โปรตีน: ประวัติการวิจัย องค์ประกอบทางเคมี คุณสมบัติ หน้าที่ทางชีวภาพ โปรตีนประกอบด้วยองค์ประกอบอะไรบ้างและมีคุณสมบัติอะไรบ้าง? มีอะไรอยู่ในโปรตีน
องค์ประกอบทางเคมีของโปรตีน
3.1. พันธะเปปไทด์
โปรตีนเป็นโพลีเมอร์ที่ผิดปกติซึ่งสร้างขึ้นจากกากกรดอะมิโน ซึ่งเป็นสูตรทั่วไปในสารละลายที่เป็นน้ำที่ค่า pH ใกล้เคียงกับค่าเป็นกลางสามารถเขียนเป็น NH 3 + CHRCOO - . สารตกค้างของกรดอะมิโนในโปรตีนเชื่อมต่อกันด้วยพันธะเอไมด์ระหว่างหมู่-อะมิโนและ-คาร์บอกซิล พันธะเปปไทด์ระหว่าง สอง-สารตกค้างของกรดอะมิโนมักถูกเรียกว่า พันธะเปปไทด์ และพอลิเมอร์ที่สร้างขึ้นจากสารตกค้างของกรด α-amino ที่เชื่อมต่อกันด้วยพันธะเปปไทด์เรียกว่า โพลีเปปไทด์ โปรตีนในฐานะโครงสร้างที่มีนัยสำคัญทางชีววิทยาสามารถเป็นได้ทั้งโพลีเปปไทด์เดี่ยวหรือโพลีเปปไทด์หลายตัวที่ก่อรูปสารเชิงซ้อนเดียวอันเป็นผลมาจากการกระทำระหว่างกันที่ไม่ใช่โควาเลนต์
3.2. องค์ประกอบองค์ประกอบของโปรตีน
จากการศึกษาองค์ประกอบทางเคมีของโปรตีนจำเป็นต้องค้นหาก่อนอื่นว่าองค์ประกอบทางเคมีประกอบด้วยอะไรและประการที่สองโครงสร้างของโมโนเมอร์ เพื่อตอบคำถามแรก กำหนดองค์ประกอบเชิงปริมาณและคุณภาพขององค์ประกอบทางเคมีของโปรตีน การวิเคราะห์ทางเคมีแสดงให้เห็น มีอยู่ในโปรตีนทั้งหมด คาร์บอน (50-55%), ออกซิเจน (21-23%), ไนโตรเจน (15-17%), ไฮโดรเจน (6-7%), กำมะถัน (0.3-2.5%) นอกจากนี้ยังพบฟอสฟอรัส ไอโอดีน เหล็ก ทองแดง และมาโครและองค์ประกอบขนาดเล็กอื่นๆ ในองค์ประกอบของโปรตีนแต่ละชนิดในปริมาณที่น้อยมาก
เนื้อหาขององค์ประกอบทางเคมีหลักในโปรตีนสามารถเปลี่ยนแปลงได้ ยกเว้นไนโตรเจน ความเข้มข้นที่มีลักษณะเฉพาะคือความคงตัวมากที่สุดและเฉลี่ย 16% นอกจากนี้ ปริมาณไนโตรเจนในสารอินทรีย์อื่นๆ ยังต่ำอีกด้วย ตามนี้ เสนอให้กำหนดปริมาณโปรตีนโดยไนโตรเจนที่เป็นส่วนประกอบ เมื่อรู้ว่าไนโตรเจน 1 กรัมบรรจุอยู่ในโปรตีน 6.25 กรัม ปริมาณไนโตรเจนที่พบจะถูกคูณด้วยค่า 6.25 และได้รับปริมาณโปรตีน
เพื่อตรวจสอบลักษณะทางเคมีของโปรตีนโมโนเมอร์ จำเป็นต้องแก้ปัญหาสองประการ: แยกโปรตีนออกเป็นโมโนเมอร์และค้นหาองค์ประกอบทางเคมีของโปรตีน การสลายโปรตีนออกเป็นส่วนๆ ทำได้โดยการไฮโดรไลซิส - การเดือดของโปรตีนเป็นเวลานานด้วยกรดแร่ที่แรง (กรดไฮโดรไลซิส)หรือบริเวณ (ไฮโดรไลซิสอัลคาไลน์). มักใช้ต้มที่อุณหภูมิ 110 C กับ HCl เป็นเวลา 24 ชั่วโมง ในขั้นตอนต่อไป สารที่ประกอบเป็นไฮโดรไลเสตจะถูกแยกออก เพื่อจุดประสงค์นี้มีการใช้วิธีการต่าง ๆ ซึ่งส่วนใหญ่มักใช้โครมาโตกราฟี (สำหรับรายละเอียดเพิ่มเติมโปรดดูบทที่ "วิธีการวิจัย ... ") กรดอะมิโนเป็นส่วนหลักของไฮโดรไลเสตที่แยกจากกัน
3.3. กรดอะมิโน
ปัจจุบันพบกรดอะมิโนที่แตกต่างกันถึง 200 ชนิดในวัตถุต่างๆ ของสัตว์ป่า ตัวอย่างเช่นในร่างกายมนุษย์มีประมาณ 60 ตัว อย่างไรก็ตามโปรตีนมีกรดอะมิโนเพียง 20 ชนิดซึ่งบางครั้งเรียกว่ากรดจากธรรมชาติ
กรดอะมิโนเป็นกรดอินทรีย์ที่อะตอมไฮโดรเจนของอะตอม -คาร์บอนถูกแทนที่ด้วยหมู่อะมิโน - NH 2 ดังนั้นโดยธรรมชาติทางเคมี เหล่านี้คือกรดอะมิโนที่มีสูตรทั่วไป:
H - C - NH 2
จากสูตรนี้จะเห็นได้ว่าองค์ประกอบของกรดอะมิโนทั้งหมดประกอบด้วยหมู่ทั่วไปดังต่อไปนี้: - CH 2, - NH 2, - COOH. โซ่ด้านข้าง (อนุมูล - R) กรดอะมิโนต่างกัน ดังที่เห็นได้จากภาคผนวก 1 ลักษณะทางเคมีของอนุมูลมีความหลากหลาย ตั้งแต่อะตอมไฮโดรเจนไปจนถึงสารประกอบไซคลิก เป็นอนุมูลที่กำหนดคุณสมบัติโครงสร้างและหน้าที่ของกรดอะมิโน
กรดอะมิโนทั้งหมด ยกเว้น glycine อะมิโนอะซิติกที่ง่ายที่สุดสำหรับคุณ (NH 3 + CH 2 COO ) มีไครัลอะตอม C และสามารถมีอยู่ในรูปของอีแนนทิโอเมอร์สองตัว (ไอโซเมอร์ออปติคัล):
ซีโอโอ-ซีโอโอ-
NH3+ RR NH3+
หลี่-ไอโซเมอร์ดี-ไอโซเมอร์
โปรตีนที่ศึกษาทั้งหมดในปัจจุบันมีเพียงกรดอะมิโนของซีรีย์ L ซึ่งหากเราพิจารณาอะตอม chiral จากด้านข้างของอะตอม H กลุ่ม NH 3 + , COO และ R Radical จะเรียงตามเข็มนาฬิกา ความจำเป็นในการสร้างโมเลกุลพอลิเมอร์ที่มีนัยสำคัญทางชีวภาพจากอีแนนชิโอเมอร์ที่กำหนดไว้อย่างเข้มงวดนั้นชัดเจน - จากส่วนผสมของราซีมิกของอีแนนชิโอเมอร์สองตัว จะได้ส่วนผสมที่ซับซ้อนอย่างคาดไม่ถึงของไดแอสเทอริโอไอโซเมอร์ คำถามที่ว่าทำไมสิ่งมีชีวิตบนโลกขึ้นอยู่กับโปรตีนที่สร้างขึ้นอย่างแม่นยำจาก L- และไม่ใช่กรดอะมิโน D- ยังคงเป็นปริศนาที่น่าสนใจ ควรสังเกตว่ากรด D-amino นั้นค่อนข้างแพร่หลายในธรรมชาติและยิ่งกว่านั้นยังเป็นส่วนหนึ่งของ oligopeptides ที่มีความสำคัญทางชีวภาพ
โปรตีนถูกสร้างขึ้นจากกรด -อะมิโนพื้นฐาน 20 ชนิด อย่างไรก็ตาม ส่วนที่เหลือ ซึ่งเป็นกรดอะมิโนที่มีความหลากหลายค่อนข้างมาก ก่อตัวขึ้นจากกรดอะมิโน 20 ชนิดที่มีอยู่แล้วในองค์ประกอบของโมเลกุลโปรตีน ในบรรดาการเปลี่ยนแปลงเหล่านี้ อันดับแรกควรสังเกตการก่อตัว สะพานซัลไฟด์ ในระหว่างการออกซิเดชันของซิสเทอีนสองตัวที่ตกค้างในองค์ประกอบของโซ่เปปไทด์ที่เกิดขึ้นแล้ว เป็นผลให้สารตกค้างของกรดไดอะมิโนไดคาร์บอกซิลิกเกิดขึ้นจากซิสเทอีนเรซิดิวสองตัว ซีสทีน (ดูภาคผนวก I) ในกรณีนี้ การเชื่อมขวางเกิดขึ้นภายในสายโพลีเปปไทด์หนึ่งสายหรือระหว่างสายโซ่ที่ต่างกันสองสาย ในฐานะที่เป็นโปรตีนขนาดเล็กที่มีสายพอลิเปปไทด์สองสายเชื่อมต่อกันด้วยไดซัลไฟด์บริดจ์ เช่นเดียวกับการเชื่อมขวางภายในหนึ่งในสายโซ่โพลีเปปไทด์:
GIVEQCCCASVCSLYQLENYCN
FVNQHLCGSHLVEALYLVCGERGFFYTPKA
ตัวอย่างที่สำคัญของการดัดแปลงเรซิดิวกรดอะมิโนคือการแปลงโพรลีนเรซิดิวไปเป็นเรซิดิว ไฮดรอกซีโพรลีน :
N-CH-CO-N-CH-CO-
CH 2 CH 2 CH 2 CH 2
CH2CHOH
การเปลี่ยนแปลงนี้เกิดขึ้นและในระดับที่มีนัยสำคัญ ระหว่างการก่อตัวขององค์ประกอบโปรตีนที่สำคัญของเนื้อเยื่อเกี่ยวพัน - คอลลาเจน .
