Hvilke organismer utviklet først et nervesystem? Hvilket dyr utviklet først et nervesystem? Generelle egenskaper ved nervesystemet
I evolusjonen har nervesystemet gjennomgått flere utviklingsstadier, som ble vendepunkter i den kvalitative organiseringen av aktivitetene. Disse stadiene er forskjellige i antall og typer nevronformasjoner, synapser, tegn på deres funksjonelle spesialisering og dannelsen av grupper av nevroner forbundet med felles funksjoner. Det er tre hovedstadier av den strukturelle organiseringen av nervesystemet: diffus, nodulær, rørformet.
Diffus Nervesystemet er det eldste, funnet i coelenterates (hydra). Et slikt nervesystem er preget av et mangfold av forbindelser mellom naboelementer, noe som gjør at eksitasjon fritt kan spre seg gjennom nervenettverket i alle retninger.
Denne typen nervesystem gir bred utskiftbarhet og dermed større pålitelighet i funksjon, men disse reaksjonene er upresise og vage.
Nodal typen nervesystem er typisk for ormer, bløtdyr og krepsdyr.
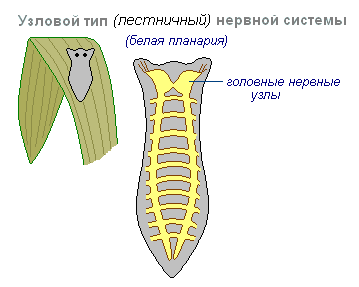
Det er preget av det faktum at forbindelsene til nerveceller er organisert på en bestemt måte, eksitasjon passerer langs strengt definerte baner. Denne organiseringen av nervesystemet viser seg å være mer sårbar. Skade på en node forårsaker dysfunksjon av hele organismen som helhet, men dens kvaliteter er raskere og mer nøyaktige.
Rørformet Nervesystemet er karakteristisk for akkordater; det inkluderer funksjoner av diffuse og nodulære. Nervesystemet til høyere dyr tok alt det beste: høy pålitelighet av den diffuse typen, nøyaktighet, lokalitet, hastighet på organisering av reaksjoner av nodaltype.
Den ledende rollen til nervesystemet
På det første stadiet av utviklingen av verden av levende vesener ble interaksjonen mellom de enkleste organismene utført gjennom vannmiljøet i det primitive havet, hvor de kjemiske stoffene som ble frigjort av dem kom inn i. Den første eldste formen for interaksjon mellom cellene i en flercellet organisme er kjemisk interaksjon gjennom metabolske produkter som kommer inn i kroppsvæskene. Slike metabolske produkter, eller metabolitter, er nedbrytningsprodukter av proteiner, karbondioksid, etc. Dette er den humorale overføringen av påvirkninger, den humorale mekanismen for korrelasjon eller forbindelser mellom organer.
Den humorale forbindelsen er preget av følgende funksjoner:
- mangel på en nøyaktig adresse som et kjemisk stoff som kommer inn i blodet eller andre kroppsvæsker sendes til;
- kjemikaliet sprer seg sakte;
- kjemikaliet virker i små mengder og brytes vanligvis raskt ned eller elimineres fra kroppen.
Humoriske forbindelser er felles for både dyre- og planteverdenen. På et visst utviklingsstadium av dyreverdenen, i forbindelse med utseendet til nervesystemet, dannes en ny, nervøs form for forbindelser og regulering, som kvalitativt skiller dyreverdenen fra planteverdenen. Jo høyere utviklingen av et dyrs organisme er, desto større rolle spiller interaksjonen mellom organer gjennom nervesystemet, som er betegnet som refleks. I høyere levende organismer regulerer nervesystemet humorale forbindelser. I motsetning til den humorale forbindelsen har nerveforbindelsen en presis retning til et spesifikt organ og til og med en gruppe celler; kommunikasjon utføres hundrevis av ganger raskere enn hastigheten på distribusjon av kjemikalier. Overgangen fra en humoral forbindelse til en nervøs forbindelse ble ikke ledsaget av ødeleggelsen av den humorale forbindelsen mellom cellene i kroppen, men av underordningen av nerveforbindelser og fremveksten av neurohumorale forbindelser.
På neste stadium av utviklingen av levende vesener vises spesielle organer - kjertler, der hormoner produseres, dannet fra matstoffer som kommer inn i kroppen. Hovedfunksjonen til nervesystemet er både å regulere aktiviteten til individuelle organer seg imellom, og i samspillet mellom kroppen som helhet og dets ytre miljø. Enhver påvirkning av det ytre miljøet på kroppen vises først av alt på reseptorene (sanseorganer) og utføres gjennom endringer forårsaket av det ytre miljøet og nervesystemet. Etter hvert som nervesystemet utvikler seg, blir dets høyeste avdeling – hjernehalvdelene – «leder og distributør av alle kroppens aktiviteter».
Struktur av nervesystemet
Nervesystemet er dannet av nervevev, som består av en enorm mengde nevroner- en nervecelle med prosesser.

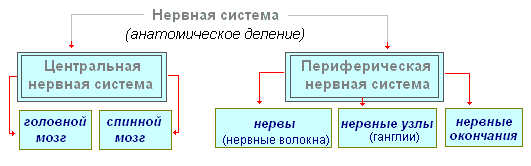
Nervesystemet er konvensjonelt delt inn i sentralt og perifert.

Sentralnervesystemet inkluderer hjernen og ryggmargen, og perifert nervesystem- nerver som strekker seg fra dem.
Hjernen og ryggmargen er en samling av nevroner. I et tverrsnitt av hjernen skilles hvit og grå substans. Grå substans består av nerveceller, og hvit substans består av nervefibre, som er prosesser av nerveceller. I forskjellige deler av sentralnervesystemet er plasseringen av hvit og grå substans forskjellig. I ryggmargen er grå materie plassert på innsiden, og hvit substans er på utsiden (hjernehalvdelene, lillehjernen), tvert imot, grå substans er på utsiden, hvit substans er på innsiden. I ulike deler av hjernen er det separate klynger av nerveceller (grå substans) plassert inne i den hvite substansen - kjerner. Klynger av nerveceller er også plassert utenfor sentralnervesystemet. De kalles noder og tilhører det perifere nervesystemet.
Refleksaktivitet i nervesystemet
Hovedformen for aktivitet i nervesystemet er refleksen. Refleks- kroppens reaksjon på endringer i det indre eller ytre miljøet, utført med deltakelse av sentralnervesystemet som svar på irritasjon av reseptorer.
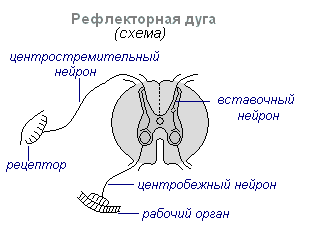
Ved enhver irritasjon overføres eksitasjon fra reseptorene langs sentripetale nervefibre til sentralnervesystemet, hvorfra det, gjennom interneuronet langs sentrifugalfibre, går til periferien til et eller annet organ, hvis aktivitet endres. Hele denne veien gjennom sentralnervesystemet til arbeidsorganet kalles refleksbue vanligvis dannet av tre nevroner: sensorisk, intercalary og motorisk. En refleks er en kompleks handling der et betydelig større antall nevroner deltar. Eksitasjon, som kommer inn i sentralnervesystemet, sprer seg til mange deler av ryggmargen og når hjernen. Som et resultat av samspillet mellom mange nevroner, reagerer kroppen på irritasjon.
Ryggmarg
Ryggmarg- en snor ca. 45 cm lang, 1 cm i diameter, plassert i ryggmargskanalen, dekket med tre hjernehinner: dura, arachnoid og myk (vaskulær).

