Inkludering av ytre stimuli i drømmer. Virkning av irriterende stoffer på menneskekroppen
Irritanter- dette er faktorer i det ytre eller indre miljøet som har en reserve av energi, og når de utsettes for vev, blir effekten deres registrert biologisk reaksjon.
Klassifisering av stimuli avhenger av hva som legges til grunn:
1.På din egen måte natur irriterende stoffer er:
kjemisk
fysisk
mekanisk
termisk
biologiske
2. Av biologisk korrespondanse, det vil si hvor mye stimulansen tilsvarer et gitt vev:
tilstrekkelig– stimuli som samsvarer av dette stoffet. For eksempel, for netthinnen i øyet, lys - alle andre stimuli tilsvarer ikke netthinnen, for muskelvev
– nerveimpuls, etc.; utilstrekkelig – stimuli som ikke stemmer overens av dette stoffet For eksempel, for netthinnen i øyet, lys - alle andre stimuli tilsvarer ikke netthinnen, for. For netthinnen i øyet vil alle stimuli unntatt lys være utilstrekkelige, men for
alle stimuli unntatt nerveimpulsen. 3. Av styrke
– Det er fem hovedirriterende stoffer: underterskelstimuli
– dette er styrken til stimulansen der ingen respons oppstår; terskel stimulus - dette er minimumskraften som forårsaker en respons med uendelig virkningsvarighet. Denne kraften kalles også rheobase
– den er unik for hvert stoff; overterskel , eller;
submaksimal maksimal stimulans - dette er minimumskraften som maksimal respons oppstår ved;
vevsreaksjon supramaksimale stimuli
– med disse stimuliene er vevsreaksjonen enten maksimal, eller avtar, eller forsvinner midlertidig. terskel Det er én terskel for hvert vev
, en maksimum og mange underterskel, suprathreshold og supramaximal. Irritasjon – dette er eventuelle effekter på vevet. Som svar på irritasjon oppstår de biologiske reaksjoner
stoffer. Irritabilitet - Dette er en universell egenskap til levende materie og gjenspeiler evnen til ethvert levende vev til å endre sin ikke-spesifikk aktivitet
under påvirkning av irritasjon.
Billett 3. Begrepene eksitabilitet og opphisselse. Det er tre funksjonelle tilstander stoffer:.
hvile, eksitasjon og hemming Tilstand fred
hvile, eksitasjon og hemming – dette er en passiv prosess der det ikke er noen ytre uttrykte manifestasjoner av spesifikk aktivitet (sammentrekning, sekresjon, etc.). spenning Og bremsing - Dette, hvor i det ene tilfellet øker den spesifikke aktiviteten til vevet (eksitasjon), og i det andre forsvinner manifestasjonen av den spesifikke aktiviteten enten fullstendig eller avtar, selv om stimulansen fortsetter å virke på vevet.
To typer biologiske reaksjoner:
spesiell
uspesifikke
Spesifikke reaksjoner karakteristisk for noe strengt definert vev (en spesifikk reaksjon av muskelvev er en sammentrekning, for kjertelvev er det frigjøring av et sekret eller hormon, for nervevev er generering og overføring av nerveimpulser). Derfor har spesialisert vev spesifikke aktiviteter.
Uspesifikke reaksjoner karakteristisk for ethvert levende vev.
For eksempel en endring i metabolsk hastighet, en endring i hvilemembranpotensialet, en endring i ionegradienten, etc. Eksitabilitet – dette er en egenskap ved spesialisert vev og reflekterer evne vev reagerer på irritasjon ved å endre deres spesifikke reaksjoner
. Eksitabiliteten til et vev bestemmes av dets terskelstyrke: jo lavere terskelstyrke, jo større eksitabilitet av vevet. - dette er minimumskraften som maksimal respons oppstår ved
Exitasjon- Dette er spesifikt
Terskel for eksitabilitet (spenning) - den minste styrken til stimulansen som forårsaker minst opphisselse. Ved terskeleksitasjon er aktiviteten til et organ eller vev ekstremt liten. Styrken til stimulus mindre enn terskelen kalles subterskel, mer enn terskelen kalles suprathreshold. Jo større eksitabilitet vevet er, jo lavere er terskelen, og omvendt. Med en sterkere stimulus er det større eksitasjon, og følgelig øker aktiviteten til det opphissede organet. For eksempel, jo sterkere irritasjonen er mer høyde- sammentrekning av skjelettmuskulatur. Jo sterkere stimulansen er, desto kortere varigheten av virkningen, noe som forårsaker minimal opphisselse, og omvendt. Nyttig tid
minst tid
virkningen av en stimulus av terskelstyrke, eller rheobase, som forårsaker minimal eksitasjon. Denne tiden er imidlertid vanskelig å bestemme, så den korteste virkningstiden for den doble rheobase-stimulusen, som kalles kronaksi, bestemmes. Billett 4. Historien om oppdagelsen av bioelektriske fenomener. Arten av opphisselse. Opprinnelsen til læren om "dyreelektrisitet", dvs. bioelektriske fenomener, som oppstår i levende vev, dateres tilbake til andre halvdel av 1700-tallet. Kort tid etter oppdagelsen av Leyden-krukken ble det vist at noen fisk (elektrisk stråle, elektrisk ål. Samtidig foreslo J. Priestley at forplantningen av en nerveimpuls er en strøm av "elektrisk væske" langs nerven, og Bertolon prøvde å bygge en teori om medisin, og forklarte forekomsten av sykdommer ved overskudd og mangel på denne væsken. i kroppen.