การดัดแปลงโปรตีนที่สำคัญมากอีกประเภทหนึ่งคือ ฟอสโฟรีเลชั่นของกลุ่มไฮดรอกโซของสารตกค้างของซีรีน ทรีโอนีน และไทโรซีน ตัวอย่างเช่น
– NH – CH – CO – – NH – CH – CO –
CH 2 โอ้ CH 2 OPO 3 2 –
กรดอะมิโนในสารละลายในน้ำอยู่ในสถานะแตกตัวเป็นไอออนเนื่องจากการแตกตัวของหมู่อะมิโนและคาร์บอกซิลที่ประกอบขึ้นเป็นอนุมูล กล่าวอีกนัยหนึ่ง พวกมันเป็นสารประกอบแอมโฟเทอริกและสามารถมีอยู่ได้ทั้งในรูปกรด (ผู้ให้โปรตอน) หรือในฐานะที่เป็นเบส (ตัวรับผู้บริจาค)
กรดอะมิโนทั้งหมดขึ้นอยู่กับโครงสร้างแบ่งออกเป็นหลายกลุ่ม:
อะไซคลิก. กรดอะมิโนโมโนอะมิโนโมโนคาร์บอกซิลิกมีเอมีนหนึ่งกลุ่มและกลุ่มคาร์บอกซิลหนึ่งกลุ่มในสารละลายที่เป็นน้ำพวกมันเป็นกลาง บางส่วนมีลักษณะโครงสร้างทั่วไปซึ่งช่วยให้พิจารณาร่วมกันได้:
ไกลซีนและอะลานีน Glycine (glycocol หรือ aminoacetic acid) ไม่ใช้งานทางสายตา - เป็นกรดอะมิโนเพียงชนิดเดียวที่ไม่มีอีแนนทิโอเมอร์ Glycine มีส่วนร่วมในการก่อตัวของนิวคลีอิกและน้ำดีถึง - t, heme เป็นสิ่งจำเป็นสำหรับการวางตัวเป็นกลางของผลิตภัณฑ์ที่เป็นพิษในตับ ร่างกายใช้อะลานีนในกระบวนการเผาผลาญคาร์โบไฮเดรตและพลังงานต่างๆ ไอโซเมอร์ -อะลานีนของมันเป็นส่วนสำคัญของวิตามิน pantothenic สำหรับคุณ โคเอ็นไซม์ A (CoA) ซึ่งเป็นสารสกัดจากกล้ามเนื้อ
ซีรีนและทรีโอนีนพวกมันอยู่ในกลุ่มของกรดไฮดรอกซีเพราะ มีหมู่ไฮดรอกซิล ซีรีนเป็นส่วนหนึ่งของเอ็นไซม์ต่าง ๆ ซึ่งเป็นโปรตีนหลักของนม - เคซีน เช่นเดียวกับไลโปโปรตีนหลายชนิด ธรีโอนีนมีส่วนร่วมในการสังเคราะห์โปรตีน ซึ่งเป็นกรดอะมิโนที่จำเป็น
ซิสเทอีนและเมไทโอนีนกรดอะมิโนที่มีอะตอมกำมะถัน ค่าของ cysteine ถูกกำหนดโดยการปรากฏตัวของกลุ่ม sulfhydryl (-SH) ในองค์ประกอบของมันซึ่งทำให้สามารถออกซิไดซ์ได้ง่ายและปกป้องร่างกายจากสารที่มีความสามารถในการออกซิไดซ์สูง (ในกรณีที่ได้รับบาดเจ็บจากรังสีฟอสฟอรัส พิษ) เมไทโอนีนมีลักษณะเด่นคือมีหมู่เมทิลเคลื่อนที่ได้ง่าย ซึ่งใช้สำหรับการสังเคราะห์สารประกอบสำคัญในร่างกาย (โคลีน ครีเอทีน ไทมีน อะดรีนาลีน เป็นต้น)
วาลีน ลิวซีน และไอโซลิวซีนพวกมันเป็นกรดอะมิโนที่แตกแขนงซึ่งเกี่ยวข้องอย่างแข็งขันในการเผาผลาญและไม่ได้สังเคราะห์ในร่างกาย
กรดอะมิโนโมโนอะมิโนไดคาร์บอกซิลิกมีหมู่อะมิโนหนึ่งหมู่และหมู่คาร์บอกซิลสองหมู่ และให้ปฏิกิริยาที่เป็นกรดในสารละลายที่เป็นน้ำ เหล่านี้รวมถึงแอสปาร์ติกและกลูตามีนสำหรับคุณ แอสพาราจีนและกลูตามีน พวกมันเป็นส่วนหนึ่งของตัวกลางไกล่เกลี่ยของระบบประสาท
กรดอะมิโนไดอะมิโนโมโนคาร์บอกซิลิกในสารละลายที่เป็นน้ำจะมีปฏิกิริยาเป็นด่างเนื่องจากการมีอยู่ของหมู่เอมีนสองกลุ่ม ที่เกี่ยวข้องกับพวกเขา ไลซีนเป็นสิ่งจำเป็นสำหรับการสังเคราะห์ฮิสโตนและในเอนไซม์จำนวนหนึ่ง อาร์จินีนมีส่วนร่วมในการสังเคราะห์ยูเรียครีเอทีน
วัฏจักร. กรดอะมิโนเหล่านี้มีนิวเคลียสอะโรมาติกหรือเฮเทอโรไซคลิกในองค์ประกอบและตามกฎแล้วจะไม่ถูกสังเคราะห์ในร่างกายมนุษย์และต้องได้รับอาหาร พวกเขามีส่วนร่วมอย่างแข็งขันในกระบวนการเผาผลาญที่หลากหลาย ดังนั้น
ฟีนิลอะลานีนเป็นแหล่งหลักของการสังเคราะห์ไทโรซีน ซึ่งเป็นสารตั้งต้นของสารสำคัญทางชีววิทยาจำนวนหนึ่ง ได้แก่ ฮอร์โมน (ไทรอกซีน อะดรีนาลีน) เม็ดสีบางชนิด ทริปโตเฟนนอกจากจะมีส่วนร่วมในการสังเคราะห์โปรตีนแล้วยังเป็นส่วนประกอบของวิตามินพีพี เซโรโทนิน ทริปตามีน และเม็ดสีอีกจำนวนหนึ่ง ฮิสทิดีนจำเป็นสำหรับการสังเคราะห์โปรตีน ซึ่งเป็นสารตั้งต้นของฮีสตามีน ซึ่งส่งผลต่อความดันโลหิตและการหลั่งน้ำย่อย
คุณสมบัติ
โปรตีนเป็นสารประกอบโมเลกุลขนาดใหญ่ เหล่านี้เป็นพอลิเมอร์ที่ประกอบด้วยโมโนเมอร์นับร้อยนับพันของกรดอะมิโน
โปรตีนมีน้ำหนักโมเลกุลสูง บางชนิดสามารถละลายได้ในน้ำ สามารถบวมได้ มีลักษณะเฉพาะด้วยกิจกรรมทางแสง การเคลื่อนที่ในสนามไฟฟ้า และคุณสมบัติอื่นๆ บางอย่าง
โปรตีนมีส่วนร่วมอย่างแข็งขันในปฏิกิริยาเคมี คุณสมบัตินี้เกิดจากการที่กรดอะมิโนที่ประกอบเป็นโปรตีนมีกลุ่มการทำงานที่แตกต่างกันซึ่งสามารถทำปฏิกิริยากับสารอื่นได้ เป็นสิ่งสำคัญที่อันตรกิริยาดังกล่าวจะเกิดขึ้นภายในโมเลกุลโปรตีน ส่งผลให้เกิดการก่อตัวของเปปไทด์ ไฮโดรเจนซัลไฟด์ และพันธะประเภทอื่นๆ เพื่ออนุมูลกรดอะมิโนและตามลำดับและ มวลโมเลกุลโปรตีนอยู่ในช่วง 10,000 - 1,000,000 ดังนั้น ribonuclease (เอ็นไซม์ที่ทำลาย RNA) มีกรดอะมิโนตกค้าง 124 ตัว และมีน้ำหนักโมเลกุลประมาณ 14,000 Myoglobin (โปรตีนของกล้ามเนื้อ) ประกอบด้วย 153 กรดอะมิโนตกค้าง มีโมเลกุล น้ำหนัก 17,000 และเฮโมโกลบิน - 64,500 (574 กรดอะมิโนตกค้าง) น้ำหนักโมเลกุลของโปรตีนอื่น ๆ สูงกว่า: -โกลบูลิน (รูปแบบแอนติบอดี) ประกอบด้วยกรดอะมิโน 1250 และมีน้ำหนักโมเลกุลประมาณ 150,000 และน้ำหนักโมเลกุลของเอนไซม์กลูตาเมตดีไฮโดรจีเนสเกิน 1,000,000
การกำหนดน้ำหนักโมเลกุลดำเนินการด้วยวิธีการต่างๆ เช่น ออสโมเมตริก การกรองเจล ออปติคัล ฯลฯ อย่างไรก็ตาม วิธีการตกตะกอนที่เสนอโดย T. Svedberg ที่แม่นยำที่สุดคือ โดยอาศัยข้อเท็จจริงที่ว่าในระหว่างการปั่นแยกเหวี่ยงด้วยความเข้มข้นสูงด้วยความเร่งสูงถึง 900,000 กรัม อัตราการตกตะกอนของโปรตีนจะขึ้นอยู่กับน้ำหนักโมเลกุลของโปรตีน
คุณสมบัติที่สำคัญที่สุดของโปรตีนคือความสามารถในการแสดงทั้งความเป็นกรดและเบส กล่าวคือทำหน้าที่เป็น แอมโฟเทอริกอิเล็กโทรไลต์ สิ่งนี้ทำให้มั่นใจได้โดยกลุ่มที่แยกตัวออกจากกันที่ประกอบขึ้นเป็นอนุมูลของกรดอะมิโน ตัวอย่างเช่น คุณสมบัติของกรดจะถูกส่งไปยังโปรตีนโดยกลุ่มคาร์บอกซิลของกรดอะมิโนแอสปาร์ติกกลูตามิก ในขณะที่คุณสมบัติอัลคาไลน์นั้นมาจากอาร์จินีน ไลซีน และอนุมูลฮิสทิดีน ยิ่งโปรตีนมีกรดอะมิโนไดคาร์บอกซิลิกมากเท่าไหร่ คุณสมบัติที่เป็นกรดของกรดก็จะยิ่งแรงขึ้นและในทางกลับกัน
กลุ่มเหล่านี้ยังมีประจุไฟฟ้าที่สร้างประจุโดยรวมของโมเลกุลโปรตีน ในโปรตีนที่กรดอะมิโนแอสปาร์ติกและกลูตามีนมีอิทธิพลเหนือ ประจุของโปรตีนจะเป็นลบ กรดอะมิโนพื้นฐานที่มากเกินไปจะให้ประจุบวกกับโมเลกุลโปรตีน เป็นผลให้ในสนามไฟฟ้า โปรตีนจะเคลื่อนที่ไปทางแคโทดหรือขั้วบวก ขึ้นอยู่กับขนาดของประจุทั้งหมด ดังนั้นในสภาพแวดล้อมที่เป็นด่าง (pH 7 - 14) โปรตีนจะบริจาคโปรตอนและกลายเป็นประจุลบ ในขณะที่ในสภาพแวดล้อมที่เป็นกรด (pH 1 - 7) การแตกตัวของกลุ่มกรดจะถูกระงับและโปรตีนจะกลายเป็นไอออนบวก
ดังนั้น ปัจจัยที่กำหนดพฤติกรรมของโปรตีนเป็นไอออนบวกหรือประจุลบคือปฏิกิริยาของตัวกลาง ซึ่งพิจารณาจากความเข้มข้นของไฮโดรเจนไอออนและแสดงโดยค่า pH อย่างไรก็ตาม ที่ค่า pH บางค่า จำนวนประจุบวกและประจุลบจะเท่ากัน และโมเลกุลจะกลายเป็นกลางทางไฟฟ้า กล่าวคือ มันจะไม่เคลื่อนที่ในสนามไฟฟ้า ค่า pH ของตัวกลางนี้ถูกกำหนดให้เป็นจุดไอโซอิเล็กทริกของโปรตีน ในกรณีนี้ โปรตีนอยู่ในสถานะที่เสถียรน้อยที่สุด และเมื่อ pH ของกรดหรือด่างเปลี่ยนแปลงไปเล็กน้อย โปรตีนก็จะตกตะกอนได้ง่าย สำหรับโปรตีนธรรมชาติส่วนใหญ่ จุดไอโซอิเล็กทริกอยู่ในสภาพแวดล้อมที่เป็นกรดเล็กน้อย (pH 4.8 - 5.4) ซึ่งบ่งชี้ถึงความเด่นของกรดอะมิโนไดคาร์บอกซิลิกในองค์ประกอบ