Ryggmarg ligger i ryggmargskanalen og er en ledning som på toppen går inn i medulla oblongata og nederst ender i nivå med den andre lumbale vertebra. Ryggmargen består av grå substans som inneholder nerveceller og hvit substans som består av nervefibre. Grå substans ligger inne i ryggmargen og er omgitt på alle sider av hvit substans.
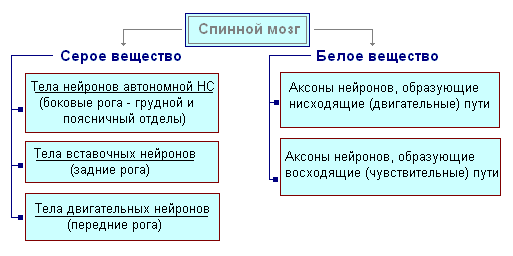
I et tverrsnitt ligner den grå substansen bokstaven H. Den skiller de fremre og bakre hornene, samt den forbindende tverrstangen, i midten av hvilken det er en smal kanal i ryggmargen som inneholder cerebrospinalvæske. I thoraxregionen er det laterale horn. De inneholder kroppene til nevroner som innerverer indre organer. Den hvite substansen i ryggmargen dannes av nerveprosesser. Korte prosesser forbinder deler av ryggmargen, og lange utgjør det ledende apparatet for bilaterale forbindelser med hjernen.

Ryggmargen har to fortykkelser - cervikal og lumbal, hvorfra nerver strekker seg til øvre og nedre ekstremiteter. 31 par spinalnerver oppstår fra ryggmargen. Hver nerve begynner fra ryggmargen med to røtter - fremre og bakre. Bakre røtter - følsom består av prosesser av sentripetale nevroner. Kroppene deres er lokalisert i spinalgangliene. Fremre røtter - motor- er prosesser av sentrifugale nevroner lokalisert i den grå substansen i ryggmargen. Som et resultat av sammensmeltningen av de fremre og bakre røttene, dannes en blandet spinalnerve. Ryggmargen inneholder sentre som regulerer de enkleste reflekshandlingene. Hovedfunksjonene til ryggmargen er refleksaktivitet og ledning av eksitasjon.
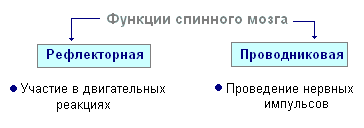
Den menneskelige ryggmargen inneholder reflekssentre for musklene i øvre og nedre ekstremiteter, svette og vannlating. Funksjonen til eksitasjon er at impulser fra hjernen til alle områder av kroppen og ryggen passerer gjennom ryggmargen. Sentrifugale impulser fra organer (hud, muskler) overføres gjennom stigende veier til hjernen. Langs nedadgående baner overføres sentrifugale impulser fra hjernen til ryggmargen, deretter til periferien, til organene. Når banene er skadet, er det tap av følsomhet i ulike deler av kroppen, et brudd på frivillige muskelsammentrekninger og evnen til å bevege seg.
Evolusjon av virveldyrhjernen
Dannelsen av sentralnervesystemet i form av et nevralrør vises først i akkordater. U lavere akkordater nevralrøret vedvarer hele livet, høyere- virveldyr - i embryonalstadiet dannes en nevrale plate på ryggsiden, som synker under huden og folder seg til et rør. I det embryonale utviklingsstadiet danner nevralrøret tre hevelser i den fremre delen - tre hjernevesikler, hvorfra deler av hjernen utvikler seg: den fremre vesikkelen gir forhjernen og diencephalon, den midterste vesikkelen blir til midthjernen, den bakre vesikkelen danner lillehjernen og medulla oblongata. Disse fem delene av hjernen er karakteristiske for alle virveldyr.
Til lavere virveldyr- fisk og amfibier - preget av en overvekt av mellomhjernen over andre deler. U amfibier Forhjernen forstørres noe og det dannes et tynt lag med nerveceller i taket av halvkulene – det primære medullærhvelvet, den eldgamle cortex. U reptiler Forhjernen øker betydelig på grunn av ansamlinger av nerveceller. Det meste av taket på halvkulene er okkupert av den gamle cortex. For første gang hos reptiler vises rudimentet til en ny cortex. Halvkulene i forhjernen kryper inn på andre deler, som et resultat av at det dannes en bøyning i diencephalon-regionen. Fra og med gamle krypdyr, blir hjernehalvdelene den største delen av hjernen.
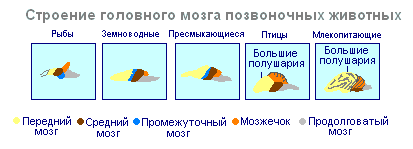
I hjernens struktur fugler og krypdyr mye til felles. På taket av hjernen er den primære cortex, mellomhjernen er godt utviklet. Men hos fugler, sammenlignet med krypdyr, øker den totale hjernemassen og den relative størrelsen på forhjernen. Lillehjernen er stor og har en foldet struktur. U pattedyr forhjernen når sin største størrelse og kompleksitet. Det meste av hjernestoffet består av neocortex, som fungerer som sentrum for høyere nervøs aktivitet. De mellomliggende og midtre delene av hjernen hos pattedyr er små. De ekspanderende halvkulene i forhjernen dekker dem og knuser dem under seg selv. Noen pattedyr har en glatt hjerne uten riller eller viklinger, men de fleste pattedyr har riller og viklinger i hjernebarken. Utseendet til spor og viklinger oppstår på grunn av veksten av hjernen med begrensede dimensjoner av skallen. Ytterligere vekst av cortex fører til utseendet av folding i form av spor og viklinger.
Hjerne
Hvis ryggmargen hos alle virveldyr er utviklet mer eller mindre likt, er hjernen betydelig forskjellig i størrelse og kompleksitet av struktur hos forskjellige dyr. Forhjernen gjennomgår spesielt dramatiske endringer under evolusjonen. Hos lavere virveldyr er forhjernen dårlig utviklet. Hos fisk er det representert av luktlappene og kjernene av grå substans i hjernens tykkelse. Den intensive utviklingen av forhjernen er assosiert med fremveksten av dyr på land. Det skiller seg inn i diencephalon og to symmetriske halvkuler, som kalles telencephalon. Grå substans på overflaten av forhjernen (cortex) vises først hos krypdyr, og utvikler seg videre hos fugler og spesielt hos pattedyr. Virkelig store forhjernehalvkuler blir bare hos fugler og pattedyr. I sistnevnte dekker de nesten alle andre deler av hjernen.
Hjernen er lokalisert i kraniehulen. Det inkluderer hjernestammen og telencephalon (hjernebarken).

