Strukturen til bindevevet til brusk. Strukturen og funksjonene til menneskelig brusk
Det er ingen hemmelighet at idrettsutøvere, selv i god fysisk form og i forhold tidlig alder faller ofte ut av trening på grunn av skade. En stor del av problemene deres er leddbånd. Deres svakeste del er bruskvev. Funksjonene til skadede ledd, viser det seg, kan gjenopprettes hvis du tar hensyn til problemet i tide og skaper passende forhold for behandling og regenerering av cellene deres.
Vev i menneskekroppen
Menneskekroppen er kompleks og fleksibelt system i stand til selvregulering. Den består av celler med ulike strukturer og funksjoner. De utfører hovedmetabolismen. Sammen med ikke-cellulære strukturer er de kombinert til vev: epitel, muskel, nervøs, bindemiddel.
Epitelceller danner grunnlaget for huden. De forer de indre hulrommene (buk, thorax, øvre luftveier, tarmkanal). Muskelvev lar en person bevege seg. Det sikrer også bevegelse av interne medier i alle organer og systemer. Musklene er delt inn i typer: glatt (vegger av bukorganer og kar), hjerte, skjelett (stripete). nervevev overfører impulser fra hjernen. Noen celler er i stand til å vokse og formere seg, noen av dem er i stand til å regenerere.
Bindevev er det indre miljøet i kroppen. Det er forskjellig i struktur, struktur og egenskaper. Den består av sterke skjelettbein, subkutant fettvev, flytende medier: blod og lymfe. Det inkluderer også brusk. Dens funksjoner er forming, demping, støtte og støtte. De spiller alle sammen viktig rolle og er nødvendige i komplekst system organisme.
struktur og funksjoner
Henne trekk- løshet i arrangementet av celler. Når du ser på dem individuelt, kan du se hvor tydelig de er atskilt fra hverandre. Båndet mellom dem er det intercellulære stoffet - matrisen. Dessuten, forskjellige typer brusk, dannes det, i tillegg til det viktigste amorfe stoffet, av forskjellige fibre (elastisk og kollagen). Selv om de har til felles protein opprinnelse, men varierer i egenskaper og, avhengig av dette, utfører forskjellige funksjoner.
Alle bein i kroppen består av brusk. Men etter hvert som de vokste, ble deres intercellulære substans fylt med saltkrystaller (hovedsakelig kalsium). Som et resultat fikk beinene styrke og ble en del av skjelettet. Brusk utfører også støttefunksjoner. I ryggraden, mellom segmentene, oppfatter de konstante belastninger (statiske og dynamiske). Auriklene, nesen, luftrøret, bronkiene - i disse områdene spiller vevet en mer formativ rolle.
Bruskens vekst og ernæring utføres gjennom perichondrium. Det er en obligatorisk del i vevet, bortsett fra leddene. De inneholder leddvæske mellom gnideflatene. Den vasker, smører og nærer dem, fjerner metabolske produkter.
Struktur
Det er få celler i brusken som er i stand til å dele seg, og det er mye plass rundt dem, fylt med et proteinstoff med ulike egenskaper. På grunn av denne funksjonen finner regenereringsprosesser ofte sted i matrisen i større grad.
Det finnes to typer vevsceller: kondrocytter (modne) og kondroblaster (unge). De er forskjellige i størrelse, plassering og metode for plassering. Kondrocytter har rund form og de er større. Arrangert i par eller i grupper på opptil 10 celler. Kondroblaster er vanligvis mindre og er lokalisert i vevet langs periferien eller enkeltvis.
I cytoplasmaet til celler under membranen akkumuleres vann, det er inneslutninger av glykogen. Oksygen og næringsstoffer kommer diffust inn i cellene. Det er en syntese av kollagen og elastin. De er nødvendige for dannelsen av intercellulær substans. Det avhenger av dens spesifisitet hvilken type bruskvev det vil være. Strukturelle egenskaper og skiller seg fra mellomvirvelskiver, inkludert innholdet av kollagen. I brusken i nesen består det intercellulære stoffet av 30 % elastin.
Slags
Hvordan det klassifiseres Dens funksjoner avhenger av overvekten av spesifikke fibre i matrisen. Hvis det er mer elastin i det intercellulære stoffet, vil bruskvevet være mer plastisk. Den er nesten like sterk, men fiberbuntene i den er tynnere. De tåler godt belastninger, ikke bare i kompresjon, men også i spenning, er i stand til deformasjon uten kritiske konsekvenser. Slik brusk kalles elastisk. Deres vev danner strupehodet, auriklene, nesen.
Dersom matrisen rundt cellene har et høyt innhold av kollagen med kompleks struktur bygge polypeptidkjeder, slik brusk kalles hyalin. Det dekker vanligvis indre overflater ledd. Det største antallet kollagen er konsentrert i den overfladiske sonen. Det spiller rollen som et rammeverk. Buntene av fibre i den minner strukturelt om tredimensjonale sammenvevde nettverk med spiralform.
Det er en annen gruppe: fibrøs eller fibrøs brusk. De, som hyaline, inneholder en stor mengde kollagen i det intercellulære stoffet, men det har en spesiell struktur. Buntene av fibrene deres har ikke en kompleks veving og er plassert langs aksen til de største belastningene. De er tykkere, har en spesiell trykkstyrke og blir dårlig restaurert under deformasjon. Intervertebrale skiver, krysset mellom sener og bein, er dannet fra slikt vev.

Funksjoner
På grunn av sine spesielle biomekaniske egenskaper er bruskvev ideell for å binde komponentene i muskel- og skjelettsystemet. Den er i stand til å ta påvirkningen av trykk- og strekkkrefter under bevegelser, omfordele dem jevnt til belastningen, absorbere eller spre seg til en viss grad.
Bruskene danner slitasjebestandige overflater. I kombinasjon med leddvæske, slike ledd med tillatte belastninger i stand til å utføre sine funksjoner normalt i lang tid.
Sener er ikke brusk. Deres funksjoner består også i å knytte seg til et felles apparat. De består også av bunter av kollagenfibre, men deres struktur og opprinnelse er forskjellig. åndedrettsorganer, aurikler, i tillegg til å utføre formende og støttende funksjoner, er stedet for feste av bløtvev. Men i motsetning til sener, har ikke musklene ved siden av dem en slik belastning.
Spesielle egenskaper
Det er svært få kar i elastisk brusk. Og dette er forståelig, fordi en sterk dynamisk belastning kan skade dem. Hvordan mates bruskbindevev? Disse funksjonene overtas av det intercellulære stoffet. Det er ingen kar i hyalin brusk i det hele tatt. Gniflatene deres er ganske harde og tette. De drives av leddvæsken i leddet.
