesi erinevatel etappidel. Jooned" on ümbritsetud loomarühmadega, kes eritavad peamiste väljaheidetena vastavalt ammoniaaki, uureat ja kusihapet. (B. Schmidt-Nielsen, 3972.)
tagasi verre ja ilmub uriinis ligikaudu samas kontsentratsioonis kui veres. Seega kaob uriiniga vaid väike kogus.
Kui tavalistel konnadel toimub karbamiidi aktiivne tubulaarne sekretsioon, siis miks krabisööv konn ei kasuta
Tabel 10.5
Karbamiidi transpordivad aktiivselt hai (aktiivne reabsorptsioon) ja konna (aktiivne sekretsioon) neerutorukesed. Kuid kolme muu sarnase ainega on nende kahe loomaliigi tulemused täiesti erinevad. See viitab sellele, et nende neerude raku transpordimehhanism on erinev. (B. Schmidt-Nielsen, Rabinovitz, 1964)

selline pump, lihtsalt muutes selle suunda vastupidiseks? Sellele küsimusele ei ole lihtne vastata, kuid näib, et aktiivse transpordi suund on konservatiivne füsioloogiline funktsioon, mida ei saa kergesti muuta. Nagu juba nägime, säilib nii konna nahas kui ka imetaja neerus naatriumkloriidi aktiivse transpordi suund väljast keha sisemusse. Kuid imetaja neerudes kasutatakse NaCl-i sissepoole tagurpidi transporti toruvedelikust kehasse paljunevas vastuvoolusüsteemis nii, et lõpptulemuseks on siiski kontsentreeritud uriin.
Tavaline seisukoht karbamiidi eritumisel imetajate neerude kaudu on see, et uurea filtreeritakse glomerulites ja lastakse seejärel passiivselt läbi tuubulite, kuigi osa sellest difundeerub oma suure difusiooni tõttu passiivselt tagasi verre. Siiski on; veenvad tõendid selle kohta, et uurea on paljuneva vastuvoolusüsteemi oluline element ja et uurea eritumise viis on imetajate neerufunktsiooni oluline element.
UREA JA METAMORFOOS KAHEPEASLASTEL
Konnade ja kärnkonnade kullesed toodavad peamiselt ammoniaaki; täiskasvanud loomad eritavad uureat. Konna juures (Rana temporaria), kärnkonnad (Bufo bufo) tritoon (Triturus uulgaris) ja teistel kahepaiksetel on metamorfoosi ajal selge üleminek ammoniaagi vabanemiselt uurea eritumisele. Lõuna-Aafrika küüniskonn aga (Xenopus), mis jääb vette ka täiskasvanuna, eraldab selles etapis jätkuvalt ammoniaaki (tabel 10.6).
Üleminek karbamiidi eritumisele poolmaise kahepaikse metamorfoosi ajal on seotud; koos kõigi ornitiinitsükli ensüümide aktiivsuse märgatava suurenemisega maksas (Brown et al., 1959).
Tabel 10.6
Ammoniaagi tootmine maakärnkonnas Bufo bufo ja täiesti vees elaval sabata kahepaiksel Xenopus laevis. Numbrid näitavad vaba ammoniaagi eraldumist protsentides erinevates arenguetappides vabanenud ammoniaagi ja karbamiidi koguhulgast. (Munro, 1953)

Huvitaval kombel on vees elavate kahepaiksete isendid Xenopus, ekstraheeritakse: mitme nädala jooksul veest koguneb uurea verre ja kudedesse. Karbamiidi kogunemist saab esile kutsuda, pannes loomad 0,9% NaCl lahusesse. Kui täiskasvanuid hoiti veest eemal, kuid niiskes samblas (dehüdratsiooni vältimiseks), tõusis uurea kontsentratsioon veres 10-,20 korda ja jõudis peaaegu 100 mmol/l-ni. Pärast loomade vette naasmist vabanes liigne karbamiid (Balinsky et al, 1961).