คุณสมบัติ amphoteric รองรับคุณสมบัติบัฟเฟอร์ของโปรตีนและการมีส่วนร่วมในการควบคุมค่า pH ของเลือด ค่า pH ของเลือดมนุษย์จะคงที่และอยู่ในช่วง 7.36 - 7.4 แม้ว่าจะมีสารต่างๆ ที่มีลักษณะเป็นกรดหรือเป็นด่าง ซึ่งให้อาหารเป็นประจำหรือเกิดขึ้นในกระบวนการเมตาบอลิซึม ดังนั้นจึงมีกลไกพิเศษในการควบคุมกรด-เบส ความสมดุลของสภาพแวดล้อมภายในร่างกาย ระบบดังกล่าวรวมถึงระบบที่พิจารณาในบทที่ ระบบบัฟเฟอร์เฮโมโกลบิน “การจำแนกประเภท” (หน้า 28) การเปลี่ยนแปลงค่า pH ของเลือดมากกว่า 0.07 บ่งชี้ถึงการพัฒนาของกระบวนการทางพยาธิวิทยา การเปลี่ยนแปลงของค่า pH ไปทางด้านกรดเรียกว่าภาวะกรด (acidosis) และด้านที่เป็นด่างเรียกว่าอัลคาโลซิส (alkalosis)
สิ่งที่สำคัญอย่างยิ่งสำหรับร่างกายคือความสามารถของโปรตีนในการดูดซับสารและไอออนบางชนิด (ฮอร์โมน วิตามิน เหล็ก ทองแดง) ซึ่งละลายได้ไม่ดีในน้ำหรือเป็นพิษ (บิลิรูบิน กรดไขมันอิสระ) โปรตีนขนส่งพวกมันผ่านทางเลือดไปยังสถานที่ที่มีการเปลี่ยนแปลงเพิ่มเติมหรือทำให้เป็นกลาง
สารละลายโปรตีนในน้ำมีลักษณะเฉพาะของตัวเอง ประการแรก โปรตีนมีความสัมพันธ์กับน้ำสูง กล่าวคือ พวกเขา ชอบน้ำซึ่งหมายความว่าโมเลกุลโปรตีน เช่น อนุภาคที่มีประจุ จะดึงดูดไดโพลของน้ำ ซึ่งอยู่รอบๆ โมเลกุลโปรตีนและก่อตัวเป็นน้ำหรือเปลือกไฮเดรต เปลือกนี้ปกป้องโมเลกุลโปรตีนจากการเกาะติดกันและตกตะกอน ขนาดของเปลือกน้ำขึ้นอยู่กับโครงสร้างของโปรตีน ตัวอย่างเช่น อัลบูมินจับกับโมเลกุลของน้ำได้ง่ายขึ้นและมีเปลือกน้ำที่ค่อนข้างใหญ่ ในขณะที่โกลบูลิน ไฟบริโนเจนจับกับน้ำได้แย่กว่า และเปลือกไฮเดรชั่นมีขนาดเล็กกว่า ดังนั้น ความคงตัวของสารละลายในน้ำของโปรตีนจึงถูกกำหนดโดยปัจจัยสองประการ: การปรากฏตัวของประจุบนโมเลกุลโปรตีนและเปลือกน้ำที่อยู่รอบๆ เมื่อปัจจัยเหล่านี้ถูกกำจัดออกไป โปรตีนจะตกตะกอน กระบวนการนี้สามารถย้อนกลับและย้อนกลับไม่ได้
การตกตะกอนของโปรตีนแบบย้อนกลับได้(การทำเกลือออก) เกี่ยวข้องกับการตกตะกอนของโปรตีนภายใต้อิทธิพลของสารบางชนิด หลังจากที่กำจัดออกซึ่งโปรตีนจะกลับสู่สถานะเดิม (ดั้งเดิม) เกลือของโลหะอัลคาไลและอัลคาไลน์เอิร์ ธ ใช้สำหรับทำเกลือโปรตีน (โซเดียมและแอมโมเนียมซัลเฟตมักใช้ในทางปฏิบัติ) เกลือเหล่านี้เอาเปลือกน้ำออก (ทำให้เกิดการคายน้ำ) และเอาประจุออก มีความสัมพันธ์โดยตรงระหว่างขนาดของเปลือกน้ำของโมเลกุลโปรตีนและความเข้มข้นของเกลือ คือ ยิ่งเปลือกน้ำมีขนาดเล็กเท่าใด เกลือก็จะยิ่งน้อยลงเท่านั้น ดังนั้นโกลบูลินซึ่งมีโมเลกุลขนาดใหญ่และหนักและเปลือกน้ำขนาดเล็กจะตกตะกอนเมื่อสารละลายอิ่มตัวด้วยเกลือและอัลบูมินอย่างไม่สมบูรณ์ เนื่องจากโมเลกุลขนาดเล็กที่ล้อมรอบด้วยเปลือกน้ำขนาดใหญ่ จะตกตะกอนเมื่อสารละลายอิ่มตัวอย่างสมบูรณ์
โมเลกุลโปรตีนพื้นเมือง
โมเลกุลโปรตีนที่แปลงสภาพ เส้นประบ่งบอกถึงพันธะในโมเลกุลโปรตีนพื้นเมืองที่แตกตัวระหว่างการเสียสภาพ


การตกตะกอนที่ไม่สามารถย้อนกลับได้นั้นสัมพันธ์กับการเปลี่ยนแปลงโครงสร้างโมเลกุลลึกในโครงสร้างของโปรตีน ซึ่งนำไปสู่การสูญเสียคุณสมบัติดั้งเดิมของพวกมัน (ความสามารถในการละลาย กิจกรรมทางชีวภาพ ฯลฯ) โปรตีนดังกล่าวเรียกว่า denature และกระบวนการ การทำให้เสียสภาพ. การสลายตัวของโปรตีนเกิดขึ้นในกระเพาะอาหารซึ่งมีสภาพแวดล้อมที่เป็นกรดอย่างแรง (pH 0.5 - 1.5) และสิ่งนี้มีส่วนช่วยในการสลายโปรตีนด้วยเอนไซม์สลายโปรตีน การเปลี่ยนสภาพของโปรตีนเป็นพื้นฐานสำหรับการรักษาพิษของโลหะหนัก เมื่อผู้ป่วยได้รับนมหรือไข่ดิบต่อ os ("ทางปาก") เพื่อให้โลหะแปลงสภาพโปรตีนของนมหรือไข่
พวกเขาถูกดูดซับบนพื้นผิวของพวกเขาและไม่ได้ทำหน้าที่เกี่ยวกับโปรตีนของเยื่อเมือกของกระเพาะอาหารและลำไส้และไม่ถูกดูดซึมเข้าสู่กระแสเลือด
ขนาดของโมเลกุลโปรตีนอยู่ในช่วง 1 µm ถึง 1 nm ดังนั้นจึงเท่ากับ คอลลอยด์อนุภาคที่สร้างสารละลายคอลลอยด์ในน้ำ สารละลายเหล่านี้มีลักษณะเฉพาะด้วยความหนืดสูง ความสามารถในการกระจายรังสีแสงที่มองเห็นได้ และไม่ผ่านเมมเบรนแบบกึ่งซึมผ่านได้
ความหนืดของสารละลายขึ้นอยู่กับน้ำหนักโมเลกุลและความเข้มข้นของตัวถูกละลาย ยิ่งน้ำหนักโมเลกุลสูง สารละลายก็จะยิ่งมีความหนืดมากขึ้น โปรตีนเป็นสารประกอบโมเลกุลขนาดใหญ่ทำให้เกิดสารละลายหนืด ตัวอย่างเช่น สารละลายไข่ขาวในน้ำ
น้ำ

อนุภาคคอลลอยด์ไม่ผ่านเยื่อใยสังเคราะห์ (กระดาษแก้ว ฟิล์มคอลลอยด์) เนื่องจากรูพรุนของพวกมันมีขนาดเล็กกว่าอนุภาคคอลลอยด์ เยื่อหุ้มชีวภาพทั้งหมดไม่สามารถซึมผ่านโปรตีนได้ คุณสมบัติของสารละลายโปรตีนนี้ใช้กันอย่างแพร่หลายในยาและเคมีเพื่อทำให้การเตรียมโปรตีนบริสุทธิ์จากสิ่งเจือปน กระบวนการแยกนี้เรียกว่าการฟอกไต ปรากฏการณ์ของการฟอกไตรองรับการทำงานของอุปกรณ์ "ไตเทียม" ซึ่งใช้กันอย่างแพร่หลายในการแพทย์เพื่อรักษาภาวะไตวายเฉียบพลัน
การฟอกไต (วงกลมสีขาวขนาดใหญ่คือโมเลกุลโปรตีน วงกลมสีดำคือโมเลกุลของโซเดียมคลอไรด์)
แร่ธาตุน้ำนม
เถ้านมมีแร่ธาตุเช่นแคลเซียม ฟอสฟอรัส แมกนีเซียม โพแทสเซียม โซเดียม คลอรีน กำมะถัน ซิลิคอน ปริมาณขององค์ประกอบแต่ละอย่างในนมนั้นพิจารณาจากปัจจัยทางพันธุกรรมเป็นหลัก การให้อาหารและปัจจัยแวดล้อมอื่น ๆ มีอิทธิพลเพียงเล็กน้อยต่อการบำรุงรักษา ปริมาณแร่ธาตุในนมจะคงที่แม้ว่าจะมีองค์ประกอบเพียงเล็กน้อยในอาหารก็ตาม ด้วยการบริโภคแร่ธาตุที่ไม่เพียงพอกับอาหาร สำรองของร่างกายจะถูกระดมและทำให้ความเข้มข้นในนมยังคงอยู่ในระดับหนึ่ง ด้วยการขาดธาตุอย่างน้อยหนึ่งธาตุอย่างมีนัยสำคัญ ปริมาณแร่ธาตุต่อหน่วยปริมาตรของนมยังคงไม่เปลี่ยนแปลงมากหรือน้อย อย่างไรก็ตาม ผลผลิตน้ำนมและปริมาณแร่ธาตุทั้งหมดในนมลดลง
|
แร่ธาตุ |
ประกอบด้วย g |
แร่ธาตุ |
|
ปริมาณรวมของธาตุในนมน้อยกว่า 0.15% เนื้อหาของธาตุในนมจะขึ้นอยู่กับการปรากฏตัวของพวกมันในอาหารสัตว์อย่างใกล้ชิด
สมบัติทางโครงสร้างและทางกลของน้ำมัน
ตาม Rebinder มีโครงสร้างหลักสองประเภท
ชนิดแรกเป็นโครงสร้างการแข็งตัวของเลือด- เป็นโครงข่ายเชิงพื้นที่ที่เกิดขึ้นจากการยึดเกาะแบบสุ่มของอนุภาคที่เล็กที่สุดของเฟสที่กระจายตัวหรือไมโครโมเลกุลผ่านชั้นบางๆ ของตัวกลางที่กำหนด
ประเภทที่สองคือโครงสร้างการตกผลึกและการควบแน่นซึ่งเกิดขึ้นจากการผสมผสานระหว่างผลึกโดยตรงกับการก่อตัวของของแข็งคริสตัลไลน์
ฐานไขมันของมาการีนอยู่ในโครงสร้างประเภทการแข็งตัวของเลือด ความสม่ำเสมอและคุณสมบัติของพลาสติกของฐานไขมันของมาการีนนั้นพิจารณาจากอัตราส่วนของเฟสของแข็งและของเหลวในไขมันที่บริโภคได้โดยเฉพาะ อัตราส่วนของเฟสของแข็งและของเหลวนี้เป็นเรื่องปกติสำหรับสภาวะการตกผลึกเฉพาะบางอย่าง (อุณหภูมิ เวลา การผสม) ในกรณีนี้ องค์ประกอบของตัวกลางต่อเนื่องและระยะกระจัดกระจาย และลักษณะของตำแหน่งของเฟสที่กระจัดกระจายในตัวกลางของเหลวแบบต่อเนื่องมีความสำคัญอย่างยิ่ง
สำหรับไขมันที่บริโภคได้บางชนิดที่อุณหภูมิหนึ่งและสภาวะการตกผลึก ปริมาณของเฟสที่กระจายตัวของของแข็งอาจเกินอัตราส่วนเฟสที่เหมาะสม จากนั้นฟิล์มบางๆ ของตัวกลางที่เป็นของเหลวแบบต่อเนื่องจะก่อตัวขึ้นบนพื้นผิวของผลึกที่ไม่สามารถ รบกวนการรวมตัวของผลึกที่วุ่นวายขนาดใหญ่เข้าด้วยกัน ในกรณีนี้ เราจะมีความแข็งมากที่สุดของฐานไขมัน ความคงตัวของคราบสกปรก และคุณสมบัติของพลาสติกที่แย่ที่สุดเสมอ
หากที่อุณหภูมิห้อง ฟิล์มของตัวกลางที่เป็นของเหลวแบบต่อเนื่องจะมีความหนาที่เหมาะสมที่สุด กล่าวคือ ที่ไม่สร้างเงื่อนไขสำหรับการหลอมเหลวของผลึกระหว่างการเก็บรักษา ภายใต้ผลกระทบทางกลหรือความร้อนในระบบ ในกรณีนี้ เราจะได้รับโครงสร้างการแข็งตัวของเลือดที่เสริมแรงเสมอ ซึ่งกำหนดคุณสมบัติพลาสติกที่ดีที่สุดของฐานไขมัน