I matrisen beveger vann seg fritt. Den inneholder alle nødvendige stoffer for metabolske prosesser. Proteoglykankomponentene i brusk er ideelle for å binde vann. Som et inkompressibelt stoff gir det stivhet og ekstra demping. Når det er lastet, tar vann på seg støtet, sprer seg gjennom det intercellulære rommet og lindrer jevnt stress, og forhindrer irreversible kritiske deformasjoner.

Utvikling
I kroppen til en voksen faller opptil 2% av massen på bruskvev. Hvor er den plassert og hvilke funksjoner utfører den? Brusk- og benvev i embryonalperioden er ikke differensiert. Embryoer har ingen bein. De utvikler seg fra brusk og dannes ved fødselstidspunktet. Men en del av det forbener seg aldri. Ører, nese, strupehode, bronkier dannes av det. Det er også tilstede i leddene i armer og ben, leddene i mellomvirvelskivene, meniskene i knærne.
Bruskutvikling skjer i flere stadier. Først blir de mesenkymale cellene mettet med vann, runde, mister prosessene og begynner å produsere stoffer til matrisen. Etter det differensierer de til kondrocytter og kondroblaster. De førstnevnte er tett omgitt av intercellulær substans. I denne tilstanden kan de dele et begrenset antall ganger. Etter slike prosesser dannes en isogen gruppe. Celler som blir igjen på overflaten av vevet blir kondroblaster. I prosessen med å produsere matrisestoffer oppstår endelig differensiering, en struktur dannes med en distinkt inndeling i en tynn kant og vevsbase.

Aldersendringer
Bruskfunksjonene endres ikke i løpet av livet. Men over tid kan tegn på aldring bli lagt merke til: muskler og sener i leddene svekkes, fleksibilitet går tapt, smerte forstyrres av en endring i været eller med en uvanlig belastning. En slik prosess betraktes som en fysiologisk norm. I en alder av 30-40 år kan symptomene på endringer allerede begynne å forårsake ulemper i større eller mindre grad. Aldringen av leddbruskvevet oppstår på grunn av tap av elastisitet. Elastisiteten til fibrene går tapt. Stoffet tørker opp og løsner.
Sprekker vises på en jevn overflate, den blir grov. Glatthet og enkel glidning er ikke lenger mulig. Skadede kanter vokser, avleiringer dannes i dem, og osteofytter dannes i vevet. Elastiske brusk eldes med akkumulering av kalsium i det intercellulære stoffet, men dette påvirker nesten ikke funksjonene deres (nese, aurikler).

Dysfunksjon av brusk og beinvev
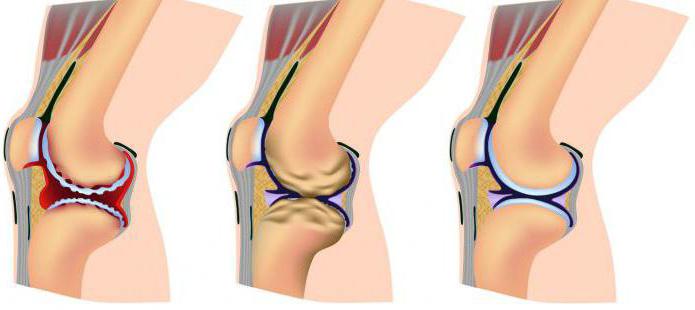
Når og hvordan kan dette skje? I stor grad avhenger det av hvilken funksjon bruskvevet utfører. I de intervertebrale skivene, hvis hovedfunksjon er stabiliserende og støttende, oppstår oftest en funksjonsfeil under utviklingen av dystrofiske eller degenerative prosesser. Situasjonen kan føre til forskyvninger, som igjen vil føre til kompresjon av det omkringliggende vevet. Hevelse, klyping av nerver, klemming av blodårer er uunngåelig.
For å gjenopprette stabiliteten prøver kroppen å bekjempe problemet. Virvelen på stedet for deformasjon "tilpasser seg" til situasjonen, vokser i form av særegne beinutvekster (whiskers). Dette gagner heller ikke det omkringliggende vevet: igjen hevelse, brudd, kompresjon. Dette problemet er komplekst. Brudd på funksjonen til det osteokondrale apparatet kalles ofte osteokondrose.
Langvarig bevegelsesbegrensning (gips mot skader) påvirker også brusken negativt. Hvis elastiske fibre under overdreven belastning degenererer til grove fibrøse bunter, slutter brusk å spise normalt med lav aktivitet. Leddvæsken blander seg ikke godt, kondrocytter mottar mindre næringsstoffer, som et resultat blir de ikke produsert. nødvendig beløp kollagen og elastin for matrisen.
Konklusjonen tyder på seg selv: for normal funksjon av leddene må brusken motta en tilstrekkelig belastning i spenning og kompresjon. For å sikre dette, må du trening, lede en sunn og aktivt bilde liv.
Bruskvev er en spesiell type bindevev og utfører en støttefunksjon i den dannede organismen. I maxillofacial regionen er brusk en del av auricle, auditive tube, nese, leddskive i kjeveleddet, og gir også en forbindelse mellom de små beinene i skallen.
Avhengig av sammensetning, metabolsk aktivitet og evne til å regenerere, er det tre typer bruskvev - hyalin, elastisk og fibrøs.
hyalin brusk dannes først på det embryonale utviklingsstadiet, og under visse forhold dannes de to andre typer brusk fra den. Dette bruskvevet finnes i kystbruskene, bruskstrukturen i nesen, og danner bruskene som dekker overflatene til leddene. Den har en høyere metabolsk aktivitet sammenlignet med de elastiske og fibrøse typene og inneholder en stor mengde karbohydrater og lipider. Dette tillater aktiv proteinsyntese og differensiering av kondrogene celler for å fornye og regenerere hyalinbrusk. Med alderen oppstår hypertrofi og apoptose av celler i hyalinbrusk, etterfulgt av forkalkning av den ekstracellulære matrisen.
Elastisk brusk har en lignende struktur som hyalin brusk. Fra slikt bruskvev dannes for eksempel auriklene, hørselsrøret og noen brusk i strupehodet. Denne typen brusk er preget av tilstedeværelsen av et nettverk av elastiske fibre i bruskmatrisen, en liten mengde lipider, karbohydrater og kondroitinsulfater. På grunn av lav metabolsk aktivitet forkalker ikke elastisk brusk og regenereres praktisk talt ikke.
fibrobrusk i sin struktur inntar den en mellomposisjon mellom senen og hyalinbrusken. Et karakteristisk trekk ved fibrocartilage er tilstedeværelsen i den intercellulære matrisen av et stort antall kollagenfibre, hovedsakelig type I, som er plassert parallelt med hverandre, og celler i form av en kjede mellom dem. Fibrøs brusk, på grunn av sin spesielle struktur, kan oppleve betydelig mekanisk belastning både i kompresjon og i strekk.