Hjernestammen består av medulla oblongata, pons, midthjernen og diencephalon.
Medulla oblongata er en direkte fortsettelse av ryggmargen og går over i bakhjernen. Den beholder i utgangspunktet formen og strukturen til ryggmargen. I tykkelsen av medulla oblongata er det ansamlinger av grå substans - kjernene til kranialnervene. Bakakselen inkluderer lillehjernen og pons. Lillehjernen ligger over medulla oblongata og har en kompleks struktur. På overflaten av hjernehalvdelene danner grå substans cortex, og inne i lillehjernen - dens kjerner. I likhet med spinal medulla oblongata, utfører den to funksjoner: refleks og ledende. Imidlertid er refleksene til medulla oblongata mer komplekse. Dette gjenspeiles i dens betydning i reguleringen av hjerteaktivitet, tilstanden til blodårer, respirasjon og svette. Sentrene for alle disse funksjonene er lokalisert i medulla oblongata. Her er sentrene for tygging, suging, svelging, spytt og magesaft. Til tross for sin lille størrelse (2,5–3 cm), er medulla oblongata en viktig del av sentralnervesystemet. Skader på det kan forårsake død på grunn av pustestopp og hjerteaktivitet. Lederfunksjonen til medulla oblongata og pons er å overføre impulser fra ryggmargen til hjernen og tilbake.
I mellomhjernen de primære (subkortikale) sentrene for syn og hørsel er lokalisert, som utfører refleksive orienterende reaksjoner på lys- og lydstimulering. Disse reaksjonene kommer til uttrykk i ulike bevegelser av overkropp, hode og øyne mot stimuli. Mellomhjernen består av cerebrale peduncles og quadrigeminalis. Mellomhjernen regulerer og distribuerer tonen (spenningen) i skjelettmuskulaturen.
Diencephalon består av to avdelinger - thalamus og hypothalamus, som hver består av et stort antall kjerner i den visuelle thalamus og subthalamus-regionen. Gjennom den visuelle thalamus overføres sentripetale impulser til hjernebarken fra alle kroppens reseptorer. Ikke en eneste sentripetalimpuls, uansett hvor den kommer fra, kan passere til cortex og omgå de visuelle fjellene. Gjennom diencephalon kommuniserer således alle reseptorer med hjernebarken. I den subtuberkulære regionen er det sentre som påvirker metabolisme, termoregulering og endokrine kjertler.
Lillehjernen ligger bak medulla oblongata. Den består av grå og hvit substans. Men i motsetning til ryggmargen og hjernestammen, er den grå substansen - cortex - lokalisert på overflaten av lillehjernen, og den hvite substansen er plassert inne, under cortex. Lillehjernen koordinerer bevegelser, gjør dem klare og jevne, spiller en viktig rolle i å opprettholde balansen i kroppen i rommet, og påvirker også muskeltonen. Når lillehjernen er skadet, opplever en person en reduksjon i muskeltonus, bevegelsesforstyrrelser og endringer i gangart, tale bremser, etc. Men etter en tid gjenopprettes bevegelse og muskeltonus på grunn av det faktum at de intakte delene av sentralnervesystemet overtar funksjonene til lillehjernen.
Store halvkuler- den største og mest utviklede delen av hjernen. Hos mennesker utgjør de hoveddelen av hjernen og er dekket med cortex over hele overflaten. Grå substans dekker utsiden av halvkulene og danner hjernebarken. Den menneskelige hjernebarken har en tykkelse på 2 til 4 mm og består av 6–8 lag dannet av 14–16 milliarder celler, forskjellige i form, størrelse og funksjon. Under cortex er en hvit substans. Den består av nervefibre som forbinder cortex med de nedre delene av sentralnervesystemet og de individuelle lappene på halvkulene med hverandre.
Hjernebarken har viklinger atskilt av riller, som øker overflaten betydelig. De tre dypeste rillene deler halvkulene i fliker. Hver halvkule har fire lober: frontal, parietal, temporal, occipital. Eksitering av forskjellige reseptorer går inn i de tilsvarende oppfattende områdene i cortex, kalt soner, og herfra blir de overført til et spesifikt organ, og får det til å handle. Følgende soner skilles ut i cortex. Auditiv sone lokalisert i tinninglappen, mottar impulser fra auditive reseptorer.