เพื่อให้ได้โครงสร้างการแข็งตัวที่เสริมความแข็งแรงด้วยคุณสมบัติของพลาสติกที่ดีที่สุด น้ำมันหมูสองประเภทที่มีจุดหลอมเหลว 32°C และ 42°C มักจะถูกนำมาใช้ในสูตรฐานไขมันในต่างประเทศ ในกรณีนี้มีการแนะนำน้ำมันพืชเหลวจำนวนมากพอสมควร ในแง่หนึ่งนี้จะสร้างอัตราส่วนที่ดีที่สุดของสถานะของแข็งและของเหลวในฐานไขมันโดยให้ความสม่ำเสมอคล้ายกับเนยและในทางกลับกันสร้างเงื่อนไขสำหรับความสม่ำเสมอของความคงตัวของมาการีนในช่วงอุณหภูมิที่ค่อนข้างกว้าง นอกจากนี้ การนำไขละลายสูงเข้าไปในฐานไขมันยังขัดแย้งกับข้อกำหนดของนักสรีรวิทยาสำหรับองค์ประกอบของไขมันในอาหาร
ประการแรกควรสังเกตว่ามีเพียงสารทำให้คงตัวอิมัลซิไฟเออร์ที่มีประสิทธิภาพสูงเท่านั้นที่ทำให้สามารถสร้างเทคโนโลยีที่ทันสมัยในการผลิตมาการีนและรับประกันการผลิตผลิตภัณฑ์ไขมันที่บริโภคได้คุณภาพสูง สารเติมแต่งที่ออกฤทธิ์บนพื้นผิวให้อิมัลชันที่กระจายตัวอย่างละเอียดในพันธะที่แข็งแกร่งระหว่างอนุภาคของเฟสที่กระจายตัวและตัวกลางแบบต่อเนื่อง (ไขมันที่เป็นของแข็งที่อุณหภูมิห้อง) ปัญหาหลักในการผลิตมาการีนคือผลกระทบของสารเติมแต่งที่พื้นผิวต่อคุณสมบัติทางโครงสร้างและทางกลของมาการีน และโดยเฉพาะอย่างยิ่งความสามารถในการละลาย
ชั้นดูดซับของอิมัลซิไฟเออร์ช่วยเพิ่มความเสถียรของอิมัลชัน โดยเฉพาะอย่างยิ่งในกรณีที่ชั้นนี้มีโครงสร้าง ทำให้เกิดฟิล์มของเจลพื้นผิวที่มีความหนืดและความแข็งแรงเพิ่มขึ้นอย่างมาก
คุณสมบัติเหล่านี้มีความสำคัญเป็นพิเศษสำหรับการผลิตมาการีน เนื่องจากผลิตภัณฑ์สำเร็จรูปเป็นอิมัลชันของอนุภาคที่เล็กที่สุดของเฟสของเหลว ซึ่งกระจายอย่างสม่ำเสมอในตัวกลางที่ต่อเนื่องของเฟสของแข็งที่อุณหภูมิห้อง
ที่เกี่ยวข้องอย่างใกล้ชิดกับปัญหาความแรงของอิมัลชันคือคำถามเกี่ยวกับประเภทของอิมัลชันที่เกิดขึ้นจากอิมัลชันที่กำหนด มีความเป็นไปได้ของการก่อตัวของสองประเภท ค่าของอัตราส่วนของปริมาตรเฟสสำหรับอิมัลชันบางประเภทที่เกิดขึ้นนั้นอธิบายได้จากข้อเท็จจริงที่ว่าการรวมตัวและการแยกตัวของอิมัลชันของประเภทหนึ่ง ๆ นั้นเกิดขึ้นอย่างเข้มข้นมากขึ้น ปริมาตรของตัวกลางที่กระจายตัวจะเล็กลงและเฟสที่กระจายตัวก็จะยิ่งใหญ่ขึ้น . หากอิมัลชันให้อิมัลชันที่คงตัวเพียงประเภทเดียว อัตราส่วนของปริมาตรจะหยุดเป็นตัวกำหนดในการกำหนดประเภทของอิมัลชัน การผกผันไม่ได้ขึ้นอยู่กับอัตราส่วนของปริมาตรเฟสเท่านั้น แต่ยังขึ้นกับความเข้มข้นและลักษณะทางเคมีของอิมัลซิไฟเออร์ด้วย
อิมัลซิไฟเออร์ต้องมีคุณสมบัติดังต่อไปนี้:
ลดแรงตึงผิว
- เร็วพอที่จะดูดซับบนเฟสอินเตอร์เฟส ป้องกันการรวมตัวของหยด;
- มีโครงสร้างโมเลกุลเฉพาะแบบมีขั้วและไม่มีขั้ว
- ส่งผลต่อความหนืดของอิมัลชัน
ประสิทธิภาพของอิมัลซิไฟเออร์เป็นคุณสมบัติเฉพาะที่ขึ้นอยู่กับลักษณะของสาร ชนิดของสารที่ทำให้เป็นอิมัลชัน อุณหภูมิ ค่า pH ของตัวกลาง ความเข้มข้น เวลาในการทำให้เป็นอิมัลชัน ฯลฯ
ประสิทธิผลของการกระทำและลักษณะของอิมัลชันกำหนดประเภทของอิมัลชัน
อิมัลซิไฟเออร์ที่ชอบน้ำ ละลายได้ดีกว่าในน้ำมากกว่าในไฮโดรคาร์บอน มีส่วนทำให้เกิดอิมัลชันน้ำมัน-น้ำ และไม่ชอบน้ำ ละลายได้ดีกว่าในไฮโดรคาร์บอน อิมัลชันน้ำ-น้ำมัน อัตราส่วนของขนาดของส่วนขั้วและส่วนที่ไม่ใช่ขั้วของโมเลกุลอิมัลซิไฟเออร์นั้นมีลักษณะเฉพาะด้วยตัวบ่งชี้พิเศษ - ความสมดุลของชอบน้ำและไลโปฟิลิก ถ้า HLB ของอิมัลชันเป็น 3-6 อิมัลชันน้ำกับน้ำมันจะถูกสร้างขึ้นโดยมีค่า HLB เท่ากับ 8-13 อิมัลชันน้ำมันและน้ำจะก่อตัวเป็นส่วนใหญ่
มาการีนเป็นอิมัลชันน้ำในน้ำมันซุปเปอร์คูล ในกรณีนี้ ความเป็นไปได้ของการก่อตัวของอิมัลชันชนิดผสมที่มีความโดดเด่นของอิมัลชันน้ำกับน้ำมันจะไม่ถูกตัดออก
หน้าที่หลักของอิมัลซิไฟเออร์:
การสร้างอิมัลชันที่มีความเสถียรสูง
- การรักษาเสถียรภาพและป้องกันการแยกความชื้นและไขมันในผลิตภัณฑ์สำเร็จรูป
- มั่นใจเสถียรภาพระหว่างการจัดเก็บ
- ให้ความสามารถในการป้องกันการกระเซ็นระหว่างการทอด
- รับรองความเป็นพลาสติก
- สร้างความมั่นใจในการสร้างรูปแบบที่มั่นคงของผลึกขัดแตะในกระบวนการสร้างโครงสร้าง
- รับรองคุณสมบัติการทำงานที่ระบุของผลิตภัณฑ์สำเร็จรูปขึ้นอยู่กับพื้นที่ที่ใช้มาการีน
ในยูเครนเป็นเวลาหลายปีที่อิมัลซิไฟเออร์ที่ผลิตในรัสเซียและของการผลิตของเราเองซึ่งผลิตในการผลิตกึ่งอุตสาหกรรมถูกนำมาใช้ ซึ่งรวมถึงอิมัลซิไฟเออร์:
T-1 เป็นผลิตภัณฑ์ของ glycerolysis ของไขมันในเนื้อหรือน้ำมันหมู
- Т-2 – ผลิตภัณฑ์พอลิเมอไรเซชันของกลีเซอรีน esterified ด้วยกรดสเตียริก
- T-F - ส่วนผสมของอิมัลซิไฟเออร์ T-1 และอาหารฟอสฟาไทด์เข้มข้นในอัตราส่วน 2: 1
- PMD - อาหาร monodiglycerides;
- CE - อิมัลซิไฟเออร์รวม - ส่วนผสมของ PMD และฟอสฟาไทด์เข้มข้นในอัตราส่วน 3: 1
อิมัลซิไฟเออร์หลากหลายชนิดของโรงงาน Nizhny Novgorod - โมโนกลีเซอไรด์กลั่นประเภทต่างๆ ในปัจจุบัน การผลิตชุดอิมัลซิไฟเออร์ชนิดใหม่ที่มีส่วนประกอบของเลซิตินเป็นส่วนประกอบหลักในนิจนีย์ นอฟโกรอด เหล่านี้คือเลซิตินมาตรฐาน เลซิตินแบบแยกส่วน - ฟอสฟาดิทิลโคลีนและฟอสฟาไดอิลเซอรีน เช่นเดียวกับเลซิตินที่ไฮโดรไลซ์
ในช่วงไม่กี่ปีที่ผ่านมา อิมัลซิไฟเออร์ของการดัดแปลงต่าง ๆ ของชุด Dimodan, Palsgaard (ที่สถานประกอบการ Quest บางแห่ง) ได้ถูกนำมาใช้เป็นหลักในยูเครน
ในช่วงเวลาที่ต่างกัน ความได้เปรียบในความต้องการอิมัลซิไฟเออร์ทั้งสองประเภทนี้เปลี่ยนจากชนิดหนึ่งไปอีกชนิดหนึ่ง เรียกได้ว่ามีคุณภาพการแข่งขัน-ราคา
Dimodan PVP (Dimodan HP), Dimodan OT (Dimodan S-T PEL/B), Dimodan СР ขึ้นอยู่กับปริมาณไขมันของเนยเทียมและขอบเขตของมัน สำหรับมาการีนที่มีปริมาณไขมันต่ำกว่า 40% ซึ่งปัจจุบันเป็นที่ต้องการของประชากร นอกจากนี้ (ยกเว้น Dimodan OT หรือ Dimodan СР. หรือ Dimodan LS) เอสเทอร์ของโพลิกลีเซอรอลและกรดริซิโนเลอิก - Grinsted PGPR90 ถูกนำมาใช้
ในการผลิตมาการีนไขมันต่ำโดยเฉพาะที่มีปริมาณไขมัน 25% หรือน้อยกว่านั้นใช้ระบบรักษาเสถียรภาพ - ไฮโดรคอลลอยด์ (แอลจิเนต, เพกติน ฯลฯ )
ควรสังเกตว่าผู้ผลิตให้คำแนะนำเกี่ยวกับการใช้อิมัลซิไฟเออร์ประเภทต่างๆและระบบการทำให้เสถียรขึ้นอยู่กับวัตถุประสงค์ของมาการีน การปฏิบัติตามคำแนะนำเหล่านี้จะช่วยให้คุณได้ผลิตภัณฑ์คุณภาพสูง
โปรตีนของกล้ามเนื้อ
เนื้อสัตว์ปีกมีโปรตีนประมาณ 20-23% โปรตีนจากกล้ามเนื้อสามารถแบ่งออกเป็นสามกลุ่มตามความสามารถในการละลาย: myofibrilar, sarcoplasmic และ stromal proteins
Myofibrillar, หรือ ละลายเกลือได้ กระรอกไม่ละลายในน้ำ แต่ส่วนใหญ่ละลายในสารละลายโซเดียมคลอไรด์ที่มีความเข้มข้นมากกว่า 1% กลุ่มนี้ประกอบด้วยโปรตีนประมาณ 20 ตัวซึ่งประกอบเป็น myofibrils ของกล้ามเนื้อหดตัว โปรตีน Myofibrilar สามารถแบ่งออกเป็นสามกลุ่มขึ้นอยู่กับหน้าที่ของพวกเขา: การหดตัวซึ่งมีหน้าที่ในการหดตัวของกล้ามเนื้อ, การควบคุม, เกี่ยวข้องกับการควบคุมกระบวนการหดตัวและโครงร่างโครงร่าง, การยึด myofibrils และช่วยรักษาความสมบูรณ์ของโครงสร้าง
โปรตีนหดตัวไมโอซินและแอคตินมีอิทธิพลอย่างมากต่อการทำงานของโปรตีนในกล้ามเนื้อ เนื่องจากแอคตินและไมโอซินมีอยู่ในรูปของแอกโตไมโอซินที่ซับซ้อนในกล้ามเนื้อแข็ง การทำงานของไมโอซินจึงเปลี่ยนแปลงไปในผลิตภัณฑ์สัตว์ปีกทั้งแบบอิมัลซิไฟด์และแบบขึ้นรูป คุณสมบัติของผลิตภัณฑ์ยังขึ้นอยู่กับอัตราส่วนรวมของแอคตินและไมโอซิน และอัตราส่วนของไมโอซินและแอคตินในสถานะอิสระ ในทางกลับกัน โปรตีน Sarcoplasmic และโปรตีน stromal ส่งผลต่อคุณสมบัติการทำงานของโปรตีน myofibrillar