Et forsøk på å konsekvent utvikle læren om "dyreelektrisitet" ble gjort av L. Galvani i hans berømte "Treatise on the Forces of Electricity in Motion" (1791). Mens han studerte den fysiologiske påvirkningen av elektriske maskinutladninger, så vel som atmosfærisk elektrisitet under lynutladninger, gjorde Galvani i sine eksperimenter brukte stoffet bakbena frosker koblet til ryggraden.
Han hang dette preparatet på en kobberkrok til jernrekkverket på balkongen, og la merke til at når froskebein svaiet i vinden, trakk musklene deres seg sammen for hver berøring av rekkverket. Basert på dette kom Galvani til den konklusjonen at rykningene i bena var forårsaket av "dyreelektrisitet" med opprinnelse i froskens ryggmarg og overført gjennom metallledere (kroken og balkongrekkverket) til musklene i bena. Galvanis eksperimenter ble gjentatt av A. Volta (1792) og fastslo at fenomenene beskrevet av Galvani ikke kan betraktes på grunn av "dyreelektrisitet"; i Galvanis eksperimenter var strømkilden ikke froskens ryggmarg, men en krets dannet av forskjellige metaller - kobber og jern. Som svar på Voltas innvendinger produserte Galvani ny opplevelse, allerede uten deltakelse av metaller. Han viste at hvis huden fjernes fra bakbenene til en frosk, så kuttes isjiasnerven på det punktet hvor røttene går ut
ryggmarg og klargjør nerven langs låret til underbenet, så når nerven kastes på de eksponerte musklene i underbenet, trekker de seg sammen. O. Dubois-Reymond kalte denne opplevelsen "den sanne grunnleggende opplevelsen av nevromuskulær fysiologi." Siden oppfinnelsen på 20-talletår XIX
århundrer galvanometer(multiplikator) og andre elektriske måleinstrumenter, var fysiologer i stand til nøyaktig å måle elektriske strømmer som oppstår i levende vev ved hjelp av spesielle fysiske instrumenter. Med hjelp av animatøren C. Matteuci (1838) først viste: Når et andre nevromuskulært preparat påføres en sammentrekkende nervemuskel, trekker muskelen seg også sammen.
Matteucis erfaring forklares med at aksjonspotensialene som oppstår i muskelen under eksitasjon er sterke nok til å forårsake eksitasjon av nerven festet til den første muskelen, og dette medfører sammentrekning av den andre muskelen. Den mest komplette læren om bioelektriske fenomener i levende vev ble utviklet på 40-50-tallet av forrige århundre av E. Dubois-Reymond. Hans spesielle fortjeneste er den tekniske uklanderligheten til eksperimentene hans. Ved hjelp av galvanometeret, induksjonsapparatet og ikke-polariserende elektroder han forbedret og tilpasset fysiologiens behov, ga Dubois-Reymond ugjendrivelige bevis på tilstedeværelsen av elektriske potensialer i levende vev både i hvile og under eksitasjon. I løpet av den andre halvdelen av 1800-tallet
og på 1900-tallet ble teknologien for registrering av biopotensiale kontinuerlig forbedret. Således ble telefonen på 80-tallet av forrige århundre brukt i elektrofysiologisk forskning av N. E. Vvedensky, kapillærelektrometeret av Lippmann, og strenggalvanometeret av V. Einthoven på begynnelsen av dette århundret. Takket være utviklingen av elektronikk har fysiologi svært avanserte elektriske måleinstrumenter med lav treghet (sløyfeoscilloskop) og til og med praktisk talt treghetsfrie (katodestrålerør). Den nødvendige graden av forbedring av biostrømmer er sikret. AC og DC elektronisk og forsterkere Mikrofysiologiske forskningsteknikker er utviklet , som tillater fjerning av potensialer fra enkelt nerve- og muskelceller og nervefibre. I denne forbindelse er bruken som gjenstand av spesiell betydning studier av gigantiske nervefibre (aksoner) til blekksprutblekkspruten . Diameteren deres når 1 mm, noe som gjør det mulig å sette inn tynne elektroder i fiberen, perfusere den med løsninger av forskjellige sammensetninger og bruke merkede ioner for å studere ionepermeabiliteten til den eksiterbare membranen.
Moderne representasjoner
om mekanismen for forekomst av biopotensialer er i stor grad basert på data innhentet i eksperimenter på slike aksoner. Billett 5. Plasmamembranen og dens rolle i metabolismen mellom cellen og miljøet. Celle (plasma) membran.
1. Membranen består av et dobbelt lag med lipidmolekyler.
De hydrofile, polare delene av molekylene (hodene) er plassert på utsiden av membranen, mens de hydrofobe, ikke-polare delene (haler) er plassert på innsiden.
2. Membranproteiner er mosaisk innebygd i lipid-dobbeltlaget.
Noen av dem passerer gjennom membranen (de kalles integrerte), andre er plassert på den ytre eller indre overflaten av membranen (de kalles perifere).
3. Lipidbasen til membranen har egenskapene til en væske (som flytende olje) og kan endre dens tetthet. Membranens viskositet avhenger av lipidsammensetning og temperatur. I denne forbindelse kan membranproteiner og lipider i seg selv bevege seg fritt langs membranen og inne i den.
