Kjemisk formel for den primære strukturen til et protein. Fysisk-kjemiske egenskaper til proteiner
>> Kjemi: Proteiner
Proteiner, eller proteinstoffer, er høymolekylære (molekylvekten varierer fra 5-10 tusen til 1 million eller mer) naturlige polymerer, hvis molekyler er bygget av aminosyrerester forbundet med en amid (peptid)binding.
Proteiner kalles også proteiner (fra det greske "protos" - det første, viktige). Antall aminosyrerester i et proteinmolekyl varierer mye og når noen ganger flere tusen. Hvert protein har sin egen iboende sekvens av aminosyrerester.
Proteiner utfører en rekke biologiske funksjoner: katalytiske (enzymer), regulatoriske (hormoner), strukturelle (kollagen, fibroin), motoriske (myosin), transport (hemoglobin, myoglobin), beskyttende (immunoglobuliner, interferon), reservedeler (kasein, albumin, gliadin) og andre. Blant proteiner er det antibiotika og stoffer som har en giftig effekt.
Proteiner er grunnlaget for biomembraner, den viktigste delen av cellen og cellekomponentene. De spiller en nøkkelrolle i cellens liv, og danner så å si det materielle grunnlaget for dens kjemiske aktivitet.
En eksepsjonell egenskap til et protein er selvorganiseringen av strukturen, dvs. dens evne til spontant å skape en spesifikk romlig struktur som bare er særegen for et gitt protein. I hovedsak er alle aktivitetene til kroppen (utvikling, bevegelse, utførelse av ulike funksjoner og mye mer) assosiert med proteinstoffer (fig. 36). Det er umulig å forestille seg et liv uten proteiner.
Proteiner er den viktigste komponenten i mat til mennesker og dyr, leverandøren av aminosyrene de trenger.
Struktur
I den romlige strukturen til proteiner er naturen til radikalene (restene) R- i aminosyremolekyler av stor betydning. Ikke-polare aminosyreradikaler er vanligvis lokalisert inne i proteinmakromolekylet og forårsaker hydrofobe (se nedenfor) interaksjoner; polare radikaler som inneholder ionogene (ionedannende) grupper er vanligvis lokalisert på overflaten av et proteinmakromolekyl og karakteriserer elektrostatiske (ioniske) interaksjoner. Polare ikke-ioniske radikaler (for eksempel inneholdende alkohol-OH-grupper, amidgrupper) kan være lokalisert både på overflaten og inne i proteinmolekylet. De deltar i dannelsen av hydrogenbindinger.
I proteinmolekyler er a-aminosyrer forbundet med peptidbindinger (-CO-NH-):
Polypeptidkjedene konstruert på denne måten eller individuelle seksjoner i polypeptidkjeden kan i noen tilfeller i tillegg være forbundet med disulfid (-S-S-) bindinger, eller, som de ofte kalles, disulfidbroer.
En viktig rolle i å skape strukturen til proteiner spilles av ioniske (salt) og hydrogenbindinger, samt hydrofob interaksjon - en spesiell type kontakt mellom de hydrofobe komponentene til proteinmolekyler i et vandig medium. Alle disse bindingene har forskjellig styrke og gir dannelsen av et komplekst, stort proteinmolekyl.
Til tross for forskjellen i strukturen og funksjonene til proteinstoffer, svinger deres elementære sammensetning litt (i % av tørr masse): karbon - 51-53; oksygen - 21,5-23,5; nitrogen - 16,8-18,4; hydrogen - 6,5-7,3; svovel - 0,3-2,5. Noen proteiner inneholder små mengder fosfor, selen og andre grunnstoffer.
Sekvensen for å forbinde aminosyrerester i polypeptidkjeden kalles proteinets primære struktur (fig. 37).
Et proteinmolekyl kan bestå av en eller flere polypeptidkjeder, som hver inneholder et forskjellig antall aminosyrerester. Gitt antallet mulige kombinasjoner, kan det sies at variasjonen av proteiner er nesten ubegrenset, men ikke alle finnes i naturen. Det totale antallet ulike typer proteiner i alle typer levende organismer er 10 10 -10 12 . For proteiner hvis struktur er ekstremt kompleks, i tillegg til den primære, skilles det også ut høyere nivåer av strukturell organisering: sekundære, tertiære og noen ganger kvaternære strukturer (tabell 9). De fleste av proteinene har en sekundær struktur, men ikke alltid gjennom hele polypeptidkjeden. Polypeptidkjeder med en viss sekundær struktur kan ordnes annerledes i rommet.
Dette romlige arrangementet kalles tertiærstrukturen (fig. 39)
I dannelsen av den tertiære strukturen, i tillegg til hydrogenbindinger, spiller ioniske og hydrofobe interaksjoner en viktig rolle. Ved arten av "emballasjen" av proteinmolekylet, er kuleformede eller sfæriske, og fibrillære, eller filamentøse, proteiner skilt.
For kuleproteiner er a-helikal struktur mer karakteristisk, heliksene er buede, "foldet". Makromolekylet har en sfærisk form. De løses opp i vann og saltløsninger for å danne kolloidale systemer. De fleste dyre-, plante- og mikroorganismeproteiner er kuleformede proteiner. 
For fibrillære proteiner er en filamentøs struktur mer karakteristisk. De oppløses vanligvis ikke i vann. Fibrillære proteiner utfører vanligvis strukturdannende funksjoner. Egenskapene deres (styrke, evne til å strekke seg) avhenger av måten polypeptidkjedene er pakket. Et eksempel på fibrillære proteiner er muskelvevsproteiner (myosin), keratin (hornvev). I noen tilfeller danner individuelle proteinunderenheter komplekse ensembler ved hjelp av hydrogenbindinger, elektrostatiske og andre interaksjoner. I dette tilfellet dannes en kvaternær struktur av proteiner.
Imidlertid bør det bemerkes nok en gang at den primære strukturen spiller en eksepsjonell rolle i organiseringen av høyere proteinstrukturer. 
Klassifisering
Det er flere klassifiseringer av proteiner. De er basert på forskjellige egenskaper:
Vanskelighetsgrad (enkel og kompleks);
Formen på molekylene (globære og fibrillære proteiner);
Løselighet i individuelle løsningsmidler (vannløselig, løselig i fortynnede saltvannsløsninger - albuminer, alkoholløselige - prolaminer, løselig i fortynnede alkalier og syrer - gluteliner);
Funksjon utført (for eksempel lagringsproteiner, skjelett, etc.).
Eiendommer
Proteiner er amfotere elektrolytter. Ved en viss pH-verdi på mediet (det kalles det isoelektriske punktet) er antallet positive og negative ladninger i proteinmolekylet det samme. Dette er en av hovedegenskapene til protein. Proteiner på dette tidspunktet er elektrisk nøytrale, og deres løselighet i vann er lavest. Proteiners evne til å redusere løseligheten når molekylene deres blir elektrisk nøytrale brukes til å isolere dem fra løsninger, for eksempel i teknologien for å skaffe proteinprodukter.
Hydrering
Hydratiseringsprosessen betyr binding av vann av proteiner, mens de viser hydrofile egenskaper: de svulmer, deres masse og volum øker. Hevelse av proteinet er ledsaget av dets delvise oppløsning. Hydrofilisiteten til individuelle proteiner avhenger av deres struktur. De hydrofile amid- (-CO-NH-, peptidbinding), amin- (NH2) og karboksyl- (COOH)-gruppene som er tilstede i sammensetningen og lokalisert på overflaten av proteinmakromolekylet tiltrekker seg vannmolekyler, og orienterer dem strengt på overflaten av molekylet . Hydratiserings- (vann)-skallet som omgir proteinkulene forhindrer aggregering og sedimentering og bidrar følgelig til stabiliteten til proteinløsninger. Ved det isoelektriske punktet har proteiner minst evne til å binde vann, hydreringsskallet rundt proteinmolekylene blir ødelagt, så de kombineres og danner store aggregater. Aggregering av proteinmolekyler oppstår også når de dehydreres med noen organiske løsemidler, for eksempel etylalkohol. Dette fører til utfelling av proteiner. Når pH i mediet endres, blir proteinmakromolekylet ladet, og dets hydreringskapasitet endres.
Med begrenset hevelse danner konsentrerte proteinløsninger komplekse systemer kalt gelé. Geléene er ikke flytende, elastiske, har plastisitet, en viss mekanisk styrke, og er i stand til å opprettholde formen. Globulære proteiner kan hydreres fullstendig ved å løse dem opp i vann (for eksempel melkeproteiner), og danne løsninger med lav konsentrasjon. De hydrofile egenskapene til proteiner, dvs. deres evne til å svelle, danne geléer, stabilisere suspensjoner, emulsjoner og skum, er av stor betydning i biologi og næringsmiddelindustrien. En veldig mobil gelé, hovedsakelig bygget av proteinmolekyler, er cytoplasmaet - det halvflytende innholdet i cellen. Høyhydrert gelé - rå gluten isolert fra hvetedeig, den inneholder opptil 65 % vann. Den forskjellige hydrofilisiteten til glutenproteiner er et av tegnene som kjennetegner kvaliteten på hvetekorn og melet som oppnås fra det (den såkalte sterke og svake hveten). Hydrofilisiteten til korn- og melproteiner spiller en viktig rolle i lagring og bearbeiding av korn, i baking. Deigen, som fås i bakeindustrien, er et protein svellet i vann, en konsentrert gelé som inneholder stivelseskorn.
Proteindenaturering
Under denaturering, under påvirkning av eksterne faktorer (temperatur, mekanisk virkning, virkningen av kjemiske midler og en rekke andre faktorer), skjer det en endring i de sekundære, tertiære og kvaternære strukturene til proteinmakromolekylet, dvs. dets opprinnelige romlig struktur. Den primære strukturen og følgelig den kjemiske sammensetningen av proteinet endres ikke. Fysiske egenskaper endres: løseligheten avtar, evnen til å hydrere, biologisk aktivitet går tapt. Formen på proteinmakromolekylet endres, aggregering skjer. Samtidig øker aktiviteten til noen kjemiske grupper, effekten av proteolytiske enzymer på proteiner lettes, og følgelig hydrolyseres den lettere.
I matteknologi er termisk denaturering av proteiner av spesiell praktisk betydning, graden av denne avhenger av temperatur, varighet av oppvarming og fuktighet. Dette må huskes når man utvikler moduser for varmebehandling av matråvarer, halvfabrikata og noen ganger ferdige produkter. Prosessene med termisk denaturering spiller en spesiell rolle i å blanchere vegetabilske råvarer, tørke korn, bake brød og skaffe pasta. Proteindenaturering kan også være forårsaket av mekanisk påvirkning (trykk, gnidning, risting, ultralyd). Til slutt fører virkningen av kjemiske reagenser (syrer, alkalier, alkohol, aceton) til denaturering av proteiner. Alle disse teknikkene er mye brukt i mat og bioteknologi.
Skummer
Prosessen med skumming forstås som proteiners evne til å danne svært konsentrerte væske-gasssystemer, kalt skum. Stabiliteten til skummet, der proteinet er et esemiddel, avhenger ikke bare av dets natur og konsentrasjon, men også av temperatur. Proteiner som skummende midler er mye brukt i konfektindustrien (marshmallow, marshmallow, soufflé). Skummets struktur har brød, og dette påvirker smaksegenskapene.
Proteinmolekyler under påvirkning av en rekke faktorer kan ødelegges eller samhandle med andre stoffer for å danne nye produkter. For næringsmiddelindustrien kan to svært viktige prosesser skilles: 1) hydrolyse av proteiner under påvirkning av enzymer og 2) interaksjon av aminogrupper av proteiner eller aminosyrer med karbonylgrupper av reduserende sukker. Under påvirkning av proteaser - enzymer som katalyserer den hydrolytiske spaltningen av proteiner, brytes sistnevnte ned til enklere produkter (poly- og dipeptider) og til slutt til aminosyrer. Hastigheten av proteinhydrolyse avhenger av sammensetningen, molekylstrukturen, enzymaktiviteten og forholdene.
Proteinhydrolyse
Hydrolysereaksjonen med dannelse av aminosyrer i generelle termer kan skrives som følger: 
Forbrenning
4. Hvilke reaksjoner kan brukes for å gjenkjenne proteiner?
5. Hvilken rolle spiller proteiner i organismenes liv?
6. Husk fra det generelle biologikurset hvilke proteiner som bestemmer organismenes immunegenskaper.
7. Fortell oss om AIDS og forebygging av denne forferdelige sykdommen.
8. Hvordan gjenkjenne et produkt laget av naturlig ull og kunstige fibre?
9. Skriv reaksjonsligningen for hydrolyse av proteiner med den generelle formelen (-NH-CH-CO-) n.
l
R
Hva er betydningen av denne prosessen i biologi og hvordan brukes den i industrien?
10. Skriv reaksjonsligninger som kan brukes til å lage følgende overganger: etan -> etylalkohol -> eddikaldehyd -> eddiksyre -> kloreddiksyre -> aminoeddiksyre -> polypeptid.
kjemisaker, oppgaver og løsninger, leksjonsnotater
PROTEINER (proteiner), en klasse av komplekse nitrogenholdige forbindelser, de mest karakteristiske og viktige (sammen med nukleinsyrer) komponentene i levende materie. Proteiner utfører mange og varierte funksjoner. De fleste proteiner er enzymer som katalyserer kjemiske reaksjoner. Mange hormoner som regulerer fysiologiske prosesser er også proteiner. Strukturelle proteiner som kollagen og keratin er hovedkomponentene i beinvev, hår og negler. De kontraktile proteinene i muskler har evnen til å endre lengden ved å bruke kjemisk energi til å utføre mekanisk arbeid. Proteiner er antistoffer som binder og nøytraliserer giftige stoffer. Noen proteiner som kan reagere på ytre påvirkninger (lys, lukt) fungerer som reseptorer i sanseorganene som oppfatter irritasjon. Mange proteiner lokalisert inne i cellen og på cellemembranen utfører regulerende funksjoner.I første halvdel av 1800-tallet mange kjemikere, og blant dem først og fremst J. von Liebig, kom etter hvert til den konklusjon at proteiner er en spesiell klasse nitrogenholdige forbindelser. Navnet "proteiner" (fra gresk.
protoer den første) ble foreslått i 1840 av den nederlandske kjemikeren G. Mulder. FYSISKE EGENSKAPER Proteiner er hvite i fast tilstand, men fargeløse i løsning, med mindre de har en kromoforgruppe (farget), som for eksempel hemoglobin. Løseligheten i vann av ulike proteiner varierer sterkt. Det varierer også med pH og med konsentrasjonen av salter i løsningen, slik at man kan velge forholdene under hvilke ett protein selektivt vil utfelles i nærvær av andre proteiner. Denne "utsaltingsmetoden" er mye brukt for å isolere og rense proteiner. Det rensede proteinet feller ofte ut av løsningen som krystaller.Sammenlignet med andre forbindelser er molekylvekten til proteiner veldig stor - fra flere tusen til mange millioner dalton. Derfor, under ultrasentrifugering, blir proteiner utfelt, og dessuten med forskjellige hastigheter. På grunn av tilstedeværelsen av positivt og negativt ladede grupper i proteinmolekyler, beveger de seg med forskjellige hastigheter i et elektrisk felt. Dette er grunnlaget for elektroforese, en metode som brukes til å isolere individuelle proteiner fra komplekse blandinger. Rensing av proteiner utføres også ved kromatografi.