Visuelt område ligger i den oksipitale regionen. Hit kommer impulser fra øyereseptorene.
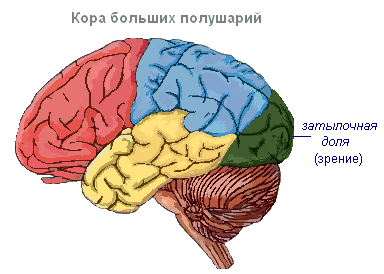
Olfaktorisk sone lokalisert på den indre overflaten av tinninglappen og er assosiert med reseptorer i nesehulen.
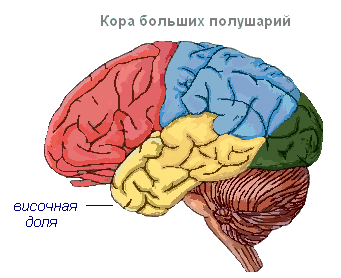
Sanse-motorisk sonen ligger i frontal- og parietallappene. Denne sonen inneholder de viktigste bevegelsessentrene for bena, overkroppen, armene, nakken, tungen og leppene. Det er også her talens sentrum ligger.
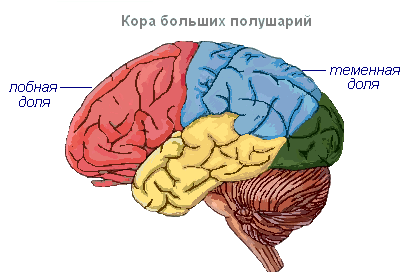
Hjernehalvdelene er den høyeste avdelingen av sentralnervesystemet, og kontrollerer funksjonen til alle organer hos pattedyr. Betydningen av hjernehalvdelene hos mennesker ligger også i det faktum at de representerer det materielle grunnlaget for mental aktivitet. I.P Pavlov viste at mental aktivitet er basert på fysiologiske prosesser som skjer i hjernebarken. Tenkning er assosiert med aktiviteten til hele hjernebarken, og ikke bare med funksjonen til dens individuelle områder.
| Hjerneavdelingen | Funksjoner | |
| Medulla oblongata | Dirigent | Forbindelse mellom spinal og overliggende deler av hjernen. |
| Refleks | Regulering av aktiviteten til respiratoriske, kardiovaskulære, fordøyelsessystemer:
|
|
| Pons | Dirigent | Kobler de lille hjernehalvdelene til hverandre og til hjernebarken. |
| Lillehjernen | Koordinasjon | Koordinering av frivillige bevegelser og opprettholdelse av kroppsposisjon i rommet. Regulering av muskeltonus og balanse |
| Midthjerne | Dirigent | Omtrentlig reflekser til visuelle og lydstimuli ( snur hodet og kroppen). |
| Refleks |
|
|
| Diencephalon | thalamus
hypothalamus
|
|
Cerebral cortex
Flate cerebral cortex hos mennesker er den omtrent 1500 cm 2, som er mange ganger større enn den indre overflaten av skallen. Denne store overflaten av cortex ble dannet på grunn av utviklingen av et stort antall spor og viklinger, som et resultat av at det meste av cortex (ca. 70%) er konsentrert i sporene. De største rillene i hjernehalvdelene er sentral, som går over begge halvkuler, og tidsmessig, som skiller tinninglappen fra resten. Hjernebarken har, til tross for sin lille tykkelse (1,5–3 mm), en svært kompleks struktur. Den har seks hovedlag, som er forskjellige i strukturen, formen og størrelsen på nevroner og forbindelser. Cortex inneholder sentrene til alle sensoriske (reseptor) systemer, representanter for alle organer og deler av kroppen. I denne forbindelse nærmer sentripetale nerveimpulser fra alle indre organer eller deler av kroppen cortex, og det kan kontrollere arbeidet deres. Gjennom hjernebarken lukkes betingede reflekser, gjennom hvilke kroppen hele tiden, gjennom hele livet, meget nøyaktig tilpasser seg de skiftende forholdene i tilværelsen, til miljøet.
Nervesystemet i en levende organisme er representert av et nettverk av kommunikasjoner som sikrer dets forbindelse med omverdenen og dens egne prosesser. Dens grunnleggende element er en nevron - en celle med prosesser (aksoner og dendritter) som overfører informasjon elektrisk og kjemisk.
Hensikten med nerveregulering
For første gang dukket nervesystemet opp i levende organismer på grunn av behovet for mer effektiv interaksjon med miljøet. Utviklingen av et enkelt nettverk for overføring av impulser bidro ikke bare til å oppfatte signaler fra utsiden. Takket være det ble det mulig å organisere sine egne livsprosesser for mer vellykket funksjon.
Under evolusjonen ble nervesystemets struktur mer kompleks: dens oppgave ble ikke bare dannelsen av en adekvat respons på ytre påvirkninger, men også organiseringen av sin egen oppførsel. I. P. Pavlov kalte denne metoden for å fungere
Interaksjon med det encellede miljøet
Nervesystemet dukket først opp i organismer som består av mer enn én celle, da det overfører signaler mellom nevroner som danner et nettverk. Men allerede i protozoer kan man observere evnen til å reagere på ytre stimuli gitt av intracellulære prosesser.
Nervesystemet til flercellede organismer er kvalitativt forskjellig fra en lignende formasjon i protozoer. Sistnevnte lokaliserer hele systemet av forbindelser innenfor metabolismen til en enkelt celle. Ciliatene "lærer" om ulike prosesser som skjer eksternt eller internt på grunn av endringer i sammensetningen av protoplasma og aktiviteten til noen andre strukturer. Flercellede levende vesener har et system bygget av funksjonelle enheter, som hver er utstyrt med sine egne metabolske prosesser.
Dermed dukker det for første gang opp et nervesystem hos noen som ikke har én, men flere celler, det vil si at prototypen er ledning av impulser i protozoer. På deres nivå av vital aktivitet produserer protoplasma strukturer som leder impulser. På samme måte, i mer komplekse levende vesener, utføres denne funksjonen av individet
Funksjoner av nervesystemet til coelenterates
Flercellede dyr som lever i kolonier deler ikke funksjoner seg imellom, og de har ennå ikke et nervenettverk. Det oppstår på det stadiet når ulike funksjoner i en flercellet organisme er differensiert.

For første gang dukker nervesystemet opp i hydra og andre coelenterater. Det er et nettverk som leder ikke-målrettede signaler. Strukturen er ennå ikke dannet, den er diffust fordelt i hele coelenteratets kropp. Ganglieceller og deres Nisslev-substans er ikke fullstendig dannet. Dette er den enkleste versjonen av nervesystemet.
Typen av dyrs motoriske aktivitet bestemmes av det diffuse nettverkslignende nervesystemet. Hydra utfører peristaltiske bevegelser, siden den ikke har spesielle kroppsdeler for bevegelse og andre bevegelser. For motorisk aktivitet krever det kontinuerlig tilkobling av de kontraktile elementene, og det krever at hoveddelen av de ledende cellene er plassert i den kontraktile delen. Hos hvilket dyr opptrer nervesystemet først i form av et diffust nettverk? De som er grunnleggerne av det menneskelige reguleringssystemet. Bevis på dette er det faktum at gastrulasjon er tilstede i utviklingen av dyreembryoer.
Funksjoner av nervesystemet til helminths
Påfølgende forbedring av nerveregulering var assosiert med utviklingen av bilateral symmetri i stedet for radial og dannelsen av klynger av nevroner i forskjellige deler av kroppen.

I form av ledninger vises nervesystemet først i 1 På dette stadiet er det representert av sammenkoblede hodefibre og dannede fibre som strekker seg fra dem. Sammenlignet med coelenterater er dette systemet mye mer komplekst. I helminths finnes grupper av nerveceller i form av noder og ganglier. Prototypen av hjernen er en ganglion i den fremre delen av kroppen som utfører regulatoriske funksjoner. Det kalles cerebral ganglion. Fra den løper to nervestammer langs hele kroppen, forbundet med hoppere.

Alle komponenter i systemet er ikke plassert utenfor, men er nedsenket i parenkymet og er dermed beskyttet mot skade. For første gang dukker nervesystemet opp hos flatormer sammen med de enkleste sanseorganene: berøring, syn og en følelse av balanse.
Funksjoner av nervesystemet til nematoder
Det neste utviklingsstadiet er dannelsen av en ringformasjon nær svelget og flere lange fibre som strekker seg fra den. Med slike egenskaper vises nervesystemet først i Peripharyngeal-ringen, som er en enkelt sirkulær ganglion og utfører funksjonene til det grunnleggende persepsjonsorganet. Ventralledningen og dorsalnerven er koblet til den.
Nervestammene til nematoder er lokalisert intraepitelialt, det vil si i hypodermale rygger. Rollen til persepsjonsorganer spilles av sensilla - setae, papiller, supplerende organer, amphider og phasmids. Alle av dem er utstyrt med blandet følsomhet.