Üksikisikute rühmas Xenopus, mis looduslikes tingimustes koges suvist põuda kuivanud tiigi ääres mudas, tõusis ka uurea kontsentratsioon 15-20 korda. Karbamiidi sünteesis osalevate ensüümide hulgas suurenes sünteesi esimese etapi eest vastutava karbamoüülfosfaadi süntetaasi hulk (vt joonis 10.13) ligikaudu kuus korda, kuid tsükli ülejäänud ensüümide aktiivsus ei muutunud. On võimalik, et karbamoüülfosfaadi süntees on karbamiidi sünteesi kiirust piirav etapp ja selle ensüümi suurenemine hoiab tõenäoliselt plasma ammoniaagi madala, kui loomad on veest väljas (Balinsky et al., 1967).
Karbamiid kopsukalades
Aafrika kopsukalades Protopterus Toimuvad absoluutselt samad muutused, mis kahepaiksetel. Tavalistes tingimustes eraldab selline kala vees elades palju ammoniaaki
(ja teatud kogus karbamiidi), aga kui ta on põua ajal kookonis kuivatatud muda sees, siis kõik selle lämmastikujäätmed muundatakse karbamiidiks, mis koguneb verre, kus selle kontsentratsioon kala lõpuks kolme- aasta kookonis viibimine võib ulatuda 3%-ni ( 500 mmol/l) (Smith, 1959).
Kõik viis ornitiinitsükli ensüümi leiti Aafrika kopsukala maksast (Janssens ja Cohen, 1966). Kahe karbamiidi sünteesi kiirust piirava ensüümi tase on selles kalas ja konnakulleses sarnane Rana catsbeia ja oluliselt madalam kui täiskasvanud konnadel. See on kooskõlas tõsiasjaga, et kopsukalad eraldavad vees olles eelistatavalt ammoniaaki. Arvutati aga, et mittetalveuneva kopsukala maksas sisalduv ornitiinitsükli ensüümide hulk on piisav, et tagada talveunes tegelikult täheldatud karbamiidi kogunemine (Forster ja Goldstein, 1966).
Austraalia kopsukalas Neoceratodus ornitiiniringe ensüümide kontsentratsioonid on madalad, mis on kooskõlas selle kala elustiiliga: ta kasutab kopse vaid täiendava hingamisorganina ja suudab õhus püsida vaid lühikest aega (kopskalade hingamise kohta vt. 2. peatükk). Karbamiidi süntees Austraalia kopsukala maksa lõikudes on sada korda aeglasem kui Aafrika kopsukalal. See on jällegi kooskõlas esimese neist puhtalt veelise eluviisiga (Goldstein et al., 1967).
KURIHAPE
Kusihappe tootmine domineerib putukatel, maismaatigudel, enamikul roomajatel ja lindudel. Kõik need on tüüpiliselt maismaaloomad ja kusihappe teket neis võib pidada tõhusaks kohanemiseks, mis säästab vett maal elades. Kuna kusihape ja selle soolad lahustuvad vees väga halvasti (selle lahustuvus on umbes 6 mg 1 liitri vee kohta), põhjustab vee reabsorptsioon uriinist kusihappe ja selle soolade sadestumist.
KUISIHAPE LINDUDEL JA PUTUKATEL
Lindude väljaheidete pooltahke valge osa on uriin ja koosneb peamiselt kusihappest; Lämmastikku sisaldavate väljaheidete väljutamiseks kulutavad linnud mõnele putukale väga vähe vett, uriiniga veekao vähenemine on jõudnud nii kaugele, et nad ei erita kusihapet üldse, vaid ladestavad seda erinevatesse kehaosadesse, peamiselt
paks keha. Seetõttu ei vaja sellised vormid lõplike lämmastikproduktide eemaldamiseks vett üldse (Kilby, 1963).
On oletatud, et kusihappe kasutamine peamise väljaheidetena annab lindudele veel ühe eelise. Kuna nad vajavad uriini tootmiseks vähe vett, arvatakse, et kusihappe sekretsioon vähendab lendavate lindude kehakaalu. Kuid see idee ei ole veenev, kuna linnud, kellel on juurdepääs veele (nii mage- kui ka mereveele), eritavad sageli suures koguses vedelat uriini.