KJEMISKE EGENSKAPER Struktur. Proteiner er polymerer, dvs. molekyler bygget som kjeder fra repeterende monomere enheter, eller underenheter, rollen som de spiller en -aminosyrer. Generell formel for aminosyrer hvor R et hydrogenatom eller en organisk gruppe.Et proteinmolekyl (polypeptidkjede) kan bestå av bare et relativt lite antall aminosyrer eller flere tusen monomerenheter. Sammenkoblingen av aminosyrer i kjeden er mulig fordi hver av dem har to forskjellige kjemiske grupper: en aminogruppe med grunnleggende egenskaper,
NH2 og en sur karboksylgruppe, COOH. Begge disse gruppene er tilknyttet en - et karbonatom. Karboksylgruppen til en aminosyre kan danne en amid(peptid)binding med aminogruppen til en annen aminosyre: Etter at to aminosyrer er koblet sammen på denne måten, kan kjeden forlenges ved å legge en tredje til den andre aminosyren, og så videre. Som man kan se fra ligningen ovenfor, frigjøres et vannmolekyl når en peptidbinding dannes. I nærvær av syrer, alkalier eller proteolytiske enzymer, fortsetter reaksjonen i motsatt retning: polypeptidkjeden spaltes til aminosyrer med tilsetning av vann. Denne reaksjonen kalles hydrolyse. Hydrolyse går spontant, og energi er nødvendig for å kombinere aminosyrer til en polypeptidkjede.
Etter at to aminosyrer er koblet sammen på denne måten, kan kjeden forlenges ved å legge en tredje til den andre aminosyren, og så videre. Som man kan se fra ligningen ovenfor, frigjøres et vannmolekyl når en peptidbinding dannes. I nærvær av syrer, alkalier eller proteolytiske enzymer, fortsetter reaksjonen i motsatt retning: polypeptidkjeden spaltes til aminosyrer med tilsetning av vann. Denne reaksjonen kalles hydrolyse. Hydrolyse går spontant, og energi er nødvendig for å kombinere aminosyrer til en polypeptidkjede. En karboksylgruppe og en amidgruppe (eller en lignende imidgruppe når det gjelder aminosyren prolin) er tilstede i alle aminosyrer, men forskjellene mellom aminosyrer bestemmes av arten til den gruppen, eller "sidekjeden", som er angitt ovenfor med bokstaven
R . Rollen til sidekjeden kan spilles av ett hydrogenatom, som i aminosyren glycin, eller av en eller annen voluminøs gruppe, som i histidin og tryptofan. Noen sidekjeder er kjemisk inerte, mens andre er svært reaktive.Mange tusen forskjellige aminosyrer kan syntetiseres, og mange forskjellige aminosyrer forekommer i naturen, men bare 20 typer aminosyrer brukes til proteinsyntese: alanin, arginin, asparagin, asparaginsyre, valin, histidin, glycin, glutamin, glutamin. syre, isoleucin, leucin, lysin, metionin, prolin, serin, tyrosin, treonin, tryptofan, fenylalanin og cystein (i proteiner kan cystein være tilstede som en dimer
cystin). Riktignok er det i noen proteiner andre aminosyrer i tillegg til de tjue som regelmessig forekommer, men de dannes som et resultat av modifikasjon av noen av de tjue som er oppført etter at det har blitt inkludert i proteinet.optisk aktivitet. Alle aminosyrer, med unntak av glycin, en Karbonatomet har fire forskjellige grupper festet. Når det gjelder geometri, kan fire forskjellige grupper festes på to måter, og følgelig er det to mulige konfigurasjoner, eller to isomerer, relatert til hverandre som et objekt til dets speilbilde, dvs. som venstre hånd til høyre. Én konfigurasjon kalles venstre- eller venstrehendt ( L ), og den andre høyre, eller høyredreiende ( D ), siden to slike isomerer er forskjellige i rotasjonsretningen til planet av polarisert lys. Finnes bare i proteiner L -aminosyrer (unntaket er glycin; det kan bare representeres i én form, siden to av de fire gruppene er like), og de har alle optisk aktivitet (siden det bare er en isomer). D -aminosyrer er sjeldne i naturen; de finnes i noen antibiotika og celleveggen til bakterier.Rekkefølgen av aminosyrer. Aminosyrer i polypeptidkjeden er ikke ordnet tilfeldig, men i en viss fast rekkefølge, og det er denne rekkefølgen som bestemmer funksjonene og egenskapene til proteinet. Ved å variere rekkefølgen på de 20 typene aminosyrer, kan du få et stort antall forskjellige proteiner, akkurat som du kan lage mange forskjellige tekster fra bokstavene i alfabetet.Tidligere tok det ofte flere år å bestemme aminosyresekvensen til et protein. Direkte bestemmelse er fortsatt en ganske møysommelig oppgave, selv om det er opprettet enheter som lar den utføres automatisk. Det er vanligvis lettere å bestemme nukleotidsekvensen til det tilsvarende genet og utlede aminosyresekvensen til proteinet fra det. Til dags dato er aminosyresekvensene til mange hundre proteiner allerede bestemt. Funksjonene til dekodede proteiner er vanligvis kjent, og dette hjelper til med å forestille seg de mulige funksjonene til lignende proteiner dannet, for eksempel i ondartede neoplasmer.
Komplekse proteiner. Proteiner som kun består av aminosyrer kalles enkle. Ofte er imidlertid et metallatom eller en kjemisk forbindelse som ikke er en aminosyre festet til polypeptidkjeden. Slike proteiner kalles komplekse. Et eksempel er hemoglobin: det inneholder jernporfyrin, som gir det sin røde farge og lar det fungere som en oksygenbærer.Navnene på de fleste komplekse proteiner inneholder en indikasjon på arten av de vedlagte gruppene: sukker er tilstede i glykoproteiner, fett i lipoproteiner. Hvis den katalytiske aktiviteten til enzymet avhenger av den vedlagte gruppen, kalles det en protesegruppe. Noen vitaminer spiller ofte rollen som en protesegruppe eller er en del av den. Vitamin A, for eksempel festet til et av proteinene i netthinnen, bestemmer følsomheten for lys.
Tertiær struktur. Det som er viktig er ikke så mye aminosyresekvensen til proteinet (primærstruktur), men måten det er lagt i rommet. Langs hele lengden av polypeptidkjeden danner hydrogenioner vanlige hydrogenbindinger, som gir den formen av en spiral eller et lag (sekundær struktur). Fra kombinasjonen av slike helikser og lag oppstår en kompakt form av neste rekkefølge - den tertiære strukturen til proteinet. Rundt bindingene som holder de monomere leddene i kjeden, er rotasjoner gjennom små vinkler mulig. Derfor, fra et rent geometrisk synspunkt, er antallet mulige konfigurasjoner for enhver polypeptidkjede uendelig stort. I virkeligheten eksisterer hvert protein normalt i bare én konfigurasjon, bestemt av dets aminosyresekvens. Denne strukturen er ikke stiv, det er som det var, « puster” svinger rundt en viss gjennomsnittlig konfigurasjon. Kjedet brettes inn i en konfigurasjon der den frie energien (evnen til å utføre arbeid) er minimal, akkurat som en frigjort fjær bare komprimeres til en tilstand som tilsvarer et minimum av fri energi. Ofte er en del av kjeden stivt knyttet til en annen disulfid ( SS) bindinger mellom to cysteinrester. Dette er delvis grunnen til at cystein blant aminosyrer spiller en spesielt viktig rolle.Kompleksiteten i strukturen til proteiner er så stor at det ennå ikke er mulig å beregne tertiærstrukturen til et protein, selv om dets aminosyresekvens er kjent. Men hvis det er mulig å oppnå proteinkrystaller, kan dens tertiære struktur bestemmes ved røntgendiffraksjon.
I strukturelle, kontraktile og enkelte andre proteiner er kjedene forlengede og flere lett foldede kjeder som ligger side om side danner fibriller; fibriller, i sin tur, foldes til større formasjoner fibre. Imidlertid er de fleste proteiner i løsning kuleformede: kjedene er kveilet i en kule, som garn i en ball. Fri energi med denne konfigurasjonen er minimal, siden hydrofobe ("vannavstøtende") aminosyrer er skjult inne i kulen, og hydrofile ("vanntiltrekkende") aminosyrer er på overflaten.
Mange proteiner er komplekser av flere polypeptidkjeder. Denne strukturen kalles proteinets kvaternære struktur. Hemoglobinmolekylet, for eksempel, består av fire underenheter, som hver er et kuleprotein.
Strukturelle proteiner på grunn av deres lineære konfigurasjon danner fibre der strekkstyrken er svært høy, mens den kuleformede konfigurasjonen tillater proteiner å inngå spesifikke interaksjoner med andre forbindelser. På overflaten av kulen, med riktig legging av kjeder, vises hulrom av en viss form, der reaktive kjemiske grupper er lokalisert. Hvis et gitt protein er et enzym, kommer et annet, vanligvis mindre, molekyl av et eller annet stoff inn i et slikt hulrom, akkurat som en nøkkel går inn i en lås; i dette tilfellet endres konfigurasjonen av elektronskyen til molekylet under påvirkning av kjemiske grupper lokalisert i hulrommet, og dette tvinger det til å reagere på en bestemt måte. På denne måten katalyserer enzymet reaksjonen. Antistoffmolekyler har også hulrom der ulike fremmedstoffer binder seg og dermed ufarliggjøres. «Nøkkel og lås»-modellen, som forklarer interaksjonen mellom proteiner og andre forbindelser, gjør det mulig å forstå spesifisiteten til enzymer og antistoffer, d.v.s. deres evne til å reagere bare med visse forbindelser.
Proteiner i ulike typer organismer. Proteiner som utfører samme funksjon i forskjellige plante- og dyrearter og derfor bærer samme navn, har også en lignende konfigurasjon. De skiller seg imidlertid noe i aminosyresekvensen. Ettersom arter avviker fra en felles stamfar, erstattes noen aminosyrer i visse posisjoner av mutasjoner med andre. Skadelige mutasjoner som forårsaker arvelige sykdommer forkastes ved naturlig utvalg, men gunstige eller i det minste nøytrale kan bevares. Jo nærmere to biologiske arter er hverandre, jo mindre forskjeller finnes i proteinene deres.Noen proteiner endres relativt raskt, andre er ganske konservative. Sistnevnte inkluderer for eksempel cytokrom Med et respirasjonsenzym som finnes i de fleste levende organismer. Hos mennesker og sjimpanser er aminosyresekvensene identiske, og i cytokrom Med hvete, bare 38 % av aminosyrene viste seg å være forskjellige. Selv sammenligne mennesker og bakterier, likheten mellom cytokromer Med(forskjellene påvirker 65 % av aminosyrene her) kan fortsatt sees, selv om den felles stamfaren til bakterier og mennesker levde på jorden for rundt to milliarder år siden. I dag brukes ofte sammenligning av aminosyresekvenser for å bygge et fylogenetisk (genealogisk) tre som gjenspeiler de evolusjonære forholdene mellom ulike organismer.
Denaturering. Det syntetiserte proteinmolekylet, folding, får sin egen konfigurasjon. Denne konfigurasjonen kan imidlertid ødelegges ved oppvarming, ved å endre pH, ved påvirkning av organiske løsningsmidler, og til og med ved ganske enkelt å agitere løsningen til det kommer bobler på overflaten. Et protein endret på denne måten kalles denaturert; den mister sin biologiske aktivitet og blir vanligvis uløselig. Velkjente eksempler på denaturert protein er kokte egg eller pisket krem. Små proteiner, som bare inneholder rundt hundre aminosyrer, er i stand til å renaturere, dvs. skaffe tilbake den opprinnelige konfigurasjonen. Men de fleste av proteinene blir ganske enkelt transformert til en masse sammenfiltrede polypeptidkjeder og gjenoppretter ikke deres tidligere konfigurasjon.En av hovedvanskene med å isolere aktive proteiner er deres ekstreme følsomhet for denaturering. Denne egenskapen til proteiner finner nyttig anvendelse i konservering av matprodukter: høy temperatur denaturerer irreversibelt enzymene til mikroorganismer, og mikroorganismene dør.
PROTEIN SYNTESE For proteinsyntese må en levende organisme ha et system av enzymer som er i stand til å feste en aminosyre til en annen. En informasjonskilde er også nødvendig som kan bestemme hvilke aminosyrer som skal kobles sammen. Siden det finnes tusenvis av typer proteiner i kroppen, og hver av dem består av i gjennomsnitt flere hundre aminosyrer, må informasjonen som kreves være virkelig enorm. Det lagres (i likhet med hvordan en post lagres på et magnetbånd) i nukleinsyremolekylene som utgjør gener. Cm . også ARVLIGHET; NUKLEINSYRER.Enzymaktivering. En polypeptidkjede syntetisert fra aminosyrer er ikke alltid et protein i sin endelige form. Mange enzymer syntetiseres først som inaktive forløpere og blir aktive først etter at et annet enzym fjerner noen få aminosyrer fra den ene enden av kjeden. Noen av fordøyelsesenzymene, som trypsin, syntetiseres i denne inaktive formen; disse enzymene aktiveres i fordøyelseskanalen som et resultat av fjerning av det terminale fragmentet av kjeden. Hormonet insulin, hvis molekyl i sin aktive form består av to korte kjeder, syntetiseres i form av en enkelt kjede, den såkalte. proinsulin. Deretter fjernes den midtre delen av denne kjeden, og de gjenværende fragmentene binder seg til hverandre og danner det aktive hormonmolekylet. Komplekse proteiner dannes først etter at en viss kjemisk gruppe er festet til proteinet, og denne tilknytningen krever ofte også et enzym.Metabolsk sirkulasjon. Etter å ha fôret et dyr med aminosyrer merket med radioaktive isotoper av karbon, nitrogen eller hydrogen, blir merket raskt inkorporert i proteinene. Hvis merkede aminosyrer slutter å komme inn i kroppen, begynner mengden etikett i proteiner å avta. Disse eksperimentene viser at de resulterende proteinene ikke lagres i kroppen før livet er slutt. Alle av dem, med noen få unntak, er i en dynamisk tilstand, dekomponerer konstant til aminosyrer og syntetiseres deretter på nytt.Noen proteiner brytes ned når cellene dør og blir ødelagt. Dette skjer hele tiden, for eksempel med røde blodceller og epitelceller langs den indre overflaten av tarmen. I tillegg skjer nedbrytning og resyntese av proteiner også i levende celler. Merkelig nok er mindre kjent om nedbrytningen av proteiner enn om deres syntese. Det som derimot er klart er at proteolytiske enzymer er involvert i nedbrytningen, lik de som bryter ned proteiner til aminosyrer i fordøyelseskanalen.
