Se hva "bark" er i andre ordbøker. Barkhøsting på begynnelsen av 1900-tallet
Cerebral cortex - ytre lag nervevev i hjernen til mennesker og andre pattedyrarter. Hjernebarken er delt av en langsgående sprekk (lat. Fissura longitudinalis) i to store deler, som kalles hjernehalvdelene eller halvkulene – høyre og venstre. Begge halvkulene er forbundet nedenfor med corpus callosum (lat. Corpus callosum). Cerebral cortex spiller en nøkkelrolle i utførelsen av hjernefunksjoner som hukommelse, oppmerksomhet, persepsjon, tenkning, tale, bevissthet.
Hos store pattedyr er hjernebarken satt sammen til mesenterier, noe som gir opphav til stort område overflaten i samme volum av skallen. Krusningene kalles viklinger, og mellom dem ligger furer og dypere - sprekker.
To tredjedeler av den menneskelige hjernen er skjult i riller og sprekker.
Cerebral cortex har en tykkelse på 2 til 4 mm.
Cortex er dannet av grå substans, som hovedsakelig består av cellelegemer, hovedsakelig astrocytter, og kapillærer. Derfor, selv visuelt, skiller det kortikale vevet seg fra den hvite substansen, som ligger dypere og består hovedsakelig av hvite myelinfibre - aksonene til nevroner.
Den ytre delen av cortex, den såkalte neocortex (lat. Neocortex), den mest evolusjonært unge delen av cortex hos pattedyr, har opptil seks cellelag. Nevroner av forskjellige lag er sammenkoblet i kortikale minikolonner. Ulike områder av cortex, kjent som Brodmanns områder, skiller seg fra hverandre i cytoarkitektonikk (histologisk struktur) og funksjonell rolle i sensitivitet, tenkning, bevissthet og erkjennelse.

Utvikling
Cerebral cortex utvikler seg fra den embryonale ektodermen, nemlig fra den fremre delen av nevrale platen. Nevralplaten folder seg og danner nevralrøret. Det ventrikulære systemet oppstår fra hulrommet inne i nevralrøret, og nevroner og glia oppstår fra epitelcellene i veggene. Fra den fremre delen av nevraleplaten dannes forhjernen, hjernehalvdelene og deretter cortex
Vekstsonen til kortikale nevroner, den såkalte "S"-sonen, ligger ved siden av ventrikkelsystemet i hjernen. Denne sonen inneholder stamceller som senere i differensieringsprosessen blir til gliaceller og nevroner. Glialfibre, dannet i de første divisjonene av forløperceller, er radialt orientert, spenner over tykkelsen av cortex fra ventrikkelsonen til pia mater (lat. Pia mater) og danner "skinner" for migrering av nevroner utover fra ventrikkelen. sone. Disse datternervecellene blir pyramideceller i cortex. Utviklingsprosessen er tydelig regulert i tid og styres av hundrevis av gener og energireguleringsmekanismer. Under utviklingen dannes en lag-for-lag-struktur av cortex.

Kortikal utvikling mellom 26 og 39 uker (menneskelig embryo)
Cellelag
Hvert av cellelagene har en karakteristisk tetthet nerveceller og forbindelser med andre områder. Det er direkte forbindelser mellom ulike områder av cortex og indirekte forbindelser, for eksempel gjennom thalamus. Et typisk mønster for kortikal laminering er stripen av Gennari i den primære visuelle cortex. Denne tråden er visuelt hvitere enn vevet, synlig for det blotte øye ved bunnen av calcarine-rillen (lat. Sulcus calcarinus) i occipitallappen (lat. Lobus occipitalis). Streken til Gennari består av aksoner som bærer visuell informasjon fra thalamus til det fjerde laget av visuell cortex.
Farging av kolonner av celler og deres aksoner tillot nevroanatomer på begynnelsen av det tjuende århundre. lage en detaljert beskrivelse av lag-for-lag-strukturen til cortex ulike typer. Etter arbeidet til Corbinian Brodmann (1909), ble nevroner i cortex gruppert i seks hovedlag - fra de ytre, ved siden av pia mater; til de interne, som grenser til den hvite substansen:
- Lag I, det molekylære laget, inneholder noen få spredte nevroner og består hovedsakelig av vertikalt (apikalt) orienterte dendritter av pyramidale nevroner og horisontalt orienterte aksoner og gliaceller. Under utviklingen inneholder dette laget Cajal-Retzius-celler og subpialceller (celler som ligger rett under det granulære laget. Ryggastrocytter finnes også noen ganger her. De apikale tuftene av dendritter antas å ha stor verdi for gjensidige forbindelser (" tilbakemelding") i hjernebarken, og er involvert i funksjonene assosiativ læring og oppmerksomhet.
- Lag II, det ytre granulære laget, inneholder små pyramidale nevroner og tallrike stjernenevroner (hvis dendritter kommer fra forskjellige sider cellekropp, danner en stjerneform).
- Lag III, det ytre pyramidale laget, inneholder overveiende små og mellomstore pyramidale og ikke-pyramidale nevroner med vertikalt orienterte intrakortikale (de i cortex). Cellelag I til III er hovedmålene for intrapulmonale afferenter, og lag III er hovedkilden til kortiko-kortikale forbindelser.
- Lag IV, det indre granulære laget, inneholder ulike typer pyramidale og stjerneformede nevroner og fungerer som hovedmålet for thalamokortikale (thalamus til cortex) afferenter.
- Lag V, det indre pyramidale laget, inneholder store pyramidale nevroner, hvis aksoner forlater cortex og stikker ut til subkortikale strukturer (som basalgangliene. I den primære motoriske cortex inneholder dette laget Betz-celler, hvis aksoner strekker seg gjennom den indre kapselen, hjernestammen og ryggmargen og danner den kortikospinale banen, som kontrollerer frivillige bevegelser.
- Lag VI, det polymorfe eller multiforme laget, inneholder få pyramidale nevroner og mange polymorfe nevroner; Efferente fibre fra dette laget går til thalamus, og etablerer en omvendt (resiprok) forbindelse mellom thalamus og cortex.

Den ytre overflaten av hjernen, som områdene er utpekt på, forsynes med blod av cerebrale arterier. Området indikert i blått tilsvarer den fremre cerebrale arterien. Delen av den bakre cerebrale arterien er indikert med gult
De kortikale lagene er ikke bare stablet én på én. Det er karakteristiske forbindelser mellom de ulike lagene og celletypene i dem som gjennomsyrer hele tykkelsen av cortex. Den grunnleggende funksjonelle enheten til cortex regnes for å være den kortikale minisøylen (en vertikal kolonne av nevroner i hjernebarken som går gjennom lagene. Minikolonnen inkluderer fra 80 til 120 nevroner i alle områder av hjernen unntatt den primære visuelle cortex av primater).
Områder av cortex uten det fjerde (interne granulære) laget kalles agranulært de med et rudimentært granulært lag. Hastigheten på informasjonsbehandlingen innenfor hvert lag er forskjellig. Så i II og III er det tregt, med en frekvens (2 Hz), mens i lag V er oscillasjonsfrekvensen mye raskere - 10-15 Hz.
Kortikale soner
Anatomisk kan cortex deles inn i fire deler, som har navn som tilsvarer navnene på hodeskallebenene som dekker:
- Frontallappen (hjerne), (lat. Lobus frontalis)
- Temporallapp, (lat. Lobus temporalis)
- Parietallapp, (lat. Lobus parietalis)
- Occipitallapp, (lat. Lobus occipitalis)
Med tanke på egenskapene til den laminære (lag-for-lag) strukturen, er cortex delt inn i neocortex og alocortex:
- Neocortex (lat. Neocortex, andre navn - isocortex, lat. Isocortex og neopallium, lat. Neopallium) er en del av den modne hjernebarken med seks cellelag. Eksempler på neocortex er Brodmann Area 4, også kjent som primær motorisk cortex, primær visuell cortex eller Brodmann Area 17. Neocortex er delt inn i to typer: isocortex (den sanne neocortex, eksempler på Brodmann Areas 24, 25 og 32) er kun omtalt) og prosocortex, som spesielt er representert ved Brodmann område 24, Brodmann område 25 og Brodmann område 32
- Alocortex (lat. Allocortex) - en del av cortex med antall cellelag mindre enn seks, er også delt i to deler: paleocortex (lat. Paleocortex) med tre lag, archicortex (lat. Archicortex) på fire til fem, og den tilstøtende perialocortex (lat. periallocortex). Eksempler på områder med en slik lagdelt struktur er olfaktorisk cortex: hvelvet gyrus (lat. Gyrus fornicatus) med kroken (lat. Uncus), hippocampus (lat. Hippocampus) og strukturer nær denne.