CLEIDOIC MUNA
Joseph Needham tegi ettepaneku, et uureat tootvate selgroogsete (imetajad ja kahepaiksed) ja kusihapet tootvate loomade (roomajad ja linnud) erinevus tuleneb peamiselt paljunemisviisist. Kahepaikse muna areneb vees, imetaja embrüo aga vedelas keskkonnas emakas, kus ainevahetuse jääkained satuvad ema verre. Seevastu roomajate ja lindude embrüonaalne areng toimub suletud, nn Cleidic muna, mis vahetab väliskeskkonnaga ainult gaase ja kõik väljaheited jäävad koore sisse. Cleidic muna veevaru on väga väike ja ammoniaak on loomulikult liiga mürgine, et embrüo taluks selle suurtes kogustes. Kui karbamiidi tekiks, jääks see munarakku ja koguneks lahustunud olekus. Samal ajal võib kusihape sadestuda ja seega sisuliselt elimineerida; see juhtub siis, kui see ladestub kristallidena allantoisis, mis toimib seega embrüonaalse põiena.
URIHAPE ROOMAJATEL
Sisalikud ja maod eritavad peamiselt kusihapet; paljud kilpkonnad eritavad kusihappe ja uurea segu, krokodillid aga peamiselt ammoniaaki (Cragg et al., 1961). See on kooskõlas üldise ideega, et lämmastiku eritumise viis on tihedalt seotud keskkonnas saadaoleva vee hulgaga.
Krokodillid ja alligaatorid eritavad ammoniaaki oma uriiniga, kus põhikatiooniks on NH4+ ja peamiseks aniooniks HCO 3 - (Coulson et al., 1950; Goulson ja Hernandez, 1955). Võimalik, et nende ioonide olemasolu uriinis aitab neid magevett
Loomadel on parem säilitada Na + ja C1 - ioone, mille kadu roojaga, muide, on samuti väga väike.
Kilpkonnade elupaiga ja nende lämmastiku eraldumise vahelises tihedas seoses võib vaevalt olla kahtlust. Tabelis Joonisel 10.7 on näidatud Londoni loomaaiast saadud kaheksa kilpkonnaliigi uriiniproovide koostis. Kõige silmatorkavama veelise eluviisiga liigid toodavad märkimisväärses koguses ammoniaaki ja uureat ning ainult kusihappe jälgi; kõige enam maismaal asuvates vormides eritub üle poole lämmastikust kusihappe kujul.
Tabel 10.7
Lämmastiku osakaal erinevate kilpkonnade uriinis (protsendina kogu eritunud lämmastikust). Veega kõige enam seotud vormid ei tooda peaaegu üldse kusihapet, kuid see aine on domineeriv kuivade piirkondade maismaaliikides. Moyle, 1949 )

Selle kohta, kas kilpkonnad eritavad peamiselt uureat või kusihapet, on vastuoluline teave. Fakt on see, et mitte ainult liigid ei erine, vaid ka ühe liigi sees võivad mõned isendid eritada valdavalt kusihapet, teised – valdavalt uureat ja kolmandad – mõlema aine segu (Khalil, Haggag, 1955). Isegi sama isik võib aja jooksul liikuda ühest ühendusest teise. Mõned
sadestunud kusihappe kogus jääb kloaaki kinni ja uriini vedel osa eritub; see muudab tekkiva kusihappe määramise ühe või mitme uriiniproovi analüüsimisel ebausaldusväärseks: kui kloaaki ei tühjendata täielikult, võib saada väga madalaid numbreid ja sellise evakueerimisega, kui mõne aja jooksul kogunenud sete väljub. , kusihapet on liiga palju.
Kilpkonna juures Testudo mauritanicaüleminek uurealt kusihappele ja tagasi näib olevat otseselt sõltuv temperatuurist ja veesisaldusest kehas. Kusihappe tootmine suureneb, kui vedeliku tasakaal on ebasoodne, kuid mehhanism, mis seda biokeemilise aktiivsuse muutust juhib, on ebaselge.