โปรตีนซาร์โคพลาสมิกละลายได้ในน้ำหรือในสารละลายที่มีความแรงไอออนิกต่ำ (
โปรตีนสโตรมามักเรียกว่าโปรตีนเนื้อเยื่อเกี่ยวพันทำหน้าที่เป็นโครงที่รองรับโครงสร้างกล้ามเนื้อ คอลลาเจนเป็นโปรตีนหลักในสโตรมา อีลาสตินและเรติคูลินเป็นส่วนเล็กๆ ของสโตรมา โปรตีนทั้งหมดเหล่านี้ไม่ละลายในน้ำและสารละลายน้ำเกลือ ความนุ่มของเนื้อมีแนวโน้มลดลงตามอายุอันเนื่องมาจากการเชื่อมโยงข้ามและการเปลี่ยนแปลงอื่นๆ ของคอลลาเจน
เลือดและเศษส่วน
เลือดครบส่วนถูกใช้เป็นวัตถุดิบหลักในการผลิตไส้กรอก กล้ามเนื้อ อาหารกระป๋อง และผลิตภัณฑ์อาหารอื่น ๆ รวมถึงสารเติมแต่งที่ให้สีดั้งเดิมแก่ผลิตภัณฑ์เมื่อใช้การเตรียมโปรตีน (0.6-1.0%) เพื่อจุดประสงค์เดียวกันจะใช้การเตรียมเฮโมโกลบินหรือส่วนผสมขององค์ประกอบที่เกิดขึ้นหลังจากการให้น้ำ (1: 1)
เมื่อเปรียบเทียบกับวัตถุดิบที่มีโปรตีนประเภทอื่น เลือดครบส่วนนั้นไม่นิยมใช้กันอย่างกว้างขวาง เนื่องจากมีสีและรสชาติเฉพาะที่ปรับเปลี่ยนลักษณะทางประสาทสัมผัสของผลิตภัณฑ์สำเร็จรูป ขณะนี้ การวิจัยกำลังดำเนินการเกี่ยวกับการชี้แจงเลือด อย่างไรก็ตาม ด้วยเหตุผลหลายประการ วิธีการที่เสนอยังไม่พบการใช้งานจริงในอุตสาหกรรม คุณสมบัติการทำงานและเทคโนโลยีของเลือดและเศษส่วน (พลาสมา, ซีรั่ม) ขึ้นอยู่กับองค์ประกอบของโปรตีนเป็นหลัก เลือดครบส่วนประกอบด้วยโปรตีนประมาณ 150 ชนิดที่มีคุณสมบัติทางเคมีกายภาพที่แตกต่างกัน ซึ่งส่วนใหญ่ประกอบด้วยโปรตีนที่เป็นองค์ประกอบ อัลบูมิน โกลบูลิน และไฟบริโนเจน ในเรื่องนี้ บนพื้นฐานของเลือดครบส่วน ขอแนะนำให้เตรียมอิมัลชันสำหรับการแนะนำในสูตรผลิตภัณฑ์จากเนื้อสัตว์ และให้ความเสถียรของระบบเนื้อสัตว์เพิ่มขึ้น คุณค่าทางโภชนาการและผลผลิต การปรับปรุงตัวบ่งชี้ทางประสาทสัมผัสและคุณสมบัติทางโครงสร้างและทางกล
ในการเตรียมโปรตีน ขอแนะนำให้ใช้ถั่วเหลืองไอโซเลตหรือโซเดียมเคซิเนต
ระดับของการนำอิมัลชันที่เตรียมจากเลือดครบส่วนเข้าสู่ระบบเนื้อสัตว์อาจสูงถึง 30-40% โดยน้ำหนักของวัตถุดิบหลัก
โปรตีนในพลาสมาในเลือดมี FCS complex ที่ไม่เหมือนใคร อัลบูมินโต้ตอบกับโปรตีนอื่นๆ ได้ง่าย สามารถเชื่อมโยงกับไขมันและคาร์โบไฮเดรต และมีความสามารถในการจับกับน้ำและทำให้เกิดฟองได้สูง
Globulins เป็นอิมัลซิไฟเออร์ที่ดี
ไฟบริโนเจน - มีความสามารถในการสร้างเจลที่เด่นชัด เปลี่ยนเป็นไฟบรินภายใต้อิทธิพลของปัจจัยหลายประการ (ค่า pH เปลี่ยนไปที่ไอโซพอยท์ การนำไอออน Ca ++ เข้าสู่พลาสมา) และสร้างกรอบเชิงพื้นที่
สารผสม คุณสมบัติเหล่านี้ของไฟบริโนเจนสามารถใช้ในการผลิตโปรตีนที่มีหลายองค์ประกอบรวมถึงพีซี, พื้นผิวคล้ายเจล, ในกระบวนการของการสร้างโครงสร้างรองของอิมัลชันเนื้อในการผลิตไส้กรอกต้ม
โปรตีนในพลาสมาทั้งหมดมีลักษณะการละลายได้ดี และด้วยเหตุนี้ ความสามารถในการจับกับน้ำและการทำให้เป็นอิมัลชันสูง จึงสามารถก่อตัวเป็นเจลเมื่อถูกความร้อน การแนะนำของเกลือแกงมีผลเสียต่อความเสถียรของอิมัลชันตามพลาสมาเลือดที่ pH 7.0 คุณสมบัติที่สำคัญที่สุดของพลาสม่าคือความสามารถในการสร้างเจลระหว่างการอบชุบด้วยความร้อน ความแข็งแรงและระดับความสามารถในการจับกับน้ำขึ้นอยู่กับความเข้มข้นของโปรตีนในระบบ ค่า pH การปรากฏตัวของเกลือ อุณหภูมิและระยะเวลาของ เครื่องทำความร้อน
การนำโปรตีนที่ไม่ใช่พลาสมา (ไข่อัลบูมิน ถั่วเหลืองไอโซเลต โซเดียมเคซิเนต) เข้าสู่พลาสมาช่วยเพิ่มความแข็งแรงของเจลและความสามารถในการดูดซับน้ำและไขมันหลังการให้ความร้อนอย่างมีนัยสำคัญ
องค์ประกอบและคุณสมบัติการทำงานและเทคโนโลยีขึ้นอยู่กับสถานะของเลือดและเงื่อนไขของการประมวลผลเบื้องต้นและดังนั้นพื้นที่การใช้งานอาจแตกต่างกันไป
การจัดระบบข้อมูลที่มีอยู่ในปัจจุบันในการประมวลผลของพีซีช่วยให้เราสามารถประเมินแนวทางที่ทันสมัยในการตระหนักถึงศักยภาพทางชีวภาพและเทคโนโลยีเชิงหน้าที่ของส่วนประกอบโปรตีนของพีซีในการผลิตอาหาร
โครงการให้แนวคิดเกี่ยวกับสถานะวิธีการประมวลผลองค์ประกอบและคุณสมบัติของการเตรียมโปรตีนที่ได้รับบนพื้นฐานของ PA กำหนดพื้นที่ของการใช้งานจริงและมัลติฟังก์ชั่นของวัตถุประสงค์ที่ตั้งใจไว้ของ PA นั้นสะท้อนให้เห็นในรูปแบบ FCS ด้วยวิธีการประมวลผลอย่างใดอย่างหนึ่ง
ควรสังเกตว่าระดับของตัวบ่งชี้ FCS แต่ละตัวที่ระบุในตารางที่ 13 และใช้ในการถอดรหัสสัญลักษณ์ที่ใช้ในโครงการนั้นสัมพันธ์กันเนื่องจากข้อเท็จจริงที่ว่าค่าที่แท้จริงของแต่ละลักษณะขึ้นอยู่กับความเข้มข้นของโปรตีน ค่า pH ใน ระบบและอุณหภูมิของตัวกลาง ความแรงของอิออน และปัจจัยอื่นๆ อีกหลายประการ
การวิเคราะห์รูปแบบการจัดหมวดหมู่แสดงให้เห็นว่าวิธีการใช้เทคโนโลยีพลาสมาวิธีหนึ่งคือการใช้พลาสมาในรูปของเหลวที่เสถียร (เช่นเดียวกับหลังจากการทำให้เย็นลงและการแช่แข็ง) ที่มีปริมาณโปรตีนค่อนข้างต่ำและ FCS ดั้งเดิมที่เก็บรักษาไว้
ในกรณีนี้ โปรตีน PC มีลักษณะเฉพาะด้วย BCC และอิมัลซิฟิเคชั่นในระดับสูง ซึ่งเกิดจากการมีโปรตีนที่ละลายน้ำได้ในตัวซึ่งสามารถก่อตัวเป็นเจลเมื่อถูกความร้อน การผสมผสานของคุณสมบัติเหล่านี้ทำให้สามารถใช้พลาสม่าได้อย่างกว้างขวาง ไม่เพียงแต่เป็นส่วนประกอบที่ปรับสมดุลองค์ประกอบทางเคมีโดยรวมของผลิตภัณฑ์สำเร็จรูปเท่านั้น แต่ยังเป็นสารเติมแต่งการทำงานในการผลิตผลิตภัณฑ์จากเนื้ออิมัลซิไฟเออร์ที่มีความชื้นสูงขั้นสุดท้าย: ไส้กรอกต้ม แฟรงค์เฟิร์ต, ไส้กรอก, ผลิตภัณฑ์กึ่งสำเร็จรูปสับ, อาหารกระป๋องสับละเอียด, ผลิตภัณฑ์แฮม ที่สมเหตุสมผลที่สุดคือการนำพลาสมา 10% มาผสมในสูตรแทนเนื้อวัว 3% หรือเนื้อหมู 2% การแนะนำของ PC 20% แทนน้ำในระหว่างการตัดช่วยให้พารามิเตอร์ทางประสาทสัมผัส โครงสร้างและทางกลดีขึ้น และเพิ่มผลผลิตของผลิตภัณฑ์สำเร็จรูป 0.3-0.5% การใช้พลาสมาเลือดเป็นตัวกลางในการเตรียมโปรตีนให้ความชุ่มชื้น (PC 3-4 ส่วนต่อ 1 ส่วนของการเตรียมโปรตีน) ได้ผลลัพธ์ที่ยอดเยี่ยม
พีซีเป็นสิ่งที่ขาดไม่ได้ในการผลิตอิมัลชันโปรตีนไขมันสารยึดเกาะระบบโปรตีนหลายองค์ประกอบที่มีองค์ประกอบที่กำหนดและคุณสมบัติการทำงานและเทคโนโลยีการเตรียมโปรตีนที่มีโครงสร้าง
ความเข้มข้นของ PA โดยการทำให้แห้ง อัลตร้าฟิลเตรชัน และความเข้มข้นของความเย็น ซึ่งทำให้สามารถเพิ่มปริมาณโปรตีนอย่างมีนัยสำคัญ นำไปสู่การดัดแปลง FCS ของสารเตรียมบางส่วน
การทำแห้งด้วยพลาสม่ามีผลที่มีนัยสำคัญอย่างยิ่งต่อระดับการเปลี่ยนแปลงใน FCS ในขณะที่ PA แบบแห้งที่เข้มข้นภายใต้การกรองด้วยอัลตราฟิลเตรชันมีคุณสมบัติการทำงานที่สูงมาก
เข้มข้นที่ได้จากวิธีการเหล่านี้จะถูกนำมาใช้อย่างประสบความสำเร็จในการผลิตผลิตภัณฑ์จากเนื้อสัตว์พร้อมกับ PC ที่เป็นของเหลว
ผู้เชี่ยวชาญชาวอเมริกันเชื่อว่าพลาสม่าจากเลือดวัวต้องขอบคุณ FCS ที่สามารถทดแทนไข่ขาวได้สำเร็จ
การตกตะกอนการตกตะกอนจากการตกตะกอนซึ่งเป็นการรวมกันของกระบวนการของโครงสร้างเทอร์โมโทรปิกการตกตะกอน (การตกตะกอน) และความเข้มข้นของโปรตีน PC ทำให้สามารถเตรียมการเตรียมที่มีความเข้มข้นของโปรตีนค่อนข้างสูงและ FCS พิเศษซึ่งช่วยให้สามารถใช้ในสูตรสำหรับ กึ่งรมควัน รมควัน อบ ไส้กรอกตับ อาหารกระป๋องหัว และผลิตภัณฑ์กึ่งสำเร็จรูปที่มีความชื้นสุดท้ายจำกัดและความสามารถในการดูดซับไขมันสูง ยากลุ่มนี้ประกอบด้วย: "โปรตีนในพลาสมาตกตะกอน", "โปรตีนตกตะกอนในพลาสมา", มีชีวิต, "ชีสพลาสม่า", พีซีแบบเม็ด