Det er også en "overgangs" (mellom alocortex og neocortex) cortex, som kalles paralimbisk, hvor cellelag 2,3 og 4 smelter sammen. Denne sonen inneholder proisocortex (fra neocortex) og perialocortex (fra alocortex).

Cerebral cortex. (ifølge Poirier fr. Poirier.). Livooruch - grupper av celler, til høyre - fibre.
Paul Brodmann
Ulike områder av cortex er involvert i å utføre forskjellige funksjoner. Du kan se og registrere denne forskjellen på ulike måter- identifisere lesjoner i visse områder, sammenligne mønstre av elektrisk aktivitet, bruke nevroavbildningsteknikker, studere cellulær struktur. Basert på disse forskjellene klassifiserer forskerne kortikale områder.
Den mest kjente og siterte i et århundre er klassifiseringen som ble opprettet i 1905-1909 av den tyske forskeren Corbinian Brodmann. Han delte hjernebarken inn i 51 regioner basert på cytoarkitekturen til nevroner, som han studerte i hjernebarken ved å bruke Nissl-farging av celler. Brodmann publiserte sine kart over kortikale områder hos mennesker, aper og andre arter i 1909.
Brodmanns felt har blitt aktivt og i detalj diskutert, diskutert, avklart og omdøpt i nesten et århundre og er fortsatt de mest kjente og ofte siterte strukturene i den cytoarkitektoniske organiseringen av den menneskelige hjernebarken.
Mange av Brodmann-feltene, opprinnelig definert utelukkende av deres nevronorganisering, ble senere assosiert med korrelasjon med forskjellige kortikale funksjoner. For eksempel er felt 3, 1 og 2 den primære somatosensoriske cortex; område 4 er den primære motoriske cortex; felt 17 er primær visuell cortex, og felt 41 og 42 er mer korrelert med primær auditiv cortex. Å bestemme korrespondansen mellom prosessene med høyere nervøs aktivitet til områder av hjernebarken og koble dem til spesifikke Brodmann-felt utføres ved hjelp av nevrofysiologiske studier, funksjonell magnetisk resonansavbildning og andre teknikker (da dette for eksempel ble gjort med å koble Brocas områder av tale og språk til Brodmann felt 44 og 45). Funksjonell avbildning kan imidlertid bare omtrentlig bestemme lokaliseringen av hjerneaktivering i Brodmanns felt. Og for å nøyaktig bestemme grensene deres i hver enkelt hjerne, er det nødvendig med en histologisk undersøkelse.

Noen av de viktige Brodmann-feltene. Hvor: Primær somatosensorisk cortex - primær somatosensorisk cortex Primær motorisk cortex - primær motorisk (motorisk) cortex; Wernicke’s area – Wernicke’s area; Primært visuelt område - primært visuelt område; Primær auditiv cortex - primær auditiv cortex; Brocas område - Brocas område.
Barktykkelse
Hos pattedyrarter med store hjernestørrelser (i absolutte termer, ikke bare i forhold til kroppsstørrelsen), har cortex en tendens til å være tykkere. Utvalget er imidlertid ikke veldig stort. Små pattedyr som spissmus har en neocortex-tykkelse på omtrent 0,5 mm; og arter med de største hjernene, som mennesker og hvaler, er 2,3-2,8 mm tykke. Det er en omtrent logaritmisk sammenheng mellom hjernevekt og kortikal tykkelse.
Magnetisk resonansavbildning (MRI) av hjernen gjør det mulig å måle intravital kortikal tykkelse og korrelere den med kroppsstørrelse. Tykkelsen på forskjellige områder varierer, men generelt er de sensoriske (sensitive) områdene i cortex tynnere enn de motoriske (motoriske) områdene. En studie viste avhengigheten av kortikal tykkelse på intelligensnivå. En annen studie viste større kortikal tykkelse hos migrenepasienter. Andre studier viser imidlertid fraværet av en slik sammenheng.
Konvolusjoner, riller og sprekker
Sammen skaper disse tre elementene - konvolusjoner, sulci og sprekker - et stort overflateareal av hjernen til mennesker og andre pattedyr. Når man ser på menneskelig hjerne, er det merkbart at to tredjedeler av overflaten er skjult i sporene. Både riller og sprekker er fordypninger i cortex, men de varierer i størrelse. Sulcus er et grunt spor som omgir gyri. Fissuren er en stor rille som deler hjernen i deler, samt i to halvkuler, for eksempel den mediale langsgående sprekken. Dette skillet er imidlertid ikke alltid entydig. For eksempel er den laterale fissuren også kjent som den laterale fissuren og som den "Sylvianske sprekken" og den "sentralfissuren", også kjent som den sentrale sprekken og som "Rolandic fissuren".
Dette er svært viktig i forhold der hjernestørrelsen er begrenset av den indre størrelsen på skallen. En økning i overflaten av hjernebarken ved hjelp av et system av konvolusjoner og sulci øker antallet celler som er involvert i utførelsen av hjernefunksjoner som hukommelse, oppmerksomhet, persepsjon, tenkning, tale, bevissthet.
Blodtilførsel
Spesielt tilførsel av arterielt blod til hjernen og cortex skjer gjennom to arterielle bassenger - de indre halspulsårene og vertebrale arterier. Den terminale delen av den indre halspulsåren forgrener seg til grener - de fremre cerebrale og midtre cerebrale arteriene. I de nedre (basale) delene av hjernen danner arterier en sirkel av Willis, på grunn av hvilken arterielt blod omfordeles mellom arteriebassengene.
Midt cerebral arterie
Den midtre cerebrale arterien (lat. A. Cerebri media) er den største grenen av den indre halspulsåren. Dårlig sirkulasjon i det kan føre til utvikling av iskemisk slag og midtre cerebralt arteriesyndrom med følgende symptomer:
- Lammelse, plegi eller parese av motsatte muskler i ansikt og armer
- Tap sensorisk følsomhet motsatte lesjoner av musklene i ansiktet og armene
- Skade på den dominerende halvkulen (ofte venstre) av hjernen og utvikling av Brocas afasi eller Wernickes afasi
- Skade på den ikke-dominante hjernehalvdelen (ofte til høyre) i hjernen fører til ensidig romlig agnosi på den avsides berørte siden
- Infarkter i området av den midtre cerebrale arterien fører til avvikskonjugering, når pupillene i øynene beveger seg mot siden av hjernelesjonen.
Fremre cerebral arterie
Den fremre cerebrale arterien er en mindre gren av den indre halspulsåren. Etter å ha nådd den mediale overflaten av hjernehalvdelene, går den fremre hjernearterie til occipitallappen. Den forsyner de mediale områdene av halvkulene til nivået av den parieto-occipitale sulcus, området til den øvre frontale gyrus, området til parietallappen, samt områder av de nedre mediale delene av orbital gyri . Symptomer på hennes nederlag:
- Parese av benet eller hemiparese med en dominerende lesjon av benet på motsatt side.
- Blokkering av de parasentrale grenene fører til monoparese av foten, som minner om perifer parese. Urinretensjon eller inkontinens kan forekomme. Reflekser av oral automatisme og gripende fenomener, patologiske fotbøyereflekser vises: Rossolimo, Bekhterev, Zhukovsky. Endringer i mental tilstand oppstår på grunn av skade på frontallappen: redusert kritikk, hukommelse, umotivert oppførsel.
Posterior cerebral arterie
Et sammenkoblet kar som leverer blod til de bakre delene av hjernen (occipitallappen). Har en anastomose med den midtre cerebrale arterien. Dens lesjoner fører til:
- Homonym (eller øvre kvadrant) hemianopsi (tap av en del av synsfeltet)
- Metamorfopsi (brudd visuell persepsjon størrelse eller form på objekter og rom) og visuell agnosia,
- Alexia,
- Sensorisk afasi,
- Forbigående (forbigående) hukommelsestap;
- Rørformet syn
- Kortikal blindhet (mens reaksjonen på lys opprettholdes),
- Prosopagnosia,
- Desorientering i rommet
- Tap av topografisk hukommelse
- Ervervet achromatopsia - mangel på fargesyn
- Korsakoffs syndrom (svekket arbeidsminne)
- Emosjonelle og affektive lidelser
Eller grener som mer eller mindre lett skilles fra den indre (mye tettere) tremassen. I botanikk er bark (i treaktige planter) samlingen av forskjellige vev som ligger mellom huden og den kambiale ringen. Cellene i kambiumet er veldig delikate, de rives lettere enn andre, og derfor rives barken lett av på dette stedet fra det underliggende treverket.