4. Membranene til de fleste intracellulære membranorganeller er fundamentalt lik plasmamembranen.:
5. Til tross for den felles strukturen til membranene til alle celler, er sammensetningen av proteiner og lipider i hver celletype og i cellen forskjellig. Sammensetningen av de ytre og indre lipidlagene er også forskjellig. Funksjoner
1) Barriere- gir regulert, selektiv, passiv og aktiv metabolisme med omgivelsene. Selektiv permeabilitet betyr at permeabiliteten til en membran for forskjellige atomer eller molekyler avhenger av størrelse, elektrisk ladning og kjemiske egenskaper. Selektiv permeabilitet sørger for at cellen og cellerommene er separert fra miljøet og tilført de nødvendige stoffene.
2) Transport
- transport av stoffer inn og ut av cellen skjer gjennom membranen. Transport gjennom membraner gir:
levering av næringsstoffer
fjerning av sluttprodukter av metabolisme
utskillelse av ulike stoffer
skape ionegradienter opprettholde optimale pH- og ionekonsentrasjoner i cellen, som er nødvendige for funksjonen til cellulære enzymer
4)3) Matrise- sikrer en viss relativ posisjon og orientering av membranproteiner, deres optimale interaksjon.
Mekanisk- sikrer cellens autonomi, dens intracellulære strukturer, samt forbindelse med andre celler (i vev).
6)Cellevegger spiller en stor rolle for å sikre mekanisk funksjon, og hos dyr den intercellulære substansen.- noen proteiner som ligger i membranen er reseptorer (molekyler ved hjelp av hvilke cellen oppfatter visse signaler).
7)Enzymatisk- membranproteiner er ofte enzymer.
8)Genererer og leder biopotensialer. Ved hjelp av membranen opprettholdes en konstant konsentrasjon av ioner i cellen: konsentrasjonen av K + ion inne i cellen er mye høyere enn utenfor, og konsentrasjonen av Na + er mye lavere, noe som er veldig viktig, siden dette sikrer opprettholdelse av potensialforskjellen på membranen og generering av en nerveimpuls.
9) Cellemerking- det er antigener på membranen som fungerer som markører - "merker" som gjør at cellen kan identifiseres. Dette er glykoproteiner (det vil si proteiner med forgrenede oligosakkaridsidekjeder festet til dem) som spiller rollen som "antenner". Ved hjelp av markører kan celler gjenkjenne andre celler og handle i samspill med dem, for eksempel i dannelsen av organer og vev. Dette tillater også
immunsystem
gjenkjenne fremmede antigener. Billett 6. Membran teori om eksitasjon.
Passiv transport av stoffer over en membran. Kalium-natrium pumpe. Membraneksitasjonsteori - i fysiologi - kommer fra ideen om at når en levende celle (nerve, muskel) er irritert, endres permeabiliteten til overflatemembranen, noe som fører til fremveksten av transmembrane ioniske strømmer. Konsentrasjonsgradient
- dette er vektor fysisk mengde
, som karakteriserer størrelsen og retningen til den største endringen i konsentrasjonen av et stoff i miljøet. Hvis vi for eksempel tar for oss to regioner med forskjellige konsentrasjoner av et stoff, atskilt med en semipermeabel membran, vil konsentrasjonsgradienten bli rettet fra regionen med lavere konsentrasjon av stoffet til regionen med høyere konsentrasjon. Passiv transport. Under driften overføres tre positive Na+-ioner fra cellen for hver to positive K-ioner inn i cellen. Dette arbeidet er ledsaget av akkumulering av forskjell på membranen elektriske potensialer. Samtidig brytes ATP ned, og gir energi. fungerer etter prinsippet om en peristaltisk pumpe.
Billett 7. Mekanismen for forekomsten av membranpotensial og dets endringer under påvirkning av ulike faktorer.
Normalt, når en nervecelle er i fysiologisk hvile og klar til å jobbe, har den allerede opplevd en omfordeling av elektriske ladninger mellom den indre og ytre siden av membranen. På grunn av dette oppsto et elektrisk felt, og et elektrisk potensial dukket opp på membranen - hvilemembranpotensial.
Hvilepotensial- dette er forskjellen i elektriske potensialer som er tilstede på den indre og ytre siden av membranen når cellen er i en tilstand av fysiologisk hvile. (cellen er utenfor +, og inni -.). Hemmeligheten bak utseendet til negativitet i en celle: først bytter den ut "sin" natrium med "fremmed" kalium (ja, noen positive ioner for andre, like positive så lekker disse "utvekslede" positive kaliumionene ut av den). , sammen med hvilke positive ladninger strømmer ut av cellen. Det viktige her er det utveksling av natrium mot kalium - ulik . For hver celle gitt tre natriumioner hun får alt
to kaliumioner .
Dette resulterer i tap av én positiv ladning med hver ionebyttehendelse. Så allerede på dette stadiet, på grunn av ulik utveksling, mister cellen flere "plusser" enn den mottar i retur. skape en forskjell mellom ute og inne.
Neste kommer Konsentrasjonspotensialet er en del av hvilepotensialet som skapes av mangelen på positive ladninger inne i cellen, dannet på grunn av lekkasje av positive kaliumioner fra den. Billett 8. Aksjonspotensial. Mekanismen for dens forekomst. Handlingspotensial - en eksitasjonsbølge som beveger seg langs membranen til en levende celle under overføring av et nervesignal. I hovedsak representerer det elektrisk utladning - rask kortsiktig endring i potensial over et lite område av membranen eksiterbar celle fysisk grunnlag nerve- eller muskelimpuls.
Billett 9. Eksitasjonsbølger, deres komponenter.