Bruskkomponent i kjeveleddet presentert i form av en skive av fibrøs brusk, som er plassert på overflaten av den artikulære prosessen mandible og skiller den fra leddfossaen til tinningbenet. Siden fibrobrusk ikke har perichondrium, får bruskcellene næring gjennom leddvæsken. Sammensetningen av leddvæsken avhenger av ekstravasasjonen av metabolitter fra blodårer synovial membran inn i leddhulen. Leddvæsken inneholder lavmolekylære komponenter - Na + , K + -ioner, urinsyre, urea, glukose, som er nært i kvantitativt forhold til blodplasma. Imidlertid er innholdet av proteiner i leddvæsken 4 ganger høyere enn i blodplasma. I tillegg til glykoproteiner, immunoglobuliner, er leddvæske rik på glykosaminoglykaner, blant hvilke hyaluronsyre, tilstede i form av natriumsalt, inntar førsteplassen.
2.1. STRUKTUR OG EGENSKAPER TIL BRISKVEV
Bruskvev, som alt annet vev, inneholder celler (kondroblaster, kondrocytter) som er innebygd i en stor intercellulær matrise. I prosessen med morfogenese differensierer kondrogene celler til kondroblaster. Kondroblaster begynner å syntetisere og skille ut proteoglykaner i bruskmatrisen, som stimulerer differensieringen av kondrocytter.
Den intercellulære matrisen av bruskvev gir dens komplekse mikroarkitektonikk og består av kollagener, proteoglykaner og ikke-kollagenproteiner - hovedsakelig glykoproteiner. Kollagenfibre er flettet sammen i et tredimensjonalt nettverk som forbinder resten av matrisekomponentene.
Cytoplasmaet til kondroblaster inneholder en stor mengde glykogen og lipider. Nedbrytningen av disse makromolekylene i oksidative fosforyleringsreaksjoner er ledsaget av dannelsen ATP-molekyler nødvendig for proteinsyntese. Proteoglykaner og glykoproteiner syntetisert i det granulære endoplasmatiske retikulumet og Golgi-komplekset pakkes inn i vesikler og frigjøres i den ekstracellulære matrisen.
Elastisiteten til bruskmatrisen bestemmes av mengden vann. Proteoglykaner er preget av en høy grad av vannbinding, som bestemmer størrelsen. Bruskmatrisen inneholder opptil 75 %
vann, som er assosiert med proteoglykaner. Høy grad hydrering bestemmer den store størrelsen på den ekstracellulære matrisen og tillater ernæring av celler. Tørket agrecan etter binding av vann kan øke i volum med 50 ganger, men på grunn av begrensningene forårsaket av kollagennettverket overstiger ikke hevelsen av brusken 20 % av maksimalt mulig verdi.
Når brusk komprimeres, fortrenges vann, sammen med ioner, fra områdene rundt de sulfaterte og karboksylgruppene i proteoglykanen, gruppene nærmer seg hverandre, og frastøtende krefter mellom deres negative ladninger forhindrer ytterligere vevskompresjon. Etter at lasten er fjernet, oppstår den elektrostatiske tiltrekningen av kationer (Na +, K +, Ca 2+), etterfulgt av tilstrømning av vann inn i den intercellulære matrisen (fig. 2.1).
Ris. 2.1.Vannbinding av proteoglykaner i bruskmatrisen. Forskyvning av vann under komprimering og restaurering av strukturen etter fjerning av lasten.
Kollagenproteiner i brusk
Styrken til bruskvev bestemmes av kollagenproteiner, som er representert av type II, VI, IX, XII, XIV kollagener og er nedsenket i makromolekylære aggregater av proteoglykaner. Type II kollagener står for omtrent 80-90% av alle kollagenproteiner i brusk. De resterende 15-20 % av kollagenproteinene er de såkalte mindre kollagenene av typene IX, XII, XIV, som kryssbinder type II kollagenfibriller og binder glykosaminoglykaner kovalent. Et trekk ved matrisen av hyalin og elastisk brusk er tilstedeværelsen av type VI kollagen.
Type IX kollagen, som finnes i hyalin brusk, sikrer ikke bare interaksjonen av type II kollagen med proteoglykaner, men regulerer også diameteren til type II kollagenfibriller. Kollagen type X har samme struktur som kollagen type IX. Denne typen kollagen syntetiseres kun av hypertrofierte vekstplatekondrocytter og akkumuleres rundt cellene. Gitt unik eiendom type X kollagen antyder deltakelsen av dette kollagenet i prosessene med beindannelse.
Proteoglykaner. Generelt når innholdet av proteoglykaner i bruskmatrisen 3%-10%. Hovedproteoglykanet i brusk er agrecan, som er aggregert med hyaluronsyre. I formen ligner agrecan-molekylet en flaskebørste og er representert av én polypeptidkjede (kjerneprotein) med opptil 100 kondroitinsulfatkjeder og ca. 30 keratansulfatkjeder festet til den (fig. 2.2).

Ris. 2.2.Proteoglykanaggregat av bruskmatrisen. Proteoglykanaggregatet består av ett hyaluronsyremolekyl og omtrent 100 agrekanmolekyler.