การใช้พลาสมาในเลือดประเภทนี้ในการผลิตเนื้อสัตว์มีจำกัด
โครงสร้างของพลาสมาเลือดโดยการเพิ่มแคลเซียมใหม่ช่วยเพิ่มความเป็นไปได้ในการใช้เทคโนโลยีอย่างมาก การแปลง PC และระบบหลายองค์ประกอบตามรูปแบบเจลทำให้สามารถรับเมทริกซ์โครงสร้างที่เลียนแบบวัตถุทางชีววิทยาตามธรรมชาติในลักษณะองค์ประกอบและคุณสมบัติ สร้างข้อกำหนดเบื้องต้นสำหรับกฎระเบียบของ Federal Customs Service ทำให้มั่นใจได้ว่าการมีส่วนร่วมของ คัดเกรดวัตถุดิบในกระบวนการผลิตและทำให้สามารถเข้าถึงแนวทางการแก้ปัญหาจากตำแหน่งใหม่ ๆ การพัฒนาผลิตภัณฑ์อาหารประเภทใหม่ มีประสิทธิภาพโดยเฉพาะอย่างยิ่งคือการใช้ PC และการเตรียมโปรตีนที่ซับซ้อน (แยกถั่วเหลือง, โซเดียมเคซิเนต, ฯลฯ ) , ความคล้ายคลึงกันของผลิตภัณฑ์จากเนื้อสัตว์
การสุกของเนื้อ
ประเด็น “การสุกของเนื้อ” ยังไม่ได้รับการชี้แจงขั้นสุดท้าย จากการสังเกตของผู้ปฏิบัติงาน เป็นที่ทราบกันดีว่าหลังจากการสิ้นสุดชีวิตของสัตว์ การเปลี่ยนแปลงทางเคมีกายภาพเกิดขึ้นในเนื้อ โดยมีลักษณะแข็งตึง จากนั้นจึงคลาย (ทำให้อ่อนลง) ของเส้นใยกล้ามเนื้อ ส่งผลให้เนื้อสัตว์ได้รับรสชาติและปรุงอาหารได้ง่ายขึ้น คุณค่าทางโภชนาการเพิ่มขึ้น การเปลี่ยนแปลงเหล่านี้ในเนื้อเยื่ออ่อนของซากสัตว์เรียกว่า "การทำให้สุก" ("การสุก") หรือ "การหมักเนื้อ"
เพื่ออธิบายกระบวนการของการสุกของเนื้อ คำสอนของ Meyerhof, Embden, Palladin และ Abdergalden เกี่ยวกับพลวัตและการเผาผลาญของคาร์โบไฮเดรตในกล้ามเนื้อในช่วงชีวิตของสัตว์สมควรได้รับความสนใจอย่างมาก
Meyerhof แสดงให้เห็นว่าไกลโคเจนที่มีอยู่ในกล้ามเนื้อถูกใช้ไปในการสร้างกรดแลคติกในระหว่างการหดตัวของกล้ามเนื้อ ระหว่างพักผ่อน
(ส่วนที่เหลือ) กล้ามเนื้อเนื่องจากการจัดหาออกซิเจนไกลโคเจนถูกสังเคราะห์อีกครั้งจากกรดแลคติค
ลุนด์สกราดแสดงให้เห็นว่ากรดครีเอติโนฟอสฟอริกอยู่ในเซลล์กล้ามเนื้อ และเมื่อลดขนาดลง จะถูกแยกออกเป็นกรดครีเอทีนและกรดฟอสฟอริก (ตาม
พัลลาดิน) ซึ่งรวมกับเฮกโซส (กลูโคส) กรดอะดีโนซิโนฟอสฟอริกที่มีอยู่ในกล้ามเนื้อยังถูกย่อยสลายเพื่อสร้างกรดอะดีโนซีนและกรดฟอสฟอริก ซึ่งเมื่อรวมกับเฮกโซส (กลูโคส) จะส่งเสริมการก่อตัวของกรดแลคติก (เอ็มเดนและซิมเมอร์มันน์)
เนื้อสัตว์ที่ฆ่าใหม่ (เนื้อสด) มีเนื้อแน่นโดยไม่มีกลิ่นเฉพาะที่เด่นชัดเมื่อปรุงสุกจะให้น้ำซุปที่ไม่มีกลิ่นขุ่นและไม่มีคุณภาพรสชาติสูง นอกจากนี้ ในชั่วโมงแรกหลังการฆ่าสัตว์ เนื้อจะแข็งและแข็ง
24-72 ชั่วโมงหลังจากการฆ่าสัตว์ (ขึ้นอยู่กับอุณหภูมิของสิ่งแวดล้อมการเติมอากาศและปัจจัยอื่น ๆ ) เนื้อสัตว์จะได้รับตัวบ่งชี้คุณภาพใหม่: ความแข็งแกร่งของมันหายไปมันได้รับความชุ่มฉ่ำและมีกลิ่นหอมเฉพาะฟิล์มหนาแน่นก่อตัวขึ้น พื้นผิวของซาก (เปลือกแห้ง) ด้วยการปรุงอาหารให้น้ำซุปที่มีกลิ่นหอมโปร่งใสกลายเป็นนุ่ม ฯลฯ
กระบวนการและการเปลี่ยนแปลงที่เกิดขึ้นในเนื้อสัตว์ซึ่งเป็นผลมาจากการได้รับตัวบ่งชี้คุณภาพที่ต้องการนั้นมักเรียกว่าการสุกของเนื้อสัตว์
การสุกของเนื้อสัตว์เป็นการผสมผสานระหว่างกระบวนการทางชีวเคมีที่ซับซ้อนในเนื้อเยื่อของกล้ามเนื้อและการเปลี่ยนแปลงในโครงสร้างคอลลอยด์ทางกายภาพของโปรตีน ซึ่งเกิดขึ้นภายใต้การกระทำของเอ็นไซม์ของมันเอง
กระบวนการที่เกิดขึ้นในเนื้อเยื่อของกล้ามเนื้อหลังจากการฆ่าสัตว์สามารถแบ่งออกเป็นสามขั้นตอนต่อไปนี้: ความเข้มงวดหลังการชันสูตรพลิกศพ การสุกเต็มที่ และการย่อยสลายอัตโนมัติ
มอร์ทิสที่รุนแรงหลังการฆ่าจะพัฒนาในซากสัตว์ในชั่วโมงแรกหลังจากการฆ่าสัตว์ ในเวลาเดียวกัน กล้ามเนื้อจะยืดหยุ่นและสั้นลงเล็กน้อย ซึ่งจะช่วยเพิ่มความแข็งแกร่งและความต้านทานในการตัดได้อย่างมาก
ความสามารถในการบวมของเนื้อสัตว์นั้นต่ำมาก ที่อุณหภูมิ 15-20 "C มอร์ทิสที่เข้มงวดอย่างสมบูรณ์เกิดขึ้น 3-5 ชั่วโมงหลังจากการฆ่าสัตว์และที่อุณหภูมิ 0-2 ° C - หลังจาก 18-20 ชั่วโมง
กระบวนการของมอร์ทิสที่รุนแรงหลังการฆ่านั้นมาพร้อมกับอุณหภูมิที่เพิ่มขึ้นเล็กน้อยในซากอันเป็นผลมาจากการปล่อยความร้อนซึ่งเกิดขึ้นจากปฏิกิริยาเคมีที่เกิดขึ้นในเนื้อเยื่อ ความเข้มงวดของเนื้อเยื่อของกล้ามเนื้อที่สังเกตได้ในชั่วโมงแรกและวันแรกหลังจากการฆ่าสัตว์นั้นเกิดจากการก่อตัวของแอคโตไมโอซินคอมเพล็กซ์ที่ไม่ละลายน้ำจากโปรตีนแอคตินและไมโอซิน ข้อกำหนดเบื้องต้นสำหรับการก่อตัวของมันคือการขาดกรดอะดีโนซีนไตรฟอสฟอริก (ATP) สภาพแวดล้อมที่เป็นกรดของเนื้อสัตว์และการสะสมของกรดแลคติกในนั้น การเปลี่ยนแปลงทางชีวเคมีในเนื้อสัตว์ทำให้เกิดเงื่อนไขเหล่านี้
การลดลงและการหายไปอย่างสมบูรณ์ของ ATP นั้นสัมพันธ์กับการสลายของมันอันเป็นผลมาจากการทำงานของเอนไซม์ของ myosin การสลายของ ATP เป็น adenosine diphosphoric (ADP, adenosine monophosphoric (AMP) และกรดฟอสฟอริกในตัวเองทำให้เกิดสภาพแวดล้อมที่เป็นกรด ในเนื้อ นอกจากนี้ในระยะนี้การสลายตัวของไกลโคเจนในกล้ามเนื้อเริ่มต้นขึ้นซึ่งนำไปสู่การสะสมของกรดแลคติคซึ่งก่อให้เกิดสภาพแวดล้อมที่เป็นกรดในนั้น
สภาพแวดล้อมที่เป็นกรดซึ่งเป็นปรากฏการณ์ทางธรรมชาติของการสลายตัวของ ATP และจุดเริ่มต้นของกระบวนการไกลโคไลซิสที่ไม่สามารถย้อนกลับได้ (การสลายตัวของไกลโคเจนในกล้ามเนื้อ) จะเพิ่มความตึงของกล้ามเนื้อ สังเกตได้ว่ากล้ามเนื้อของสัตว์ที่ตายระหว่างการชักจะเกร็งเร็วขึ้น ความเข้มงวดที่ไม่มีการสะสมของกรดแลคติคนั้นมีลักษณะเป็นความตึงเครียดของกล้ามเนื้อที่อ่อนแอและการแก้ปัญหาอย่างรวดเร็วของกระบวนการ
อย่างไรก็ตาม ก่อนที่ระยะความเข้มงวดจะสิ้นสุดลง เนื้อสัตว์จะพัฒนากระบวนการที่เกี่ยวข้องกับระยะของการสุกและการย่อยสลายของเนื้อมันเอง
สองกระบวนการเป็นผู้นำสำหรับพวกเขา - การสลายตัวของไกลโคเจนในกล้ามเนื้ออย่างเข้มข้นซึ่งนำไปสู่การเปลี่ยนแปลงค่า pH ของเนื้อสัตว์ไปทางด้านกรดอย่างรวดเร็วรวมถึงการเปลี่ยนแปลงองค์ประกอบทางเคมีและโครงสร้างคอลลอยด์ทางกายภาพของโปรตีน
เนื่องจากความจริงที่ว่ากล้ามเนื้อของเนื้อสัตว์ไม่ได้รับออกซิเจนและกระบวนการออกซิเดชั่นถูกยับยั้งทำให้กรดแลคติคและฟอสฟอริกสะสมมากเกินไปในเนื้อสัตว์ ตัวอย่างเช่น เมื่อร่างกายเมื่อยล้าของกล้ามเนื้อ (ตลอดอายุขัย) กรดแลคติกจะไปถึงสูงสุด 0.25% และความเข้มงวดในการชันสูตรพลิกศพจะสะสมได้ถึง 0.82% ปฏิกิริยาแอคทีฟของตัวกลาง (pH) ในกรณีนี้เปลี่ยนจาก 7.26 เป็น 6.02 จากการสะสมของกรดแลคติกจะเกิดการหดตัวอย่างรวดเร็วของกล้ามเนื้อ (ความแข็งแกร่ง) พร้อมกับการแข็งตัวของโปรตีน (Saxl) ในกรณีนี้ Actomyosin จะสูญเสียความสามารถในการละลาย โปรตีนคงตัว และแคลเซียมตกตะกอนจากคอลลอยด์โปรตีนและไปเป็นสารละลาย (น้ำเนื้อ) เนื่องจากกรดแลคติกมีมากเกินไป การบวมของสารแอนไอโซโทรปิกคอลลอยด์ (ดิสก์มืด) ของเส้นใยกล้ามเนื้อจึงเกิดขึ้นเป็นครั้งแรก จากนั้นเมื่อความเข้มข้นของกรดแลคติกเพิ่มขึ้นและโปรตีนจับตัวเป็นก้อน สารนี้ก็จะอ่อนตัวลง โปรตีนที่จับตัวเป็นก้อนจะสูญเสียคุณสมบัติคอลลอยด์ ทำให้ไม่สามารถจับ (กัก) น้ำได้ และสูญเสียตัวกลางที่กระจัดกระจายไปในระดับหนึ่ง (น้ำ): แทนที่จะบวมครั้งแรก คอลลอยด์เซลล์จะหดตัว (หดตัว) และกล้ามเนื้อจะอ่อนตัว (ความละเอียดของความแข็งกระด้าง) ).