Hvis på levende vev handle med en stimulans av tilstrekkelig styrke og varighet, så oppstår det eksitasjon i den, som manifesterer seg i endringer i den elektriske tilstanden til membranen. Settet med suksessive endringer i den elektriske tilstanden til membranen kalles en eksitasjonsbølge. For første gang ble en eksitasjonsbølge registrert av K. Cole, H. Curtis (1938-1939), som satte inn én elektrode i prosessen nervecelle blekksprut, og den andre ble plassert i sjøvann, hvor skuddet ble nedsenket. Etter å ha koblet elektrodene med riktig utstyr, registrerte de først MF, og under stimulering, en eksitasjonsbølge.
Komponentene i eksitasjonsbølgen er:
Terskelpotensial;
Handlingspotensial - AP;
Spor potensialer. Årsaken til eksitasjonsbølgen er en endring i den ioniske permeabiliteten til membranen. Når den utsettes for et irriterende middel, permeabilitet cellemembran for Na+-økninger diffunderer natriumioner inn i cellen. I samsvar med reduksjonen i den elektropositive ladningen på yttersiden av membranen, avtar den elektronegative ladningen på innsiden av membranen. Depolarisering av membranen oppstår - en nedgang i MP. I det første øyeblikket skjer depolarisering sakte, MP reduseres bare med 15-25 Go. Den første depolariseringen kalles den lokale (lokale) responsen. Depolarisering fortsetter og når kritiske ( terskelnivå til null og endrer fortegn til det motsatte. Toppen av aksjonspotensialet inntreffer i perioden da membranen lades opp - potensialreversering. Den ytre siden av membranen lades negativt, den indre - positivt. Etter dette begynner repolariseringsfasen - gjenoppretting av det opprinnelige polarisasjonsnivået. Permeabiliteten til membranen for Na+-ioner avtar, og for K+-ioner øker den. K+ ioner diffunderer fra cellen til den ytre overflaten av membranen, og lader den positivt. I perioden når membranpermeabiliteten for K+ avtar under repolarisering, og repolarisering skjer langsommere enn i den synkende delen av J-toppen, observeres hypopolarisering av membranen (negativt sporpotensial).
Den opprinnelige MP-verdien gjenopprettes. Etter dette, i mange celler, i noen tid, observeres økt permeabilitet av membranen til K+, i forbindelse med dette begynner MP å vokse - hyperpolarisering av membranen oppstår (et positivt sporpotensiale oppstår som genererer J, cellen). mottar hver gang en viss mengde Na+ og mister K+. Konsentrasjonen av ioner i cellen og det intercellulære stoffet utjevnes imidlertid ikke, noe som skyldes virkningen av natrium-kalium-pumpen, som fjerner Na+ fra cellen og slipper K+ inn i cellen.
Billett 10. Absolutte og relative ildfaste faser.
Under eksitasjonsprosessen endres vevseksitabilitet. Det er perioder med eksitabilitet:
1. Innledende økning i eksitabilitet. Observert under lokale (lokale) svar.
2. Refraktær - midlertidig reduksjon i vevseksitabilitet. Det er faser:
Absolutt ildfast - fullstendig inexcitability i løpet av vekstperioden C kan ikke forårsake spenning i denne fasen, selv om stimulansen virker over terskelstyrken.
Relativ refraktæritet - redusert eksitabilitet i perioden med redusert AP for å forårsake eksitasjon er det nødvendig å handle med en stimulus av overterskelstyrke.
2. Supernormal - økt eksitabilitet, eksitasjon kan være forårsaket av en veldig svak stimulans av underterskelstyrke.
Oppfyller spor negativt potensial.
3. Subnormal - redusert eksitabilitet sammenlignet med dets opprinnelige nivå. Sammenfaller med det positive sporpotensialet. Deretter gjenopprettes det innledende nivået av eksitabilitet. Billett 11. Begrepet labilitet, eller funksjonell mobilitet Labilitet (funksjonell mobilitet) er en egenskap per tidsenhet. Labilitet karakteriserer også hastigheten på oppstart og opphør av nerveprosessen.
Frekvensen av forekomst av elementære sykluser av eksitasjon i nerve- og muskelvev.
Konseptet ble introdusert av den russiske fysiologen N. E. Vvedensky, som anså målet på L. for å være den høyeste frekvensen av vevsirritasjon gjengitt av det uten å konvertere rytmen. L. gjenspeiler tiden hvor vevet gjenoppretter sin ytelse etter neste eksitasjonssyklus.
De største L. skiller seg ut Axon s , i stand til å reprodusere opptil 500-1000 pulser per 1 sek; mindre labil Synapser(for eksempel motor nerveenden kan ikke overføre mer enn 100-150 eksitasjoner per 1 sek).
L. er en variabel verdi. Således, i hjertet, under påvirkning av hyppige irritasjoner, øker L Dette fenomenet ligger til grunn for den såkalte. mestre rytme. Læren om L. er viktig for å forstå mekanismene for nervøs aktivitet, nervesentres og analysatorers arbeid, både normalt og ved ulike smertefulle abnormiteter.
Billett 12. Oppsummering og dens typer.
Oppsummering- interaksjon av synoptiske prosesser (eksitatorisk og hemmende) på nevronmembranen eller muskelcelle, preget av en økning i effekten av irritasjon til en refleksreaksjon. Fenomenet S. as karakteristisk egenskap
nervesentre ble først beskrevet av I. M. Sechenov i 1868.:
På systemnivå skilles det mellom summering
Romlig
Midlertidig Romlig S.
oppdages ved samtidig handling av flere. romlig adskilte afferente stimuli, som hver er ineffektiv for forskjellige reseptorer i samme mottakelige sone. Midlertidig S. består av interaksjon nervøse påvirkninger, kommer med en viss intervall til de samme eksitable strukturene langs de samme nervekanalene. På
cellenivå
et slikt skille mellom typene S. er ikke berettiget, derfor kalles det.
romlig-tidsmessig. S. er en av mekanismene for å implementere koordinering.