Halveringstiden til ulike proteiner er forskjellig fra flere timer til mange måneder. Det eneste unntaket er kollagenmolekylet. Når de er dannet, forblir de stabile og blir ikke fornyet eller erstattet. Over tid endres imidlertid noen av egenskapene deres, spesielt elastisiteten, og siden de ikke fornyes, er visse aldersrelaterte endringer et resultat av dette, for eksempel utseendet av rynker på huden.
syntetiske proteiner. Kjemikere har for lengst lært hvordan man polymeriserer aminosyrer, men aminosyrene kombineres tilfeldig, slik at produktene fra en slik polymerisering har liten likhet med naturlige. Riktignok er det mulig å kombinere aminosyrer i en gitt rekkefølge, noe som gjør det mulig å oppnå noen biologisk aktive proteiner, spesielt insulin. Prosessen er ganske komplisert, og på denne måten er det mulig å oppnå bare de proteinene hvis molekyler inneholder omtrent hundre aminosyrer. Det er å foretrekke i stedet å syntetisere eller isolere nukleotidsekvensen til et gen som tilsvarer den ønskede aminosyresekvensen, og deretter introdusere dette genet i en bakterie, som ved replikasjon vil produsere en stor mengde av det ønskede produktet. Denne metoden har imidlertid også sine ulemper. Cm . Se også GENETEKNIKK. PROTEINER OG NÆRING Når proteiner i kroppen brytes ned til aminosyrer, kan disse aminosyrene gjenbrukes til proteinsyntese. Samtidig er selve aminosyrene utsatt for forfall, slik at de ikke utnyttes fullt ut. Det er også klart at under vekst, graviditet og sårheling må proteinsyntesen overstige nedbrytningen. Kroppen mister kontinuerlig noen proteiner; disse er proteinene i hår, negler og overflatelaget på huden. Derfor, for syntese av proteiner, må hver organisme motta aminosyrer fra mat. Grønne planter syntetiseres fra CO 2 , vann og ammoniakk eller nitrater er alle 20 aminosyrer som finnes i proteiner. Mange bakterier er også i stand til å syntetisere aminosyrer i nærvær av sukker (eller noe tilsvarende) og fiksert nitrogen, men sukker leveres til slutt av grønne planter. Hos dyr er evnen til å syntetisere aminosyrer begrenset; de får aminosyrer ved å spise grønne planter eller andre dyr. I fordøyelseskanalen brytes de absorberte proteinene ned til aminosyrer, sistnevnte absorberes, og proteinene som er karakteristiske for den gitte organismen bygges av dem. Ingen av det absorberte proteinet er inkorporert i kroppsstrukturer som sådan. Det eneste unntaket er at hos mange pattedyr kan en del av mors antistoffer passere intakt gjennom morkaken inn i fosterets sirkulasjon, og gjennom morsmelk (spesielt hos drøvtyggere) overføres til den nyfødte umiddelbart etter fødselen.Behov for proteiner. Det er klart at for å opprettholde livet må kroppen få en viss mengde protein fra maten. Størrelsen på dette behovet avhenger imidlertid av en rekke faktorer. Kroppen trenger mat både som energikilde (kalorier) og som materiale for å bygge strukturene. I første omgang er behovet for energi. Dette betyr at når det er lite karbohydrater og fett i kosten, brukes diettproteiner ikke til syntese av deres egne proteiner, men som en kilde til kalorier. Ved langvarig faste brukes til og med dine egne proteiner for å dekke energibehovet. Hvis det er nok karbohydrater i kosten, kan proteininntaket reduseres.nitrogenbalanse. I gjennomsnitt ca. 16 % av den totale proteinmassen er nitrogen. Når aminosyrene som utgjør proteiner brytes ned, skilles nitrogenet som finnes i dem ut fra kroppen i urinen og (i mindre grad) i avføringen i form av ulike nitrogenholdige forbindelser. Derfor er det praktisk å bruke en slik indikator som nitrogenbalanse for å vurdere kvaliteten på proteinernæring, dvs. forskjellen (i gram) mellom mengden nitrogen som tas inn i kroppen og mengden nitrogen som skilles ut per dag. Med normal ernæring hos en voksen er disse mengdene like. I en voksende organisme er mengden av utskilt nitrogen mindre enn mengden innkommende, dvs. balansen er positiv. Ved mangel på protein i kosten er balansen negativ. Hvis det er nok kalorier i kosten, men proteinene er helt fraværende i det, sparer kroppen proteiner. Samtidig bremses proteinmetabolismen, og gjenbruken av aminosyrer i proteinsyntesen går så effektivt som mulig. Tap er imidlertid uunngåelig, og nitrogenholdige forbindelser skilles fortsatt ut i urinen og delvis i avføringen. Mengden nitrogen som skilles ut fra kroppen per dag under proteinsult kan tjene som et mål på den daglige mangelen på protein. Det er naturlig å anta at ved å innføre i kosten en mengde protein tilsvarende denne mangelen, er det mulig å gjenopprette nitrogenbalansen. Det er det imidlertid ikke. Etter å ha mottatt denne mengden protein, begynner kroppen å bruke aminosyrer mindre effektivt, så noe ekstra protein er nødvendig for å gjenopprette nitrogenbalansen.Hvis mengden protein i kosten overstiger det som er nødvendig for å opprettholde nitrogenbalansen, ser det ut til at det ikke er noen skade av dette. Overflødige aminosyrer brukes ganske enkelt som en energikilde. Et spesielt slående eksempel er eskimoen, som inntar lite karbohydrater og omtrent ti ganger mer protein enn det som kreves for å opprettholde nitrogenbalansen. I de fleste tilfeller er det imidlertid ikke gunstig å bruke protein som energikilde, siden du kan få i deg mange flere kalorier fra en gitt mengde karbohydrater enn fra samme mengde protein. I fattige land får befolkningen de nødvendige kaloriene fra karbohydrater og inntar en minimumsmengde protein.
Hvis kroppen får det nødvendige antall kalorier i form av ikke-proteinprodukter, er minimumsmengden protein som opprettholder nitrogenbalansen ca. 30 g per dag. Omtrent like mye protein finnes i fire brødskiver eller 0,5 liter melk. En litt større mengde anses vanligvis som optimal; anbefalt fra 50 til 70 g.
Essensielle aminosyrer. Til nå har protein blitt sett på som en helhet. I mellomtiden, for at proteinsyntese skal finne sted, må alle nødvendige aminosyrer være tilstede i kroppen. Noen av aminosyrene er kroppen til dyret selv i stand til å syntetisere. De kalles utskiftbare fordi de ikke trenger å være tilstede i kosten, det er bare viktig at generelt sett er inntaket av protein som nitrogenkilde tilstrekkelig; da, med mangel på ikke-essensielle aminosyrer, kan kroppen syntetisere dem på bekostning av de som er tilstede i overkant. De gjenværende "essensielle" aminosyrene kan ikke syntetiseres og må inntas sammen med mat. Viktige for mennesker er valin, leucin, isoleucin, treonin, metionin, fenylalanin, tryptofan, histidin, lysin og arginin. (Selv om arginin kan syntetiseres i kroppen, anses det som en essensiell aminosyre fordi nyfødte og voksende barn produserer utilstrekkelige mengder av det. På den annen side, for en person i moden alder, inntak av noen av disse aminosyrene fra mat kan bli valgfritt.)Denne listen over essensielle aminosyrer er omtrent den samme hos andre virveldyr og til og med hos insekter. Ernæringsverdien til proteiner bestemmes vanligvis ved å mate dem til voksende rotter og overvåke vektøkningen til dyrene.
Den ernæringsmessige verdien av proteiner. Næringsverdien til et protein bestemmes av den essensielle aminosyren som er mest mangelfull. La oss illustrere dette med et eksempel. Proteinene i kroppen vår inneholder i gjennomsnitt ca. 2 % tryptofan (i vekt). La oss si at kostholdet inneholder 10 g protein som inneholder 1% tryptofan, og at det er nok andre essensielle aminosyrer i den. I vårt tilfelle tilsvarer 10 g av dette defekte proteinet i hovedsak 5 g av et komplett; de resterende 5 g kan bare tjene som en energikilde. Merk at siden aminosyrer praktisk talt ikke lagres i kroppen, og for at proteinsyntese skal finne sted, må alle aminosyrer være tilstede samtidig, kan effekten av inntak av essensielle aminosyrer bare oppdages hvis de alle kommer inn i kroppen på samme tid.. Den gjennomsnittlige sammensetningen av de fleste animalske proteiner er nær den gjennomsnittlige sammensetningen av proteinene i menneskekroppen, så vi vil neppe møte aminosyremangel hvis kostholdet vårt er rikt på mat som kjøtt, egg, melk og ost. Imidlertid er det proteiner, for eksempel gelatin (et produkt av kollagen denaturering), som inneholder svært få essensielle aminosyrer. Vegetabilske proteiner, selv om de er bedre enn gelatin i denne forstand, er også fattige på essensielle aminosyrer; spesielt lite i dem lysin og tryptofan. Likevel er et rent vegetarisk kosthold slett ikke skadelig, med mindre det forbruker en litt større mengde vegetabilske proteiner, tilstrekkelig til å gi kroppen essensielle aminosyrer. Mest protein finnes i planter i frøene, spesielt i frøene til hvete og ulike belgfrukter. Unge skudd, som asparges, er også rike på protein.Syntetiske proteiner i kosten. Ved å tilsette små mengder syntetiske essensielle aminosyrer eller proteiner rike på dem til ufullstendige proteiner, som for eksempel maisproteiner, er det mulig å øke næringsverdien til sistnevnte betydelig, dvs. og dermed øke mengden protein som konsumeres. En annen mulighet er å dyrke bakterier eller gjær på petroleumshydrokarboner med tilsetning av nitrater eller ammoniakk som nitrogenkilde. Det mikrobielle proteinet oppnådd på denne måten kan tjene som fôr til fjørfe eller husdyr, eller kan konsumeres direkte av mennesker. Den tredje, mye brukte metoden bruker drøvtyggeres fysiologi. Hos drøvtyggere, i den innledende delen av magen, den såkalte. Vom er bebodd av spesielle former for bakterier og protozoer som omdanner defekte planteproteiner til mer komplette mikrobielle proteiner, og disse blir igjen etter fordøyelse og absorpsjon til animalske proteiner. Urea, en billig syntetisk nitrogenholdig forbindelse, kan tilsettes husdyrfôr. Mikroorganismer som lever i vomma bruker ureanitrogen til å omdanne karbohydrater (som det er mye mer av i fôret) til protein. Omtrent en tredjedel av alt nitrogen i husdyrfôr kan komme i form av urea, som i hovedsak betyr til en viss grad kjemisk proteinsyntese. I USA spiller denne metoden en viktig rolle som en av måtene å få protein på.LITTERATUR Murray R, Grenner D, Meyes P, Rodwell W. menneskelig biokjemi, tt. 12. M., 1993Alberts B., Bray D., Lewis J. et al. Molekylærbiologi av cellen, tt. 1. 3. M., 1994
Blant organisk materiale ekorn, eller proteiner, er de mest tallrike, mest mangfoldige og av største betydning biopolymerer. De står for 50 - 80% tørrvekt av cellen.
Proteinmolekyler er store, og det er derfor de kalles makromolekyler. I tillegg til karbon, oksygen, hydrogen og nitrogen kan proteiner inneholde svovel, fosfor og jern. Proteiner skiller seg fra hverandre i antall (fra hundre til flere tusen), sammensetning og sekvens av monomerer. Proteinmonomerer er aminosyrer (fig. 1)
Et uendelig utvalg av proteiner skapes av forskjellige kombinasjoner av alt 20 aminosyrer. Hver aminosyre har sitt eget navn, spesielle struktur og egenskaper. Deres generelle formel kan representeres som følger:

Et aminosyremolekyl består av to identiske deler for alle aminosyrer, hvorav den ene er en aminogruppe ( -NH2) med grunnleggende egenskaper, den andre med en karboksylgruppe ( -COOH) med sure egenskaper. Den delen av molekylet som kalles radikalet ( R), forskjellige aminosyrer har forskjellige strukturer. Tilstedeværelsen av basiske og sure grupper i ett aminosyremolekyl bestemmer deres høye reaktivitet. gjennom disse gruppene kobles aminosyrer sammen for å danne et protein. I dette tilfellet dukker det opp et vannmolekyl, og de frigjorte elektronene danner en peptidbinding. Det er derfor proteiner kalles polypeptider.
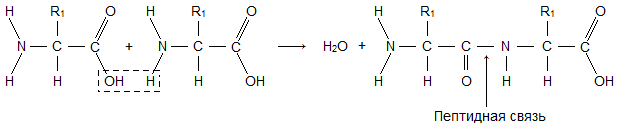
Proteinmolekyler kan ha forskjellige romlige konfigurasjoner, og fire nivåer av strukturell organisering skiller seg ut i deres struktur.
Sekvensen av aminosyrer i en polypeptidkjede er primær struktur ekorn. Det er unikt for ethvert protein og bestemmer dets form, egenskaper og funksjoner.
De fleste proteiner er spiralformet som et resultat av dannelsen av hydrogenbindinger mellom -CO- og -NH- grupper av forskjellige aminosyrerester i polypeptidkjeden. Hydrogenbindinger er svake, men i kombinasjon gir de en ganske sterk struktur. Denne spiralen er sekundær struktur ekorn.
Tertiær struktur- tredimensjonal romlig "pakking" av polypeptidkjeden. Som et resultat oppstår en bisarr, men spesifikk konfigurasjon for hvert protein - kule. Styrken til den tertiære strukturen er gitt av ulike bindinger som oppstår mellom aminosyreradikaler.
Kvartær struktur ikke typisk for alle proteiner. Det oppstår som et resultat av kombinasjonen av flere makromolekyler med en tertiær struktur til et komplekst kompleks. For eksempel er humant blodhemoglobin et kompleks av fire proteinmakromolekyler.
Denne kompleksiteten til strukturen til proteinmolekyler er assosiert med en rekke funksjoner som er iboende i disse biopolymerene.
Krenkelse av den naturlige strukturen til proteinet kalles denaturering. Det kan oppstå under påvirkning av temperatur, kjemikalier, strålingsenergi og andre faktorer. Med en svak innvirkning desintegrerer bare den kvartære strukturen, med en sterkere, den tertiære, og deretter den sekundære, og proteinet forblir i form av en polypeptidkjede.
Denne prosessen er delvis reversibel: hvis den primære strukturen ikke er forstyrret, er det denaturerte proteinet i stand til å gjenopprette strukturen. Det følger at alle trekk ved strukturen til et proteinmakromolekyl bestemmes av dets primære struktur.
Unntatt enkle proteiner, som bare består av aminosyrer, er det også komplekse proteiner
Ekorner høymolekylære organiske forbindelser bygget av 20 aminosyrerester. I henhold til deres struktur tilhører de polymerer. Molekylene deres er i form av lange kjeder som består av repeterende molekyler - monomerer. For å danne et polymermolekyl må hver av monomerene ha minst to reaktive bindinger med andre monomerer.
Proteinet ligner i strukturen på polymer nylon: begge polymerene er en kjede av monomerer. Men det er en betydelig forskjell mellom dem. Nylon består av to typer monomerer, mens protein består av 20 forskjellige monomerer kalt aminosyrer. Avhengig av rekkefølgen av veksling av monomerer, dannes mange forskjellige typer proteiner.