จากการสะสมของแลคติก ฟอสฟอริก และกรดอื่น ๆ ในเนื้อสัตว์ ความเข้มข้นของไฮโดรเจนไอออนเพิ่มขึ้น อันเป็นผลมาจากการที่ pH ลดลงเป็น 5.8-5.7 (และต่ำกว่านั้น) ในตอนท้ายของวัน
ในสภาพแวดล้อมที่เป็นกรด การสลายของ ATP, ADP, AMP และกรดฟอสฟอริกส่งผลให้เกิดการสะสมของฟอสฟอรัสอนินทรีย์บางส่วน สภาพแวดล้อมที่เป็นกรดอย่างรุนแรงและการปรากฏตัวของฟอสฟอรัสอนินทรีย์ถือเป็นสาเหตุของการแยกตัวของแอคโตไมโอซินคอมเพล็กซ์ไปเป็นแอคตินและไมโอซิน การสลายตัวของคอมเพล็กซ์นี้ช่วยขจัดปรากฏการณ์ของความแข็งและความแข็งแกร่งของเนื้อ ดังนั้นระยะที่เข้มงวดจึงไม่สามารถแยกออกจากระยะอื่นๆ ได้ และต้องถือว่าเป็นหนึ่งในขั้นตอนในกระบวนการสุกของเนื้อ
รูปแบบของการเปลี่ยนแปลงทางชีวเคมีในกระบวนการสุกเนื้อสามารถแสดงได้ดังนี้
สภาพแวดล้อมที่เป็นกรดนั้นทำหน้าที่ในการฆ่าเชื้อแบคทีเรียและแม้กระทั่งการฆ่าเชื้อแบคทีเรีย ดังนั้นเมื่อค่า pH เปลี่ยนไปทางด้านกรด จะทำให้เกิดสภาวะที่ไม่เอื้ออำนวยต่อการพัฒนาของจุลินทรีย์ในเนื้อสัตว์
ในที่สุด สภาพแวดล้อมที่เป็นกรดนำไปสู่การเปลี่ยนแปลงบางอย่างในองค์ประกอบทางเคมีและโครงสร้างทางกายภาพคอลลอยด์ของโปรตีน มันเปลี่ยนการซึมผ่านของเยื่อหุ้มกล้ามเนื้อและระดับของการกระจายตัวของโปรตีน กรดทำปฏิกิริยากับแคลเซียมโปรตีนและแคลเซียมแยกออกจากโปรตีน
การเปลี่ยนผ่านของแคลเซียมไปเป็นสารสกัดทำให้การกระจายตัวของโปรตีนลดลง อันเป็นผลมาจากการสูญเสียน้ำที่กักเก็บไฮเดรต ดังนั้นน้ำเนื้อสามารถแยกออกจากเนื้อสุกได้บางส่วนโดยการหมุนเหวี่ยง
น้ำที่กักเก็บไฮเดรตที่ปล่อยออกมา การทำงานของเอนไซม์ย่อยโปรตีนและสภาพแวดล้อมที่เป็นกรดทำให้เกิดเงื่อนไขในการคลายเนื้อเยื่อซาร์โคเลมมาของเส้นใยกล้ามเนื้อ และประการแรกคือการคลายและบวมของคอลลาเจน สิ่งนี้มีส่วนอย่างมากในการเปลี่ยนแปลงความสอดคล้องของเนื้อสัตว์และความชุ่มฉ่ำที่เด่นชัดยิ่งขึ้น เห็นได้ชัดว่าด้วยการบวมของคอลลาเจนและการปล่อยความชื้นบางส่วนจากพื้นผิวของซากไปสู่สิ่งแวดล้อม การก่อตัวของเปลือกแห้งบนพื้นผิวของมันควรจะสัมพันธ์กัน
ระยะของการเจริญเติบโตของตัวเองส่วนใหญ่จะกำหนดความเข้มของการไหลของกระบวนการคอลลอยด์ทางกายภาพและการเปลี่ยนแปลงโครงสร้างจุลภาคในเส้นใยกล้ามเนื้อซึ่งเกิดขึ้นในระยะ autolysis การสลายตัวของเนื้ออัตโนมัติในระหว่างการสุกของเนื้อจะลดลงในความหมายกว้างของคำและไม่เพียงเกี่ยวข้องกับการสลายตัวของโปรตีนเท่านั้น แต่ยังรวมถึงกระบวนการสลายตัวของส่วนประกอบต่างๆ ของเซลล์ด้วย ในเรื่องนี้ ไม่สามารถแยกหรือแยกกระบวนการที่เกิดขึ้นในระยะของการสุกของตัวมันเองออกจากกระบวนการในกระบวนการ autolysis อย่างไรก็ตาม อันเป็นผลมาจากความซับซ้อนของเหตุผล (การกระทำของเอนไซม์โปรตีโอไลติก, สภาพแวดล้อมที่เป็นกรดอย่างรวดเร็ว, ผลิตภัณฑ์จากการสลายตัวอัตโนมัติของสารที่ไม่ใช่โปรตีน ฯลฯ ) การสลายตัวอัตโนมัติของเส้นใยกล้ามเนื้อออกเป็นส่วนที่แยกจากกัน
การสุกของเนื้อจะเกิดขึ้นภายใน 24-72 ชั่วโมงที่อุณหภูมิ +4°
อย่างไรก็ตามไม่สามารถทนต่อเนื้อสัตว์ได้เสมอที่ +4 ° บางครั้งจำเป็นต้องเก็บไว้ภายใต้สภาวะปกติ (ไม่ใช่ในที่เย็น) ที่อุณหภูมิ + 6-8 °ขึ้นไป ที่อุณหภูมิสูง กระบวนการของการตายที่เข้มงวดและความละเอียดของกล้ามเนื้อจะดำเนินไปเร็วขึ้น อัตราการสุกของเนื้อสัตว์ยังขึ้นอยู่กับชนิดและสภาวะสุขภาพของสัตว์ที่ถูกฆ่า ความอ้วนและอายุของมันด้วย แต่คำถามเหล่านี้ต้องการการสังเกตและศึกษาเพิ่มเติม
ในระหว่างการสุกของเนื้อ นิวเคลียสบางส่วนจะถูกผ่าออก
(สารสกัดจากไนโตรเจน). เกิดสารระเหย เอสเทอร์ และอัลดีไฮด์ ซึ่งทำให้เนื้อมีรสชาติ กรด Adenylic และ inosinic, adenine, xanthine, hypoxanthine ซึ่งขึ้นอยู่กับรสชาติของเนื้อสัตว์ ปฏิกิริยาของสภาพแวดล้อมของเนื้อสัตว์เปลี่ยนแปลงไปสู่ความเป็นกรด (pH 6.2-
5.8) สิ่งนี้มีส่วนทำให้เกิดการบวมของคอลลอยด์โปรโตพลาสซึม เนื่องจากเนื้อจะนุ่ม นุ่ม และหุงต้มได้ดี
เนื้อสัตว์ที่มีคุณภาพนี้จะได้รับหลังจากเก็บรักษา 1-3 วันที่อุณหภูมิ 4 ถึง 12 ° (ขึ้นอยู่กับความสามารถขององค์กร)
ในขั้นตอนแรกของกระบวนการนี้ การแบ่งส่วนในเส้นใยกล้ามเนื้อแต่ละส่วนจะถูกตรวจพบโดยที่ยังคงรักษาเส้นใยเอนโดไมเซียลไว้ ในเวลาเดียวกันโครงสร้างของนิวเคลียส, แนวขวางและแนวยาวจะถูกเก็บรักษาไว้ในส่วน
ในขั้นตอนที่สอง เส้นใยกล้ามเนื้อส่วนใหญ่จะถูกแบ่งส่วน
เช่นเดียวกับในระยะแรก เอ็นโดมิเซียมของเส้นใย และในส่วน โครงสร้างของนิวเคลียส แนวขวาง และแนวยาวจะยังคงถูกรักษาไว้ ในที่สุด ในระยะที่สาม (ระยะของการ autolysis ลึก) ตรวจพบการสลายตัวของเซ็กเมนต์เป็น myofibrils และ myofibrils เป็น sarcomeres
Sarcomeres ในส่วนเล็ก ๆ ที่ทำจากเนื้อสัตว์ดังกล่าวจะถูกมองว่าเป็นมวลเม็ดเล็ก ๆ ที่ล้อมรอบด้วยเอนโดมิเซียม
การเปลี่ยนแปลงทางสัณฐานวิทยาและโครงสร้างจุลภาคในเนื้อเยื่อยังทำให้เนื้อนิ่มและคลายตัวในระหว่างการสุก เนื่องจากน้ำย่อยอาหารสามารถแทรกซึมเข้าไปในซาร์โคพลาสซึมได้อิสระมากขึ้น ซึ่งช่วยปรับปรุงการย่อยได้ ควรสังเกตว่าโปรตีนจากเนื้อเยื่อเกี่ยวพันแทบไม่ได้รับกระบวนการสลายโปรตีนในระหว่างการสุกของเนื้อสัตว์ ดังนั้นภายใต้สภาวะที่สุกงอมเท่ากัน ความอ่อนโยนของการตัดเนื้อที่แตกต่างกันของสัตว์ตัวเดียวกัน เช่นเดียวกับการตัดที่เหมือนกันของสัตว์ที่แตกต่างกัน จึงไม่เหมือนกัน ความอ่อนโยนของเนื้อสัตว์ที่มีเนื้อเยื่อเกี่ยวพันจำนวนมากมีขนาดเล็กและเนื้อของสัตว์เล็กจะนุ่มกว่าเนื้อเก่า
อันเป็นผลมาจากความซับซ้อนของการเปลี่ยนแปลงอัตโนมัติของส่วนประกอบต่างๆ ของเนื้อสัตว์ ในระหว่างการสุก สารจะก่อตัวและสะสมซึ่งกำหนดกลิ่นและรสชาติของเนื้อสุก เนื้อสัตว์ที่สุกแล้วจะได้รับรสชาติและกลิ่นที่หอมหวานด้วยสารสกัดที่มีไนโตรเจน เช่น ไฮโปแซนทีน ครีเอทีน และครีเอตินีน ซึ่งเกิดขึ้นระหว่างการแตกตัวของเอทีพี เช่นเดียวกับการสะสมกรดอะมิโนอิสระ (กรดกลูตามิก อาร์จินีน ทรีโอนีน ฟีนิลอะลานีน เป็นต้น) . เห็นได้ชัดว่ากรดไพรูวิกและแลคติคมีส่วนเกี่ยวข้องกับการก่อตัวของรสชาติและกลิ่นหอม
I. A. Smorodintsev เสนอว่ารสชาติและกลิ่นขึ้นอยู่กับการสะสมของสารที่ละลายได้ง่ายและระเหยง่าย เช่น เอสเทอร์ อัลดีไฮด์ และคีโตนในเนื้อสุก ต่อจากนั้น ผลการศึกษาจำนวนหนึ่งแสดงให้เห็นว่าคุณสมบัติในการหอมของเนื้อสุกดีขึ้นเมื่อปริมาณสารรีดิวซ์ที่ระเหยได้ทั้งหมดสะสมอยู่ในเนื้อนั้น ปัจจุบันโดยใช้แก๊สโครมาโตกราฟีและการวิเคราะห์แมสสเปกโตรเมตรีพบว่าสารประกอบที่ทำให้เกิดกลิ่นของเนื้อต้ม ได้แก่ อะซีตัลดีไฮด์ อะซิโตน เมทิล เอทิล คีโตน เมทานอล เมทิล เมอร์แคปแทน ไดเมทิล ซัลไฟด์ เอทิล เมอร์แคปแทน เป็นต้น
ด้วยอุณหภูมิที่เพิ่มขึ้น (สูงถึง 30 ° C) เช่นเดียวกับการได้รับเนื้อสัตว์เป็นเวลานาน (มากกว่า 20-26 วัน) ที่อุณหภูมิบวกต่ำ กระบวนการสุกของเอนไซม์จะลึกมากจนปริมาณของผลิตภัณฑ์สลายโปรตีนในรูปแบบ ของเปปไทด์ขนาดเล็กและกรดอะมิโนอิสระ ในขั้นตอนนี้เนื้อสัตว์จะได้สีน้ำตาลปริมาณของเอมีนและแอมโมเนียไนโตรเจนเพิ่มขึ้นทำให้เกิดการสลายตัวของไขมันไฮโดรไลติกที่เห็นได้ชัดเจนซึ่งช่วยลดคุณภาพทางการค้าและคุณค่าทางโภชนาการลงอย่างรวดเร็ว
กระบวนการทางชีวเคมีที่เกิดขึ้นระหว่างการสุกในเนื้อสัตว์ที่เป็นโรคนั้นแตกต่างจากกระบวนการทางชีวเคมีในเนื้อสัตว์ที่มีสุขภาพดี
ด้วยไข้และการทำงานหนักเกินไป กระบวนการพลังงานในร่างกายจะเพิ่มขึ้น
กระบวนการออกซิเดชันในเนื้อเยื่อได้รับการปรับปรุง การเปลี่ยนแปลงของการเผาผลาญคาร์โบไฮเดรตในระหว่างการเจ็บป่วยและการทำงานหนักเกินไปนั้นมีลักษณะเฉพาะโดยการลดระดับไกลโคเจนในกล้ามเนื้ออย่างรวดเร็ว ดังนั้นเกือบทุกกระบวนการทางพยาธิวิทยาในร่างกายของสัตว์ปริมาณไกลโคเจนในกล้ามเนื้อจึงลดลง เนื่องจากมีไกลโคเจนในเนื้อสัตว์ที่ป่วยน้อยกว่าในเนื้อสัตว์ที่มีสุขภาพดี ปริมาณของผลิตภัณฑ์ที่สลายไกลโคเจน (กลูโคส กรดแลคติก ฯลฯ) ในเนื้อสัตว์ที่ป่วยจึงไม่มีนัยสำคัญเช่นกัน
นอกจากนี้ในช่วงที่มีโรคร้ายแรงแม้ในช่วงชีวิตของสัตว์ผลิตภัณฑ์ระดับกลางและขั้นสุดท้ายของการเผาผลาญโปรตีนจะสะสมในกล้ามเนื้อ ในกรณีเหล่านี้ ในชั่วโมงแรกหลังจากการฆ่าสัตว์ จะพบปริมาณเอมีนและแอมโมเนียไนโตรเจนเพิ่มขึ้นในเนื้อสัตว์
การสะสมของกรดเล็กน้อยและปริมาณโพลีเปปไทด์ กรดอะมิโน และแอมโมเนียที่เพิ่มขึ้นเป็นสาเหตุของการลดลงของความเข้มข้นของไฮโดรเจนไอออนในระหว่างการสุกของเนื้อสัตว์ที่ป่วย ปัจจัยนี้ส่งผลต่อการทำงานของเอ็นไซม์เนื้อ ในกรณีส่วนใหญ่ ความเข้มข้นของไฮโดรเจนไอออนซึ่งเกิดขึ้นจากการสุกของเนื้อสัตว์ที่ป่วย เป็นที่นิยมมากกว่าสำหรับการกระทำของเปปไทเดสและโปรตีเอส
เป็นผลให้การสะสมของสารไนโตรเจนที่สกัดได้ในเนื้อสัตว์ที่ป่วยและการขาดการเปลี่ยนแปลงอย่างรวดเร็วของค่า pH ไปทางด้านกรดถือเป็นเงื่อนไขที่ดีสำหรับการพัฒนาของจุลินทรีย์
การเปลี่ยนแปลงที่เกิดขึ้นในเนื้อสัตว์ที่ป่วยยังส่งผลต่อธรรมชาติของโครงสร้างคอลลอยด์ทางกายภาพของเนื้อสัตว์ในลักษณะที่ต่างออกไป ความเป็นกรดที่น้อยลงทำให้เกิดการตกตะกอนของเกลือแคลเซียมเล็กน้อย ซึ่งในทางกลับกัน ก็เป็นสาเหตุของการเปลี่ยนแปลงเล็กน้อยในระดับของการกระจายตัวของโปรตีนและลักษณะการเปลี่ยนแปลงอื่นๆ ของพวกมันในระหว่างการสุกปกติของเนื้อสัตว์ ค่า pH ที่ค่อนข้างสูง การสะสมของผลิตภัณฑ์จากการย่อยสลายโปรตีน และสภาวะที่เอื้ออำนวยต่อการพัฒนาของจุลินทรีย์จะกำหนดความต้านทานที่ต่ำกว่าของเนื้อสัตว์ที่ป่วยในระหว่างการเก็บรักษา เครื่องหมายที่ระบุเป็นลักษณะเนื้อของสัตว์ป่วยหนักแต่ละชนิด พวกเขาเป็นสาเหตุของความสม่ำเสมอที่รู้จักกันดีในการเปลี่ยนแปลงพารามิเตอร์ทางเคมีกายภาพของเนื้อสัตว์ที่ได้จากสัตว์ที่ถูกฆ่าตายในระหว่างกระบวนการทางพยาธิวิทยาโดยไม่คำนึงถึงลักษณะของโรค บทบัญญัตินี้ไม่ได้ปฏิเสธการเปลี่ยนแปลงเฉพาะในองค์ประกอบของเนื้อสัตว์ในบางโรค แต่ให้เหตุผลที่จะพูดถึงรูปแบบทั่วไปของการเจริญเติบโตของเนื้อสัตว์ในพยาธิสภาพในร่างกายของสัตว์
สารประกอบที่มีน้ำหนักโมเลกุลสูง ที่ สารประกอบรู้กันหมด โปรตีน...คงรูปร่างและ เคมี สารประกอบแม้จะต่อเนื่อง...