To typer interneuronforbindelser bør skilles:
1) lokal – synaptisk
2) "diffus, ikke-synaptisk", utført gjennom påvirkning på omkringliggende celler av nevroaktive stoffer som sirkulerer i de intercellulære rommene.
De har en modulerende effekt på elektrogenese og mange vitale prosesser i nerveceller.
Sherrington kalte de eksisterende interneuronforbindelsene synapser. bremsing Synapse strukturell utdanning , hvor overgangen av en nervefiber til en annen skjer, eller overgangen av en nerve til en nevron og en muskel. Den synaptiske delen av aksonet er preget av en opphopning av små runde legemer - synaptiske vesikler (vesikler) med en diameter på 10 til 20 nm. Disse vesiklene inneholder et spesifikt stoff som frigjøres når aksonet er opphisset og kalles formidler. Enden på et akson med vesikler kalles presynaptisk membran . Området til en nerve, nevron eller muskel som overføringen overføres direkte til eksitasjon ringte postsynaptisk membran. Mellom disse to strukturene er det et lite gap (ikke mer enn 50 nm), som kalles synaptisk spalte. Så hvem som helst).
synapse består av tre deler: presynaptisk membran, synaptisk spalte og postsynaptisk membran
Av ovenstående følger det at i synapser utføres overføringen av eksitasjon
kjemisk
og dette skjer gjennom tre prosesser:
stoffer. 1) frigjøring av mediatoren fra boblene; 2) diffusjon av senderen inn i synaptisk spalte 3) forbindelsen til denne mediatoren med spesifikke reaktive strukturer i den postsynaptiske membranen, noe som fører til dannelsen av en ny impuls.
Irritanter- dette er alle levende tings eiendom å reagere på
, en maksimum og mange underterskel, suprathreshold og supramaximal. ytre påvirkninger
For eksempel en endring i metabolsk hastighet, en endring i hvilemembranpotensialet, en endring i ionegradienten, etc. endringer i struktur og funksjoner. Alle celler og vev er irritable.
– Dette er miljøfaktorer som kan forårsake respons i en levende formasjon.
fysisk (mekanisk, termisk, stråling, lydstimulering)
kjemisk (syrer, alkalier, giftstoffer, medisinske stoffer)
biologiske (virus, ulike mikroorganismer)
Interne stimuli inkluderer stoffer dannet i kroppen selv (hormoner, biologisk aktive stoffer).
I henhold til deres biologiske betydning er stimuli delt inn i tilstrekkelig og utilstrekkelig. Tilstrekkelige stimuli inkluderer de som virker under naturlige forhold på eksitable systemer, for eksempel: lys for synsorganet; lyd for hørselsorganet; lukt for luktesansen.
Upassende tid. For å forårsake spenning må en utilstrekkelig respons være mange ganger sterkere enn en adekvat respons for persepsjonsapparatet. Eksitasjon er et sett med fysiske og kjemiske prosesser i vev.
7. Hvilepotensial, handlingspotensial. Lokal respons.
Hvilepotensial.
Når en celle eller fiber er i ro, er det indre potensial(membranpotensial) varierer fra -50 til -90 millivolt og er konvensjonelt tatt som null. Tilstedeværelsen av dette potensialet skyldes ulikheten i konsentrasjonene av Na +, K +, Cl -, Ca 2+ ioner i og utenfor cellen, samt ulik permeabilitet av membraner for disse ionene. Det er 30-50 ganger mer kalium inne i cellen enn utenfor. Samtidig er permeabiliteten til membranen til en ueksitert celle for kaliumioner 25 ganger høyere enn for natriumioner. Derfor forlater kalium cellen til utsiden. På dette tidspunktet passerer anionene i cellens cytoplasma, spesielt de ytre anionene, mindre godt gjennom membranen og er konsentrert på overflaten, og skaper et "-"-potensial. Kaliumioner frigjort fra cellen holdes på den ytre overflaten av membranen av en elektrostatisk motsatt ladning.
Denne potensialforskjellen kalles membranpotensialet eller hvilepotensialet. Over tid, i en slik situasjon, kan flertallet av kaliumioner forlate cellen og forskjellen i konsentrasjonene deres ute og inne vil jevne seg ut, men dette skjer ikke, fordi det er en natrium-kalium-pumpe i cellen. På grunn av hvilket kalium strømmer tilbake fra vevsvæsken inn i cellen og natriumioner frigjøres mot konsentrasjonsgradienten (og det er mer natrium utenfor cellen)
Handlingspotensial
Hvis en nerve eller muskelfiber påvirkes av en endring, endres membranens permeabilitet umiddelbart. Den øker for natriumioner, siden konsentrasjonen av natrium i vevsvæsken er høyere, suser ionene inn i syren og reduserer den til null membranpotensial. I noen tid vises en potensiell forskjell med motsatt fortegn (membranpotensialreversering).
a) depolariseringsfase
b) repolariseringsfase
c) spor repolariseringsfase (potensial)
Endringen i membranpermeabilitet for Na+ varer ikke lenge. Den begynner å øke for K+ og avta for Na+. Dette tilsvarer repolariseringsfasen. Den synkende delen av kurven tilsvarer sporpotensialet og reflekterer gjenopprettingsprosessene som skjer etter irritasjon.