Tabell 2.1
Ikke-kollagenøse bruskproteiner
Navn | Egenskaper og funksjoner |
Kondrokalsin | Kalsiumbindende protein, som er et C-propeptid av type II kollagen. Proteinet inneholder 3 rester av 7-karboksyglutaminsyre. Syntetisert av hypertrofiske kondroblaster og gir mineralisering av bruskmatrisen |
Gla protein | I motsetning til beinvev, inneholder brusk et Gla-protein med høy molekylvekt, som inneholder 84 aminosyrerester (i bein - 79 aminosyrerester) og 5 rester av 7-karboksyglutaminsyre. Det er en hemmer av bruskmineralisering. Hvis syntesen forstyrres under påvirkning av warfarin, dannes foci av mineralisering, etterfulgt av forkalkning av bruskmatrisen. |
Chondroaderin | Glykoprotein med mol. veier 36 kDa, rik på leucin. Korte oligosakkaridkjeder, bestående av sialinsyrer og heksosaminer, er festet til serinrester. Kondroaderin binder type II kollagener og proteoglykaner til kondrocytter og kontrollerer den strukturelle organiseringen av den ekstracellulære bruskmatrisen |
Bruskprotein (CILP) | Glykoprotein med mol. som veier 92 kDa, og inneholder en oligosakkaridkjede koblet til proteinet med en N-glykosidbinding. Proteinet syntetiseres av kondrocytter, deltar i nedbrytningen av proteoglykanaggregater og er nødvendig for å opprettholde konstansen i bruskvevsstrukturen. |
Matrilin-1 | Adhesivt glykoprotein med en mol. veier 148 kDa, bestående av tre polypeptidkjeder koblet med disulfidbindinger. Det er flere isoformer av dette proteinet - matrilin -1, -2, -3, -4. I friskt modent bruskvev finnes ikke matrilin. Det syntetiseres i prosessen med bruskvevsmorfogenese og av hypertrofiske kondrocytter. Dens aktivitet er manifestert i revmatoid artritt. Med utviklingen av den patologiske prosessen binder den fibrillære fibre av type II kollagen med proteoglykanaggregater og bidrar dermed til å gjenopprette strukturen til bruskvev |
I strukturen til agrecan-kjerneproteinet er et N-terminalt domene isolert, som sikrer binding av agrecan til hyaluronsyre og lavmolekylære bindende proteiner, og et C-terminalt domene, som binder agrecan til andre molekyler i den ekstracellulære matrisen . Syntese av komponentene i proteoglykanaggregater utføres av kondrocytter, og den endelige dannelsesprosessen fullføres i den ekstracellulære matrisen.
Sammen med store proteoglykaner er små proteoglykaner tilstede i bruskmatrisen: decorin, biglykan og fibromodulin. De utgjør bare 1-2 % av den totale tørrstoffmassen av brusk, men deres rolle er veldig stor. Decorin, bindende i visse områder med type II kollagenfibre, er involvert i prosessene med fibrillogenese, og biglykan er involvert i dannelsen av bruskproteinmatrisen under embryogenese. Med veksten av embryoet avtar mengden biglykan i bruskvevet, og etter fødselen forsvinner denne proteoglykanen helt. Regulerer diameteren til type II kollagen fibromodulin.
I tillegg til kollagener og proteoglykaner inneholder den ekstracellulære matrisen av brusk uorganiske forbindelser og en liten mengde ikke-kollagenproteiner, som er karakteristiske ikke bare for brusk, men også for andre vev. De er nødvendige for bindingen av proteoglykaner til kollagenfibre, celler og individuelle komponenter i bruskmatrisen til et enkelt nettverk. Dette er klebende proteiner - fibronektin, laminin og integriner. De fleste av de spesifikke ikke-kollagenproteinene i bruskmatrisen er kun tilstede i perioden med morfogenese, forkalkning av bruskmatrisen, eller vises under patologiske forhold(Tabell 2.1). Oftest er dette kalsiumbindende proteiner som inneholder 7-karboksyglutaminsyrerester, samt glykoproteiner rike på leucin.
2.2. DANNING AV BRISKVEV
På tidlig stadie embryonal utvikling, bruskvev består av udifferensierte celler inneholdt i form av en amorf masse. I prosessen med morfogenese begynner cellene å differensiere, den amorfe massen øker og tar form av fremtidig brusk (fig. 2.3).
I den ekstracellulære matrisen til det utviklende bruskvevet endres sammensetningen av proteoglykaner, hyaluronsyre, fibronektin og kollagenproteiner kvantitativt og kvalitativt. Overfør fra
Ris. 2.3.Stadier av dannelse av bruskvev.
prechondrogene mesenkymale celler til kondroblaster er preget av sulfatering av glykosaminoglykaner, en økning i mengden hyaluronsyre og går foran begynnelsen av syntesen av en bruskspesifikk stor proteoglykan (agrecan). Ved primær

stadier av morfogenese syntetiseres høymolekylære bindende proteiner, som senere gjennomgår begrenset proteolyse med dannelse av lavmolekylære proteiner. Molekyler av agrecan binder seg til hyaluronsyre ved hjelp av lavmolekylære bindende proteiner og proteoglykanaggregater dannes. Deretter avtar mengden hyaluronsyre, noe som er assosiert med både en reduksjon i syntesen av hyaluronsyre og en økning i aktiviteten til hyaluronidase. Til tross for reduksjonen i mengden hyaluronsyre, øker lengden på dens individuelle molekyler, nødvendig for dannelsen av proteoglykanaggregater under kondrogenese. Syntesen av type II kollagen av kondroblaster skjer senere enn syntesen av proteoglykaner. Til å begynne med syntetiserer prekondrogene celler kollagener I og III type derfor finnes type I kollagen i cytoplasmaet til modne kondrocytter. Videre, i prosessen med kondrogenese, er det en endring i komponentene i den ekstracellulære matrisen som kontrollerer morfogenesen og differensieringen av kondrogene celler.
Brusk som en forløper til bein
Alle bokmerkene til beinskjelettet går gjennom tre stadier: mesenkymalt, brusk og bein.
Mekanismen for bruskforkalkning er en svært kompleks prosess og er ennå ikke fullt ut forstått. Ossifikasjonspunkter, langsgående septa i den nedre hypertrofiske sonen av brusk-rudimenter, samt laget av leddbrusk ved siden av beinet er gjenstand for fysiologisk forkalkning. Den sannsynlige årsaken til denne utviklingen av hendelser er tilstedeværelsen av alkalisk fosfatase på overflaten av hypertrofiske kondrocytter. I matrisen som er utsatt for forkalkning, dannes såkalte matriksvesikler som inneholder fosfatase. Det antas at disse vesiklene tilsynelatende er det primære området for bruskmineralisering. Rundt kondrocytter øker den lokale konsentrasjonen av fosfationer, noe som bidrar til vevsmineralisering. Hypertrofiske kondrocytter syntetiserer og frigjør i bruskmatrisen et protein - kondrokalsin, som har evnen til å binde kalsium. Områder utsatt for mineralisering er preget av høye konsentrasjoner fosfolipider. Deres tilstedeværelse stimulerer dannelsen av hydroksyapatittkrystaller på disse stedene. I sonen for bruskforkalkning skjer delvis nedbrytning av proteoglykaner. De av dem som ikke har blitt påvirket av nedbrytning bremser forkalkningen.
Brudd på induktive forhold, så vel som en endring (forsinkelse eller akselerasjon) i tidspunktet for utseendet og synostese av ossifikasjonssentre i sammensetningen av individuelle beinanlag, forårsaker dannelsen av strukturelle defekter i hodeskallen i det menneskelige embryoet.