De mest komplekse persepsjonsorganene til nematoder er amphider. De er sammenkoblet, kan være forskjellige i form og er plassert foran. Deres hovedoppgave er å gjenkjenne kjemiske midler som befinner seg langt fra kroppen. Noen rundormer har også reseptorer som oppfatter indre og ytre mekaniske påvirkninger. De kalles metanemes.
Funksjoner av ringlet nervesystemet
Dannelsen av ganglier i nervesystemet utvikler seg videre i annelider. I de fleste av dem skjer ganglionisering av bukstammene på en slik måte at hvert segment av ormen har et par nerveknuter som er forbundet med fibre til nabosegmenter. har en ventral nervestreng dannet av medullær ganglion og et par ledninger som kommer fra den. De strekker seg langs bukplanet. Oppfattende elementer er plassert foran og er representert av de enkleste øynene, luktceller, ciliære fossae og locatorer. Med parede noder dukket nervesystemet først opp i annelids, men senere utvikler det seg i leddyr. De har en forstørrelse av gangliene i hodet og en kombinasjon av noder i kroppen.
Elementer av det diffuse nettverket i det menneskelige nervesystemet
Toppen av den evolusjonære utviklingen av nervesystemet er utseendet til hjernen og ryggmargen hos mennesker. Men selv i nærvær av slike komplekse strukturer, er den opprinnelige diffuse organisasjonen bevart. Dette nettverket vikler inn hver eneste celle i kroppen: hud, blodårer, etc. Men med slike egenskaper dukker det for første gang opp et nervesystem hos noen som ikke engang hadde muligheten til å differensielt oppfatte miljøet.

Takket være disse "resterende" strukturelle enhetene har en person muligheten til å føle ulike påvirkninger selv i mikroskopiske områder. Kroppen kan reagere på utseendet til den minste fremmede agenten ved å utvikle beskyttende reaksjoner. Tilstedeværelsen av et diffust nettverk i det menneskelige nervesystemet bekreftes av laboratorieforskningsmetoder basert på innføring av et fargestoff.
Generell utviklingslinje for nervesystemet under evolusjon
De evolusjonære prosessene i nervesystemet fant sted i tre stadier:
- diffust nettverk;
- gangilia;
- ryggmarg og hjerne.

Strukturen og funksjonen til CNS er veldig forskjellig fra tidligere typer. Dens sympatiske inndeling inneholder ganglion og retikulære elementer. I sin fylogenetiske utvikling ble nervesystemet stadig mer fragmentert og differensiert. Det ganglioniske utviklingsstadiet skilte seg fra det retikulære stadiet i nærvær av nevroner som fremdeles er plassert over ledningssystemet.
Enhver levende organisme er i hovedsak en monolitt, bestående av forskjellige organer og deres systemer, som konstant og kontinuerlig interagerer med hverandre og med det ytre miljøet. Nervesystemet dukket først opp i coelenterater, det var et diffust nettverk som sørget for elementær ledning av impulser.
FOREDRAG OM EMNET: MENNESKELIG NERVESYSTEM
Nervesystemet er et system som regulerer aktivitetene til alle menneskelige organer og systemer. Dette systemet bestemmer: 1) den funksjonelle enheten til alle menneskelige organer og systemer; 2) hele organismens forbindelse med miljøet.
Fra synspunktet om å opprettholde homeostase, sikrer nervesystemet: opprettholdelse av parametrene til det indre miljøet på et gitt nivå; inkludering av atferdsreaksjoner; tilpasning til nye forhold hvis de vedvarer over lengre tid.
Nevron(nervecelle) - det viktigste strukturelle og funksjonelle elementet i nervesystemet; Mennesker har mer enn hundre milliarder nevroner. Et nevron består av en kropp og prosesser, vanligvis en lang prosess - et akson og flere kort forgrenede prosesser - dendritter. Langs dendritter følger impulser til cellekroppen, langs et akson – fra cellekroppen til andre nevroner, muskler eller kjertler. Takket være prosessene kontakter nevroner hverandre og danner nevrale nettverk og sirkler som nerveimpulser sirkulerer gjennom.
Et nevron er en funksjonell enhet i nervesystemet. Nevroner er mottakelige for stimulering, det vil si at de er i stand til å bli opphisset og overføre elektriske impulser fra reseptorer til effektorer. Basert på retningen for impulsoverføring skilles afferente nevroner (sensoriske nevroner), efferente nevroner (motoriske nevroner) og interneuroner.
Nervevev kalles eksiterbart vev. Som svar på en viss påvirkning oppstår en prosess med eksitasjon og sprer seg i den - rask opplading av cellemembraner. Fremveksten og forplantningen av eksitasjon (nerveimpuls) er den viktigste måten nervesystemet utfører sin kontrollfunksjon på.
Hovedforutsetningene for forekomsten av eksitasjon i celler: eksistensen av et elektrisk signal på membranen i hviletilstand - hvilemembranpotensialet (RMP);
evnen til å endre potensialet ved å endre permeabiliteten til membranen for visse ioner.
Cellemembranen er en semipermeabel biologisk membran, den har kanaler som lar kaliumioner passere gjennom, men det er ingen kanaler for intracellulære anioner, som holdes tilbake på den indre overflaten av membranen, og skaper en negativ ladning av membranen fra innsiden dette er hvilemembranpotensialet, som er gjennomsnittlig - – 70 millivolt (mV). Det er 20-50 ganger flere kaliumioner i cellen enn utenfor, dette opprettholdes gjennom hele livet ved hjelp av membranpumper (store proteinmolekyler som er i stand til å transportere kaliumioner fra det ekstracellulære miljøet til innsiden). MPP-verdien bestemmes av overføringen av kaliumioner i to retninger:
1. fra utsiden inn i cellen under påvirkning av pumper (med et stort energiforbruk);
2. fra cellen til utsiden ved diffusjon gjennom membrankanaler (uten energiforbruk).
I eksitasjonsprosessen spilles hovedrollen av natriumioner, som alltid er 8-10 ganger mer rikelig utenfor cellen enn inne. Natriumkanaler er lukket når cellen er i ro for å åpne dem, er det nødvendig å handle på cellen med en tilstrekkelig stimulans. Hvis stimuleringsterskelen nås, åpnes natriumkanalene og natrium kommer inn i cellen. I løpet av tusendeler av et sekund vil membranladningen først forsvinne og deretter endres til det motsatte - dette er den første fasen av aksjonspotensialet (AP) - depolarisering. Kanalene lukkes - toppen av kurven, så gjenopprettes ladningen på begge sider av membranen (på grunn av kaliumkanaler) - repolariseringsstadiet. Eksitasjonen stopper og mens cellen er i ro bytter pumpene ut natriumet som kom inn i cellen med kalium, som forlot cellen.

En PD fremkalt på et hvilket som helst tidspunkt på en nervefiber i seg selv blir irriterende for naboseksjoner av membranen, og forårsaker PD i dem, som igjen eksiterer flere og flere deler av membranen, og dermed sprer seg gjennom hele cellen. I fibre dekket med myelin vil AP bare forekomme i områder fri for myelin. Derfor øker hastigheten på signalutbredelsen.