Den generelle formelen for aminosyrene som utgjør et protein er:
Denne formelen viser at fire forskjellige grupper er knyttet til det sentrale karbonatomet. Tre av dem - hydrogenatomet H, den alkaliske aminogruppen H N og karboksylgruppen COOH - er like for alle aminosyrer. I henhold til sammensetningen og strukturen til den fjerde gruppen, utpekt
R aminosyrer er forskjellige fra hverandre. I de enkleste tilfellene, i et glyserolmolekyl - en slik gruppe er et hydrogenatom, i et alaninmolekyl - CH, etc.Kjemisk binding (- CO -
NH -), som forbinder aminogruppen til en aminosyre med karboksylgruppen til en annen i proteinmolekyler, kalles peptidbinding(se fig.7.5).
Alle aktive organismer, enten det er planter, dyr, bakterier eller virus, inneholder proteiner bygget av de samme aminosyrene. Derfor inneholder enhver type mat de samme aminosyrene som er en del av proteinene til organismer som spiser mat.
Definisjonen "proteiner er polymerer bygget av 20 forskjellige aminosyrer" inneholder en ufullstendig karakterisering av proteiner. Under laboratorieforhold er det ikke vanskelig å oppnå peptidbindinger i en løsning av aminosyrer og dermed danne lange molekylkjeder. Imidlertid vil arrangementet av aminosyrer i slike kjeder være kaotisk, og de resulterende molekylene vil avvike fra hverandre. Samtidig, i hvert av de naturlige proteinene, er arrangementet av individuelle typer aminosyrer alltid det samme. Og dette betyr at under proteinsyntese i et levende system brukes informasjon, i samsvar med hvilken en veldefinert sekvens av aminosyrer dannes for hvert protein.
Sekvensen av aminosyrer i et protein bestemmer dets romlige struktur. De fleste proteiner fungerer som katalysatorer. I deres romlige struktur er det aktive sentre i form av fordypninger med en veldefinert form. Molekyler, hvis transformasjon er katalysert av dette proteinet, kommer inn i slike sentre. Proteinet, som i dette tilfellet fungerer som et enzym, kan bare katalysere reaksjonen hvis formen på det transformerende molekylet og det aktive senteret faller sammen. Dette bestemmer den høye selektiviteten til protein-enzymet.
Det aktive senteret til et enzym kan dannes som et resultat av foldingen av deler av proteinkjeden som er svært fjernt fra hverandre. Derfor kan substitusjon av en aminosyre med en annen, selv i liten avstand fra det aktive stedet, enten påvirke selektiviteten til enzymet eller fullstendig ødelegge stedet. Ved å lage forskjellige sekvenser av aminosyrer, kan du få et bredt utvalg av aktive sentre. Dette er en av de viktigste egenskapene til proteiner som fungerer som enzymer.
Innholdet i artikkelen
PROTEINER (artikkel 1)- en klasse av biologiske polymerer som finnes i hver levende organisme. Med deltakelse av proteiner foregår hovedprosessene som sikrer kroppens vitale aktivitet: pust, fordøyelse, muskelsammentrekning, overføring av nerveimpulser. Benvev, hud, hår, hornformasjoner av levende vesener er sammensatt av proteiner. For de fleste pattedyr skjer veksten og utviklingen av organismen på grunn av produkter som inneholder proteiner som matkomponent. Rollen til proteiner i kroppen og følgelig deres struktur er veldig mangfoldig.
Sammensetningen av proteiner.
Alle proteiner er polymerer, hvis kjeder er satt sammen av fragmenter av aminosyrer. Aminosyrer er organiske forbindelser som i sin sammensetning (i samsvar med navnet) inneholder en NH 2 aminogruppe og en organisk syre, dvs. karboksyl, COOH-gruppe. Av all mangfoldet av eksisterende aminosyrer (teoretisk sett er antallet mulige aminosyrer ubegrenset), er det bare de som har bare ett karbonatom mellom aminogruppen og karboksylgruppen som deltar i dannelsen av proteiner. Generelt kan aminosyrene som er involvert i dannelsen av proteiner representeres med formelen: H2N–CH(R)–COOH. R-gruppen festet til karbonatomet (den mellom amino- og karboksylgruppene) bestemmer forskjellen mellom aminosyrene som utgjør proteiner. Denne gruppen kan kun bestå av karbon- og hydrogenatomer, men inneholder oftere, i tillegg til C og H, forskjellige funksjonelle (i stand til ytterligere transformasjoner) grupper, for eksempel HO-, H 2 N- osv. Det er også en alternativ når R \u003d H.
Organismer av levende vesener inneholder mer enn 100 forskjellige aminosyrer, men ikke alle brukes i konstruksjonen av proteiner, men bare 20, de såkalte "fundamental". I tabellen. 1 viser navnene deres (de fleste av navnene har utviklet seg historisk), strukturformelen, samt den mye brukte forkortelsen. Alle strukturformler er ordnet i tabellen slik at hovedfragmentet av aminosyren er til høyre.
| Navn | Struktur | Betegnelse |
| GLYSIN | GLI | |
| ALANIN | ALA | |
| VALIN | AKSEL | |
| LEUCINE | LEI | |
| ISOLEUCINE | ILE | |
| SERIN | SER | |
| THREONINE | TRE | |
| CYSTEINE | CIS | |
| METIONINE | MET | |
| LYSIN | LIZ | |
| ARGININ | ARG | |
| ASPARGESYRE | ACH | |
| ASPARAGIN | ACH | |
| GLUTAMSYRE | GLU | |
| GLUTAMIN | GLN | |
| fenylalanin | hårføner | |
| TYROSIN | TIR | |
| tryptofan | TRE | |
| HISTIDINE | GIS | |
| PROLINE | PRO | |
| I internasjonal praksis aksepteres den forkortede betegnelsen på de listede aminosyrene ved å bruke latinske tre- eller enbokstavsforkortelser, for eksempel glycin - Gly eller G, alanin - Ala eller A. | ||
Blant disse tjue aminosyrene (tabell 1) inneholder kun prolin en NH-gruppe (i stedet for NH 2) ved siden av COOH-karboksylgruppen, siden den er en del av det sykliske fragmentet.
Åtte aminosyrer (valin, leucin, isoleucin, treonin, metionin, lysin, fenylalanin og tryptofan), plassert i bordet på en grå bakgrunn, kalles essensielle, siden kroppen hele tiden må motta dem med proteinmat for normal vekst og utvikling.
Et proteinmolekyl dannes som et resultat av den sekvensielle koblingen av aminosyrer, mens karboksylgruppen til en syre interagerer med aminogruppen til nabomolekylet, som et resultat dannes en –CO–NH– peptidbinding og et vann molekylet frigjøres. På fig. 1 viser serieforbindelsen av alanin, valin og glycin.
Ris. en SERIEKOBLING AV AMINOSYRER under dannelsen av et proteinmolekyl. Veien fra den terminale aminogruppen H 2 N til den terminale karboksylgruppen COOH ble valgt som hovedretningen til polymerkjeden.
For å kompakt beskrive strukturen til et proteinmolekyl, brukes forkortelsene for aminosyrer (tabell 1, tredje kolonne) som er involvert i dannelsen av polymerkjeden. Fragmentet av molekylet vist i fig. 1 er skrevet som følger: H 2 N-ALA-VAL-GLY-COOH.
Proteinmolekyler inneholder fra 50 til 1500 aminosyrerester (kortere kjeder kalles polypeptider). Individualiteten til et protein bestemmes av settet med aminosyrer som utgjør polymerkjeden og, ikke mindre viktig, av rekkefølgen av deres veksling langs kjeden. For eksempel består insulinmolekylet av 51 aminosyrerester (det er et av de korteste kjedeproteinene) og består av to sammenkoblede parallelle kjeder med ulik lengde. Sekvensen av aminosyrefragmenter er vist i fig. 2.
Ris. 2 INSULINMOLEKYL, bygget fra 51 aminosyrerester, er fragmenter av de samme aminosyrene merket med den tilsvarende bakgrunnsfargen. Cystein-aminosyrerestene (forkortet betegnelse CIS) i kjeden danner disulfidbroer -S-S-, som forbinder to polymermolekyler, eller danner jumpere i en kjede.
Molekyler av aminosyren cystein (tabell 1) inneholder reaktive sulfhydridgrupper -SH, som interagerer med hverandre og danner disulfidbroer -S-S-. Cysteins rolle i proteinverdenen er spesiell, med dens deltakelse dannes det tverrbindinger mellom polymere proteinmolekyler.
Assosiasjonen av aminosyrer til en polymerkjede skjer i en levende organisme under kontroll av nukleinsyrer, det er de som gir en streng monteringsrekkefølge og regulerer den faste lengden på polymermolekylet ( cm. NUKLEINSYRER).
Strukturen til proteiner.
Sammensetningen av proteinmolekylet, presentert i form av alternerende aminosyrerester (fig. 2), kalles proteinets primære struktur. Hydrogenbindinger oppstår mellom iminogruppene HN som er tilstede i polymerkjeden og karbonylgruppene CO ( cm. HYDROGEN BOND), som et resultat får proteinmolekylet en viss romlig form, kalt den sekundære strukturen. Det vanligste er to typer sekundærstruktur i proteiner.
Det første alternativet, kalt α-helix, implementeres ved bruk av hydrogenbindinger i ett polymermolekyl. De geometriske parameterne til molekylet, bestemt av bindingslengdene og bindingsvinklene, er slik at dannelsen av hydrogenbindinger er mulig for H-N og C=O gruppene, mellom hvilke det er to peptidfragmenter H-N-C=O (fig. 3). .
Sammensetningen av polypeptidkjeden vist i fig. 3 er skrevet i forkortet form som følger:
H 2 N-ALA VAL-ALA-LEY-ALA-ALA-ALA-ALA-VAL-ALA-ALA-ALA-COOH.
Som et resultat av den sammentrekkende virkningen av hydrogenbindinger tar molekylet form av en helix - den såkalte α-helixen, den er avbildet som et buet spiralformet bånd som går gjennom atomene som danner polymerkjeden (fig. 4).
Ris. fire 3D-MODELL AV ET PROTEINMOLEKYL i form av en α-helix. Hydrogenbindinger vises som grønne stiplede linjer. Den sylindriske formen til spiralen er synlig ved en viss rotasjonsvinkel (hydrogenatomer er ikke vist i figuren). Fargen på individuelle atomer er gitt i samsvar med internasjonale regler, som anbefaler svart for karbonatomer, blått for nitrogen, rødt for oksygen og gult for svovel (hvit farge anbefales for hydrogenatomer som ikke er vist i figuren, i dette tilfellet hele strukturen avbildet på en mørk bakgrunn).
En annen variant av sekundærstrukturen, kalt β-strukturen, dannes også med deltakelse av hydrogenbindinger, forskjellen er at H-N og C=O-gruppene til to eller flere polymerkjeder plassert parallelt samhandler. Siden polypeptidkjeden har en retning (fig. 1), er varianter mulige når retningen til kjedene er den samme (parallell β-struktur, fig. 5), eller de er motsatte (antiparallell β-struktur, fig. 6). .
Polymerkjeder av forskjellige sammensetninger kan delta i dannelsen av β-strukturen, mens de organiske gruppene som rammer inn polymerkjeden (Ph, CH 2 OH, etc.) i de fleste tilfeller spiller en sekundær rolle, den gjensidige ordningen av H-N og C =O-grupper er avgjørende. Siden H-N- og C=O-gruppene er rettet i forskjellige retninger i forhold til polymerkjeden (opp og ned i figuren), blir det mulig for tre eller flere kjeder å samhandle samtidig.
Sammensetningen av den første polypeptidkjeden i fig. 5:
H 2 N-LEI-ALA-FEN-GLI-ALA-ALA-COOH
Sammensetningen av den andre og tredje kjeden:
H 2 N-GLY-ALA-SER-GLY-TRE-ALA-COOH
Sammensetningen av polypeptidkjedene vist i fig. 6, det samme som i fig. 5, er forskjellen at den andre kjeden har motsatt (i sammenligning med fig. 5) retning.
Det er mulig å danne en β-struktur i ett molekyl, når et kjedefragment i en bestemt seksjon viser seg å være rotert med 180°, i dette tilfellet har to grener av ett molekyl motsatt retning, som et resultat, en antiparallell Det dannes β-struktur (fig. 7).
Strukturen vist i fig. 7 i et flatt bilde, vist i fig. 8 i form av en tredimensjonal modell. Seksjoner av β-strukturen er vanligvis betegnet på en forenklet måte med et flatt bølget bånd som passerer gjennom atomene som danner polymerkjeden.
I strukturen til mange proteiner veksler deler av α-helix og båndlignende β-strukturer, så vel som enkeltpolypeptidkjeder. Deres gjensidige arrangement og veksling i polymerkjeden kalles den tertiære strukturen til proteinet.
Metoder for å skildre strukturen til proteiner er vist nedenfor ved å bruke planteproteinet crambin som et eksempel. Strukturformler av proteiner, som ofte inneholder opptil hundrevis av aminosyrefragmenter, er komplekse, tungvinte og vanskelige å forstå, derfor brukes noen ganger forenklede strukturformler - uten symboler for kjemiske elementer (fig. 9, alternativ A), men samtidig gang de beholder fargen på valensslag i samsvar med internasjonale regler (fig. 4). I dette tilfellet presenteres formelen ikke i et flatt, men i et romlig bilde, som tilsvarer den virkelige strukturen til molekylet. Denne metoden gjør det for eksempel mulig å skille mellom disulfidbroer (lik de som finnes i insulin, fig. 2), fenylgrupper i siderammen av kjeden osv. Bildet av molekyler i form av tredimensjonale modeller (kuler forbundet med stenger) er noe klarere (fig. 9, alternativ B). Begge metodene tillater imidlertid ikke å vise den tertiære strukturen, så den amerikanske biofysikeren Jane Richardson foreslo å avbilde α-strukturer som spiralvridde bånd (se fig. 4), β-strukturer som flate bølgete bånd (fig. 8) og forbindende dem enkeltkjeder - i form av tynne bunter har hver type struktur sin egen farge. Denne metoden for å avbilde den tertiære strukturen til et protein er nå mye brukt (fig. 9, variant B). Noen ganger, for større informasjonsinnhold, vises en tertiær struktur og en forenklet strukturformel sammen (fig. 9, variant D). Det er også modifikasjoner av metoden foreslått av Richardson: α-helikser er avbildet som sylindre, og β-strukturer er i form av flate piler som indikerer retningen til kjeden (fig. 9, alternativ E). Mindre vanlig er metoden hvor hele molekylet er avbildet som en bunt, hvor ulik struktur utmerker seg ved forskjellige farger, og disulfidbroer er vist som gule broer (fig. 9, variant E).
Alternativ B er det mest hensiktsmessige for persepsjon, når de strukturelle egenskapene til proteinet (aminosyrefragmenter, deres vekslende rekkefølge, hydrogenbindinger) ikke er indikert når det avbildes tertiærstrukturen, mens det antas at alle proteiner inneholder "detaljer" tatt fra et standardsett med tjue aminosyrer (tabell 1). Hovedoppgaven i å skildre en tertiær struktur er å vise det romlige arrangementet og vekslingen av sekundære strukturer.
Ris. 9 ULIKE VERSJONER AV BILDET AV STRUKTUREN AV KRUMMINPROTEINET.