Japansk lønnbark.

Vekster på skadestedet i et ungt asketre.
Basert på strukturen og mønsteret til barken, skilles følgende ut:
- trær med glatt bark (bøk, agnbøk, ailanthus, kirsebær);
- trær med lamellbark (kastanje, platan, eukalyptus);
- trær med fint sprukket bark (pære, lyriodendron, ginkgo);
- trær med dypt sprukket bark (stilt eik, hvit selje).
Struktur
Skjelne primær Og sekundær bark Den primære cortex representerer den perifere delen av det såkalte grunnvevet; dette er permanent vev. Sekundærbarken er født fra kambium og, takket være aktiviteten til sistnevnte, blir den gradvis tykkere [En liten fortykkelse av primærbarken oppstår bare når den såkalte kambium dannes i den. periderm. De kaller henne noe annet floem eller bast. Under etternavnet vil det bli diskutert mer detaljert.
For informasjon om den sekundære cortex, se Lubs artikkelPrimær cortex
Den primære cortex er sammensatt av parenkymceller, som ofte inneholder klorofyll og stivelseskorn. I den er det på sin side ofte mulig å skille mellom ytre og indre deler. Den første er ved siden av huden og består av celler som er tett lukket til hverandre, ofte omdannet til bunter av collenchyma eller sclerenchyma. Den andre delen består av tynnveggede celler, løst sammenkoblet, med små intercellulære mellomrom. Hos vann- og myrplanter når sistnevnte betydelige størrelser og smelter ofte sammen, og danner store lufthulrom og kanaler, vanligvis atskilt fra hverandre med bare ett lag med celler (intercellulære rom). Noen ganger strekker slike kanaler seg kontinuerlig over hele internoden (for eksempel hos representanter for slektene Hippuris, Ceratophyllum, Myriophyllum), i andre tilfeller blir de avbrutt av tverrgående skillevegger ( Marsilia, Potamogeton, Butomus osv.).
Endoderm
Det innerste laget av celler i den primære cortex danner den såkalte. endoderm, ellers beskyttende eller beskyttende lag eller beskyttende vagina, omslutter hele den sekundære cortex, og samtidig hele systemet av vaskulære-fibrøse bunter eller, i terminologien til Fan-Tighem og hans tilhengere, den "sentrale sylinderen". Endoderm finnes i stengler og røtter, men i sistnevnte kommer det til uttrykk mye mer fremtredende. Endodermale celler har en spesiell struktur: på deres radielle vegger (som de er tett ved siden av hverandre) er karakteristiske bølgete folder merkbare, som i tverrsnitt vises i form av spesielt typiske mørke flekker (Caspari-flekker). Til å begynne med blir tynne, rene cellulosecellevegger gradvis kutikulære; i dette tilfellet begynner prosessen med cuticularization med foldene som nettopp er nevnt og dekker til slutt (i de fleste tilfeller) hele skallet. Tettpakkede celler skiller systemet til den primære cortex fra systemet av vaskulære bunter, og tjener som en "beskyttende kappe" for sistnevnte. Dette har betydelig fysiologisk betydning. Atmosfærisk luft, som penetrerer gjennom stomata inn i de intercellulære hulrommene i primærbarken, kan, takket være endodermen, fra sistnevnte komme inn i vaskulærbuntenes vannveier og her forstyrre bevegelsen av vann. Av samme grunn kan luften i de histologiske elementene i karbuntene ha en annen sammensetning og være under et annet trykk enn atmosfæren som omgir planten. Endodermis forhindrer imidlertid ikke fri utveksling av vann og stoffer oppløst i den mellom primærbarken og "sentralsylinderen" (i roten strømmer for eksempel vann fritt til sistnevnte, absorbert av rothår). Ofte, over tid, blir membranene til endodermale celler mer eller mindre kraftig fortykkede og treaktige, vanligvis mer på de indre veggene enn på de laterale (radiale) veggene. Men i noen celler forblir membranene tynne, tykner ikke og er ikke kutikulære: under bevegelse av stoffer spiller slike celler rollen som passasjer eller inngangsporter (Durchlasszellen) i endodermen. Bølgefolder på de radielle veggene, kutikularisering av membranen og rikelig akkumulering av stivelse er tegnene som skiller cellene i den beskyttende skjeden fra naboene. I luftstammer, hvor endodermen, som allerede nevnt, ikke kommer så tydelig til uttrykk som i røttene, er en stor ansamling av stivelse et spesielt typisk trekk, på grunn av hvilket endoderm ofte kalles stivelse i skjeden. I følge Sachs og noen andre botanikere skjer bevegelsen av stivelse gjennom hele planten hovedsakelig gjennom denne skjeden.
Fotogalleri
Bruk
I følge tsjekkisk forskning fra Rectoris er prosentandelen bark av den totale vedmassen:
Eikegrener gir (ifølge Grebe) 8-10 % mer bjeff enn stammer. Hvor mye det gitte forholdet endres med størrelsen på trærne kan sees fra Walters Hessian-studier: for innsatser varierer det mellom 15,5-28,6 %, et gjennomsnitt på 27,2 %, abbor (0,1-0,2 kubikkmeter) 10, 2-14,5 %, snitt 12,0, tynne stammer (0,2-0,7 kubikkmeter) 7,0-15,2 %, snitt 11,6 og store stammer (0,7-2,0 kubikkm) 7,7-12,4 %, snitt 9,8. I eikebark (ifølge Grebe) 24,3 volumprosent av totalmassen; glatt "speil", 21,4% bra sprukket og 16,8% grov, lav verdi.
I de fleste tilfeller har bark nesten ingen verdi, selv om det noen ganger fungerer som et materiale for forskjellige produkter - det er utsatt for mekanisk eller kjemisk behandling; Således brukes bark av eik, mange vier, gran, lerk og delvis bjørk (egentlig "grønn" uten bjørkebark, i Sibir og Norge) og or som "bark-eik", "tauben" eller "eik" ved garving skinn (se. Tanning materialer) og deretter, etter bløtlegging, drar til utlandet for å forberede briketter.
Barken til korkeiken gir korken; Bjørkebark er et materiale for en lang rekke produkter og for utvinning av tjære. Barken av lind, vier og alm gir bast til bastproduksjon og i tillegg den første - bast til matter og sekker, skinne til diverse håndverk. Overflate for selvflyter og not er laget av sargbark. Tak laget av furubark (Novgorod-provinsen) er mye bedre enn tekke; granbark brukes som underlag for shingeltak. Salicin utvinnes fra pilbark (Salix purpurea og Salix helix), det lages tau til not osv. Al- og valnøttbark egner seg til å male svart og brunt; fra kost bast ((Citysus laburnum og (Citysus alpinus) vakre hatter osv. veves i Italia.
I Nordvest-Russland varierer andelen granbark fra 9-10,5 %.
Innhøsting av bark på begynnelsen av 1900-tallet
Den beste tiden for å høste bark er når sevjen renner helt i treet, når den lett henger etter veden - "fjernet med en strømpe" eller i ett stykke. «Barking» eller «sliping» utføres, avhengig av formålet med å høste barken, både på felte og voksende trær, for eksempel ved høsting av bast, bjørkebark, eik og til bast kan du bruke trær som står på roten , lage langsgående og tverrgående snitt bark med en øks eller en "peshka" - en jernmeisel på et langt skaft - og rive barken av treverket med en øks eller en enkel pinne med et spiss skrått kutt i enden, eller direkte med hendene dine. Ved avrivning av en dobbelbark skilles den ikke fra resten av barken med et øvre snitt, men får henge og tørke på stammen, under beskyttelse av trekronen. I voksende trær fjernes barken bare fra stammen, mens den på grener og kvister, ofte mer verdifulle i sine kvaliteter, forblir og blir bortkastet uproduktivt. Bjørkebark for produkter, lindved til skinne, og furu og gran til tak høstes mer hensiktsmessig fra felte trær på samme måte som fra voksende trær, og verktøyene som brukes til å fjerne eikeved, generelt lignende, finnes i Tyskland og Østerrike -Ungarn veldig forskjellig størrelse, form og type laget av jern, tre og bein og navn. eikeskjeer, kuttere, meisler o.l.