Mainisime juba 9. peatükis, et Aafrika konn Chiromantis xerampelina kaotab vett läbi naha väga aeglaselt, umbes sama kiirusega kui roomajad. See sarnaneb roomajatega selle poolest, et eritab peamiselt kusihapet, mitte karbamiidi, nagu on tavaliselt omane täiskasvanud kahepaiksetele. See on sensatsiooniline fakt, kuna see on vastuolus üldtunnustatud ideega kahepaiksete lämmastiku eritumisest. Selle aruande täpsus on väljaspool kahtlust, kuna kusihapet määrati uriinis Chiromantis sellele ainele spetsiifilise ensümaatilise meetodiga ja leiti, et see moodustab kuni 60–75% uriini kuivkaalust (Loveridge, 1970).
Lõuna-Aafrika konn Phyllotnedusa sauvagii selles osas sarnaneb ta ka roomajatega. Veekadu läbi naha on samas suurusjärgus kui roomajatel nende kuiva nahaga ja uriin sisaldab suures koguses pooltahket uraadisetet (Shoemaker et al., 1972). Uraatide kujul Phyllomedusa 80% kogu lämmastikust vabaneb ja suurenenud veetarbimine ei muuda uraadi moodustumise intensiivsust. See liik jätkab peamiselt kusihappe eritumist isegi siis, kui seal on liigne vesi. Kui konn peab vett säästma, muutub väga oluliseks kusihappe eritumine (karbamiidi asemel). Arvutatakse, et kui selle konna eritusproduktiks oleks uurea, siis uriini moodustamiseks vajaks see umbes 60 ml vett päevas 1 kg kehakaalu kohta. Vahepeal tänu sellele, et P. sauvagii eritab kusihapet, kaotab see uriiniga vaid 3,8 ml vett päevas 1 kg kehakaalu kohta (Shoemaker, McClanahan, 1975).
AMMONIAAK JA NEEREFUNKTSIOON
Kõigest ülaltoodust võib tunduda, et ammoniaaki eraldavad peamiselt veeloomad, kuid see pole päris tõsi. Ammoniaaki leidub tavaliselt ka maismaaloomade uriinis, kus see aitab reguleerida uriini pH-d. Kui uriin muutub happeliseks happeliste jääkainete eraldumise tõttu, lisatakse selle neutraliseerimiseks ammoniaaki.
Liigne hape tekib tavaliselt valkude metabolismi käigus, kuna väävlit sisaldava aminohappe tsüsteiini oksüdatsiooni lõppsaadus on väävelhape. Mida happelisem on uriin, seda rohkem lisatakse ammoniaaki. Happelise uriini neutraliseerimiseks kasutatav ammoniaak moodustub neerudes aminohappest glutamiinist. Neerud sisaldavad glutaminaasi ja see on seal spetsiaalselt ammoniaagi tootmiseks. Seetõttu ei ole imetaja uriinis leiduv ammoniaak otseselt seotud aminohapete deaminatsiooni käigus maksas tekkiva ammoniaagiga ning selles mõttes ei tohiks seda pidada valkude ainevahetuse normaalseks lõpp-produktiks.
NULEIINHAPPED JA LÄMMASTIKU VÄLJENDAMINE
Nukleiinhapped sisaldavad kahte rühma lämmastikuühendeid: puriine (adeniin ja guaniin) ja pürimidiine (tsütosiin ja tümiin). Mõnedel loomadel erituvad puriinid kusihappena (mis ise on puriin); teistel loomadel laguneb puriini struktuur vaheühenditeks või ammoniaagiks, millest igaüks võib organismist erituda.
Puriinide metaboolset lagunemist ja selle lõppproduktide vabanemist ei ole uuritud nii põhjalikult kui valgu 13ot metabolismi. Kõige olulisemad andmed on toodud tabelis. 10.8. Lindudel, maismaa roomajatel ja putukatel lagundatakse puriinid kusihappeks ja viimane eritub organismist. Need on loomad, kellel kusihapet sünteesitakse amiinlämmastikust; Ilmselgelt oleks mõttetu, et loom sünteesiks kusihapet ja omaks samal ajal mehhanisme selle lagundamiseks. Seetõttu ei saa eeldada puriinide edasist lagunemist loomadel, kus kusihape on valkude metabolismi lõppsaadus.