A er en strukturformel i et romlig bilde.
B - struktur i form av en tredimensjonal modell.
B er den tertiære strukturen til molekylet.
G - en kombinasjon av alternativene A og B.
E - forenklet bilde av tertiærstrukturen.
E - tertiær struktur med disulfidbroer.
Det mest praktiske for persepsjon er en tredimensjonal tertiær struktur (alternativ B), frigjort fra detaljene i strukturformelen.
Et proteinmolekyl som har en tertiær struktur, får som regel en viss konfigurasjon, som er dannet av polare (elektrostatiske) interaksjoner og hydrogenbindinger. Som et resultat tar molekylet form av en kompakt spole - kuleformede proteiner (kuler, lat. ball), eller filamentøse - fibrillære proteiner (fibra, lat. fiber).
Et eksempel på en kulestruktur er proteinet albumin, proteinet til et kyllingegg tilhører klassen albuminer. Den polymere kjeden av albumin er hovedsakelig satt sammen av alanin, asparaginsyre, glycin og cystein, alternerende i en viss rekkefølge. Den tertiære strukturen inneholder α-helikser forbundet med enkeltkjeder (fig. 10).
Ris. ti GLOBULAR STRUKTUR AV ALBUMIN
Et eksempel på en fibrillær struktur er fibroinproteinet. De inneholder en stor mengde glycin-, alanin- og serinrester (hver andre aminosyrerest er glycin); cysteinrester som inneholder sulfhydridgrupper er fraværende. Fibroin, hovedkomponenten i naturlig silke og spindelvev, inneholder β-strukturer forbundet med enkeltkjeder (fig. 11).
Ris. elleve FIBRILLÆR PROTEIN FIBROIN
Muligheten for å danne en tertiær struktur av en viss type er iboende i proteinets primære struktur, dvs. bestemt på forhånd av rekkefølgen av alternering av aminosyrerester. Fra visse sett av slike rester oppstår hovedsakelig α-helikser (det er ganske mange slike sett), et annet sett fører til utseendet av β-strukturer, enkeltkjeder er preget av deres sammensetning.
Noen proteinmolekyler, mens de beholder en tertiær struktur, er i stand til å kombineres til store supramolekylære aggregater, mens de holdes sammen av polare interaksjoner, så vel som hydrogenbindinger. Slike formasjoner kalles proteinets kvaternære struktur. For eksempel danner ferritinproteinet, som hovedsakelig består av leucin, glutaminsyre, asparaginsyre og histidin (ferricin inneholder alle de 20 aminosyrerester i varierende mengder) en tertiær struktur av fire parallelllagte α-helikser. Når molekyler kombineres til et enkelt ensemble (fig. 12), dannes det en kvartær struktur, som kan omfatte opptil 24 ferritinmolekyler.
Fig.12 DANNING AV DEN KVARNÆRE STRUKTUREN TIL DET GLOBULAR PROTEINET FERRITIN
Et annet eksempel på supramolekylære formasjoner er strukturen til kollagen. Det er et fibrillært protein hvis kjeder hovedsakelig er bygget opp av glycin vekslende med prolin og lysin. Strukturen inneholder enkeltkjeder, trippel α-helikser, alternerende med båndlignende β-strukturer stablet i parallelle bunter (fig. 13).
Fig.13 SUPRAMOLEKULÆR STRUKTUR AV KOLLAGENFIBRILLÆR PROTEIN
Kjemiske egenskaper til proteiner.
Under påvirkning av organiske løsemidler, avfallsprodukter fra noen bakterier (melkesyregjæring) eller med en økning i temperatur, ødelegges sekundære og tertiære strukturer uten å skade dens primære struktur, som et resultat mister proteinet løselighet og mister biologisk aktivitet, dette prosess kalles denaturering, det vil si tap av naturlige egenskaper, for eksempel curdling av sur melk, det koagulerte proteinet til et kokt kyllingegg. Ved forhøyede temperaturer denaturerer proteinene til levende organismer (spesielt mikroorganismer) raskt. Slike proteiner er ikke i stand til å delta i biologiske prosesser, som et resultat dør mikroorganismer, så kokt (eller pasteurisert) melk kan lagres lenger.
Peptidbindinger H-N-C=O, som danner polymerkjeden til proteinmolekylet, hydrolyseres i nærvær av syrer eller alkalier, og polymerkjeden brytes, noe som til slutt kan føre til de opprinnelige aminosyrene. Peptidbindinger inkludert i α-helikser eller β-strukturer er mer motstandsdyktige mot hydrolyse og ulike kjemiske angrep (sammenlignet med de samme bindingene i enkeltkjeder). En mer delikat demontering av proteinmolekylet til dets inngående aminosyrer utføres i et vannfritt medium ved bruk av hydrazin H 2 N–NH 2, mens alle aminosyrefragmenter, bortsett fra det siste, danner de såkalte karboksylsyrehydrazider som inneholder fragmentet C (O)–HN–NH 2 (fig. 14).
Ris. fjorten. POLYPEPTIDSPLYTNING
En slik analyse kan gi informasjon om aminosyresammensetningen til et protein, men det er viktigere å kjenne deres rekkefølge i et proteinmolekyl. En av metodene som er mye brukt til dette formålet er virkningen av fenylisotiocyanat (FITC) på polypeptidkjeden, som i et alkalisk medium fester seg til polypeptidet (fra enden som inneholder aminogruppen), og når reaksjonen til mediet endres. til sur, løsner den fra kjeden og tar med seg et fragment av en aminosyre (fig. 15).
Ris. femten SEKVENSIELL POLYPEPTID-spaltning
Mange spesielle metoder er utviklet for en slik analyse, inkludert de som begynner å "demontere" et proteinmolekyl i dets bestanddeler, med utgangspunkt i karboksylenden.
Kryssdisulfidbroer S-S (dannet ved interaksjon av cysteinrester, fig. 2 og 9) spaltes, og gjør dem til HS-grupper ved påvirkning av forskjellige reduksjonsmidler. Virkningen av oksidasjonsmidler (oksygen eller hydrogenperoksid) fører igjen til dannelse av disulfidbroer (fig. 16).
Ris. 16. Spaltning av disulfidbroer
For å lage ytterligere tverrbindinger i proteiner, brukes reaktiviteten til amino- og karboksylgrupper. Mer tilgjengelig for ulike interaksjoner er aminogruppene som er i siderammen av kjeden - fragmenter av lysin, asparagin, lysin, prolin (tabell 1). Når slike aminogrupper interagerer med formaldehyd, oppstår kondensasjonsprosessen og kryssbroer –NH–CH2–NH– oppstår (fig. 17).
Ris. 17 OPPRETTELSE AV EKSTRA TRANSVERSALE BROER MELLOM PROTEINMOLEKYLER.
De terminale karboksylgruppene i proteinet er i stand til å reagere med komplekse forbindelser av noen flerverdige metaller (kromforbindelser brukes oftere), og tverrbindinger forekommer også. Begge prosessene brukes i skinngarving.
Rollen til proteiner i kroppen.
Rollen til proteiner i kroppen er mangfoldig.
Enzymer(gjæring lat. - fermentering), deres andre navn er enzymer (en zumh gresk. - i gjær) - dette er proteiner med katalytisk aktivitet, de er i stand til å øke hastigheten på biokjemiske prosesser tusenvis av ganger. Under påvirkning av enzymer brytes matens bestanddeler: proteiner, fett og karbohydrater ned til enklere forbindelser, hvorfra nye makromolekyler syntetiseres, som er nødvendige for en viss type kropp. Enzymer deltar også i mange biokjemiske synteseprosesser, for eksempel i syntesen av proteiner (noen proteiner hjelper til med å syntetisere andre). Cm. ENZYMER
Enzymer er ikke bare svært effektive katalysatorer, men også selektive (diriger reaksjonen strengt i den gitte retningen). I deres nærvær fortsetter reaksjonen med nesten 100 % utbytte uten dannelse av biprodukter, og samtidig er strømningsforholdene milde: normalt atmosfærisk trykk og temperatur for en levende organisme. Til sammenligning utføres syntesen av ammoniakk fra hydrogen og nitrogen i nærvær av en aktivert jernkatalysator ved 400–500 °C og et trykk på 30 MPa, utbyttet av ammoniakk er 15–25 % per syklus. Enzymer regnes som uovertruffen katalysatorer.
Intensive studier av enzymer begynte på midten av 1800-tallet; mer enn 2000 forskjellige enzymer har nå blitt studert; dette er den mest mangfoldige klassen av proteiner.
Navnene på enzymer er som følger: navnet på reagenset som enzymet interagerer med, eller navnet på den katalyserte reaksjonen, legges til med endelsen -aza, for eksempel, arginase dekomponerer arginin (tabell 1), dekarboksylase katalyserer dekarboksylering, dvs. eliminering av CO 2 fra karboksylgruppen:
– COOH → – CH + CO 2
Ofte, for mer nøyaktig å indikere rollen til et enzym, er både objektet og typen reaksjon angitt i navnet, for eksempel er alkoholdehydrogenase et enzym som dehydrogenerer alkoholer.
For noen enzymer oppdaget for ganske lenge siden, har det historiske navnet (uten endelsen -aza) blitt bevart, for eksempel pepsin (pepsis, gresk. fordøyelse) og trypsin (trypsis gresk. flytendegjøring), bryter disse enzymene ned proteiner.
For systematisering er enzymer kombinert i store klasser, klassifiseringen er basert på typen reaksjon, klassene er navngitt etter det generelle prinsippet - navnet på reaksjonen og slutten - aza. Noen av disse klassene er listet opp nedenfor.
Oksidoreduktase er enzymer som katalyserer redoksreaksjoner. Dehydrogenasene som inngår i denne klassen utfører protonoverføring, for eksempel oksiderer alkoholdehydrogenase (ADH) alkoholer til aldehyder, den påfølgende oksidasjonen av aldehyder til karboksylsyrer katalyseres av aldehyddehydrogenaser (ALDH). Begge prosessene skjer i kroppen under bearbeiding av etanol til eddiksyre (fig. 18).
Ris. atten TO-Trinns OKSIDERING AV ETANOL til eddiksyre
Det er ikke etanol som har en narkotisk effekt, men mellomproduktet acetaldehyd, jo lavere aktiviteten til ALDH-enzymet er, jo langsommere går det andre stadiet - oksidasjonen av acetaldehyd til eddiksyre, og jo lengre og sterkere blir ruseffekten ved inntak. av etanol. Analysen viste at mer enn 80 % av representantene for den gule rasen har en relativt lav aktivitet av ALDH og derfor en markant mer alvorlig alkoholtoleranse. Årsaken til denne medfødte reduserte aktiviteten til ALDH er at en del av glutaminsyrerestene i det "svekkede" ALDH-molekylet er erstattet av lysinfragmenter (tabell 1).
Overføringer- enzymer som katalyserer overføringen av funksjonelle grupper, for eksempel transiminase katalyserer overføringen av en aminogruppe.
Hydrolaser er enzymer som katalyserer hydrolyse. De tidligere nevnte trypsin og pepsin hydrolyserer peptidbindinger, og lipaser spalter esterbindingen i fett:
–RC(O)OR 1 + H 2 O → –RC(O)OH + HOR 1
Liase- enzymer som katalyserer reaksjoner som foregår på en ikke-hydrolytisk måte, som et resultat av slike reaksjoner brytes C-C, C-O, C-N bindinger og nye bindinger dannes. Enzymet dekarboksylase tilhører denne klassen
Isomeraser- enzymer som katalyserer isomerisering, for eksempel omdannelse av maleinsyre til fumarsyre (fig. 19), dette er et eksempel på cis-trans-isomerisering (se ISOMERIA).
Ris. 19. ISOMERISERING AV MALEINSYRE til fumarsyre i nærvær av enzymet.
I arbeidet med enzymer observeres det generelle prinsippet, ifølge hvilket det alltid er en strukturell samsvar mellom enzymet og reagenset til den akselererte reaksjonen. I følge det figurative uttrykket til en av grunnleggerne av læren om enzymer, E. Fisher, nærmer reagenset seg enzymet som en nøkkel til en lås. I denne forbindelse katalyserer hvert enzym en viss kjemisk reaksjon eller en gruppe reaksjoner av samme type. Noen ganger kan et enzym virke på en enkelt forbindelse, for eksempel urease (uron gresk. - urin) katalyserer bare hydrolyse av urea:
(H 2 N) 2 C \u003d O + H 2 O \u003d CO 2 + 2NH 3
Den fineste selektiviteten vises av enzymer som skiller mellom optisk aktive antipoder - venstre- og høyrehendte isomerer. L-arginase virker kun på venstredreiende arginin og påvirker ikke den høyredreiende isomeren. L-laktatdehydrogenase virker bare på de venstredreiende estere av melkesyre, de såkalte laktatene (lactis). lat. melk), mens D-laktatdehydrogenase bare bryter ned D-laktater.
De fleste av enzymene virker ikke på én, men på en gruppe beslektede forbindelser, for eksempel "foretrekker" trypsin å spalte peptidbindingene dannet av lysin og arginin (tabell 1.)
De katalytiske egenskapene til noen enzymer, som hydrolaser, bestemmes utelukkende av strukturen til selve proteinmolekylet, en annen klasse enzymer - oksidoreduktaser (for eksempel alkoholdehydrogenase) kan bare være aktive i nærvær av ikke-proteinmolekyler assosiert med dem - vitaminer som aktiverer Mg, Ca, Zn, Mn og fragmenter av nukleinsyrer (fig. 20).
Ris. tjue ALKOHOLDDEHYDROGENASEMOLEKYL
Transportproteiner binder og transporterer ulike molekyler eller ioner gjennom cellemembraner (både innenfor og utenfor cellen), samt fra ett organ til et annet.
For eksempel binder hemoglobin oksygen når blod passerer gjennom lungene og leverer det til forskjellige vev i kroppen, hvor oksygen frigjøres og deretter brukes til å oksidere matkomponenter, denne prosessen fungerer som en energikilde (noen ganger bruker de begrepet "brenning" mat i kroppen).
I tillegg til proteindelen inneholder hemoglobin en kompleks forbindelse av jern med et syklisk porfyrinmolekyl (porphyros gresk. - lilla), som bestemmer den røde fargen på blodet. Det er dette komplekset (fig. 21, venstre) som spiller rollen som en oksygenbærer. I hemoglobin er jernporfyrinkomplekset lokalisert inne i proteinmolekylet og holdes tilbake av polare interaksjoner, samt av en koordinasjonsbinding med nitrogen i histidin (tabell 1), som er en del av proteinet. O2-molekylet, som bæres av hemoglobin, festes via en koordinasjonsbinding til jernatomet fra den motsatte siden av den som histidin er festet til (fig. 21, høyre).
Ris. 21 STRUKTUR AV JERNKOMPLEKSET
Strukturen til komplekset er vist til høyre i form av en tredimensjonal modell. Komplekset holdes i proteinmolekylet av en koordinasjonsbinding (stiplet blå linje) mellom Fe-atomet og N-atomet i histidin, som er en del av proteinet. O 2-molekylet, som bæres av hemoglobin, er koordinert (rød stiplet linje) til Fe-atomet fra det motsatte landet til det plane komplekset.