Amplituden og arten av midlertidige endringer i aksjonspotensialet (AP) avhenger lite av styrken til handlingen. Det er viktig at denne kraften har en viss kritisk verdi, som kalles irritasjon eller rheobase. Etter å ha oppstått på stedet for irritasjon, sprer aksjonspotensialet seg langs nerven eller muskelfiberen uten å endre amplituden. Tilstedeværelsen av en terskel for irritasjon og uavhengigheten av amplituden til handlingspotensialet fra styrken til stimulansen kalles "alt" eller "ingen"-loven. I tillegg til styrken til irritasjonen, er varigheten av dens virkning også viktig. For kort handlingstid fører ikke til opphisselse. Det er vanskelig å bestemme metodisk. Derfor introduserte forskeren Lapin begrepet "chronopsia". Dette er minimumstiden som kreves for å forårsake vevseksitasjon ved en kraft lik to reobaser.
Forekomsten av et aksjonspotensial innledes ved stimuleringspunktet av en muskel eller nerve som er aktiv under terskelendringer i membranpotensialet. De vises i skjemaet lokale(lokal) svare.
En lokal respons er preget av:
avhengighet av styrken av irritasjon
gradvis øke omfanget av responsen.
ikke-forplantning langs nervefiberen.
De første tegnene på en lokal respons oppdages når stimuli er 50-70 % av terskelverdien. Den lokale responsen, i likhet med aksjonspotensialet, skyldes en økning i natriumpermeabilitet. Denne økningen var imidlertid ikke tilstrekkelig til å utløse et handlingspotensial.
Aksjonspotensialet oppstår når membrandepolarisering når kritisk nivå. Men den lokale responsen er viktig. Den forbereder vevet for påfølgende eksponeringer.
Leder eksitasjon langs nerve- og muskelfibre. Fasenaturen til endringer i nervefibrenes eksitabilitet.
Gjennomføring av eksitasjon
Eksitasjon sprer seg langs nerve- og muskelfibre på grunn av dannelsen av aksjonspotensialer og lokale elektriske strømmer i dem. Hvis det oppstår et aksjonspotensial i noen del av nervefiberen på grunn av virkningen av en handling, vil membranen i denne delen bli ladet "+". Det tilstøtende uspennede området er “–”.
En lokal strøm oppstår, som depolariserer membranen og fremmer fremveksten av et aksjonspotensial i dette området. At. eksitasjon forplanter seg langs fiberen.
Under naturlige forhold forplanter eksitasjon seg langs fiberen i form av intermitterende pulser en viss frekvens. Dette skyldes det faktum at etter hver impuls blir nervefiberen uopphetet i en kort periode. Endringer i eksitabilitet studeres ved å bruke 2 stimuli som virker med et visst intervall.
Følgende endringer i eksitabilitet er etablert.
Tegning Under en lokal respons øker eksitabiliteten. Under depolariseringsfasen noteres fullstendig ikke-eksitabilitet av nerven. Dette er den såkalte absolutte ildfaste fasen. Varigheten av denne fasen for nervefibre er 0,2-0,4 ml, for muskler 2,5-4 ml. Dette etterfølges av en fase med relativ ildfasthet. Det tilsvarer repolariseringsfasen.
Nerve- og muskelfiber reagerer med spenning på sterke irritasjoner. Fasen varer lenger enn den relative refraksjonsfasen. og er 1,2 ml.
I samme vev varierer varigheten av refraktæritet, spesielt ved funksjonelle forstyrrelser i NS eller under sykdommen.
I løpet av sporpotensialfasen utvikler opphøyelsesfasen eller den supernormale fasen, det vil si at det oppstår en sterk respons på handlinger av noe slag. Sist for nervefibre 12-30 ml, i muskler 50 ml eller mer.
Stimulus er en ekstern eller intern faktor i forhold til den eksitable strukturen i miljøet, som, når den handler eller endrer handlingen, er i stand til å forårsake eksitasjon.
Naturligvis snakker vi om å definere begrepet stimulus i sammenheng med fysiologien til eksitable vev.
La meg minne deg på at strukturen kan reagere på virkningen av et irritasjonsmiddel (stimulus) med irritasjon (en uspesifikk reaksjon) og eksitasjon (en spesifikk elektrisk reaksjon). Eksitasjon oppstår når de tilsvarende lovene for irritasjon er oppfylt. For en irritasjonsreaksjon i de samme eksitable strukturene, er oppfyllelsen av lovene vi vurderer i dag slett ikke nødvendig.
Bare eksitable vev, deres komponenter og organer som består av dem kan reagere på irritasjon med eksitasjon. For eksempel muskelfiber, muskelvev, muskel (organ). La meg minne deg på det eksitable vev inkluderer nervøs, muskulær og kjertel.
I økende grad, i stedet for begrepet "irriterende" brukes begrepet "stimulus". Dette er synonymer. Og i fremtiden vil vi bruke begrepet stimulus veldig ofte. Men husk! I fysiologien til eksitable vev er det begrepet eksitasjon, men det er ikke noe begrep om patogen. Eksitasjon oppstår som respons på virkningen av et irritasjonsmiddel (stimulus).
Så, ifølge definisjonen, kan en stimulus være en faktor som ikke tidligere har virket på den eksitable strukturen. For eksempel rørte en nabo hånden din. Hvis du følte dette, oppsto det spenning i visse eksitable strukturer.
Et annet eksempel. I reseptorer som kontrollerer gasssammensetning blod, spenning oppstår når oksygenkonsentrasjonen endres eller karbondioksid i blodet.