Overføringen av eksitasjon fra celle til en annen skjer gjennom en kjemisk synapse, som er representert ved kontaktpunktet til to celler. Synapsen er dannet av presynaptiske og postsynaptiske membraner og den synaptiske kløften mellom dem. Eksitasjon i cellen som følge av AP når området av den presynaptiske membranen der synaptiske vesikler er lokalisert, hvorfra et spesielt stoff, transmitteren, frigjøres. Senderen som kommer inn i gapet beveger seg til den postsynaptiske membranen og binder seg til den. Porer åpner seg i membranen for ioner, de beveger seg inn i cellen og eksitasjonsprosessen skjer

I cellen blir altså det elektriske signalet omdannet til et kjemisk, og det kjemiske signalet igjen til et elektrisk. Signaloverføring i en synapse skjer langsommere enn i en nervecelle, og er også ensidig, siden transmitteren frigjøres kun gjennom den presynaptiske membranen, og kun kan binde seg til reseptorer i den postsynaptiske membranen, og ikke omvendt.
Mediatorer kan forårsake ikke bare eksitasjon, men også hemming i celler. I dette tilfellet åpner porene seg på membranen for ioner som styrker den negative ladningen som fantes på membranen i hvile. En celle kan ha mange synaptiske kontakter. Et eksempel på en mediator mellom et nevron og en skjelettmuskelfiber er acetylkolin.
Nervesystemet er delt inn i sentralnervesystemet og det perifere nervesystemet.
I sentralnervesystemet skilles det mellom hjernen, hvor hovednervesentrene og ryggmargen er konsentrert, og her er det lavere nivåsentre og veier til perifere organer.
Perifert snitt - nerver, nerveganglier, ganglier og plexuser.
Hovedmekanismen for aktiviteten til nervesystemet er refleks. En refleks er enhver reaksjon fra kroppen på en endring i det ytre eller indre miljøet, som utføres med deltakelse av sentralnervesystemet som svar på irritasjon av reseptorer. Det strukturelle grunnlaget for refleksen er refleksbuen. Den inkluderer fem påfølgende lenker:
1 - Reseptor - en signaleringsenhet som oppfatter påvirkning;
2 - Afferent nevron - bringer et signal fra reseptoren til nervesenteret;
3 - Interneuron - sentral del av buen;
4 - Efferent nevron - signalet kommer fra sentralnervesystemet til den utøvende strukturen;
5 - Effektor - en muskel eller kjertel som utfører en bestemt type aktivitet

Hjerne består av klynger av nervecellelegemer, nervekanaler og blodårer. Nervekanaler danner den hvite substansen i hjernen og består av bunter av nervetråder som leder impulser til eller fra ulike deler av den grå substansen i hjernen – kjerner eller sentre. Baner forbinder ulike kjerner, så vel som hjernen og ryggmargen.
Funksjonelt kan hjernen deles inn i flere seksjoner: forhjernen (bestående av telencephalon og diencephalon), mellomhjernen, bakhjernen (bestående av lillehjernen og pons) og medulla oblongata. Medulla oblongata, pons og midthjernen kalles samlet hjernestammen.

Ryggmarg plassert i ryggmargskanalen, og beskytter den pålitelig mot mekanisk skade.
Ryggmargen har en segmentell struktur. To par fremre og bakre røtter strekker seg fra hvert segment, som tilsvarer en ryggvirvel. Det er totalt 31 par nerver.
Ryggrøttene er dannet av sensoriske (afferente) nevroner, deres kropper er lokalisert i gangliene, og aksonene går inn i ryggmargen.
De fremre røttene er dannet av aksonene til efferente (motoriske) nevroner, hvis kropper ligger i ryggmargen.
Ryggmargen er konvensjonelt delt inn i fire seksjoner - cervikal, thorax, lumbal og sakral. Den lukker et stort antall refleksbuer, noe som sikrer regulering av mange kroppsfunksjoner.
Det grå sentrale stoffet er nerveceller, det hvite er nervefibre.