For å redusere kostnadene ved å fjerne bark og produsere den om vinteren, når en arbeidsdag koster mindre, ble kunstig barking ved hjelp av damp oppfunnet. Metraapparat er en type dampmaskin der damp fra en del av en horisontal kjele går over til en annen, hvor materialet som skal avbarkes legges i en boks; Etter damping skiller barken seg lett fra veden. Enda bedre er et annet Nosmaison-apparat, bestående av en bevegelig kjelsylinder på to hjul, på bunnen av hvilken damp produseres, som etter overoppheting i den øvre delen til 200-250 ° kommer inn i tønnene der materialet som skal avbarkes er plassert. Opptil 1000 kg bark ryddes per dag. Det ble også brukt Morges apparat, som representerer besparelser i arbeid sammenlignet med manuell barking, i forholdet 1:6.
I følge Grebes undersøkelser, jo demper plassering, jo mindre er barken i volum, men den er tyngre. Likeledes er mengden eikejord den samme på sand- og leirholdig jord, men på førstnevnte er den tyngre. Kvaliteten på opptaket påvirkes
- type jordbruk (lavtvoksende, spesielt i form av skogskornoppdrett, fortjener preferanse);
- Skjæringsomsetningen, som bør være nær debutalderen for størst årlig vekst og for å få K. alene, er 14-20 år. I dette tilfellet er inntekten fra å kutte tiende i gjennomsnitt 515 pund. og 13 cu. sot;
- fullstendighet av plantinger - rettidig, i riktig grad, rydding, tynning og rydding;
- sammensetningen av beplantningen: med en betydelig blanding av mindre treslag er eiketre tynnere og derfor oppnås mindre fra hvert tre. I følge Neubrand øker inntekten i snitt til 21 % ved overgang fra en blandet beplantning til en ren, og på grunn av påfyll av plantingen, etter hogst, med eik, er det mulig å få K. med en gjennomsnittlig felling på 50 % (Eickmeyer). Tvert imot, når selv et svært moderat antall trær blir stående "til gjengroing" eller i form av "fyrtårn" (gjennomsnittlig gård), reduseres inntekten i form av kontanter med 30-35%. Med den mest forsiktige bevaringen av eiken endres tannininnholdet i den, selv i løpet av kort tid, betydelig. Generelt er «barkoppdrett» i eikeskog en av de mest lønnsomme, og er i stand til å gi, selv på dårlig jord (Holland), opptil 7 % av kapitalen som er bundet til den. Selv om prisene på eikebark på slutten av åttitallet falt kraftig i utlandet, på grunn av introduksjonen av et surrogat der - garving av skinn med mineralske stoffer, men for tiden begynner etterspørselen etter barkbark å øke igjen [I Tyskland, 410.000 des. . under eikeskoger, men for å tilfredsstille hele Europas behov for eikeskog, kreves et areal med eikeskog på ca. 35 000 0 0 des.].
ons. Hohenstein, "Die Eichensch äl-Wirthschaft" (); Tribolin, "Der Eichenschä lwaldbetrieb" (); F v. Hohnel, "Die Gerberinden" (); F. Arnold, "Dublo og eikeskoger" ("Journal of Manufacture and Trade", VI); V. Sobichevsky, "På returen av lavstammede eiketrær for å oppnå en dobbel for garving av skinn" ("Notes of the Imperial General. Agricultural and Economic. Southern Russia", vol. II); M. Yoelson, "Om tilbakekomsten av lavstammede eiketrær for å oppnå speilkvalitet for garving," ("Forest Journal"). Willow K. - "zadubye" eller "skorpe" (St. Petersburg-provinsen), "kvast" - brukes i vårt land hovedsakelig som en dobbel, av og til - som et dekorativt materiale for noe flettverk, for eksempel. bast sko; En skikkelig seljebarkgård er ennå ikke etablert. Det er tre varianter av pil i handelen (Penza-provinsen): ramen eller skogbark - den høyeste karakteren, høstet fra hårete eller hårete vier (Salix caprea S. aurita, S. cinerea); høy eller grøftbark - K. høye eller pilepiler (S. viminalis, S. fragliis, S. pentandra, etc.) med et glatt blad, og pilbark; sistnevnte har en rødlig farge og en brun underside - "fôret" - og derfor gjenkjennes dens blanding med de høyeste grader av gråhvit farge med hvit fôr, ofte praktisert av handelsmenn, lett for øyet. I mangel av presise data kan mengden pil vi forbruker estimeres omtrentlig; for eksempel fra Finland alene bringes den årlig til St. Petersburg. mer enn en halv million pund. Eksporten av «treved», hovedsakelig eik, fra Russland til utlandet er ubetydelig; eksportert: by - 57064 pd. (44135 r.), by - 55232 p.d. (RUR 34.102) og by - 84.300 p.d. (57 935 RUB). Det er sannsynlig at de ovennevnte vekstforholdene for plantinger også påvirker kvaliteten på bjørkebark og lind K., selv om de ennå ikke er studert mye. Om regnskapsføring av bjørkebark, se bjørkebark. Linden K. fjernes både fra unge trær og fra middelaldrende og gamle trær, men dens tekniske kvaliteter, og derfor bruken, er forskjellige. For å samle unge K. - "bast", "lukon" eller "smychin" - fjernes K. fra voksende trær, og etterlater nakne stengler ved røttene, eller de blir først kuttet ned. Bast skilles: a) i henhold til høstingstidspunktet: førstekjerne - strippet om våren - og dranet, strippet om vinteren; b) etter type arbeidsstykke: "rør" - solid K., fjernet fra hele stammen, og "stripet" eller "stripe" - fjernet i strimler eller "bånd". Regnskap for bast ved salg er svært variert i forskjellige provinser; her må vi begrense oss til å bare sitere noen navn: i grupper (Minsk, Volyn) - 60 bunter på 40 bast hver (Ryaz. lepper); "politi": 60 "oppheng", 30 "vridninger" hver; rullet opp i "tusenvis" (Penz.), "hundrevis" (nord-vestlige og vestlige provinser, Vladim.), med forskjeller i varianter, "bunter" (Vladimirskaya), "noshami" og "noshitsy" - i 100 stykker ( Vitebsk, Tula), 60 store eller 100 små - to "polunochki". Noen ganger selges bast etter vekt - i pund. Fra kube favner liten lindeved er 40 pd. rå bast, og fra børsteved - 15 pund. I kubikk sazhen lyk (veier 71 pd.) 104 bunter, 30 stykker i hver (Vladim.). Av de middelaldrende 40-50 åringene - "hage" - er linden ennå ikke helt grov K. (faktisk
Moderne forskere vet med sikkerhet at takket være hjernens funksjon, evner som bevissthet om signaler som mottas fra ytre miljø, mental aktivitet, memorering av tenkning.
Evnen til et individ til å realisere sine egne forhold til andre mennesker er direkte relatert til prosessen med eksitasjon av nevrale nettverk. Dessuten vi snakker om spesielt om de nevrale nettverkene som er lokalisert i cortex. Den representerer det strukturelle grunnlaget for bevissthet og intelligens.
I denne artikkelen skal vi se på hvordan hjernebarken er bygget opp i detalj.
Neocortex
Cortex inneholder omtrent fjorten milliarder nevroner. Det er takket være dem at hovedsonene fungerer. De aller fleste nevroner, opptil nitti prosent, danner neocortex. Det er en del av den somatiske NS og dens høyeste integrerende avdeling. De viktigste funksjonene Hjernebarken er ansvarlig for oppfatning, bearbeiding og tolkning av informasjon som en person mottar ved hjelp av ulike sanser.
I tillegg styrer neocortex komplekse bevegelser muskelsystemer i menneskekroppen. Den inneholder sentre som deltar i prosessen med tale, minnelagring og abstrakt tenkning. De fleste prosessene som skjer i den danner det nevrofysiske grunnlaget for menneskelig bevissthet.
Hvilke andre deler består hjernebarken av? Vi vil vurdere områdene av hjernebarken nedenfor.
Paleocortex
Det er en annen stor og viktig del av cortex. Sammenlignet med neocortex har paleocortex en enklere struktur. Prosessene som foregår her gjenspeiles sjelden i bevisstheten. De høyere vegetative sentrene er lokalisert i denne delen av cortex.