Kan opphisselse oppstå uten ytre stimulans? Ja, som et resultat av spontan depolarisering av cellen. Disse prosessene er karakteristiske for pacemakerceller i hjertemuskelen og mage-tarmkanalen.
Typer av stimuli
Tegn som skiller irriterende stoffer:
1. Natur (modalitet, valens): fysisk, kjemisk, etc.
2. Biologisk betydning (tilstrekkelig, utilstrekkelig)
3. Forholdet mellom påvirkningskraften og eksitasjonsterskelen (underterskel, terskel, superterskel).
4. Enkelt eller seriell
Av natur er stimuli delt inn i kjemiske, mekaniske, strålende, temperaturer, elektriske osv. I dette tilfellet snakker de om modaliteten til stimulus.
Stimuli av samme modalitet varierer i valens. For eksempel kan kjemiske (modalitet) stimuli være salt, søtt, bittert, surt (valens) Begrepet modalitet brukes oftere i felten sensorisk fysiologi angående reseptorer og analysatorer generelt. Og når de snakker om modaliteten til en stimulus, mener de naturen til sensasjonene forårsaket av stimulansen. Men la oss ikke glemme at reseptorer, og analysatorer generelt, er eksitable strukturer.
Innenfor hver modalitet kan stimulansens valens skilles ut. For eksempel kan et kjemisk irritasjonsmiddel være en syre, en alkali eller et salt.
Ved biologisk betydning Uavhengig av modalitet deles stimuli inn i tilstrekkelig og utilstrekkelig.
Tilstrekkelig stimuli er i stand til å forårsake en eksitasjonsreaksjon når de utsettes for visse eksiterbare strukturer.
Med andre ord kan en stimulus, som virker på forskjellige biologiske strukturer, forårsake eksitasjon bare i noen av dem. For disse strukturene vil denne stimulansen være tilstrekkelig. For eksempel forårsaker virkningen av lys bare stimulering i visse strukturer i netthinnen. Det er tilstrekkelig for dem.
Det er ikke nødvendig, når man snakker om tilstrekkelig stimuli, å begrense seg innenfor rammen av "naturlige forhold" og å identifisere begrepene "naturlig stimulus" og "adekvat stimulus." For eksempel forårsaker effekten av matkjemikalier på smaksløkene opphisselse. Kjemikalier mat, selvfølgelig, i dette tilfellet er både naturlige og tilstrekkelig irriterende. Men hvis vi er med laboratorieforhold la oss handle på de samme reseptorene elektrisk støt, kan også agitasjon forekomme. I dette tilfellet vil stimulansen ikke være naturlig, men vil være tilstrekkelig for de aktuelle reseptorene.
La oss sitere en annen definisjon av tilstrekkelig stimuli. "Tilstrekkelige stimuli er de som virker under naturlige forhold på strengt definerte reseptorer og begeistrer dem [++484+ p238]." Du bør forstå hvorfor definisjonen som er gitt er mildt sagt upresis.
Upassende stimuli Når de utsettes for visse eksitable strukturer, er de i stand til å forårsake en eksitasjonsreaksjon, men dette krever betydelig større energiforbruk enn når eksitasjon av de samme strukturene fra en adekvat stimulus.
For eksempel er synlig lys for reseptorene i netthinnen eller lyd i området for dets oppfatning for reseptorene til den auditive analysatoren en tilstrekkelig stimulans. Imidlertid kan følelsen av et lysglimt (fosfen, "gnister fra øynene") eller en hørbar lyd (ringing i ørene) oppstå når de utsettes for mekaniske (et slag mot hodet) og andre stimuli med tilstrekkelig kraft. I i dette tilfellet Eksitasjon forekommer også i henholdsvis de visuelle eller auditive analysatorene, men under påvirkning av utilstrekkelige stimuli som ikke er typiske for dem.
Stimulans tilstrekkelighet manifesteres i det faktum at terskelstyrken er betydelig lavere sammenlignet med terskelstyrken til en utilstrekkelig stimulus. For eksempel oppstår følelsen av lys i en person når minimumsintensiteten til lysstimulusen bare er 10 -17 - 10 -18 W, og mer enn den mekaniske. 10 -4 W, dvs. forskjellen mellom lys og mekaniske terskelstimuli for menneskelige øyereseptorer når 13-14 størrelsesordener.
La meg understreke nok en gang at utilstrekkelig stimuli også kan forårsake begeistring. Når vi snakker om utilstrekkelige stimuli for enhver eksitabel struktur, mener vi at det er tilstrekkelige stimuli for samme struktur.
Kan stimuli av samme modalitet, men med forskjellig valens, være forskjellig i deres tilstrekkelighet til den eksitable strukturen? Ja, det kan de. For eksempel er slike kjemiske (modalitets-) stimuli som sukker, salt (valens) tilstrekkelige for forskjellige smaksreseptorer på tungen.
Basert på forholdet mellom styrken til stimulansen og eksitasjonsterskelen, skilles subterskel, terskel og overterskel. Mer om dette den viktigste egenskapen Vi vil snakke om irritanten senere, og analysere "styrkeloven" av irritasjon.
Stimuli kan være enkelt eller seriell.
Enkeltstimuli variere i styrke, varighet, form, økningshastighet og reduksjon i styrke (gradient) (fig. 809141947).

Ris. 809141947. Forskjeller i parameterne for enkeltstimuli (stimuli): a - etter styrke, b - etter varighet, c - etter økningshastigheten i styrke (gradient), d - etter form (den første er rektangulær, de to neste er trapesformet).