Nervesystemet er delt inn i somatisk og autonomt.
TIL somatisk nervøs system (fra det latinske ordet "soma" - kropp) refererer til en del av nervesystemet (både cellelegemer og deres prosesser), som kontrollerer aktiviteten til skjelettmuskulatur (kropp) og sanseorganer. Denne delen av nervesystemet styres i stor grad av vår bevissthet. Det vil si at vi er i stand til å bøye eller rette ut en arm, et ben osv. Vi klarer imidlertid ikke bevisst å slutte å oppfatte for eksempel lydsignaler.
Autonom nervøs system (oversatt fra latin "vegetativ" - plante) er en del av nervesystemet (både cellelegemer og deres prosesser), som kontrollerer prosessene for metabolisme, vekst og reproduksjon av celler, det vil si funksjoner som er felles for både dyr og planteorganismer . Det autonome nervesystemet er for eksempel ansvarlig for aktiviteten til indre organer og blodårer.
Det autonome nervesystemet er praktisk talt ikke kontrollert av bevissthet, det vil si at vi ikke er i stand til å lindre en spasme i galleblæren etter ønske, stoppe celledeling, stoppe tarmaktivitet, utvide eller trekke sammen blodkar.
Som kjent opptrer nervesystemet først hos lavere flercellede virvelløse dyr. Fremveksten av nervesystemet er en viktig milepæl i utviklingen av dyreverdenen, og i denne forbindelse er selv primitive flercellede virvelløse dyr kvalitativt forskjellige fra protozoer. Et viktig poeng her er den skarpe akselerasjonen av eksitasjonsledning i nervevev: i uprotoplasma overstiger ikke eksitasjonsledningshastigheten 1-2 mikron per sekund, men selv i det mest primitive nervesystemet, bestående av nerveceller, er det 0,5 meter per sekund!
Nervesystemet eksisterer i lavere flercellede organismer i svært forskjellige former: retikulert (for eksempel i hydra), ring (maneter), radial (sjøstjerne) og bilateral. Den bilaterale formen er representert i nedre (tarm) flatorm og primitive bløtdyr (kiton) bare av et nettverk som ligger nær overflaten av kroppen, men flere langsgående ledninger utmerker seg ved kraftigere utvikling. Etter hvert som nervesystemet utvikler seg gradvis, synker det under muskelvevet, og de langsgående snorene blir mer uttalte, spesielt på den ventrale siden av kroppen. Samtidig blir den fremre enden av kroppen stadig viktigere, hodet vises (prosessen med cephalization), og med det hjernen - akkumulering og komprimering av nerveelementer i fremre ende. Til slutt, i høyere ormer, erverver sentralnervesystemet allerede fullt ut den typiske strukturen til "nervestigen", der hjernen er plassert over fordøyelseskanalen og er forbundet med to symmetriske kommissurer ("periofaryngeal ring") med de subfaryngeale gangliene lokalisert på abdominalsiden og deretter med parede abdominale nervestammer. De essensielle elementene her er gangliene, og det er derfor de også snakker om det ganglioniske nervesystemet, eller "ganglionstigen." Hos noen representanter for denne dyregruppen (for eksempel igler), kommer nervestammene så tett sammen at en "nervekjede" oppnås.
Kraftige ledende fibre går fra gangliene, som utgjør nervestammene. I gigantiske fibre utføres nerveimpulser mye raskere på grunn av deres store diameter og små antall synaptiske forbindelser (kontaktsteder mellom aksonene til noen nerveceller og dendrittene og cellekroppene til andre celler). Når det gjelder de cefaliske gangliene, dvs. hjerne, så er de mer utviklet hos mer aktive dyr, som også har de mest utviklede reseptorsystemene.
Opprinnelsen og utviklingen av nervesystemet bestemmes av behovet for å koordinere de forskjellige kvalitetsfunksjonelle enhetene til en flercellet organisme, harmonisere prosessene som skjer i forskjellige deler av den når de samhandler med det ytre miljøet, og sikre aktiviteten til en kompleks organisme som et enkelt integrert system. Bare et koordinerende og organiserende senter, som sentralnervesystemet, kan gi fleksibilitet og variasjon i kroppens respons i en flercellet organisasjon.
Prosessen med cephalisapia var også av stor betydning i denne forbindelse, dvs. separasjon av hodeenden av organismen og det tilhørende utseendet til hjernen. Bare i nærvær av en hjerne er virkelig sentralisert "koding" av signaler som kommer fra periferien og dannelse av integrerte "programmer" av medfødt atferd mulig, for ikke å nevne en høy grad av koordinering av all ekstern aktivitet til dyret.
Selvfølgelig avhenger nivået av mental utvikling ikke bare av strukturen til nervesystemet. For eksempel har hjuldyr, nært beslektet med annelider, også, i likhet med dem, et bilateralt nervesystem og hjerne, samt spesialiserte sensoriske og motoriske nerver. Imidlertid skiller hjuldyrene seg lite fra ciliater i størrelse, utseende og livsstil, hjuldyr er svært like sistnevnte i oppførsel og viser ikke høyere mentale evner enn ciliater. Dette viser igjen at den ledende faktoren for utvikling av mental aktivitet ikke er den generelle strukturen, men de spesifikke leveforholdene til dyret, arten av dets relasjoner og interaksjoner med miljøet. Samtidig viser dette eksemplet nok en gang hvor nøye man må nærme seg vurderingen av "høyere" og "lavere" karakterer når man sammenligner organismer som inntar forskjellige fylogenetiske posisjoner, spesielt når man sammenligner protozoer og flercellede virvelløse dyr.
3.1. Opprinnelse og funksjoner til nervesystemet.
Nervesystemet hos alle dyr er av ektodermal opprinnelse. Den utfører følgende funksjoner:
Kommunikasjon av organismen med miljøet (oppfatning, overføring av irritasjon og respons på irritasjon);
Sammenkoblingen av alle organer og organsystemer til en enkelt helhet;
Nervesystemet ligger til grunn for dannelsen av høyere nerveaktivitet.
3.2. Evolusjon av nervesystemet blant virvelløse dyr.
Nervesystemet dukket først opp i coelenterates og hadde diffus eller retikulær type nervesystemet, dvs. Nervesystemet er et nettverk av nerveceller fordelt over hele kroppen og forbundet med tynne prosesser. Den har en typisk struktur i hydra, men allerede i maneter og polypper vises klynger av nerveceller på visse steder (nær munnen, langs kantene av paraplyen), disse klynger av nerveceller er forløperne til sanseorganer.
Videre følger utviklingen av nervesystemet banen for konsentrasjon av nerveceller på visse steder i kroppen, dvs. langs banen for dannelse av nerveknuter (ganglia). Disse nodene oppstår først og fremst der celler som oppfatter irritasjon fra omgivelsene befinner seg. Med radiell symmetri oppstår således en radial type nervesystem, og med bilateral symmetri oppstår konsentrasjonen av nerveganglier i den fremre enden av kroppen. Parede nervestammer som strekker seg langs kroppen strekker seg fra hodenodene. Denne typen nervesystem kalles ganglionstamme.
Denne typen nervesystem har en typisk struktur hos flatorm, d.v.s. i den fremre enden av kroppen er det parede ganglier, hvorfra nervefibre og sanseorganer strekker seg fremover, og nervestammer som løper langs kroppen.
Hos rundorm går de cefaliske gangliene sammen til en perifaryngeal nervering, hvorfra nervestammene også strekker seg langs kroppen.
I annelider dannes en nervekjede, d.v.s. Uavhengige parede nerveknuter dannes i hvert segment. Alle av dem er forbundet med både langsgående og tverrgående tråder. Som et resultat får nervesystemet en stigelignende struktur. Ofte kommer begge kjedene nærmere hverandre, og kobles sammen langs den midtre delen av kroppen til en uparet abdominal nervekjede.
Leddyr har samme type nervesystem, men antallet nerveganglier avtar og størrelsen øker, spesielt i hodet eller cephalothorax, d.v.s. prosessen med cefalisering er i gang.