Forbindelse av cortex med andre deler av hjernen
Det er viktig å vurdere sammenhengen som eksisterer mellom de underliggende delene av hjernen og hjernebarken, for eksempel med thalamus, pons, mediale pons og basalganglia. Denne forbindelsen utføres ved hjelp av store bunter av fibre som danner den interne kapselen. Bunter av fibre er representert av brede lag, som er sammensatt av hvitt stoff. De inneholder et stort antall nervefibre. Noen av disse fibrene gir overføring av nervesignaler til cortex. Resten av buntene overfører nerveimpulser til nervesentrene som ligger under.
Hvordan er hjernebarken bygget opp? Områdene i hjernebarken vil bli presentert nedenfor.
Struktur av cortex
Den største delen av hjernen er dens cortex. Dessuten er kortikale soner bare én type deler som skilles ut i cortex. I tillegg er cortex delt inn i to halvkuler - høyre og venstre. Halvkulene er forbundet med hverandre med bunter av hvit substans som danner corpus callosum. Dens funksjon er å sikre koordinering av aktivitetene til begge halvkuler.
Klassifisering av cerebrale cortex soner etter deres plassering
Til tross for at barken har et stort antall folder, i generell plassering dens individuelle viklinger og spor er konstant De viktigste er en retningslinje for å identifisere områder av cortex. Slike soner (lober) inkluderer occipital, temporal, frontal, parietal. Selv om de er klassifisert etter sted, har hver sine egne spesifikke funksjoner.
Auditiv cortex
For eksempel er den temporale sonen senteret der den kortikale delen av hørselsanalysatoren er plassert. Hvis det oppstår skade på denne delen av cortex, kan døvhet oppstå. I tillegg til dette, i auditiv sone Wernickes talesenter ligger. Hvis det er skadet, mister personen evnen til å oppfatte muntlig tale. En person oppfatter det som enkel støy. Også i tinninglappen er det nevrale sentre som tilhører det vestibulære apparatet. Hvis de blir skadet, blir balansefølelsen forstyrret.

Taleområder i hjernebarken
Taleområder er konsentrert i frontallappen av cortex. Her ligger også talemotorsenteret. Hvis det oppstår skade i høyre hjernehalvdel, mister personen evnen til å endre klangen og intonasjonen til sin egen tale, som blir monoton. Hvis det oppstår skade på talesenteret i venstre hjernehalvdel, forsvinner artikulasjonen og evnen til å artikulere tale og sang. Hva annet består hjernebarken av? Områdene i hjernebarken har forskjellige funksjoner.
Visuelle soner
I occipitallappen er det en visuell sone, der det er et senter som reagerer på synet vårt som sådan. Oppfatningen av verden rundt oss skjer nettopp med denne delen av hjernen, og ikke med øynene. Det er occipital cortex som er ansvarlig for synet, og skade på det kan føre til delvis eller fullstendig tap av synet. Det visuelle området av hjernebarken undersøkes. Hva er neste?
Parietallappen har også sine egne spesifikke funksjoner. Det er denne sonen som er ansvarlig for evnen til å analysere informasjon som er relatert til taktil, temperatur og smertefølsomhet. Hvis det oppstår skade på parietalregionen, forstyrres hjernens reflekser. En person kan ikke gjenkjenne gjenstander ved berøring.
Motorsone
La oss snakke om motorsonen separat. Det skal bemerkes at denne sonen av cortex ikke korrelerer på noen måte med lappene diskutert ovenfor. Det er en del av cortex som inneholder direkte forbindelser til motoriske nevroner i ryggmargen. Dette navnet er gitt til nevroner som direkte kontrollerer aktiviteten til musklene i kroppen.

Det viktigste motoriske området til hjernebarken er lokalisert i en gyrus kalt den presentrale gyrusen. Denne gyrusen er et speilbilde av sanseområdet i mange aspekter. Mellom dem er det kontralateral innervasjon. For å si det på en annen måte, er innerveringen rettet mot musklene som sitter på den andre siden av kroppen. Unntaket er ansiktsområdet, som er preget av bilateral muskelkontroll lokalisert på kjeven og nedre del av ansiktet.
Litt under hovedmotorsonen ligger en tilleggssone. Forskere mener at den har uavhengige funksjoner som er forbundet med prosessen med å sende ut motorimpulser. Det supplerende motoriske området er også studert av spesialister. Eksperimenter utført på dyr viser at stimulering av denne sonen provoserer forekomsten av motoriske reaksjoner. Det særegne er at slike reaksjoner oppstår selv om det motoriske hovedområdet er isolert eller fullstendig ødelagt. Den er også involvert i motorisk planlegging og talemotivasjon i den dominerende halvkulen. Forskere tror at hvis tilbehørsmotoren er skadet, kan dynamisk afasi oppstå. Hjernereflekser lider.
Klassifisering i henhold til strukturen og funksjonene til hjernebarken
Fysiologiske eksperimenter og kliniske forsøk, som ble utført på slutten av det nittende århundre, gjorde det mulig å etablere grensene mellom områdene som ulike reseptoroverflater projiseres til. Blant dem er sensoriske organer som er rettet mot omverdenen (hudfølsomhet, hørsel, syn), reseptorer innebygd direkte i bevegelsesorganene (motoriske eller kinetiske analysatorer).
Kortikale områder hvor ulike analysatorer er plassert kan klassifiseres etter struktur og funksjon. Så det er tre av dem. Disse inkluderer: primære, sekundære, tertiære soner i hjernebarken. Utviklingen av embryoet innebærer dannelse av bare primære soner, preget av enkel cytoarkitektur. Deretter kommer utviklingen av sekundære, tertiære utvikles sist. Tertiære soner er preget av de fleste kompleks struktur. La oss se litt mer detaljert på hver av dem.
Sentrale felt
Gjennom mange år med klinisk forskning har forskere klart å samle betydelig erfaring. Observasjoner gjorde det for eksempel mulig å fastslå at skade på ulike felt, innenfor de kortikale seksjonene av ulike analysatorer, kan ha en helt annen effekt på totalen. klinisk bilde. Hvis vi vurderer alle disse feltene, kan vi blant dem skille ut en som inntar en sentral posisjon i kjernefysisk sone. Dette feltet kalles sentralt eller primært. Den er lokalisert samtidig i den visuelle sonen, i den kinestetiske sonen og i den auditive sonen. Skader på primærfeltet medfører svært alvorlige konsekvenser. En person kan ikke oppfatte og utføre den mest subtile differensieringen av stimuli som påvirker de tilsvarende analysatorene. Hvordan ellers er områder av hjernebarken klassifisert?

Primærsoner
I primærsonene er det et kompleks av nevroner som er mest disponert for å gi bilaterale forbindelser mellom kortikale og subkortikale soner. Det er dette komplekset som forbinder hjernebarken med ulike sanseorganer på den mest direkte og korteste måten. I denne forbindelse har disse sonene evnen til å identifisere stimuli på en veldig detaljert måte.
Viktig fellestrekk Den funksjonelle og strukturelle organiseringen av primærområdene er at de alle har en tydelig somatisk projeksjon. Dette betyr at individuelle perifere punkter, for eksempel hudoverflater, netthinnen, skjelettmuskulaturen, cochleae i det indre øret, har egen projeksjon til strengt begrensede, korresponderende punkter som er lokalisert i de primære sonene i cortex av de tilsvarende analysatorene. I denne forbindelse fikk de navnet projeksjonssoner til hjernebarken.
Sekundære soner
På en annen måte kalles disse sonene perifere. Dette navnet ble ikke gitt dem ved en tilfeldighet. De er lokalisert i de perifere delene av cortex. Sekundære soner skiller seg fra de sentrale (primære) sonene i deres nevrale organisering, fysiologiske manifestasjoner og arkitektoniske trekk.
La oss prøve å finne ut hvilke effekter som oppstår hvis sekundærsonene påvirkes av en elektrisk stimulus eller om de er skadet. Effektene som oppstår angår hovedsakelig de fleste komplekse arter prosesser i psyken. I tilfelle det oppstår skade på sekundærsonene, forblir de elementære følelsene relativt intakte. I utgangspunktet er det forstyrrelser i evnen til riktig å reflektere gjensidige relasjoner og hele komplekser av elementer som utgjør de forskjellige objektene vi oppfatter. For eksempel, hvis de sekundære sonene til den visuelle og auditive cortex er skadet, kan fremveksten av auditive og visuelle hallusinasjoner observeres, som utspiller seg i en viss tidsmessig og romlig sekvens.