Bruskregenerering
Brusktransplantasjon innen samme art (såkalte allogene transplantasjoner) er vanligvis ikke ledsaget av symptomer på en avvisningsreaksjon hos mottakeren. Denne effekten kan ikke oppnås med hensyn til andre vev, siden transplantatene til disse vevene blir angrepet og ødelagt av celler i immunsystemet. Den vanskelige kontakten av donorens kondrocytter med cellene i mottakerens immunsystem skyldes først og fremst tilstedeværelsen av en stor mengde intercellulær substans i brusken.
Hyalinbrusken har den høyeste regenerative kapasiteten, som er assosiert med den høye metabolske aktiviteten til kondrocytter, samt tilstedeværelsen av perichondrium, et tett fibrøst uformet bindevev som omgir brusken og inneholder et stort antall blodkar. Type I kollagen er tilstede i det ytre laget av perichondrium, mens det indre laget er dannet av kondrogene celler.
På grunn av disse funksjonene praktiseres bruskvevstransplantasjon i plastisk kirurgi, for eksempel for rekonstruksjon av en vansiret nesekontur. I dette tilfellet er allogen transplantasjon av kondrocytter alene, uten det omkringliggende vevet, ledsaget av graftavvisning.
Regulering av bruskmetabolismen
Dannelsen og veksten av bruskvev reguleres av hormoner, vekstfaktorer og cytokiner. Kondroblaster er målceller for tyroksin, testosteron og somatotropin, som stimulerer veksten av bruskvev. Glukokortikoider (kortisol) hemmer celleproliferasjon og differensiering. En viss rolle i reguleringen av funksjonstilstanden til bruskvev spilles av kjønnshormoner som hemmer frigjøringen av proteolytiske enzymerødelegger bruskmatrisen. I tillegg syntetiserer brusken selv proteinasehemmere som undertrykker aktiviteten til proteinaser.
En rekke vekstfaktorer - TGF-(3, fibroblastvekstfaktor, insulinlignende vekstfaktor-1 stimulerer vekst og utvikling
bruskvev. Ved å binde seg til kondrocyttmembranreseptorer aktiverer de syntesen av kollagener og proteoglykaner og bidrar derved til å opprettholde bruskmatrisens konstans.
Brudd på hormonell regulering er ledsaget av overdreven eller utilstrekkelig syntese av vekstfaktorer, noe som fører til en rekke defekter i dannelsen av celler og ekstracellulær matrise. Så, revmatoid artritt, slitasjegikt og andre sykdommer er assosiert med økt dannelse av skjelettceller, og brusk begynner å bli erstattet av bein. Under påvirkning av blodplatevekstfaktor begynner kondrocytter selv å syntetisere IL-1α og IL-1(3), hvis akkumulering hemmer syntesen av proteoglykaner og kollagen type II og IX. Dette bidrar til kondrocytthypertrofi og til syvende og sist forkalkning av den intercellulære matrisen til bruskvev Destruktive endringer er også assosiert aktivering av matrisemetalloproteinaser involvert i nedbrytningen av bruskmatrisen.
Aldersrelaterte endringer i brusk
Med aldring oppstår degenerative endringer i brusken, den kvalitative og kvantitative sammensetningen av glykosaminoglykaner endres. Dermed er kjedene av kondroitinsulfat i proteoglykanmolekylet syntetisert av unge kondrocytter nesten 2 ganger lengre enn kjedene produsert av mer modne celler. Jo lengre kondroitinsulfatmolekylene er i proteoglykanen, jo mer vann strukturerer proteoglykanen. I denne forbindelse binder proteoglykanen til gamle kondrocytter mindre vann, slik at bruskmatrisen til eldre blir mindre elastisk. Endringer i mikroarkitektonikken til den ekstracellulære matrisen i enkeltsaker er årsaken til slitasjegikt. Sammensetningen av proteoglykaner syntetisert av unge kondrocytter inneholder også en stor mengde kondroitin-6-sulfat, mens hos eldre mennesker, tvert imot, dominerer kondroitin-4-sulfater i bruskmatrisen. Tilstanden til bruskmatrisen bestemmes også av lengden på glykosaminoglykankjeder. Hos unge mennesker syntetiserer kondrocytter kortkjedet keratansulfat, og med alderen forlenges disse kjedene. En reduksjon i størrelsen på proteoglykanaggregater er også observert på grunn av forkortningen av ikke bare glykosaminoglykankjeder, men også lengden på kjerneproteinet i ett proteoglykanmolekyl. Med aldring øker innholdet av hyaluronsyre i brusk fra 0,05 til 6%.
En karakteristisk manifestasjon av degenerative endringer i bruskvev er dens ikke-fysiologiske forkalkning. Det forekommer vanligvis hos eldre og er preget av primær degenerasjon av leddbrusken etterfulgt av skade på leddkomponentene i leddet. Strukturen til kollagenproteiner endres og bindingssystemet mellom kollagenfibrene blir ødelagt. Disse endringene er assosiert med både kondrocytter og matrisekomponenter. Den resulterende hypertrofien av kondrocytter fører til en økning i bruskmasse i området med bruskhulrom. Type II kollagen forsvinner gradvis, som erstattes av type X kollagen, som deltar i prosessene med beindannelse.
Sykdommer forbundet med misdannelser av bruskvev
I tannlegepraksis utføres manipulasjoner oftest på over- og underkjeven. Det er en rekke trekk ved deres embryonale utvikling, som er assosiert med forskjellige utviklingsveier for disse strukturene. I det menneskelige embryoet tidlige stadier embryogenese, brusk finnes i sammensetningen av over- og underkjeven.
På den 6-7. uken av intrauterin utvikling begynner dannelsen av beinvev i mesenkymet til mandibulære prosesser. Overkjeven utvikler seg sammen med beinene i ansiktsskjelettet og gjennomgår forbening mye tidligere enn underkjeven. Ved 3 måneders alder inneholder den fremre overflaten av beinet ikke lenger sammensmeltningen av overkjeven med hodeskallens bein.
Ved den 10. uken av embryogenese dannes sekundær brusk i de fremtidige grenene av underkjeven. En av dem tilsvarer kondylprosessen, som midt i fosterutviklingen erstattes av beinvev i henhold til prinsippet om endokondral ossifikasjon. Sekundær brusk dannes også langs den fremre kanten av koronoidprosessen, som forsvinner like før fødselen. På stedet for fusjon av de to halvdelene av underkjeven er det en eller to øyer med bruskvev, som forbenes i de siste månedene av intrauterin utvikling. Ved den 12. uken av embryogenese vises kondylbrusken. Ved 16. uke kommer kondylen til underkjevegrenen i kontakt med tinningbeinet. Det skal bemerkes at føtal hypoksi, fraværet eller svak bevegelse av embryoet bidrar til forstyrrelse av dannelsen av leddrom eller fullstendig sammensmelting av epifysene til de motsatte benkantene. Dette fører til deformasjon av underkjeveprosessene og deres fusjon med tinningbenet (ankylose).