Hemoglobin er et av de mest studerte proteinene, det består av a-helikser forbundet med enkeltkjeder og inneholder fire jernkomplekser. Dermed er hemoglobin som en voluminøs pakke for overføring av fire oksygenmolekyler på en gang. Formen for hemoglobin tilsvarer globulære proteiner (fig. 22).
Ris. 22 GLOBULAR FORM AV HEMOGLOBIN
Den største "fordelen" med hemoglobin er at tilsetningen av oksygen og dets påfølgende spaltning under overføring til ulike vev og organer skjer raskt. Karbonmonoksid, CO (karbonmonoksid), binder seg enda raskere til Fe i hemoglobin, men danner i motsetning til O 2 et kompleks som er vanskelig å bryte ned. Som et resultat er slikt hemoglobin ikke i stand til å binde O 2, noe som fører (når store mengder karbonmonoksid inhaleres) til kroppens død ved kvelning.
Den andre funksjonen til hemoglobin er overføring av utåndet CO 2, men ikke jernatomet, men H 2 i N-gruppen til proteinet er involvert i prosessen med midlertidig binding av karbondioksid.
"Ytelsen" til proteiner avhenger av deres struktur, for eksempel å erstatte den eneste aminosyreresten av glutaminsyre i hemoglobinpolypeptidkjeden med en valinrest (en sjelden observert medfødt anomali) fører til en sykdom som kalles sigdcelleanemi.
Det finnes også transportproteiner som kan binde fett, glukose, aminosyrer og frakte dem både i og utenfor cellene.
Transportproteiner av en spesiell type bærer ikke selve stoffene, men fungerer som en "transportregulator", som passerer visse stoffer gjennom membranen (cellens yttervegg). Slike proteiner kalles ofte membranproteiner. De har form som en hul sylinder og, ved å være innebygd i membranveggen, sørger de for bevegelse av noen polare molekyler eller ioner inn i cellen. Et eksempel på et membranprotein er porin (fig. 23).
Ris. 23 PORIN PROTEIN
Mat- og lagringsproteiner, som navnet tilsier, tjener som kilder til intern ernæring, oftere for embryoene til planter og dyr, så vel som i de tidlige stadiene av utviklingen av unge organismer. Kostholdsproteiner inkluderer albumin (fig. 10) - hovedkomponenten i eggehvite, samt kasein - hovedproteinet i melk. Under påvirkning av enzymet pepsin stivner kasein i magen, noe som sikrer oppbevaring i fordøyelseskanalen og effektiv absorpsjon. Kasein inneholder fragmenter av alle aminosyrene som kroppen trenger.
I ferritin (fig. 12), som finnes i vevet til dyr, lagres jernioner.
Myoglobin er også et lagringsprotein, som ligner hemoglobin i sammensetning og struktur. Myoglobin er hovedsakelig konsentrert i musklene, dens hovedrolle er lagring av oksygen, som hemoglobin gir det. Det mettes raskt med oksygen (mye raskere enn hemoglobin), og overfører det deretter gradvis til forskjellige vev.
Strukturelle proteiner utfører en beskyttende funksjon (hud) eller støtte - de holder kroppen sammen og gir den styrke (brusk og sener). Hovedkomponenten deres er det fibrillære proteinet kollagen (fig. 11), det vanligste proteinet i dyreverdenen, i kroppen til pattedyr, det utgjør nesten 30 % av den totale massen av proteiner. Kollagen har høy strekkfasthet (styrken på huden er kjent), men på grunn av det lave innholdet av tverrbindinger i hudkollagen er dyreskinn lite egnet i sin rå form for fremstilling av ulike produkter. For å redusere hevelse av huden i vann, krymping under tørking, samt for å øke styrken i vannet tilstand og øke elastisiteten i kollagen, skapes ytterligere tverrbindinger (fig. 15a), dette er den s.k. skinnbruningsprosess.
I levende organismer blir kollagenmolekyler som har oppstått i prosessen med vekst og utvikling av organismen ikke oppdatert og erstattes ikke av nylig syntetiserte. Etter hvert som kroppen eldes, øker antallet tverrbindinger i kollagen, noe som fører til en reduksjon i elastisiteten, og siden fornyelse ikke skjer, vises aldersrelaterte endringer - en økning i skjørheten til brusk og sener, utseende av rynker på huden.
Leddbånd inneholder elastin, et strukturelt protein som lett strekker seg i to dimensjoner. Resilinproteinet, som er lokalisert ved hengslene til vingene hos noen insekter, har størst elastisitet.
Hornformasjoner - hår, negler, fjær, bestående hovedsakelig av keratinprotein (fig. 24). Hovedforskjellen er det merkbare innholdet av cysteinrester, som danner disulfidbroer, som gir høy elastisitet (evnen til å gjenopprette sin opprinnelige form etter deformasjon) til hår, samt ullstoffer.
Ris. 24. FRAGMENT AV FIBRILLAR PROTEIN KERATIN
For en irreversibel endring i formen til et keratinobjekt, må du først ødelegge disulfidbroene ved hjelp av et reduksjonsmiddel, gi det en ny form, og deretter gjenskape disulfidbroene ved hjelp av et oksidasjonsmiddel (fig. . 16), slik utføres for eksempel permanent hår.
Med en økning i innholdet av cysteinrester i keratin og følgelig en økning i antall disulfidbroer, forsvinner evnen til å deformere, men samtidig vises høy styrke (horn av hovdyr og skilpaddeskall inneholder opptil 18 % av cysteinfragmentene). Pattedyr har opptil 30 forskjellige typer keratin.
Det keratinrelaterte fibrillære proteinet fibroin som skilles ut av silkeormslarver under kokongkrølling, samt av edderkopper under nettveving, inneholder kun β-strukturer forbundet med enkeltkjeder (fig. 11). I motsetning til keratin har ikke fibroin tverrgående disulfidbroer, det har en veldig sterk strekkfasthet (styrken per enhetstverrsnitt av noen baneprøver er høyere enn for stålkabler). På grunn av fraværet av tverrbindinger er fibroin uelastisk (det er kjent at ullstoffer er nesten uutslettelige, og silkestoffer blir lett rynkete).
regulatoriske proteiner.
Regulerende proteiner, oftere referert til som hormoner, er involvert i ulike fysiologiske prosesser. For eksempel består hormonet insulin (fig. 25) av to α-kjeder forbundet med disulfidbroer. Insulin regulerer metabolske prosesser som involverer glukose, fraværet fører til diabetes.
Ris. 25 PROTEININSULIN
Hypofysen i hjernen syntetiserer et hormon som regulerer veksten av kroppen. Det er regulatoriske proteiner som styrer biosyntesen av ulike enzymer i kroppen.
Kontraktile og motoriske proteiner gir kroppen evnen til å trekke seg sammen, endre form og bevege seg, først og fremst snakker vi om muskler. 40 % av massen av alle proteiner som finnes i musklene er myosin (mys, myos, gresk. - muskler). Molekylet inneholder både en fibrillær og en kuleformet del (fig. 26)
Ris. 26 MYOSINMOLEKYL
Slike molekyler kombineres til store aggregater som inneholder 300–400 molekyler.
Når konsentrasjonen av kalsiumioner endres i rommet rundt muskelfibrene, oppstår en reversibel endring i konformasjonen til molekylene - en endring i formen på kjeden på grunn av rotasjonen av individuelle fragmenter rundt valensbindingene. Dette fører til muskelsammentrekning og avspenning, signalet om å endre konsentrasjonen av kalsiumioner kommer fra nerveendene i muskelfibrene. Kunstig muskelsammentrekning kan være forårsaket av virkningen av elektriske impulser, noe som fører til en skarp endring i konsentrasjonen av kalsiumioner, dette er grunnlaget for å stimulere hjertemuskelen for å gjenopprette hjertets arbeid.
Beskyttende proteiner lar deg beskytte kroppen mot invasjon av angripende bakterier, virus og fra penetrasjon av fremmede proteiner (det generaliserte navnet på fremmedlegemer er antigener). Rollen til beskyttende proteiner utføres av immunglobuliner (deres andre navn er antistoffer), de gjenkjenner antigener som har trengt inn i kroppen og binder seg fast til dem. I kroppen til pattedyr, inkludert mennesker, er det fem klasser av immunglobuliner: M, G, A, D og E, deres struktur, som navnet tilsier, er kuleformet, i tillegg er de alle bygget på lignende måte. Den molekylære organiseringen av antistoffer er vist nedenfor ved bruk av klasse G immunglobulin som et eksempel (fig. 27). Molekylet inneholder fire polypeptidkjeder forbundet med tre S-S disulfidbroer (i fig. 27 er de vist med fortykkede valensbindinger og store S-symboler), i tillegg inneholder hver polymerkjede disulfidbroer i kjeden. To store polymerkjeder (uthevet i blått) inneholder 400–600 aminosyrerester. De to andre kjedene (uthevet i grønt) er nesten halvparten så lange, og inneholder omtrent 220 aminosyrerester. Alle fire kjedene er plassert på en slik måte at terminal H 2 N-gruppene er rettet i én retning.
Ris. 27 SKJEMATISK TEGNING AV STRUKTUREN TIL IMMUNOGLOBULIN
Etter at kroppen kommer i kontakt med et fremmed protein (antigen), begynner cellene i immunsystemet å produsere immunglobuliner (antistoffer), som samler seg i blodserumet. På det første trinnet utføres hovedarbeidet av kjedeseksjoner som inneholder terminal H 2 N (i fig. 27 er de tilsvarende seksjonene merket med lyseblått og lysegrønt). Dette er antigenfangststeder. I prosessen med immunglobulinsyntese dannes disse stedene på en slik måte at deres struktur og konfigurasjon samsvarer så mye som mulig med strukturen til antigenet som nærmer seg (som en nøkkel til en lås, som enzymer, men oppgavene i dette tilfellet er forskjellig). For hvert antigen dannes det således et strengt individuelt antistoff som en immunrespons. Ikke et eneste kjent protein kan endre strukturen så "plastisk" avhengig av eksterne faktorer, i tillegg til immunglobuliner. Enzymer løser problemet med strukturell overensstemmelse med reagenset på en annen måte - ved hjelp av et gigantisk sett av forskjellige enzymer for alle mulige tilfeller, og immunglobuliner bygger hver gang opp igjen "arbeidsverktøyet". Dessuten gir hengselregionen til immunglobulinet (fig. 27) de to fangstområdene en viss uavhengig mobilitet, som et resultat av dette kan immunglobulinmolekylet umiddelbart "finne" de to mest hensiktsmessige områdene for fangst i antigenet for sikkert å fiksere det, dette ligner handlingene til en krepsdyr.
Deretter slås en kjede av påfølgende reaksjoner av kroppens immunsystem på, immunglobuliner fra andre klasser kobles sammen, som et resultat deaktiveres det fremmede proteinet, og deretter blir antigenet (fremmed mikroorganisme eller toksin) ødelagt og fjernet.
Etter kontakt med antigenet nås den maksimale konsentrasjonen av immunglobulin (avhengig av antigenets natur og de individuelle egenskapene til selve organismen) innen noen få timer (noen ganger flere dager). Kroppen beholder minnet om slik kontakt, og når de blir angrepet igjen med det samme antigenet, akkumuleres immunglobuliner i blodserumet mye raskere og i større mengder - ervervet immunitet oppstår.
Ovennevnte klassifisering av proteiner er til en viss grad betinget, for eksempel er trombinproteinet, nevnt blant beskyttende proteiner, i hovedsak et enzym som katalyserer hydrolysen av peptidbindinger, det vil si at det tilhører klassen av proteaser.
Beskyttende proteiner blir ofte referert til som slangegiftproteiner og de giftige proteinene til noen planter, siden deres oppgave er å beskytte kroppen mot skade.
Det finnes proteiner hvis funksjoner er så unike at det gjør det vanskelig å klassifisere dem. For eksempel er proteinet monellin, som finnes i en afrikansk plante, veldig søtt og har vært gjenstand for studier som et ikke-giftig stoff som kan brukes i stedet for sukker for å forhindre fedme. Blodplasmaet til noen antarktiske fisker inneholder proteiner med frostvæskeegenskaper som forhindrer at blodet til disse fiskene fryser.
Kunstig syntese av proteiner.
Kondensering av aminosyrer som fører til en polypeptidkjede er en godt studert prosess. Det er mulig å utføre for eksempel kondensasjonen av en hvilken som helst aminosyre eller en blanding av syrer og oppnå henholdsvis en polymer som inneholder de samme enhetene, eller forskjellige enheter, alternerende i tilfeldig rekkefølge. Slike polymerer har liten likhet med naturlige polypeptider og har ikke biologisk aktivitet. Hovedoppgaven er å koble aminosyrer i en strengt definert, forhåndsplanlagt rekkefølge for å reprodusere sekvensen av aminosyrerester i naturlige proteiner. Den amerikanske vitenskapsmannen Robert Merrifield foreslo en original metode som gjorde det mulig å løse et slikt problem. Essensen av metoden er at den første aminosyren er festet til en uløselig polymergel som inneholder reaktive grupper som kan kombineres med –COOH – grupper i aminosyren. Tverrbundet polystyren med klormetylgrupper innført i det ble tatt som et slikt polymert substrat. For at aminosyren som tas for reaksjonen ikke reagerer med seg selv, og slik at den ikke forbinder H 2 N-gruppen til substratet, er aminogruppen til denne syren pre-blokkert med en voluminøs substituent [(C 4 H 9) 3] 3 OS (O) -gruppe. Etter at aminosyren har festet seg til den polymere bæreren, fjernes blokkeringsgruppen og en annen aminosyre introduseres i reaksjonsblandingen, der H 2 N-gruppen også tidligere er blokkert. I et slikt system er bare interaksjonen mellom H 2 N-gruppen til den første aminosyren og –COOH-gruppen til den andre syren mulig, som utføres i nærvær av katalysatorer (fosfoniumsalter). Deretter gjentas hele skjemaet, og introduserer den tredje aminosyren (fig. 28).
Ris. 28. SYNTESESKEMA AV POLYPEPTIDKJEDER
I det siste trinnet separeres de resulterende polypeptidkjedene fra polystyrenbæreren. Nå er hele prosessen automatisert, det er automatiske peptidsynthesizere som opererer i henhold til det beskrevne skjemaet. Mange peptider brukt i medisin og landbruk har blitt syntetisert ved denne metoden. Det var også mulig å oppnå forbedrede analoger av naturlige peptider med selektiv og forbedret virkning. Noen små proteiner har blitt syntetisert, som hormonet insulin og noen enzymer.
Det er også metoder for proteinsyntese som replikerer naturlige prosesser: de syntetiserer fragmenter av nukleinsyrer konfigurert til å produsere visse proteiner, deretter settes disse fragmentene inn i en levende organisme (for eksempel i en bakterie), hvoretter kroppen begynner å produsere ønsket protein. På denne måten oppnås nå betydelige mengder vanskelig tilgjengelige proteiner og peptider, samt deres analoger.
Proteiner som matkilder.