Sekundære områder er av betydelig betydning i implementeringen av gjensidige forbindelser mellom stimuli, som tildeles ved hjelp av primærområder i cortex. I tillegg spiller de en betydelig rolle i integreringen av funksjoner som utføres av kjernefysiske felter til forskjellige analysatorer som et resultat av å kombineres til komplekse komplekser av mottak.
Dermed er sekundærsoner av særlig betydning for gjennomføringen mentale prosesser i mer komplekse former som krever koordinering og er forbundet med en detaljert analyse av sammenhengene mellom objektive stimuli. Under denne prosessen etableres spesifikke forbindelser, som kalles assosiative. Afferente impulser som kommer inn i cortex fra reseptorer til ulike eksterne sanseorganer når sekundære felt gjennom mange ekstra brytere i assosiasjonskjernen til thalamus, som også kalles thalamus-optikken. Afferente impulser som går til primærsonene, i motsetning til impulser som går til sekundærsonene, når dem via en kortere rute. Det implementeres gjennom en relékjerne i den visuelle thalamus.
Vi fant ut hva hjernebarken er ansvarlig for.
Hva er thalamus?
Fibre fra thalamuskjernene når hver lapp i hjernehalvdelene. Thalamus er en visuell thalamus som ligger i den sentrale delen av forhjernen og består av stor mengde kjerner, som hver overfører impulser til visse områder av cortex.
Alle signaler som kommer inn i cortex (med unntak av olfaktoriske signaler) passerer gjennom reléet og integrerende kjerner i den visuelle thalamus. Fra kjernene i thalamus ledes fibre til sensoriske områder. Smak og somatosensoriske soner er lokalisert i parietallappen, auditiv sensorisk sone er i temporallappen, og synssonen er i occipitallappen.

Impulser til dem kommer henholdsvis fra de ventro-basale kompleksene, mediale og laterale kjerner. Motoriske områder er koblet til de ventrale og ventrolaterale kjernene i thalamus.
EEG desynkronisering
Hva skjer hvis en person som er i en tilstand av fullstendig hvile blir utsatt for en veldig sterk stimulans? Naturligvis vil en person fullt ut konsentrere seg om denne stimulansen. Overgangen av mental aktivitet, som skjer fra hviletilstand til aktivitetstilstand, reflekteres på EEG av beta-rytmen, som erstatter alfarytmen. Svingninger blir hyppigere. Denne overgangen kalles EEG-desynkronisering, den vises som et resultat av sensorisk stimulering som kommer inn i cortex fra uspesifikke kjerner i thalamus.
Aktiverer retikulært system
Det diffuse nervesystemet består av uspesifikke kjerner. Dette systemet er plassert i de mediale delene av thalamus. Det er den fremre delen av det aktiverende retikulære systemet, som regulerer excitabiliteten til cortex. En rekke sensoriske signaler kan aktiveres dette systemet. Sensoriske signaler kan være både visuelle og olfaktoriske, somatosensoriske, vestibulære, auditive. Det aktiverende retikulære systemet er en kanal som overfører signaldata til det overfladiske laget av cortex gjennom uspesifikke kjerner lokalisert i thalamus. Eksitering av ARS er nødvendig for at en person skal kunne opprettholde en tilstand av våkenhet. Hvis det oppstår forstyrrelser i dette systemet, kan det oppstå komatøse søvnlignende tilstander.
Tertiære soner
Mellom analysatorene av hjernebarken er det funksjonelle relasjoner, som har enda flere kompleks struktur enn den som er beskrevet ovenfor. Under vekstprosessen overlapper feltene til analysatorene hverandre. Slike overlappingssoner som dannes i endene av analysatorene kalles tertiære soner. De er de mest komplekse typene av å kombinere aktivitetene til de auditive, visuelle og hudkinestetiske analysatorene. Tertiære soner er plassert utenfor grensene til analysatorenes egne soner. I denne forbindelse har ikke skaden deres en uttalt effekt.

Tertiære soner er spesielle kortikale områder hvor spredte elementer fra forskjellige analysatorer samles. De okkuperer et veldig stort territorium, som er delt inn i regioner.
Den øvre parietalregionen integrerer bevegelsene til hele kroppen med den visuelle analysatoren og danner et kroppsdiagram. Den nedre parietale regionen kombinerer generaliserte former for signalering som er assosiert med differensierte objekt- og talehandlinger.
Ikke mindre viktig er den temporo-parietale-occipitale regionen. Hun er ansvarlig for den komplekse integrasjonen av auditive og visuelle analysatorer med muntlig og skriftlig tale.
Det er verdt å merke seg at sammenlignet med de to første sonene er de tertiære sonene preget av de mest komplekse interaksjonskjedene.
Hvis vi stoler på alt materialet presentert ovenfor, kan vi konkludere med at de primære, sekundære og tertiære sonene i den menneskelige cortex er høyt spesialiserte. Hver for seg er det verdt å understreke det faktum at alle tre kortikale soner som vi vurderte, i en normalt fungerende hjerne, sammen med forbindelsessystemer og subkortikale formasjoner, fungerer som en enkelt differensiert helhet.
Vi undersøkte i detalj sonene og delene av hjernebarken.
Bark
Luktende hjerne
Den utvikler seg fra den ventrale delen av telencephalon og består av to seksjoner: sentral og perifer.
Perifer avdeling (olfaktorisk lapp), plassert ved bunnen av hjernen, består av: luktepære, luktekanal, lukttrekant, fremre perforert substans.
Sentralavdeling representert ved den hvelvede gyrusen, hippocampus. , dentate gyrus.
Strukturene til telencephalon som ligger over striatum (tak, laterale og mediale vegger i laterale ventrikler) kalles regnfrakk(pallium). Det er kappen, som vokser betydelig, danner folder på overflaten og dekker nesten alle deler av hjernen. Det overfladiske laget av kappen, som består av grå substans, kalles hjernebarken. Overflatearealet til begge halvkuler er omtrent 1650 cm2. Hver halvkule har tre flater: superolateral (den mest tilgjengelige for observasjon), medial (halvkulene deres er rettet mot hverandre) og lavere. Stor furer hver halvkule er delt inn i lapper. Sentral eller Rolands sulcus, plassert i den øvre delen av sideflaten av halvkulen og skiller frontallappen (lobus frontalis) fra parietallappen (lobus parietalis). Lateral, eller Sylvian sporet går også langs den laterale overflaten av halvkulen og skiller tinninglappen (lobus temporalis) fra frontal og parietal. Parieto-occipital sulcus skiller parietal- og occipitallappene (lobus occipitalis) langs den mediale overflaten av halvkulene. I dypet av den sylviske sprekken ligger insula lobe, lukket på alle sider av deler av bark som har "krøpet" inn på den. I tillegg identifiseres ofte en annen lapp, som ligger dypt i den mediale overflaten av halvkulen og dekker diencephalon på en bueformet måte. Dette er den limbiske lappen.
Mindre riller deler lappene inn i viklinger(gyrus). Noen av disse sporene er konstante (observert i alle individer), den andre er individuelle (ikke observert hos alle og ikke alltid), 2/3 av overflaten av cortex danner sideveggene til sporene og bare 1/3 er på overflaten av gyri.
Cerebral cortex er heterogen i opprinnelse og struktur. Det meste av den menneskelige cortex er okkupert av ny cortex - neocortex(neocortex), fylogenetisk den yngste kortikale formasjonen. Fylogenetisk tidligere kortikale strukturer - eldgammel bark(paleocortex) og gammel bark(archicortex) - okkuperer en liten del av overflaten av halvkulene. Dannelsen av ny bark dannes i de laterale delene av kappen. Den nye cortex utvikler seg intensivt og skyver den gamle cortex til bunnen av halvkulene, hvor den er bevart i form av en smal stripe av olfactory cortex og okkuperer 0,6% av overflaten av cortex på den ventrale overflaten av halvkulene, og den gamle cortex beveger seg til de mediale overflatene av halvkulene, okkuperer 2,2% av overflaten av cortex og er representert av hippocampus og dentate gyrus. I opprinnelse og cellestruktur skiller den nye cortex seg fra den gamle og gamle cortex. Det er imidlertid ingen skarpe cytoarkitektoniske grenser mellom dem. Overgangen fra en kortikal formasjon til en annen i cellestrukturen skjer gradvis. Overgangsbarken kalles interstitiell bark; den opptar 1,3 % av det totale barkarealet. Dermed er det meste av overflaten av cortex (95,6%) okkupert av den nye cortex.