70. Bruskvev. Klassifikasjon, utvikling, struktur, histokjemiske egenskaper og funksjon. Bruskvekst, regenerering og aldersrelaterte endringer.
bruskaktig og beinvev utvikle seg fra sklerotomi mesenchyme, tilhører vev Internt miljø og, som alle andre vev i det indre miljøet, er sammensatt av celler og intercellulær substans. Den intercellulære substansen her er tett, så disse vevene utfører en støttemekanisk funksjon.
bruskvev(textuscartilagineus). De er klassifisert i hyalin, elastisk og fibrøs. Klassifiseringen er basert på funksjonene i organiseringen av det intercellulære stoffet. Brusk består av 80% vann, 10-15% organisk materiale og 5-7 % uorganiske stoffer.
Bruskutvikling, eller kondrogenese, består av 3 stadier: 1) dannelsen av kondrogene holmer; 2) dannelse av primært bruskvev, 3) differensiering av bruskvev.
Under 1. trinn mesenkymale celler kombineres til kondrogene øyer, hvis celler formerer seg, differensierer til kondroblaster. De dannede kondroblastene inneholder granulær EPS, Golgi-komplekset og mitokondrier. Kondroblastene differensierer deretter til kondrocytter.
Under 2. trinn i kondrocytter er granulær EPS, Golgi-komplekset og mitokondrier godt utviklet. Kondrocytter syntetiserer aktivt fibrillært protein (kollagen type II), hvorfra det dannes en intercellulær substans som farges oksyfilt.
På begynnelsen 3. trinn i kondrocytter utvikles granulær ER mer intensivt, hvorpå det produseres både fibrillære proteiner og kondroitinsulfater (kondroitin svovelsyre), som er farget med basiske fargestoffer. Derfor er den viktigste intercellulære substansen i bruskvevet rundt disse kondrocyttene farget basofilt.
Et perichondrium dannes rundt det bruskformede rudimentet fra mesenkymale celler, bestående av 2 lag: 1) ytre, tettere eller fibrøst, og 2) indre, løsere eller kondrogene, som inneholder prekondroblaster og kondroblaster.
apposisjonell vekst av brusk eller vekst ved superposisjon, kjennetegnes ved at kondroblaster frigjøres fra perichondrium, som er lagt over hovedstoffet i brusken, differensierer til kondrocytter og begynner å produsere den intercellulære substansen i bruskvevet.
Interstitiell vekst bruskvev utføres på grunn av kondrocytter som ligger inne i brusken, som for det første deler seg ved mitose og for det andre produserer et intercellulært stoff, på grunn av hvilket volumet av bruskvev øker.
Bruskceller(kondrocytus). Kondrocyttdifferonet er sammensatt av: stamcelle, halvstamcelle (prechondroblast), kondroblast, kondrocytt.
Kondroblaster (chondroblastus) er lokalisert i det indre laget av perichondrium, har organeller av generell betydning: granulær ER, Golgi-kompleks, mitokondrier. Funksjoner av kondroblaster:
1) skiller ut intercellulær substans (fibrillære proteiner);
2) i prosessen med differensiering blir de til kondrocytter;
3) har evnen til mitotisk deling.
Kondrocytter ligger i bruskhull. I lakunaen er det først 1 kondrocytt, deretter, i prosessen med dens mitotiske deling, dannes 2, 4, 6, etc. celler. Alle av dem er lokalisert i samme lakune og danner en isogen gruppe av kondrocytter.
Kondrocytter av den isogene gruppen er delt inn i 3 typer: I, II, III.
Type I kondrocytter har evne til mitotisk deling, inneholder Golgi-komplekset, mitokondrier, granulært ER og frie ribosomer, har en stor kjerne og en liten mengde cytoplasma (stort nukleært-cytoplasmatisk forhold). Disse kondrocyttene er lokalisert i ung brusk.
Type II kondrocytter lokalisert i moden brusk, reduseres deres nukleære-cytoplasmatiske forhold noe, ettersom volumet av cytoplasmaet øker; de mister evnen til mitose. I deres cytoplasma er granulær ER godt utviklet; de skiller ut proteiner og glykosaminoglykaner (kondroitinsulfater), så det viktigste intercellulære stoffet rundt dem farges basofilt.
Type III kondrocytter befinner seg i den gamle brusken, mister evnen til å syntetisere glykosaminoglykaner og produserer kun proteiner, så det intercellulære stoffet rundt dem flekker oksyfilt. Derfor er en ring farget oksyfilt (proteiner er isolert av type III kondrocytter) synlig rundt en slik isogen gruppe, en basofil farget ring er synlig utenfor denne ringen (glykosaminoglykaner skilles ut av type II kondrocytter) og selve den ytre ringen farges igjen oksyfilt (proteiner isoleres på et tidspunkt da brusk inneholdt bare unge type I kondrocytter). Dermed karakteriserer disse 3 forskjellig fargede ringene rundt isogene grupper prosessen med dannelse og funksjon av kondrocytter av 3 typer.
Intercellulær substans av bruskvev. Inneholder organiske stoffer (hovedsakelig type II kollagen), glykosaminoglykaner, proteoglykaner og ikke-kollagen type proteiner. Jo flere proteoglykaner, jo mer hydrofilt det intercellulære stoffet, jo mer elastisk og mer permeabelt er det. Gasser, vannmolekyler, saltioner og mikromolekyler trenger diffust gjennom hovedstoffet fra siden av perichondrium. Makromolekyler trenger imidlertid ikke gjennom. Makromolekyler har antigene egenskaper, men siden de ikke trenger inn i brusk, slår brusk transplantert fra en person til en annen godt rot (ingen immunavstøtningsreaksjon oppstår).
I grunnstoffet av brusk er det kollagenfibre, bestående av type II kollagen. Orienteringen av disse fibrene avhenger av kraftlinjer, og retningen til sistnevnte avhenger av den mekaniske effekten på brusken. Det er ingen blod- og lymfekar i det intercellulære stoffet i bruskvevet, derfor utføres ernæringen av bruskvevet ved diffust inntak av stoffer fra karene i perichondrium.