Imetajatest moodustavad erirühma inimesed, inimahvid ja saksa dogi: nemad eritavad kusihapet, teised imetajad aga allantoiini. Allantoiin moodustub kusihappest ühe muundamise teel ensüümi urikaasi juuresolekul. Inimestel ja inimahvidel seda ensüümi pole. Oma vähese lahustuvuse tõttu ladestub kusihape mõnikord inimkehasse, põhjustades liigeste turset ja väga valusat haigust -
Tabel 10.8
Puriinide metabolismi lämmastikku sisaldavad lõpp-produktid erinevatel loomadel. (Keilin, 1959)

podagra Kui inimestel säiliks ensüüm urikaasi, poleks podagra olemas.
Kuigi dalmaatsia koer toodab palju rohkem kusihapet kui teised koerad, ei ole see tingitud mingist ainevahetushäirest. Iga koera maks sisaldab urikaasi ja toodab veidi allantoiini. Kuid dogil on neerudefekt, mis takistab kusihappe tubulaarset reabsorptsiooni (mis esineb teistel imetajatel, sealhulgas inimestel); seetõttu kaob dogi kusihape uriiniga kiiremini, kui maks selle allantoiiniks töötleb (Yu et al., I960). Paljud tõendid viitavad sellele, et dalmaatsia taani kusihape ei filtreerita mitte ainult glomerulites, vaid eritub ka aktiivse transpordi teel tuubulites (Keilin, 1959).
Puriinid adeniin ja guaniin on struktuurilt sarnased kusihappega: need sisaldavad ühte kuue- ja ühte viieliikmelist ringi. Kuid pürimidiinid (tsütosiin ja tümiin) on üksikud kuueliikmelised tsüklid, mis sisaldavad kahte lämmastikuaatomit. Kõrgematel selgroogsetel lõhustatakse pürimidiinid selle tsükli purustamisel, et saada üks ammoniaagi molekul ja üks β-aminohappe molekul. Viimane metaboliseeritakse seejärel tavapärase deaminatsiooniskeemi järgi.
Nukleiinhapete metabolismi kõige silmatorkavam omadus on see, et tabeli alguses on loetletud "kõrgemad" loomad. 10.8, puuduvad täielikult lagundamiseks vajalikud ensüümid
puriinid. "Madalamate" loomade hulgast leiame üha keerukamaid biokeemilisi ja ensümaatilisi süsteeme, mis lagundavad puriine veelgi, nii et "madalamatel" vormidel on kõige täielikum ensümaatiline aparaat.
MUUD LÄMMASTIKUÜHENDID
Ämblikul on peamine eritumine guaniin Ilmselt sünteesitakse see amiinlämmastikust, kuigi kogu selle moodustumise tee pole teada. Mõned ämblikud, sealhulgas linnutoidulised tarantlid, eritavad pärast söömist enam kui 90% kogu lämmastikust guaniini kujul (Peschen, 1939). Ühises aias ämblik Epeira diadeem guaniini identifitseerimist kinnitati väga spetsiifilise ensümaatilise meetodiga (Vajropala, 1935).
Guaniin on üsna levinud ka paljudel teistel loomadel. Näiteks kalasoomuste hõbedane läige on tingitud guaniinikristallide ladestumisest. aia tigu Helix vabastab guaniini, kuid ainult umbes 20% ulatuses kogu eritunud puriinide kogusest ja ülejäänud 80% on kusihape. Võimalik, et see fraktsioon on nukleiinhapete metabolismi produkt ja kusihape tekib valkude metabolismi tulemusena.
Aminohapped ei oma tähtsat kohta lämmastiku ainevahetuse saaduste hulgas, kuid neid leidub väikestes kogustes paljude loomade uriinis. Tundub, et loomal on kasulikum aminohappe deamineerida, tavapärasel viisil ammoniaaki eritada ja saadud orgaanilist hapet kasutada energiavahetuses. Kuid kuna aminohapetel on lämmastiku eritumisel vaid väike roll, siis seda küsimust siin ei käsitleta.