Gammel og gammel bark.
For den eldgamle barken preget av fraværet av en lagdelt struktur. Det domineres av store nevroner gruppert i cellulære øyer. gammel bark har tre cellelag. Nøkkelstrukturen til den gamle cortex er hippocampus. Hippocampus (hippocampus), eller Ammons horn, Hippocampus (hippocampus), eller Ammons horn, ligger mediobasalt i dypet av tinninglappene. Den har en særegen buet form (hippocampus er oversatt som en sjøhest) og langs nesten hele lengden danner den en invaginasjon inn i hulrommet til det nedre hornet i sideventrikkelen. Hippocampus er faktisk en fold (gyrus) av den gamle cortex. Dentate gyrus er smeltet sammen med den og vikler seg over den. Som en del av den gamle cortex har hippocampus en lagdelt struktur. Ved siden av dentate gyrus er et lag med terminal forgrening av de apikale dendrittene til pyramidale celler i hippocampus. Her danner de et molekylært lag. Ulike afferente fibre ender på endegrenene til de apikale dendrittene og deres baser. De apikale dendrittene danner selv den neste - det radielle laget. Videre, mot det nedre hornet av den laterale ventrikkelen er det et lag med pyramidale cellelegemer og deres basale dendritter, så er det et lag med polymorfe celler. Laget av hvit substans i hippocampus (alveus) grenser til veggen av den laterale ventrikkelen Det består av begge aksoner av pyramidale nevroner i hippocampus (efferente fibre i hippocampus, som strekker seg som en del av fimbria inn i fornix), og av. afferente fibre som kommer langs fornix fra septum. Hippocampus har omfattende forbindelser med mange andre hjernestrukturer. Det er den sentrale strukturen i det limbiske systemet i hjernen.
Alle områder av neocortex er bygget etter samme prinsipp.
Den opprinnelige typen er sekslags bark. Lagene er representert som følger:
♦ Lag I er det mest overfladiske, ca. 0,2 mm tykt, kalt molekylært (lamina molecularis). Den består av fibre av apikale dendritter og aksoner som stiger opp fra cellene i de nedre lagene, som er i kontakt med hverandre. Det er et lite antall nevroner i det molekylære laget. Dette er små horisontale celler og granulatceller. Alle prosesser i cellene i det molekylære laget er lokalisert i samme lag.
♦ II-lag - eksternt granulært (lamina granulans externa). ♦ II-lag - eksternt granulært (lamina granulans externa). Tykkelsen på det ytre granulære laget er 0,10 mm. Den består av. små pyramidale og stjerneformede nevroner. Aksonene til disse nevronene ender på nevronene i lag III, V og VI.
♦ Lag III - pyramidalt (lamina pyramidalis), ♦ Lag III - pyramidalt (lamina pyramidalis), ca. 1 mm tykt, består av små og mellomstore pyramidale celler. En typisk pyramideformet nevron har formen av en trekant, hvis toppunkt er rettet oppover. En apikal dendritt strekker seg fra toppen og forgrener seg i de overliggende lagene. Aksonet til pyramidecellen strekker seg fra bunnen av cellen og går nedover. Dendrittene til lag III-celler ledes til det andre laget. Aksonene til celler i lag III ender på cellene i de underliggende lagene eller danner assosiative fibre.
♦ IV-lag - internt granulært (lamina granulans internus). ♦ IV-lag - internt granulært (lamina granulans internus). Den består av stjerneceller med korte prosesser og små pyramider. Dendrittene til lag IV-celler strekker seg inn i det molekylære laget av cortex, og deres kollateraler forgrener seg i sitt eget lag. Aksonene til lag IV-celler kan stige opp til de overliggende lagene eller gå inn i den hvite substansen som assosiasjonsfibre. Tykkelsen på IV-laget er fra 0,12 til 0,3 mm.
♦ V-lag - ganglion (lamina ganglionaris) - lag med store pyramider. De største cellene i cortex er lokalisert i dette laget ( gigantiske pyramider Betz fremre sentral gyrus) (se fig. 49B). Deres apikale dendritter når det molekylære laget, og de basale dendrittene er fordelt i laget deres. Aksonene til lag V-celler forlater cortex og er assosiasjons-, kommissurale eller projeksjonsfibre. Tykkelsen på V-laget når 0,5 mm. 93
♦ VI lag av cortex - polymorf (lamina multiformis). Den inneholder celler i ulike former og størrelser, og har en tykkelse fra 0,1 til 0,9 mm. Noen av dendrittene til cellene i dette laget når det molekylære laget, mens andre forblir innenfor lag IV og V. Aksonene til lag VI-celler kan stige opp til de øvre lagene eller forlate cortex som korte eller lange assosiasjonsfibre. Celler i samme lag av cortex utfører en lignende funksjon i informasjonsbehandling. Lag I og IV er stedet for forgrening av assosiative og kommissurelle fibre, dvs. motta informasjon fra andre kortikale strukturer. Lag III og IV er input, afferente for projeksjonsfelt, siden det er i disse lagene thalamusfibrene slutter. V-laget av celler utfører en efferent funksjon dets aksoner bærer informasjon til underliggende hjernestrukturer. Lag VI er også et utgangslag, men dets aksoner forlater ikke cortex, men er assosiative. Det grunnleggende prinsippet for funksjonell organisering av cortex er kombinasjonen av nevroner i kolonner. Søylen er plassert vinkelrett på overflaten av cortex og dekker alle lagene fra overflaten til den hvite substansen. Forbindelser mellom celler i samme kolonne gjøres vertikalt langs kolonnens akse. De laterale prosessene til cellene er korte. Forbindelsen mellom søylene til tilstøtende soner utføres gjennom fibre som går dypt og deretter går inn i en annen sone, dvs. korte assosiasjonsfibre. Den funksjonelle organiseringen av cortex i form av søyler finnes i de somatosensoriske, visuelle, motoriske og assosiative cortexene.
Individuelle soner i cortex har en fundamentalt identisk cellulær struktur, men det er også forskjeller, spesielt i strukturen til lag III, IV og V, som kan deles inn i flere underlag. I tillegg er betydelige cytoarkitektoniske trekk tettheten og størrelsen på celler, tilstedeværelsen av spesifikke typer nevroner, plasseringen og retningen til myelinfibre.
Cytoarkitektoniske egenskaper gjorde det mulig å dele hele overflaten av cortex inn i 11 cytoarkitektoniske regioner, inkludert 52 felt (ifølge Brodmann). Hvert cytoarkitektonisk felt er utpekt på hjernekart med et nummer, som ble tildelt det i beskrivelsesrekkefølgen. Det bør bemerkes at det ikke er noen skarpe grenser mellom de cytoarkitektoniske feltene, og cellelagene endrer strukturen jevnt når de beveger seg fra ett felt til et annet. Hvert kortikale felt utfører en bestemt funksjon. Noen av de kortikale feltene er sensoriske. De projeksjonsafferente fibrene ender i de primære sansefeltene. Fra de primære sansefeltene overføres informasjon langs korte assosiasjonsfibre til de sekundære projeksjonsfeltene som ligger ved siden av dem. Således er felt 1 og 3, som okkuperer de mediale og laterale overflatene av den bakre sentrale gyrusen, de primære projeksjonsfeltene for kutan følsomhet av den motsatte halvdelen av kroppsoverflaten. Hudområder som ligger ved siden av hverandre projiseres også ved siden av hverandre på den kortikale overflaten. Denne organiseringen av projeksjoner kalles aktuell. De nedre lemmene er representert i den mediale delen, og projeksjoner av reseptorfeltene på hudens overflate av hodet er plassert lavest på den laterale delen av gyrusen. I dette tilfellet projiseres områder av kroppsoverflaten som er rikt forsynt med reseptorer (fingre, lepper, tunge) på et større område av cortex enn områder med færre reseptorer (lår, rygg, skulder). Felt 2, som ligger i den inferolaterale delen av samme gyrus, er det sekundære projeksjonsfeltet for kutan følsomhet. Felt 17-19, lokalisert i occipitallappen, er det visuelle sentrum av cortex. felt 17, som opptar selve occipitalpolen, er primært. Det 18. og 19. felt ved siden av det utfører funksjonen som sekundære assosiative felt og mottar input fra det 17. feltet. De auditive projeksjonsfeltene er lokalisert i tinninglappene. Ved siden av dem, på grensen til tinning-, occipital- og parietallappene, ligger den 37., 39. og 40., kun karakteristisk for den menneskelige hjernebarken. For de fleste er det i disse feltene i venstre hjernehalvdel et talesenter, som er ansvarlig for oppfatningen av muntlig og skriving. Felt 43, okkuperer nederste del bakre sentrale gyrus, mottar smaksafferenter. Strukturene for luktfølsomhet u sender sine signaler til hjernebarken uten å bytte i andre deler av sentralnervesystemet. Lukteløkene er plassert under den nedre frontallappen. Fra dem begynner luktekanalen, som er det første paret kraniale nerver (s. olfact) o rius). Kortikale projeksjoner av luktfølsomhet er strukturer i den gamle cortex.