Aldersrelaterte endringer i brusk. De største endringene observeres i alderdommen, når antall kondroblaster i perichondrium og antall delende bruskceller reduseres. I kondrocytter avtar mengden av granulær EPS, Golgi-komplekset og mitokondrier, kondrocyttenes evne til å syntetisere glykosaminoglykaner og proteoglykaner går tapt. En reduksjon i mengden proteoglykaner fører til en reduksjon i hydrofilisiteten til bruskvevet, en svekkelse av bruskens permeabilitet og tilførsel av næringsstoffer. Dette fører til forkalkning av brusken, penetrering av blodkar inn i den og dannelse av beinstoff inne i brusken.
bruskvev , som bein, refererer til skjelettvev med en muskel-skjelettfunksjon. I henhold til klassifiseringen skilles tre typer bruskvev - hyalin, elastisk og fibrøs. Strukturelle funksjoner forskjellige typer brusk avhenger av dens plassering i kroppen, mekaniske forhold, individets alder.
Typer bruskvev: 1 - hyalin brusk; 2 - elastisk brusk; 3 - fibrøs brusk
Mest bred bruk mottatt fra en personhyalint bruskvev.
Det er en del av luftrøret, noen brusk i strupehodet, store bronkier, temafyser av bein, finnes i krysset mellom ribbeina og brystbenet og i noen andre områder av kroppen. Elastisk bruskvev er en del av auricleen, mellomstore bronkier og noen brusk i strupehodet. Fibrøs brusk er ofte funnet i krysset mellom sener og leddbånd med hyalin brusk, for eksempel mellomvirvelskiver.
Strukturen til alle typer brusk i generelt lignende: de inneholder celler og en intercellulær substans (matrise). En av egenskapene til det intercellulære stoffet i bruskvevet er dets høye vanninnhold: vanninnholdet varierer normalt fra 60 til 80%. Området okkupert av det intercellulære stoffet mer område okkupert av celler. Det intercellulære stoffet i bruskvev produseres av celler (kondroblaster og unge kondrocytter) og har en kompleks kjemisk sammensetning. Den er delt inn i den amorfe hovedsubstansen og den fibrillære komponenten, som utgjør omtrent 40 % av den tørre massen til den intercellulære substansen og er representert i det hyaline bruskvevet av kollagenfibriller dannet av type II kollagen, som diffunderer i forskjellige retninger. På histologiske preparater er fibriller usynlige, siden de har samme brytningsindeks som et amorft stoff. I det elastiske bruskvevet, sammen med kollagenfibriller, er det mange elastiske fibre som består av elastinprotein, som også produseres av bruskceller. Fibrøs brusk inneholder et stort antall bunter av kollagenfibre, bestående av type I og type II kollagen.
Ledende kjemiske forbindelser, som danner den viktigste amorfe substansen i bruskvev (kondromukoid), er sulfaterte glykosaminoglykaner (keratosulfater og kondroitinsulfater A og C) og nøytrale mukopolysakkarider, hvorav de fleste er representert av komplekse supramolekylære komplekser. I brusk har forbindelser av hyaluronsyremolekyler med proteoglykaner og med spesifikke sulfaterte glykosaminoglykaner blitt utbredt. Dette sikrer spesielle egenskaper bruskvev - mekanisk styrke og samtidig permeabilitet for organiske forbindelser, vann og andre stoffer som er nødvendige for å sikre den vitale aktiviteten til cellulære elementer. Markørforbindelsene som er mest spesifikke for det intercellulære stoffet i brusk er keratosulfater og visse varianter av kondroitinsulfater. De utgjør omtrent 30 % av den tørre massen av brusk.
Hovedcellene i bruskvev -kondroblaster og kondrocytter.
Kondroblasterer unge, udifferensierte celler. De ligger nær perichondrium, ligger enkeltvis og er preget av en avrundet eller oval form med ujevne kanter. En stor kjerne opptar en betydelig del av cytoplasmaet. Blant celleorganeller synteseorganeller dominerer - ribosomer og polysomer, granulært endoplasmatisk retikulum, Golgi-kompleks, mitokondrier; preget av inneslutninger av glykogen. Med generell histologisk farging av preparater med hematoksylin og eosin er kondroblaster svakt basofile. Strukturen til kondroblaster indikerer at disse cellene viser høy metabolsk aktivitet, spesielt assosiert med syntesen av intercellulær substans. Det er vist at i kondroblaster er syntesen av kollagen og ikke-kollagenproteiner romlig atskilt. Hele syklusen med syntese og utskillelse av høymolekylære komponenter av det intercellulære stoffet i funksjonelt aktive humane kondroblaster tar mindre enn en dag. Nydannede proteiner, proteoglykaner og glykosaminoglykaner befinner seg ikke direkte nær celleoverflaten, men spres diffust i betydelig avstand fra cellen i det tidligere dannede intercellulære stoffet. Blant kondroblaster er det også funksjonelt inaktive celler, hvis struktur er karakterisert dårlig utvikling syntetiske apparater. I tillegg mistet ikke en del av kondroblastene som ligger rett under perichondrium evnen til å dele seg.
Kondrocytter- Modne celler av bruskvev - okkuperer hovedsakelig de sentrale delene av brusken. Den syntetiske evnen til disse cellene er betydelig lavere enn for kondroblaster. Differensierte kondrocytter ligger oftest i bruskvev ikke alene, men i grupper på 2, 4, 8 celler. Dette er de såkalte isogene cellegruppene, som ble dannet som et resultat av deling av én bruskcelle. Strukturen til modne kondrocytter indikerer at de ikke er i stand til deling og en merkbar syntese av intercellulær substans. Men noen forskere mener at under visse forhold er mitotisk aktivitet i disse cellene fortsatt mulig. Funksjonen til kondrocytter er å opprettholde et visst nivå av metabolsk metabolske prosesser i bruskvev.
Isogene grupper av celler er lokalisert i bruskhulrom omgitt av en matrise. Formen på bruskceller i isogene grupper kan være forskjellig - rund, oval, fusiform, trekantet - avhengig av plasseringen på et bestemt område av brusk. Bruskhulene er omgitt av en smal, lettere enn hovedstoffet, stripe, og danner så å si et skall av bruskhulen. Dette skallet, som er preget av oksyfilisitet, kalles det cellulære territoriet, eller territoriell matrise. De fjernere områdene av det intercellulære stoffet kalles den interstitielle matrisen. Territoriale og interstitielle matriser er områder av intercellulær substans med forskjellige strukturelle og funksjonelle egenskaper. Innenfor den territoriale matrisen er kollagenfibriller orientert rundt overflaten av isogene cellegrupper. Vev av kollagenfibriller danner en vegg av lakuner. Mellomrommene mellom cellene i lakunaene er fylt med proteoglykaner. Den interstitielle matrisen er preget av svakt basofil eller oksyfil farge og tilsvarer de eldste delene av det intercellulære stoffet.