KOKKUVÕTETEOORIA
Üldiselt arvati, et lämmastiku eritumine arenevas tibu embrüos on ajas erinev ja läbib rea piike: esmalt on põhiproduktiks ammoniaak, seejärel uurea ja lõpuks kusihape. Eeldati, et selline areng võtab kokku evolutsiooni etapid, mis lindudel lõppevad kusihappe vabanemisega. On teatatud, et ammoniaagi tootmine tibu embrüos saavutab maksimumi 4 päeva, uurea 9 päeva ja kusihappe tootmine 11 päeva pärast inkubatsiooni algust (Baldwin, 1949).
Uuemad tööd näitavad, et lämmastiku eritumine tibu embrüos erineb järsult sellest eelnevalt kirjeldatud pildist (Clark ja Fischer, 1957). Kõik kolm peamist eritusprodukti – ammoniaak, uurea ja kusihape – moodustuvad ja esinevad juba embrüonaalse arengu algusest peale. Lõpu poole
Kusihappe inkubatsiooniperiood on palju pikem kui kahel teisel tootel. Kuid karbamiidi ja ammoniaagi kogus kasvab kogu inkubatsiooni ajal ning koorumise ajaks on mõlemat ainet ligikaudu võrdsetes kogustes. Inkubatsiooni lõpuks jõuab eralduva lämmastiku kogus 40 mg-ni, millest 23% jaguneb võrdselt karbamiidi ja ammoniaagi vahel ning ülejäänu on kusihape (joon. 10.15).

Mis on saadud tulemuste lahknevuse põhjus? Varasemad andmed võisid primitiivsemate analüüsimeetodite tõttu olla vähem täpsed, kuid see ei selgita tõenäoliselt üksikuid täheldatud piike. Peamine põhjus on lihtsalt see, et tulemusi väljendati iga eritusprodukti kogustes embrüo massiühiku kohta. Ja kuna embrüo on pidevalt ja mida kaugemal, seda kiiremini selle suurus suureneb,
seejärel jagades iga aine koguse embrüo massiga, tekib kunstlik piim.
Tegelikult on kõik kolm eritusprodukti olemas juba algusest peale ja nende arv kasvab järk-järgult kogu embrüonaalse arengu jooksul, kuid pärast 10. inkubatsioonipäeva suureneb ammoniaagi hulk veidi. Embrüo toodetav karbamiid ei sünteesita aminohappelisest lämmastikust ornitiini tsüklis, vaid arginaasi toimel arginiinile (Eakin ja Fisher, 1958). Seega ei toeta ei ammoniaagi moodustumine ega uurea süntees tibu embrüos ideed, et biokeemiliste mehhanismide ontogenees kordab lämmastiku eritumise evolutsioonilugu.
Uurisime erinevaid eritusorganeid ja kirjeldasime nende üldisi tunnuseid. Need organid eemaldavad ainevahetusjäägid, aitavad säilitada õiget soolade ja muude lahustunud ainete kontsentratsiooni ning reguleerivad keha veesisaldust, säästes hoolikalt vett, kui keha on vähe, ja eemaldades liigsed kogused.
Veenvad tõendid näitavad aga, et see ei vasta alati tõele. Mõned uurijad (nt Costa et al., 1968, 1974) on teatanud gaasilise lämmastiku moodustumisest imetajatel, keda toidetakse suures koguses valku. See teave peaks muutma mõningaid meie ideid valkude metabolismi ja lämmastikku sisaldavate lõpptoodete kohta.
Kreeka keeles tähendab kleisto suletud, kleis tähendab võtit.
Siin kirjeldatud kaks konnaliiki elavad kuivadel poolkõrbealadel. - Ligikaudu toim.
 Inimese ja looduse suhe Inimese ja looduse suhe
Inimese ja looduse suhe Inimese ja looduse suhe Ühendusvokaaliga liitsõnad
Ühendusvokaaliga liitsõnad Kusihape veres: normid ja kõrvalekalded, miks see suureneb, dieet selle vähendamiseks Lämmastiku ainevahetuse lõpp-produkt on
Kusihape veres: normid ja kõrvalekalded, miks see suureneb, dieet selle vähendamiseks Lämmastiku ainevahetuse lõpp-produkt on