Proteiner i en levende organisme brytes hele tiden ned til deres opprinnelige aminosyrer (med uunnværlig deltagelse av enzymer), noen aminosyrer går over i andre, deretter syntetiseres proteiner igjen (også med deltagelse av enzymer), dvs. kroppen fornyer seg hele tiden. Noen proteiner (kollagen i huden, hår) fornyes ikke, kroppen mister dem kontinuerlig og syntetiserer i stedet nye. Proteiner som matkilder utfører to hovedfunksjoner: de forsyner kroppen med byggemateriale for syntese av nye proteinmolekyler og forsyner i tillegg kroppen med energi (kalorikilder).
Kjøttetende pattedyr (inkludert mennesker) får de nødvendige proteinene fra plante- og dyremat. Ingen av proteinene som oppnås fra mat er integrert i kroppen i uendret form. I fordøyelseskanalen brytes alle absorberte proteiner ned til aminosyrer, og proteiner som er nødvendige for en bestemt organisme er allerede bygget av dem, mens de resterende 12 kan syntetiseres fra 8 essensielle syrer (tabell 1) i kroppen hvis de ikke er det. tilføres i tilstrekkelige mengder med mat, men essensielle syrer må tilføres mat uten feil. Svovelatomer i cystein oppnås av kroppen med den essensielle aminosyren metionin. En del av proteinene brytes ned, frigjør energien som er nødvendig for å opprettholde livet, og nitrogenet som finnes i dem skilles ut fra kroppen med urin. Vanligvis mister menneskekroppen 25–30 g protein per dag, så proteinmat må alltid være tilstede i riktig mengde. Minste daglige behov for protein er 37 g for menn og 29 g for kvinner, men anbefalt inntak er nesten dobbelt så høyt. Når du vurderer matvarer, er det viktig å vurdere proteinkvaliteten. I fravær eller lavt innhold av essensielle aminosyrer anses proteinet for å ha lav verdi, så slike proteiner bør inntas i større mengder. Så proteinene i belgfrukter inneholder lite metionin, og proteinene i hvete og mais har lite lysin (begge aminosyrer er essensielle). Animalske proteiner (unntatt kollagener) er klassifisert som fullmat. Et komplett sett med alle essensielle syrer inneholder melkekasein, samt cottage cheese og ost tilberedt av det, så et vegetarisk kosthold, hvis det er veldig strengt, dvs. "meierifri", krever økt inntak av belgfrukter, nøtter og sopp for å tilføre kroppen essensielle aminosyrer i riktig mengde.
Syntetiske aminosyrer og proteiner brukes også som matprodukter, og legger dem til fôr, som inneholder essensielle aminosyrer i små mengder. Det er bakterier som kan behandle og assimilere oljehydrokarboner, i dette tilfellet, for full syntese av proteiner, må de mates med nitrogenholdige forbindelser (ammoniakk eller nitrater). Proteinet som oppnås på denne måten brukes som fôr til husdyr og fjørfe. Et sett med enzymer, karbohydraser, tilsettes ofte dyrefôr, som katalyserer hydrolysen av karbohydratmatkomponenter som er vanskelige å dekomponere (celleveggene til kornavlinger), som et resultat av at plantemat absorberes mer fullstendig.
Mikhail Levitsky
PROTEINER (artikkel 2)
(proteiner), en klasse av komplekse nitrogenholdige forbindelser, de mest karakteristiske og viktige (sammen med nukleinsyrer) komponentene i levende materie. Proteiner utfører mange og varierte funksjoner. De fleste proteiner er enzymer som katalyserer kjemiske reaksjoner. Mange hormoner som regulerer fysiologiske prosesser er også proteiner. Strukturelle proteiner som kollagen og keratin er hovedkomponentene i beinvev, hår og negler. De kontraktile proteinene i muskler har evnen til å endre lengden ved å bruke kjemisk energi til å utføre mekanisk arbeid. Proteiner er antistoffer som binder og nøytraliserer giftige stoffer. Noen proteiner som kan reagere på ytre påvirkninger (lys, lukt) fungerer som reseptorer i sanseorganene som oppfatter irritasjon. Mange proteiner lokalisert inne i cellen og på cellemembranen utfører regulerende funksjoner.
I første halvdel av 1800-tallet mange kjemikere, og blant dem først og fremst J. von Liebig, kom etter hvert til den konklusjon at proteiner er en spesiell klasse nitrogenholdige forbindelser. Navnet "proteiner" (fra det greske protos - det første) ble foreslått i 1840 av den nederlandske kjemikeren G. Mulder.
FYSISKE EGENSKAPER
Proteiner er hvite i fast tilstand, men fargeløse i løsning, med mindre de har en kromoforgruppe (farget), som for eksempel hemoglobin. Løseligheten i vann av ulike proteiner varierer sterkt. Det varierer også med pH og med konsentrasjonen av salter i løsningen, slik at man kan velge forholdene under hvilke ett protein selektivt vil utfelles i nærvær av andre proteiner. Denne "utsaltingsmetoden" er mye brukt for å isolere og rense proteiner. Det rensede proteinet feller ofte ut av løsningen som krystaller.
Sammenlignet med andre forbindelser er molekylvekten til proteiner veldig stor - fra flere tusen til mange millioner dalton. Derfor, under ultrasentrifugering, blir proteiner utfelt, og dessuten med forskjellige hastigheter. På grunn av tilstedeværelsen av positivt og negativt ladede grupper i proteinmolekyler, beveger de seg med forskjellige hastigheter i et elektrisk felt. Dette er grunnlaget for elektroforese, en metode som brukes til å isolere individuelle proteiner fra komplekse blandinger. Rensing av proteiner utføres også ved kromatografi.
KJEMISKE EGENSKAPER
Struktur.
Proteiner er polymerer, dvs. molekyler bygget som kjeder fra repeterende monomerenheter, eller underenheter, hvis rolle spilles av alfa-aminosyrer. Generell formel for aminosyrer
hvor R er et hydrogenatom eller en eller annen organisk gruppe.
Et proteinmolekyl (polypeptidkjede) kan bestå av bare et relativt lite antall aminosyrer eller flere tusen monomerenheter. Sammenkoblingen av aminosyrer i en kjede er mulig fordi hver av dem har to forskjellige kjemiske grupper: en aminogruppe med grunnleggende egenskaper, NH2, og en sur karboksylgruppe, COOH. Begge disse gruppene er knyttet til et karbonatom. Karboksylgruppen til en aminosyre kan danne en amid(peptid)binding med aminogruppen til en annen aminosyre:
Etter at to aminosyrer er koblet sammen på denne måten, kan kjeden forlenges ved å legge en tredje til den andre aminosyren, og så videre. Som man kan se fra ligningen ovenfor, frigjøres et vannmolekyl når en peptidbinding dannes. I nærvær av syrer, alkalier eller proteolytiske enzymer, fortsetter reaksjonen i motsatt retning: polypeptidkjeden spaltes til aminosyrer med tilsetning av vann. Denne reaksjonen kalles hydrolyse. Hydrolyse går spontant, og energi er nødvendig for å kombinere aminosyrer til en polypeptidkjede.
En karboksylgruppe og en amidgruppe (eller en imidgruppe som ligner på den - når det gjelder aminosyren prolin) er tilstede i alle aminosyrer, mens forskjellene mellom aminosyrer bestemmes av arten til den gruppen, eller "siden" kjede", som er indikert ovenfor med bokstaven R. Rollen til sidekjeden kan spilles av et hydrogenatom, som aminosyren glycin, og noen voluminøse grupperinger, som histidin og tryptofan. Noen sidekjeder er kjemisk inerte, mens andre er svært reaktive.
Mange tusen forskjellige aminosyrer kan syntetiseres, og mange forskjellige aminosyrer forekommer i naturen, men bare 20 typer aminosyrer brukes til proteinsyntese: alanin, arginin, asparagin, asparaginsyre, valin, histidin, glycin, glutamin, glutamin. syre, isoleucin, leucin, lysin, metionin, prolin, serin, tyrosin, treonin, tryptofan, fenylalanin og cystein (i proteiner kan cystein være tilstede som en dimer - cystin). Riktignok er det i noen proteiner andre aminosyrer i tillegg til de tjue som regelmessig forekommer, men de dannes som et resultat av modifikasjon av noen av de tjue som er oppført etter at det har blitt inkludert i proteinet.
optisk aktivitet.
Alle aminosyrer, med unntak av glycin, har fire forskjellige grupper knyttet til α-karbonatomet. Når det gjelder geometri, kan fire forskjellige grupper festes på to måter, og følgelig er det to mulige konfigurasjoner, eller to isomerer, relatert til hverandre som et objekt til dets speilbilde, dvs. som venstre hånd til høyre. Den ene konfigurasjonen kalles venstre- eller venstrehendt (L), og den andre høyrehendt eller høyrehendt (D), fordi de to slike isomerer er forskjellige i rotasjonsretningen til planet av polarisert lys. Bare L-aminosyrer forekommer i proteiner (unntaket er glycin; det kan bare representeres i én form, siden to av de fire gruppene er like), og de har alle optisk aktivitet (siden det bare er en isomer). D-aminosyrer er sjeldne i naturen; de finnes i noen antibiotika og celleveggen til bakterier.
Rekkefølgen av aminosyrer.
Aminosyrer i polypeptidkjeden er ikke ordnet tilfeldig, men i en viss fast rekkefølge, og det er denne rekkefølgen som bestemmer funksjonene og egenskapene til proteinet. Ved å variere rekkefølgen på de 20 typene aminosyrer, kan du få et stort antall forskjellige proteiner, akkurat som du kan lage mange forskjellige tekster fra bokstavene i alfabetet.
Tidligere tok det ofte flere år å bestemme aminosyresekvensen til et protein. Direkte bestemmelse er fortsatt en ganske møysommelig oppgave, selv om det er opprettet enheter som lar den utføres automatisk. Det er vanligvis lettere å bestemme nukleotidsekvensen til det tilsvarende genet og utlede aminosyresekvensen til proteinet fra det. Til dags dato er aminosyresekvensene til mange hundre proteiner allerede bestemt. Funksjonene til dekodede proteiner er vanligvis kjent, og dette hjelper til med å forestille seg de mulige funksjonene til lignende proteiner dannet, for eksempel i ondartede neoplasmer.
Komplekse proteiner.
Proteiner som kun består av aminosyrer kalles enkle. Ofte er imidlertid et metallatom eller en kjemisk forbindelse som ikke er en aminosyre festet til polypeptidkjeden. Slike proteiner kalles komplekse. Et eksempel er hemoglobin: det inneholder jernporfyrin, som gir det sin røde farge og lar det fungere som en oksygenbærer.
Navnene på de fleste komplekse proteiner inneholder en indikasjon på arten av de vedlagte gruppene: sukker er tilstede i glykoproteiner, fett i lipoproteiner. Hvis den katalytiske aktiviteten til enzymet avhenger av den vedlagte gruppen, kalles det en protesegruppe. Noen vitaminer spiller ofte rollen som en protesegruppe eller er en del av den. Vitamin A, for eksempel festet til et av proteinene i netthinnen, bestemmer følsomheten for lys.
Tertiær struktur.
Det som er viktig er ikke så mye aminosyresekvensen til proteinet (primærstruktur), men måten det er lagt i rommet. Langs hele lengden av polypeptidkjeden danner hydrogenioner vanlige hydrogenbindinger, som gir den formen av en spiral eller et lag (sekundær struktur). Fra kombinasjonen av slike helikser og lag oppstår en kompakt form av neste rekkefølge - den tertiære strukturen til proteinet. Rundt bindingene som holder de monomere leddene i kjeden, er rotasjoner gjennom små vinkler mulig. Derfor, fra et rent geometrisk synspunkt, er antallet mulige konfigurasjoner for enhver polypeptidkjede uendelig stort. I virkeligheten eksisterer hvert protein normalt i bare én konfigurasjon, bestemt av dets aminosyresekvens. Denne strukturen er ikke stiv, den ser ut til å "puste" - den svinger rundt en viss gjennomsnittlig konfigurasjon. Kjedet brettes inn i en konfigurasjon der den frie energien (evnen til å utføre arbeid) er minimal, akkurat som en frigjort fjær bare komprimeres til en tilstand som tilsvarer et minimum av fri energi. Ofte er den ene delen av kjeden stivt knyttet til den andre av disulfid (–S–S–) bindinger mellom to cysteinrester. Dette er delvis grunnen til at cystein blant aminosyrer spiller en spesielt viktig rolle.
Kompleksiteten i strukturen til proteiner er så stor at det ennå ikke er mulig å beregne tertiærstrukturen til et protein, selv om dets aminosyresekvens er kjent. Men hvis det er mulig å oppnå proteinkrystaller, kan dens tertiære struktur bestemmes ved røntgendiffraksjon.
I strukturelle, kontraktile og enkelte andre proteiner er kjedene forlengede og flere lett foldede kjeder som ligger side om side danner fibriller; fibriller, på sin side, brettes til større formasjoner - fibre. Imidlertid er de fleste proteiner i løsning kuleformede: kjedene er kveilet i en kule, som garn i en ball. Fri energi med denne konfigurasjonen er minimal, siden hydrofobe ("vannavstøtende") aminosyrer er skjult inne i kulen, og hydrofile ("vanntiltrekkende") aminosyrer er på overflaten.
Mange proteiner er komplekser av flere polypeptidkjeder. Denne strukturen kalles proteinets kvaternære struktur. Hemoglobinmolekylet, for eksempel, består av fire underenheter, som hver er et kuleprotein.
Strukturelle proteiner på grunn av deres lineære konfigurasjon danner fibre der strekkstyrken er svært høy, mens den kuleformede konfigurasjonen tillater proteiner å inngå spesifikke interaksjoner med andre forbindelser. På overflaten av kulen, med riktig legging av kjeder, vises hulrom av en viss form, der reaktive kjemiske grupper er lokalisert. Hvis dette proteinet er et enzym, kommer et annet, vanligvis mindre, molekyl av et eller annet stoff inn i et slikt hulrom, akkurat som en nøkkel går inn i en lås; i dette tilfellet endres konfigurasjonen av elektronskyen til molekylet under påvirkning av kjemiske grupper lokalisert i hulrommet, og dette tvinger det til å reagere på en bestemt måte. På denne måten katalyserer enzymet reaksjonen. Antistoffmolekyler har også hulrom der ulike fremmedstoffer binder seg og dermed ufarliggjøres. «Nøkkel og lås»-modellen, som forklarer interaksjonen mellom proteiner og andre forbindelser, gjør det mulig å forstå spesifisiteten til enzymer og antistoffer, d.v.s. deres evne til å reagere bare med visse forbindelser.
Proteiner i ulike typer organismer.
Proteiner som utfører samme funksjon i forskjellige plante- og dyrearter og derfor bærer samme navn, har også en lignende konfigurasjon. De skiller seg imidlertid noe i aminosyresekvensen. Ettersom arter avviker fra en felles stamfar, erstattes noen aminosyrer i visse posisjoner av mutasjoner med andre. Skadelige mutasjoner som forårsaker arvelige sykdommer forkastes ved naturlig utvalg, men gunstige eller i det minste nøytrale kan bevares. Jo nærmere to biologiske arter er hverandre, jo mindre forskjeller finnes i proteinene deres.