De motoriske områdene av cortex er lokalisert i den presentrale gyrusen til frontallappen (foran projeksjonssonene med kutan følsomhet). Denne delen av cortex er okkupert av felt 4 og 5. Fra V-laget til disse feltene utgår en pyramideformet bane som ender ved motorneuroner ryggmarg. Plasseringen og forholdet til innervasjonssoner ligner på projeksjonsrepresentasjonen til hudanalysatoren, dvs. har en somatotopisk organisasjon. I de mediale delene av gyrusen er det søyler som regulerer aktiviteten til benmusklene, i den nedre delen, ved sidesporet - musklene i ansiktet og hodet motsatt side kropper.
Afferente og efferente projeksjonssoner skorpene opptar et relativt lite område. Det meste av overflaten av cortex er okkupert av tertiære eller interanalyzersoner, kalt assosiative soner.
Foreningssoner cortex opptar et betydelig rom mellom frontal, occipital og temporal cortex (60-70% av neocortex). De mottar multimodale input fra sensoriske områder.
Foreningssoner gi integrering sanseinntrykk og lek betydelig rolle i prosessene med høyere nervøs og mental aktivitet.
Cerebral cortex - lag grå materie på overflaten av hjernehalvdelene, 2-5 mm tykke, og danner mange spor og viklinger som betydelig øker området. Cortex er dannet av kroppene til nevroner og gliaceller arrangert i lag («skjerm»-type organisasjon). Under ligger hvit materie
representert av nervetråder.
Cortex er den yngste fylogenetisk og den mest komplekse i morfofunksjonelle organisering av hjernen. Dette er stedet for høyere analyse og syntese av all informasjon som kommer inn i hjernen. Det er her integrasjonen av alle komplekse former for atferd skjer. Hjernebarken er ansvarlig for bevissthet, tenkning, hukommelse, "heuristisk aktivitet" (evnen til å generalisere og gjøre funn). Cortex inneholder mer enn 10 milliarder nevroner og 100 milliarder gliaceller. Kortikale nevroner når det gjelder antall prosesser, er de bare multipolare, men når det gjelder deres plass i refleksbuene og funksjonene de utfører, er de alle interkalære og assosiative. Basert på funksjon og struktur, skilles mer enn 60 typer nevroner i cortex. Basert på deres form er det to hovedgrupper: pyramideformet og ikke-pyramideformet. nevroner er hovedtypen av nevroner i cortex. Størrelsene på deres perikaryoner varierer fra 10 til 140 mikron i tverrsnitt de har en pyramideformet form. En lang (apikal) dendritt strekker seg oppover fra deres øvre hjørne, som er delt i en T-form i det molekylære laget. Laterale dendritter strekker seg fra sideflatene til nevronkroppen. Dendrittene og cellekroppen til nevronet har mange synapser med andre nevroner. Et akson strekker seg fra bunnen av cellen, som enten går til andre deler av cortex, eller til andre deler av hjernen og ryggmargen. Blant nevronene i hjernebarken er det assosiativ- forbinder områder av cortex innenfor en halvkule, kommissær– deres aksoner går til den andre halvkule, og projeksjon– deres aksoner går til de underliggende delene av hjernen.
Mellom ikke-pyramideformet De vanligste typene nevroner er stjerneceller og spindelceller. Stjerneformet nevroner er små celler med korte, sterkt forgrenede dendritter og aksoner som danner intrakortikale forbindelser. Noen av dem har en hemmende effekt, mens andre har en eksitatorisk effekt på pyramidale nevroner. Fusiform nevroner har et langt akson som kan gå i vertikal eller horisontal retning. Cortex bygges iht skjerm type, det vil si at nevroner som ligner i struktur og funksjon er ordnet i lag (fig. 9-7). Det er seks slike lag i cortex:
1.Molekylær lag - den mest ytre. Den inneholder et plexus av nervefibre som ligger parallelt med overflaten av cortex. Hoveddelen av disse fibrene er grener av de apikale dendrittene til pyramidale nevroner i de underliggende lagene i cortex. Afferente fibre fra den visuelle thalamus kommer også hit, og regulerer eksitabiliteten til kortikale nevroner. Nevroner i det molekylære laget er stort sett små og fusiforme.
2. Ytre kornet lag. Består av et stort antall stjerneceller. Dendrittene deres strekker seg inn i det molekylære laget og danner synapser med thalamo-kortikale afferente nervefibre. Laterale dendritter kommuniserer med nærliggende nevroner i samme lag. Aksoner danner assosiasjonsfibre som beveger seg gjennom den hvite substansen til nærliggende områder av cortex og danner synapser der.
3. Ytre lag av pyramidale nevroner(pyramideformet lag). Den er dannet av mellomstore pyramidale nevroner. Akkurat som nevronene i det andre laget, går dendrittene deres til det molekylære laget, og aksonene deres går til den hvite substansen.
4. Indre granulært lag. Den inneholder mange stjernenevroner. Dette er assosiative, afferente nevroner. De danner mange forbindelser med andre kortikale nevroner. Her er et annet lag med horisontale fibre.
5. Indre lag av pyramidale nevroner(ganglionisk lag). Den er dannet av store pyramidale nevroner. De sistnevnte er spesielt store i den motoriske cortex (presentral gyrus), hvor de måler opp til 140 mikron og kalles Betz-celler. Deres apikale dendritter stiger inn i det molekylære laget, laterale dendritter danner forbindelser med nærliggende Betz-celler, og aksoner er projeksjonsefferente fibre som går til medulla oblongata og ryggmargen.
6. Lag av fusiforme nevroner(lag av polymorfe celler) består hovedsakelig av spindelneuroner. Dendrittene deres går til det molekylære laget, og aksonene deres går til de visuelle åsene.
Den seks-lags typen struktur av cortex er karakteristisk for hele cortex, men i forskjellige deler av den varierer alvorligheten til lagene, samt formen og plasseringen av nevroner og nervefibre betydelig. Basert på disse egenskapene identifiserte K. Brodman 50 cytoarkitektoner i cortex felt. Disse feltene er også forskjellige i funksjon og metabolisme.
Den spesifikke organiseringen av nevroner kalles cytoarkitektonikk. Således, i de sensoriske sonene i cortex, er pyramide- og ganglionlagene dårlig uttrykt, og de granulære lagene er godt uttrykt. Denne typen bark kalles granulært. I de motoriske sonene, tvert imot, er de granulære lagene dårlig utviklet, og de pyramidale er godt utviklet. Dette agranulær type bark.
I tillegg er det et konsept myeloarkitektur. Dette er en spesifikk organisering av nervefibre. Således er det i hjernebarken vertikale og tre horisontale bunter av myeliniserte nervefibre. Blant nervefibrene i hjernebarken er det assosiativ- forbindende områder av cortex på en halvkule, kommissær– forbinder cortex forskjellige halvkuler Og projeksjon fibre - forbinder cortex med kjernene i hjernestammen.
Ris. 9-7. Cortex av de store halvkulene i den menneskelige hjernen.
A, B. Celleplassering (cytoarkitektur).
B. Plassering av myelinfibre (myeloarkitektur).
 Forholdet mellom menneske og natur Forholdet mellom menneske og natur
Forholdet mellom menneske og natur Forholdet mellom menneske og natur Sammensatte ord med en forbindelsesvokal
Sammensatte ord med en forbindelsesvokal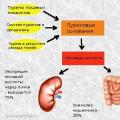 Urinsyre i blodet: normer og avvik, hvorfor det øker, kosthold for å redusere det Sluttproduktet av nitrogenmetabolismen er
Urinsyre i blodet: normer og avvik, hvorfor det øker, kosthold for å redusere det Sluttproduktet av nitrogenmetabolismen er