Serieirriterende stoffer variere i frekvens, meander (mønster, mønster) (fig.).

Ris. . Forskjellen i parametrene til serielle stimuli (stimuli): A - i frekvens, B - i forholdet mellom varigheten av stimulansen og varigheten av pausen (duty factor), C - i arten og rekkefølgen til pulsene ( meander).
Vær oppmerksom på at alle de ovennevnte egenskapene gjelder for stimuli av enhver modalitet.
Oppmerksomhet! Slike insentiver, som studenter ofte fremstiller, kan ikke eksistere.
I. E. Wolperts teknikk, utviklet i vårt laboratorium, er blottet for ulempene ved Lenz sin teknikk, siden innholdet i drømmen ikke er foreslått. Det er fysiologisk mer nøyaktig enn Klein-teknikken, siden en streng dosering av ekstern stimulans utføres når det gjelder styrke og varighet. I tillegg er studiene våre ledsaget av objektiv registrering av prosessen med hypnotisk søvn ved å bruke de ovennevnte elektrofysiologiske teknikkene. Vår største fordel fremfor amerikansk arbeid er at vi eksperimenterer på grunnlag av . Dette er en betydelig teoretisk fordel.
I. E. Volpert brukte metoden for brøkanalyse av foreslåtte drømmer i hypnose. Under hypnotisk søvn forteller hypnotisøren til motivet "du drømmer" og produserer samtidig en slags irritasjon. Etter 2 min. legen vekker emnet og spør om drømmen. Observanden rapporterer om en drøm hun nettopp hadde. Det fortsetter igjen. Etter en tid foreslås søvn igjen med påføring av irritasjon. Etter 2 min. motivet blir vekket, og hun forteller drømmen hun så under den andre søvnperioden. Det samme gjøres en tredje gang. Noen tidligere trente individer får en slags irritasjon, men forslaget "du drømmer" gis ikke. Etter slutten av den hypnotiske økten blir personen avhørt angående alle hennes opplevelser under hypnotisk søvn.
Denne metoden for drømmeforskning representerer en ytterligere eksperimentell forbedring av metoden for foreslåtte drømmer i hypnose. Som et eksempel gir vi den beskrevne studien.
I dette eksemplet kan man se hvordan irritasjonen produsert av forskeren (i dette tilfellet kutan-proprioseptiv) kommer inn i innholdet i drømmen, som består av en kombinasjon av elementer av irritasjon og elementer fra fortiden livserfaring. Det er ingenting uforståelig fra synspunktet om årsaksanalyse i disse drømmene.
Under hypnotisk søvn og under naturlig søvn er det således et samspill mellom tilgjengelige stimuli og nervespor. tidligere irritasjoner under utviklingen av drømmer. Samtidig har de stor verdi individuelle egenskaper og type nervesystemet(som vil bli omtalt nærmere i avsnitt XII). I denne forbindelse er Pavlovs doktrine om analysatorer av stor betydning for den fysiologiske forståelsen av drømmer. Rollen til individuelle kortikale analysatorer er forskjellig i forskjellige personer. Dermed har artister en mer utviklet visuell analysator, mens musikere har en mer utviklet auditiv. Denne fysiologiske forskjellen gjenspeiles i drømmene deres. Noen nevrotikere (spesielt hysterikere) har ofte luktdrømmer. Pasient G. hadde således økt luktesans og opplevde ofte luktdrømmer. Hun sa om seg selv at hun «levde i lydens og luktenes rike hele livet».
Det som presenteres i denne delen fører oss til følgende konklusjoner. Ekstern og indre stimuli, som virker under søvn, spiller rollen som den første impulsen i utplasseringen av kjeden av desinhibering av nervespor. I dette tilfellet er en mekanisme for summering av langtidsvirkende irritasjon som fører til desinhibering av spor mulig.
Effekten av ytre og indre irritasjoner under søvn kommer ned til følgende alternativer:
1) til en generell desinhibering av søvn og utseendet til grunne faser av søvn, som er assosiert med utvikling av drømmer på grunn av reproduksjon av nervespor; i dette tilfellet forårsaker eksisterende irritasjoner desinhibering av søvn, men forårsaker ikke direkte drømmer;
2) til desinhibering og fremveksten av en drøm med deltakelse av denne analysatoren; i dette tilfellet forårsaker eksisterende stimuli desinhibering, forårsaker en drøm og går inn i innholdet;
3) til desinhibering og fremveksten av en drøm på grunn av en annen analysator eller andre analysatorer; i dette tilfellet forårsaker eksisterende irritasjoner desinhibering, forårsaker en drøm, men er ikke en del av innholdet;
4) i drømmer kan det oppstå en forvrengning av makt ytre stimuli basert på mønsteret av den paradoksale hypnotiske fasen.*
Alt det ovennevnte belyser bare én side av drømmenes fysiologi. Den andre siden er desinhibering av nervespor uten deltakelse av eksisterende stimuli.
* OM nervøs mekanisme Vi vil diskutere drømmer basert på Pavlovske hypnotiske faser videre, i avsnitt VIII.
 "En kveld med gåter basert på verkene til S.
"En kveld med gåter basert på verkene til S. Om spørsmålet om tidlig bestått Unified State-eksamen Tidlig sertifisering av Unified State-eksamenen
Om spørsmålet om tidlig bestått Unified State-eksamen Tidlig sertifisering av Unified State-eksamenen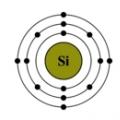 Silisium i naturen (25,8 % i jordskorpen)
Silisium i naturen (25,8 % i jordskorpen)