Noen proteiner endres relativt raskt, andre er ganske konservative. Sistnevnte inkluderer for eksempel cytokrom c, et respiratorisk enzym som finnes i de fleste levende organismer. Hos mennesker og sjimpanser er aminosyresekvensene identiske, mens i cytokrom c av hvete viste det seg at bare 38 % av aminosyrene var forskjellige. Selv når man sammenligner mennesker og bakterier, kan likheten mellom cytokromer med (forskjellene her påvirker 65 % av aminosyrene) fortsatt sees, selv om den felles stamfaren til bakterier og mennesker levde på jorden for rundt to milliarder år siden. I dag brukes ofte sammenligning av aminosyresekvenser for å bygge et fylogenetisk (genealogisk) tre som gjenspeiler de evolusjonære forholdene mellom ulike organismer.
Denaturering.
Det syntetiserte proteinmolekylet, folding, får sin egen konfigurasjon. Denne konfigurasjonen kan imidlertid ødelegges ved oppvarming, ved å endre pH, ved påvirkning av organiske løsningsmidler, og til og med ved ganske enkelt å agitere løsningen til det kommer bobler på overflaten. Et protein endret på denne måten kalles denaturert; den mister sin biologiske aktivitet og blir vanligvis uløselig. Velkjente eksempler på denaturert protein er kokte egg eller pisket krem. Små proteiner, som bare inneholder rundt hundre aminosyrer, er i stand til å renaturere, dvs. skaffe tilbake den opprinnelige konfigurasjonen. Men de fleste av proteinene blir ganske enkelt transformert til en masse sammenfiltrede polypeptidkjeder og gjenoppretter ikke deres tidligere konfigurasjon.
En av hovedvanskene med å isolere aktive proteiner er deres ekstreme følsomhet for denaturering. Denne egenskapen til proteiner finner nyttig anvendelse i konservering av matprodukter: høy temperatur denaturerer irreversibelt enzymene til mikroorganismer, og mikroorganismene dør.
PROTEIN SYNTESE
For proteinsyntese må en levende organisme ha et system av enzymer som er i stand til å feste en aminosyre til en annen. En informasjonskilde er også nødvendig som kan bestemme hvilke aminosyrer som skal kobles sammen. Siden det finnes tusenvis av typer proteiner i kroppen, og hver av dem består av i gjennomsnitt flere hundre aminosyrer, må informasjonen som kreves være virkelig enorm. Det lagres (i likhet med hvordan en post lagres på et magnetbånd) i nukleinsyremolekylene som utgjør gener.
Enzymaktivering.
En polypeptidkjede syntetisert fra aminosyrer er ikke alltid et protein i sin endelige form. Mange enzymer syntetiseres først som inaktive forløpere og blir aktive først etter at et annet enzym fjerner noen få aminosyrer fra den ene enden av kjeden. Noen av fordøyelsesenzymene, som trypsin, syntetiseres i denne inaktive formen; disse enzymene aktiveres i fordøyelseskanalen som et resultat av fjerning av det terminale fragmentet av kjeden. Hormonet insulin, hvis molekyl i sin aktive form består av to korte kjeder, syntetiseres i form av en enkelt kjede, den såkalte. proinsulin. Deretter fjernes den midtre delen av denne kjeden, og de gjenværende fragmentene binder seg til hverandre og danner det aktive hormonmolekylet. Komplekse proteiner dannes først etter at en viss kjemisk gruppe er festet til proteinet, og denne tilknytningen krever ofte også et enzym.
Metabolsk sirkulasjon.
Etter å ha fôret et dyr med aminosyrer merket med radioaktive isotoper av karbon, nitrogen eller hydrogen, blir merket raskt inkorporert i proteinene. Hvis merkede aminosyrer slutter å komme inn i kroppen, begynner mengden etikett i proteiner å avta. Disse eksperimentene viser at de resulterende proteinene ikke lagres i kroppen før livet er slutt. Alle av dem, med noen få unntak, er i en dynamisk tilstand, dekomponerer konstant til aminosyrer og syntetiseres deretter på nytt.
Noen proteiner brytes ned når cellene dør og blir ødelagt. Dette skjer hele tiden, for eksempel med røde blodceller og epitelceller langs den indre overflaten av tarmen. I tillegg skjer nedbrytning og resyntese av proteiner også i levende celler. Merkelig nok er mindre kjent om nedbrytningen av proteiner enn om deres syntese. Det som derimot er klart er at proteolytiske enzymer er involvert i nedbrytningen, lik de som bryter ned proteiner til aminosyrer i fordøyelseskanalen.
Halveringstiden til ulike proteiner er forskjellig – fra flere timer til mange måneder. Det eneste unntaket er kollagenmolekyler. Når de er dannet, forblir de stabile og blir ikke fornyet eller erstattet. Over tid endres imidlertid noen av egenskapene deres, spesielt elastisiteten, og siden de ikke fornyes, er visse aldersrelaterte endringer et resultat av dette, for eksempel utseendet av rynker på huden.
syntetiske proteiner.
Kjemikere har for lengst lært hvordan man polymeriserer aminosyrer, men aminosyrene kombineres tilfeldig, slik at produktene fra en slik polymerisering har liten likhet med naturlige. Riktignok er det mulig å kombinere aminosyrer i en gitt rekkefølge, noe som gjør det mulig å oppnå noen biologisk aktive proteiner, spesielt insulin. Prosessen er ganske komplisert, og på denne måten er det mulig å oppnå bare de proteinene hvis molekyler inneholder omtrent hundre aminosyrer. Det er å foretrekke i stedet å syntetisere eller isolere nukleotidsekvensen til et gen som tilsvarer den ønskede aminosyresekvensen, og deretter introdusere dette genet i en bakterie, som ved replikasjon vil produsere en stor mengde av det ønskede produktet. Denne metoden har imidlertid også sine ulemper.
PROTEINER OG NÆRING
Når proteiner i kroppen brytes ned til aminosyrer, kan disse aminosyrene gjenbrukes til proteinsyntese. Samtidig er selve aminosyrene utsatt for forfall, slik at de ikke utnyttes fullt ut. Det er også klart at under vekst, graviditet og sårheling må proteinsyntesen overstige nedbrytningen. Kroppen mister kontinuerlig noen proteiner; disse er proteinene i hår, negler og overflatelaget på huden. Derfor, for syntese av proteiner, må hver organisme motta aminosyrer fra mat.
Kilder til aminosyrer.
Grønne planter syntetiserer alle de 20 aminosyrene som finnes i proteiner fra CO2, vann og ammoniakk eller nitrater. Mange bakterier er også i stand til å syntetisere aminosyrer i nærvær av sukker (eller noe tilsvarende) og fiksert nitrogen, men sukker leveres til slutt av grønne planter. Hos dyr er evnen til å syntetisere aminosyrer begrenset; de får aminosyrer ved å spise grønne planter eller andre dyr. I fordøyelseskanalen brytes de absorberte proteinene ned til aminosyrer, sistnevnte absorberes, og proteinene som er karakteristiske for den gitte organismen bygges av dem. Ingen av det absorberte proteinet er inkorporert i kroppsstrukturer som sådan. Det eneste unntaket er at hos mange pattedyr kan en del av mors antistoffer passere intakt gjennom morkaken inn i fosterets sirkulasjon, og gjennom morsmelk (spesielt hos drøvtyggere) overføres til den nyfødte umiddelbart etter fødselen.
Behov for proteiner.
Det er klart at for å opprettholde livet må kroppen få en viss mengde protein fra maten. Størrelsen på dette behovet avhenger imidlertid av en rekke faktorer. Kroppen trenger mat både som energikilde (kalorier) og som materiale for å bygge strukturene. I første omgang er behovet for energi. Dette betyr at når det er lite karbohydrater og fett i kosten, brukes diettproteiner ikke til syntese av deres egne proteiner, men som en kilde til kalorier. Ved langvarig faste brukes til og med dine egne proteiner for å dekke energibehovet. Hvis det er nok karbohydrater i kosten, kan proteininntaket reduseres.
nitrogenbalanse.
I gjennomsnitt ca. 16 % av den totale proteinmassen er nitrogen. Når aminosyrene som utgjør proteiner brytes ned, skilles nitrogenet som finnes i dem ut fra kroppen i urinen og (i mindre grad) i avføringen i form av ulike nitrogenholdige forbindelser. Derfor er det praktisk å bruke en slik indikator som nitrogenbalanse for å vurdere kvaliteten på proteinernæring, dvs. forskjellen (i gram) mellom mengden nitrogen som tas inn i kroppen og mengden nitrogen som skilles ut per dag. Med normal ernæring hos en voksen er disse mengdene like. I en voksende organisme er mengden av utskilt nitrogen mindre enn mengden innkommende, dvs. balansen er positiv. Ved mangel på protein i kosten er balansen negativ. Hvis det er nok kalorier i kosten, men proteinene er helt fraværende i det, sparer kroppen proteiner. Samtidig bremses proteinmetabolismen, og gjenbruken av aminosyrer i proteinsyntesen går så effektivt som mulig. Tap er imidlertid uunngåelig, og nitrogenholdige forbindelser skilles fortsatt ut i urinen og delvis i avføringen. Mengden nitrogen som skilles ut fra kroppen per dag under proteinsult kan tjene som et mål på den daglige mangelen på protein. Det er naturlig å anta at ved å innføre i kosten en mengde protein tilsvarende denne mangelen, er det mulig å gjenopprette nitrogenbalansen. Det er det imidlertid ikke. Etter å ha mottatt denne mengden protein, begynner kroppen å bruke aminosyrer mindre effektivt, så noe ekstra protein er nødvendig for å gjenopprette nitrogenbalansen.
Hvis mengden protein i kosten overstiger det som er nødvendig for å opprettholde nitrogenbalansen, ser det ut til at det ikke er noen skade av dette. Overflødige aminosyrer brukes ganske enkelt som en energikilde. Et spesielt slående eksempel er eskimoen, som inntar lite karbohydrater og omtrent ti ganger mer protein enn det som kreves for å opprettholde nitrogenbalansen. I de fleste tilfeller er det imidlertid ikke gunstig å bruke protein som energikilde, siden du kan få i deg mange flere kalorier fra en gitt mengde karbohydrater enn fra samme mengde protein. I fattige land får befolkningen de nødvendige kaloriene fra karbohydrater og inntar en minimumsmengde protein.
Hvis kroppen får det nødvendige antall kalorier i form av ikke-proteinprodukter, er minimumsmengden protein som opprettholder nitrogenbalansen ca. 30 g per dag. Omtrent like mye protein finnes i fire brødskiver eller 0,5 liter melk. En litt større mengde anses vanligvis som optimal; anbefalt fra 50 til 70 g.
Essensielle aminosyrer.
Til nå har protein blitt sett på som en helhet. I mellomtiden, for at proteinsyntese skal finne sted, må alle nødvendige aminosyrer være tilstede i kroppen. Noen av aminosyrene er kroppen til dyret selv i stand til å syntetisere. De kalles utskiftbare, siden de ikke trenger å være tilstede i kosten - det er bare viktig at generelt inntak av protein som nitrogenkilde er tilstrekkelig; da, med mangel på ikke-essensielle aminosyrer, kan kroppen syntetisere dem på bekostning av de som er tilstede i overkant. De gjenværende "essensielle" aminosyrene kan ikke syntetiseres og må inntas sammen med mat. Viktige for mennesker er valin, leucin, isoleucin, treonin, metionin, fenylalanin, tryptofan, histidin, lysin og arginin. (Selv om arginin kan syntetiseres i kroppen, anses det som en essensiell aminosyre fordi nyfødte og voksende barn produserer utilstrekkelige mengder av det. På den annen side, for en person i moden alder, inntak av noen av disse aminosyrene fra mat kan bli valgfritt.)
Denne listen over essensielle aminosyrer er omtrent den samme hos andre virveldyr og til og med hos insekter. Ernæringsverdien til proteiner bestemmes vanligvis ved å mate dem til voksende rotter og overvåke vektøkningen til dyrene.
Den ernæringsmessige verdien av proteiner.
Næringsverdien til et protein bestemmes av den essensielle aminosyren som er mest mangelfull. La oss illustrere dette med et eksempel. Proteinene i kroppen vår inneholder i gjennomsnitt ca. 2 % tryptofan (i vekt). La oss si at kostholdet inneholder 10 g protein som inneholder 1% tryptofan, og at det er nok andre essensielle aminosyrer i den. I vårt tilfelle tilsvarer 10 g av dette defekte proteinet i hovedsak 5 g av et komplett; de resterende 5 g kan bare tjene som en energikilde. Merk at siden aminosyrer praktisk talt ikke er lagret i kroppen, og for at proteinsyntese skal finne sted, må alle aminosyrer være tilstede samtidig, kan effekten av inntak av essensielle aminosyrer bare oppdages hvis alle kommer inn i kroppen på samme tid.
Den gjennomsnittlige sammensetningen av de fleste animalske proteiner er nær den gjennomsnittlige sammensetningen av proteinene i menneskekroppen, så vi vil neppe møte aminosyremangel hvis kostholdet vårt er rikt på mat som kjøtt, egg, melk og ost. Imidlertid er det proteiner, for eksempel gelatin (et produkt av kollagen denaturering), som inneholder svært få essensielle aminosyrer. Vegetabilske proteiner, selv om de er bedre enn gelatin i denne forstand, er også fattige på essensielle aminosyrer; spesielt lite i dem lysin og tryptofan. Likevel er et rent vegetarisk kosthold slett ikke skadelig, med mindre det forbruker en litt større mengde vegetabilske proteiner, tilstrekkelig til å gi kroppen essensielle aminosyrer. Mest protein finnes i planter i frøene, spesielt i frøene til hvete og ulike belgfrukter. Unge skudd, som asparges, er også rike på protein.
Syntetiske proteiner i kosten.
Ved å tilsette små mengder syntetiske essensielle aminosyrer eller proteiner rike på dem til ufullstendige proteiner, som for eksempel maisproteiner, er det mulig å øke næringsverdien til sistnevnte betydelig, dvs. og dermed øke mengden protein som konsumeres. En annen mulighet er å dyrke bakterier eller gjær på petroleumshydrokarboner med tilsetning av nitrater eller ammoniakk som nitrogenkilde. Det mikrobielle proteinet oppnådd på denne måten kan tjene som fôr til fjørfe eller husdyr, eller kan konsumeres direkte av mennesker. Den tredje, mye brukte metoden bruker drøvtyggeres fysiologi. Hos drøvtyggere, i den innledende delen av magen, den såkalte. Vom er bebodd av spesielle former for bakterier og protozoer som omdanner defekte planteproteiner til mer komplette mikrobielle proteiner, og disse blir igjen etter fordøyelse og absorpsjon til animalske proteiner. Urea, en billig syntetisk nitrogenholdig forbindelse, kan tilsettes husdyrfôr. Mikroorganismer som lever i vomma bruker ureanitrogen til å omdanne karbohydrater (som det er mye mer av i fôret) til protein. Omtrent en tredjedel av alt nitrogen i husdyrfôr kan komme i form av urea, som i hovedsak betyr til en viss grad kjemisk proteinsyntese.
 Hvordan komme i gang som veileder?
Hvordan komme i gang som veileder? Fjernundervisning i engelsk
Fjernundervisning i engelsk Vanlige og uregelmessige engelske verb
Vanlige og uregelmessige engelske verb