Dermed er det definitive bruskvevet preget av en strengt polarisert fordeling av celler avhengig av graden av deres differensiering. Nær perichondrium er de minst differensierte cellene - kondroblaster, som ser ut som celler forlenget parallelt med perichondrium. De syntetiserer aktivt det intercellulære stoffet og beholder den mitotiske evnen. Jo nærmere sentrum av brusken, jo mer differensierte cellene er, de er lokalisert i isogene grupper og er preget av en kraftig reduksjon i syntesen av komponenter i det intercellulære stoffet og fravær av mitotisk aktivitet.
I moderne vitenskapelig litteratur en annen type bruskvevsceller er beskrevet -kondroklaster. De forekommer bare under ødeleggelsen av bruskvev, og under forholdene i dets normale liv blir ikke oppdaget. I størrelse er kondroklaster mye større enn kondrocytter og kondroblaster, siden de inneholder flere kjerner i cytoplasmaet. Funksjonen til kondroklaster er assosiert med aktivering av bruskdegenerasjonsprosesser og deltakelse i fagocytose og lysis av fragmenter av ødelagte bruskceller og bruskmatrisekomponenter. Med andre ord, kondroklaster er makrofager av bruskvev som er en del av et enkelt makrofag-fagocyttsystem i kroppen.
Leddsykdommer
I OG. Mazurov
Bruskvev er funksjonelt iboende i støtterollen. Det fungerer ikke i spenning, som et tett bindevev, men takket være indre spenning godt motstår kompresjon og fungerer som en støtdemper for beinapparatet.
Dette spesielle vevet tjener til fast forbindelse av bein, og danner synkondrose. Dekker de artikulære overflatene til beinene, myker opp bevegelsen og friksjonen i leddene.
Bruskvev er veldig tett og samtidig ganske elastisk. Dens biokjemiske sammensetning er rik på tett amorft materiale. Brusk utvikler seg fra mellomliggende mesenkym.
På stedet for fremtidens brusk formerer mesenkymale celler seg raskt, prosessene deres forkortes og cellene er i nær kontakt med hverandre.
Deretter vises et mellomstoff, på grunn av hvilke mononukleære seksjoner er tydelig synlige i rudimentet, som er de primære bruskcellene - kondroblaster. De formerer seg og gir stadig flere masser av mellomstoffet.
Reproduksjonshastigheten av bruskceller i denne perioden er sterkt redusert, og på grunn av den store mengden mellomstoff er de langt fra hverandre. Snart mister cellene evnen til å dele seg ved mitose, men beholder fortsatt evnen til å dele seg amitotisk.
Men nå skiller ikke dattercellene seg langt, ettersom mellomstoffet som omgir dem har kondensert.
Derfor er bruskceller lokalisert i massen til hovedstoffet i grupper på 2-5 eller flere celler. Alle kommer fra én innledende celle.
En slik gruppe celler kalles isogen (isos - lik, identisk, genesis - forekomst).
Ris. en.
A - hyalin brusk i luftrøret;
B - elastisk brusk av kalvens auricle;
B - fibrocartilage av mellomvirvelskiven til kalven;
a - perichondrium; b ~ brusk; i - en eldre del av brusk;
- 1 - kondroblast; 2 - kondrocytt;
- 3 - isogen gruppe av kondrocytter; 4 - elastiske fibre;
- 5 - bunter av kollagenfibre; 6 - hovedstoffet;
- 7 - kondrocyttkapsel; 8 - basofil og 9 - oksyfil sone av hovedstoffet rundt den isogene gruppen.
Celler av den isogene gruppen deler seg ikke ved mitose, de gir lite mellomsubstans av en litt annen kjemisk oppbygning, som danner bruskkapsler rundt individuelle celler, og felt rundt den isogene gruppen.
Bruskkapsel som avslørt elektronmikroskopi, dannet av tynne fibriller konsentrisk plassert rundt cellen.
Følgelig, i begynnelsen av utviklingen av bruskvevet til dyr, skjer dens vekst ved å øke massen av brusken fra innsiden.
Da slutter den eldste delen av brusken, hvor cellene ikke formerer seg og det ikke dannes noe mellomstoff, å øke i størrelse, og bruskceller degenererer til og med.
Imidlertid stopper ikke veksten av brusk som helhet. Rundt den foreldede brusken skiller et lag med celler seg fra det omkringliggende mesenkymet, som blir til kondroblaster. De skiller ut mellomstoffet i brusken rundt seg og tykner gradvis med det.
Samtidig som de utvikler seg, mister kondroblaster evnen til å dele seg ved mitose, danner mindre mellomstoff og blir til kondrocytter. På brusklaget som er dannet på denne måten, på grunn av det omkringliggende mesenkymet, blir flere og flere lag av det lagt over hverandre. Følgelig vokser brusk ikke bare fra innsiden, men også fra utsiden.
Hos pattedyr er det: hyalin (glasaktig), elastisk og fibrøs brusk.
Hyalinbrusk (fig. 1--A) er den vanligste, melkehvit og noe gjennomskinnelig, så den kalles ofte glassaktig.
Den dekker leddflatene til alle bein; kystbrusk, brusk i luftrøret og noen brusk i strupehodet dannes fra den. Hyalinbrusk består, som alt vev i det indre miljøet, av celler og et mellomstoff.
Bruskceller er representert av kondroblaster og kondrocytter. Den skiller seg fra hyalinbrusk ved den sterke utviklingen av kollagenfibre, som danner bunter som ligger nesten parallelt med hverandre, som i sener!
Det er mindre amorft stoff i fibrøs brusk enn i hyalin. Avrundede lette celler av fibrobrusk ligger mellom fibrene i parallelle rader.
På steder hvor fibrobrusk er lokalisert mellom hyalinbrusk og dannet tett bindevev, observeres en gradvis overgang fra en type vev til en annen i strukturen. Nærmere bindevevet danner således kollagenfibre i brusk grove parallelle bunter, og bruskceller ligger i rader mellom dem, som fibrocytter av tett bindevev. Nærmere den hyaline brusken deles buntene inn i individuelle kollagenfibre som danner et delikat nettverk, og cellene mister sin riktige plassering.
 Oratorium som en prototype på journalistikk
Oratorium som en prototype på journalistikk Sitater om Napoleon - dslinkov — LiveJournal
Sitater om Napoleon - dslinkov — LiveJournal Meg hevn Mannen på bulldoseren ødela byen
Meg hevn Mannen på bulldoseren ødela byen