Hos bløtdyr er nervesystemet representert av noder i forskjellige deler av kroppen, forbundet med hverandre med ledninger og nerver som strekker seg fra nodene. Gastropoder har pedal-, cerebrale og pleura-viscerale noder; i muslinger - pedal og pleural-visceral; hos blæksprutter - pleural-viscerale og cerebrale nerveganglier. En opphopning av nervevev observeres rundt svelget til blæksprutter.
3.3. Evolusjon av nervesystemet i akkordater.
Nervesystemet til akkordater er representert av nevralrøret, som differensierer til hjernen og ryggmargen.
I nedre chordater har nevralrøret utseende som et hult rør (neurocoel) med nerver som strekker seg fra røret. I lansetten dannes en liten utvidelse i hodedelen - hjernens rudiment. Denne utvidelsen kalles ventrikkelen.
I høyere chordater dannes tre hevelser i den fremre enden av nevralrøret: fremre, midtre og bakre vesikler. Fra den første cerebrale vesikkelen dannes deretter forhjernen og diencephalon, fra den midtre cerebrale vesikkelen - mesencephalon, fra bakre - cerebellum og medulla oblongata, som går inn i ryggmargen.
I alle klasser av virveldyr består hjernen av 5 seksjoner (fremre, mellomliggende, midtre, bakre og medulla), men graden av deres utvikling er ikke den samme hos dyr av forskjellige klasser.
Således, i cyclostomer, er alle deler av hjernen plassert etter hverandre i et horisontalt plan. Medulla oblongata går direkte inn i ryggmargen med den sentrale kanalen i nutria.
Hos fisk er hjernen mer differensiert sammenlignet med cyclostomer. Volumet av forhjernen økes, spesielt hos lungefisk, men forhjernen er ennå ikke delt inn i halvkuler og fungerer funksjonelt som det høyeste luktsenteret. Taket på forhjernen er tynt, den består kun av epitelceller og inneholder ikke nervevev. I diencephalon, som pineal- og hypofysen er forbundet med, er hypothalamus lokalisert, som er sentrum av det endokrine systemet. Den mest utviklede hos fisk er mellomhjernen. De optiske lappene kommer godt til uttrykk i den. I området av midthjernen er det en bøy karakteristisk for alle høyere virveldyr. I tillegg er mellomhjernen et analysesenter. Lillehjernen, som er en del av bakhjernen, er godt utviklet på grunn av kompleksiteten til bevegelse hos fisk. Det er et senter for koordinering av bevegelse. Størrelsen varierer avhengig av bevegelsesaktiviteten til forskjellige fiskearter. Medulla oblongata gir kommunikasjon mellom de høyere delene av hjernen og ryggmargen og inneholder sentrene for respirasjon og sirkulasjon.
10 par kraniale nerver kommer ut fra fiskehjernen.
Denne typen hjerne, der det høyeste integrasjonssenteret er midthjernen, kalles ikthyopsid.
Hos amfibier er nervesystemet i sin struktur nær nervesystemet til lungefisk, men kjennetegnes ved betydelig utvikling og fullstendig separasjon av sammenkoblede langstrakte halvkuler, samt svak utvikling av lillehjernen, som skyldes den lave mobiliteten til amfibier. og monotonien i deres bevegelser. Men amfibier utviklet et tak for forhjernen, kalt det primære medullærhvelvet - archipallium. Antall kraniale nerver, som hos fisk, er ti. Og hjernetypen er den samme, dvs. iktyopsid.
Dermed har alle anamni (syklostomer, fisk og amfibier) en ichthyopsid type hjerne.
I strukturen til hjernen til reptiler som tilhører høyere virveldyr, dvs. for fostervann er trekkene til en progressiv organisasjon tydelig uttrykt. Forhjernehalvdelene har en betydelig overvekt over andre deler av hjernen. Ved basen deres er det store ansamlinger av nerveceller - striatum. Øyene i den gamle cortex, archicortex, vises på laterale og mediale sider av hver halvkule. Størrelsen på mellomhjernen reduseres, og den mister sin betydning som et ledende senter. Bunnen av forhjernen blir analysesenteret, dvs. stripete kropper. Denne typen hjerne kalles sauropsid eller striatal. Lillehjernen øker i størrelse på grunn av de forskjellige bevegelsene til krypdyr. Medulla oblongata danner en skarp bøyning, karakteristisk for alle fostervann. Det er 12 par kraniale nerver som forlater hjernen.
Samme type hjerne er karakteristisk for fugler, men med noen funksjoner. Forhjernehalvdelene er relativt store. Luktlappene hos fugler er dårlig utviklet, noe som indikerer luktens rolle i fuglenes liv. I kontrast er mellomhjernen representert av store optiske fliker. Lillehjernen er godt utviklet, 12 par nerver kommer ut av hjernen.
Hjernen hos pattedyr når sin maksimale utvikling. Hemisfærene er så store at de dekker mellomhjernen og lillehjernen. Hjernebarken er spesielt utviklet, området er økt på grunn av viklinger og riller. Cortex har en svært kompleks struktur og kalles den nye cortex - neocortex. Et sekundært medullærhvelv, neopallium, dukker opp. Store luktelapper er plassert foran halvkulene. Diencephalon inkluderer, som andre klasser, pinealkjertelen, hypofysen og hypothalamus. Mellomhjernen er relativt liten, den består av fire tuberkler - quadrigeminalen. Den fremre cortex er forbundet med den visuelle analysatoren, den bakre med den auditive. Sammen med forhjernen utvikler lillehjernen seg mye. Det er 12 par kraniale nerver som forlater hjernen. Analysesenteret er hjernebarken. Denne typen hjerne kalles brystkreft.
3.4. Anomalier og misdannelser i nervesystemet hos mennesker.
1. Acefali- fravær av hjerne, hvelv, hodeskalle og ansiktsskjelett; denne lidelsen er assosiert med underutvikling av det fremre nevralrøret og er kombinert med defekter i ryggmargen, bein og indre organer.
2. Anencefali- fravær av hjernehalvdelene og skalletaket med underutvikling av hjernestammen og er kombinert med andre utviklingsdefekter. Denne patologien er forårsaket av ikke-lukking (dysraphism) av hodet av nevralrøret. I dette tilfellet utvikler ikke beinene på taket av skallen seg, og beinene i bunnen av skallen viser forskjellige anomalier. Anencefali er uforenlig med livet, gjennomsnittlig frekvens er 1/1500, og er mer vanlig hos kvinnelige fostre.
3. Atelencefali- stans av utviklingen (heterokroni) av den fremre delen av nevralrøret på stadiet av tre vesikler. Som et resultat dannes ikke hjernehalvdelene og subkortikale kjerner.
4. Prosencefali– telencephalon er delt av et langsgående spor, men i dybden forblir begge halvkulene forbundet med hverandre.
5. Holoprosencefali– telencephalon er ikke delt inn i halvkuler og ser ut som en halvkule med et enkelt hulrom (ventrikkel).
6. Alobar prosencefali– delingen av telencephalon er bare i den bakre delen, og frontallappene forblir udelte.
7. Aplasi eller hypoplasi av corpus callosum– helt eller delvis fravær av en kompleks kommissur i hjernen, dvs. corpus callosum.
8. Hydroencefali- atrofi av hjernehalvdelene i kombinasjon med hydrocephalus.
9. Agiriya- fullstendig fravær av riller og viklinger (glatt hjerne) i hjernehalvdelene.
10. Mikrogyria- reduksjon i antall og volum av furer.
11. Medfødt hydrocephalus- obstruksjon av en del av det ventrikulære systemet i hjernen og dets utganger, det er forårsaket av en primær forstyrrelse i utviklingen av nervesystemet.
12. Spina bifida- en defekt i lukkingen og separasjonen av spinalnevralrøret fra hudektodermen. Noen ganger er denne anomalien ledsaget av diplomyelia, der ryggmargen er delt langs en viss lengde i to deler, hver med sin egen sentrale fordypning.
13. Iniencephaly- en sjelden anomali, uforenlig med livet, forekommer oftere hos kvinnelige fostre. Dette er en grov anomali i bakhodet og hjernen. Hodet snus slik at ansiktet vender oppover. Dorsalt fortsetter hodebunnen inn i huden i den lumbodorsale eller sakrale regionen.
 Slaget om tankskipet Kolobanov, som gikk ned i historien
Slaget om tankskipet Kolobanov, som gikk ned i historien Hvordan beveger asteroider seg? Bevegelse av asteroider. Form og rotasjon av asteroider
Hvordan beveger asteroider seg? Bevegelse av asteroider. Form og rotasjon av asteroider Berserkers: The Vikings Who Became the "Berserker Dogs of War" We Know
Berserkers: The Vikings Who Became the "Berserker Dogs of War" We Know