เคมี สารประกอบและคุณสมบัติทางกายภาพของตัวอสุจิ
รายงาน >> ยา สุขภาพเคมี สารประกอบและคุณสมบัติทางกายภาพของสเปิร์ม สเปิร์มเป็นส่วนผสมของ ... , A) มาโครและไมโครอิลิเมนต์ เคมี สารประกอบอสุจิ: 1) น้ำ - 75% 2) วัตถุแห้ง - 25%: - กระรอก- 85% - ไขมัน...
กรดอะมิโน - ส่วนประกอบโครงสร้าง โปรตีน โปรตีนหรือ โปรตีน(โปรโตสกรีก - สำคัญยิ่ง) เป็นเฮเทอโรพอลิเมอร์ทางชีวภาพซึ่งโมโนเมอร์คือกรดอะมิโน
กรดอะมิโนเป็นสารประกอบอินทรีย์ที่มีน้ำหนักโมเลกุลต่ำซึ่งประกอบด้วยกลุ่มคาร์บอกซิล (-COOH) และอะมิโน (-NH 2) ที่เชื่อมโยงกับอะตอมของคาร์บอนเดียวกัน โซ่ข้างติดอยู่กับอะตอมของคาร์บอน - อนุมูลที่ให้คุณสมบัติบางอย่างของกรดอะมิโนแต่ละชนิด สูตรทั่วไปของกรดอะมิโนคือ:

กรดอะมิโนส่วนใหญ่มีหมู่คาร์บอกซิลหนึ่งหมู่และหมู่อะมิโนหนึ่งหมู่ กรดอะมิโนเหล่านี้เรียกว่า เป็นกลาง.แต่ก็มี กรดอะมิโนพื้นฐาน- มีหมู่อะมิโนมากกว่า 1 หมู่ และยัง กรดอะมิโนที่เป็นกรด- มีหมู่คาร์บอกซิลมากกว่าหนึ่งกลุ่ม
ทราบว่ามีกรดอะมิโนประมาณ 200 ชนิดเกิดขึ้นในสิ่งมีชีวิต แต่มีเพียง 20 ชนิดเท่านั้นที่เป็นส่วนหนึ่งของโปรตีน สิ่งเหล่านี้เรียกว่า ขั้นพื้นฐาน,หรือ การสร้างโปรตีน(โปรตีน) กรดอะมิโน
ขึ้นอยู่กับชนิดของอนุมูล กรดอะมิโนพื้นฐานแบ่งออกเป็นสามกลุ่ม: 1) ไม่มีขั้ว (อะลานีน, เมไทโอนีน, วาลีน, โปรไลน์, ลิวซีน, ไอโซลิวซีน, ทริปโตเฟน, ฟีนิลอะลานีน); 2) ขั้วไม่มีประจุ (แอสพาราจีน, กลูตามีน, ซีรีน, ไกลซีน, ไทโรซีน, ทรีโอนีน, ซิสเทอีน); 3) ประจุมีขั้ว (อาร์จินีน, ฮิสติดีน, ไลซีน - บวก; กรดแอสปาร์ติกและกลูตามิก - ลบ)
สายด้านข้างของกรดอะมิโน (radical) สามารถเป็นแบบไม่ชอบน้ำหรือชอบน้ำ ซึ่งทำให้โปรตีนมีคุณสมบัติที่สอดคล้องกันซึ่งปรากฏระหว่างการก่อตัวของโครงสร้างโปรตีนทุติยภูมิ ตติยภูมิ และควอเทอร์นารี
ในพืช ทั้งหมดกรดอะมิโนที่จำเป็นถูกสังเคราะห์จากผลิตภัณฑ์หลักของการสังเคราะห์ด้วยแสง มนุษย์และสัตว์ไม่สามารถสังเคราะห์กรดอะมิโนที่สร้างโปรตีนได้จำนวนหนึ่งและต้องได้รับกรดอะมิโนสำเร็จรูปพร้อมอาหาร กรดอะมิโนดังกล่าวเรียกว่า ไม่สามารถถูกแทนที่ได้ ถึงพวกเขารวมถึงไลซีน, วาลีน, ลิวซีน, ไอโซลิวซีน, ทรีโอนีน, ฟีนิลอะลานีน, ทริปโตเฟน, เมไทโอนีน; เช่นเดียวกับอาร์จินีนและฮิสติดีน - ที่ขาดไม่ได้สำหรับเด็ก
ในสารละลาย กรดอะมิโนสามารถทำหน้าที่เป็นทั้งกรดและเบส กล่าวคือ เป็นสารประกอบแอมโฟเทอริก หมู่คาร์บอกซิล -COOH สามารถบริจาคโปรตอนซึ่งทำหน้าที่เป็นกรด และหมู่เอมีน - NH2 - เพื่อรับโปรตอน ดังนั้นจึงแสดงคุณสมบัติของเบส
เปปไทด์ หมู่อะมิโนของกรดอะมิโนหนึ่งสามารถทำปฏิกิริยากับหมู่คาร์บอกซิลของกรดอะมิโนอื่นได้
โมเลกุลที่ได้คือไดเปปไทด์ และพันธะ -CO-NH- เรียกว่าพันธะเปปไทด์:

ที่ปลายด้านหนึ่งของโมเลกุลไดเปปไทด์คือหมู่อะมิโนอิสระ และที่ปลายอีกด้านหนึ่งคือหมู่คาร์บอกซิลอิสระ ด้วยเหตุนี้ไดเปปไทด์จึงสามารถเกาะกับกรดอะมิโนอื่น ๆ เพื่อสร้างโอลิโกเปปไทด์ได้ หากกรดอะมิโนจำนวนมาก (มากกว่าสิบ) เชื่อมต่อกันในลักษณะนี้ ปรากฎว่า โพลีเปปไทด์
เปปไทด์มีบทบาทสำคัญในร่างกาย โอลิโกและโพลีเปปไทด์จำนวนมากเป็นฮอร์โมน ยาปฏิชีวนะ สารพิษ
Oligopeptides ได้แก่ oxytocin, vasopressin, thyrotropin รวมทั้ง bradykinin (pain peptide) และยาหลับในบางชนิด ("ยาธรรมชาติ" ของบุคคล) ที่ทำหน้าที่บรรเทาอาการปวด การเสพยาทำลายระบบฝิ่นของร่างกาย ดังนั้นผู้ติดยาที่ไม่ได้รับยาจะประสบกับความเจ็บปวดอย่างรุนแรง - "การถอนตัว" ซึ่งปกติจะบรรเทาได้ด้วยฝิ่น Oligopeptides ยังรวมถึงยาปฏิชีวนะบางชนิด (เช่น gramicidin S)
ฮอร์โมนหลายชนิด (อินซูลิน ฮอร์โมนอะดรีโนคอร์ติโคทรอปิก ฯลฯ) ยาปฏิชีวนะ (เช่น แกรมซิซิดิน เอ) สารพิษ (เช่น พิษคอตีบ) เป็นโพลีเปปไทด์
โปรตีนคือโพลีเปปไทด์ ซึ่งโมเลกุลประกอบด้วยกรดอะมิโนตั้งแต่ห้าสิบถึงหลายพันตัวที่มีน้ำหนักโมเลกุลสัมพัทธ์มากกว่า 10,000
โครงสร้างของโปรตีน โปรตีนแต่ละชนิดในสภาพแวดล้อมที่กำหนดมีลักษณะเฉพาะด้วยโครงสร้างเชิงพื้นที่พิเศษ เมื่อกำหนดลักษณะโครงสร้างเชิงพื้นที่ (สามมิติ) จะแยกแยะการจัดระเบียบโมเลกุลโปรตีนสี่ระดับ (รูปที่ 1.1)
|
โกหก—glu—tre—ala—ala—ala—lis—fen—glu—arg—gln—gis—met—asp—ser— |
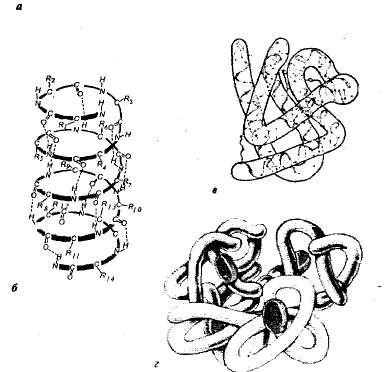
ข้าว. 1.1.ระดับการจัดโครงสร้างโปรตีน: a — โครงสร้างหลักคือลำดับกรดอะมิโนของโปรตีนไรโบนิวคลีเอส (124 หน่วยของกรดอะมิโน) ข — โครงสร้างรอง — โซ่โพลีเปปไทด์บิดเป็นเกลียว ใน— โครงสร้างระดับอุดมศึกษาของโปรตีน myoglobin; G — โครงสร้างควอเทอร์นารีของเฮโมโกลบิน
โครงสร้างหลัก- ลำดับของกรดอะมิโนในสายโซ่พอลิเปปไทด์ โครงสร้างดังกล่าวมีความเฉพาะเจาะจงสำหรับโปรตีนแต่ละชนิดและถูกกำหนดโดยข้อมูลทางพันธุกรรม กล่าวคือ ขึ้นอยู่กับลำดับของนิวคลีโอไทด์ในบริเวณโมเลกุลดีเอ็นเอที่เข้ารหัสโปรตีนนี้ คุณสมบัติและหน้าที่ทั้งหมดของโปรตีนขึ้นอยู่กับโครงสร้างหลัก การแทนที่กรดอะมิโนเดี่ยวในองค์ประกอบของโมเลกุลโปรตีนหรือการละเมิดลำดับในการจัดเรียงมักจะทำให้เกิดการเปลี่ยนแปลงในการทำงานของโปรตีน
เมื่อพิจารณาว่าโปรตีนประกอบด้วยกรดอะมิโน 20 ชนิด จำนวนตัวแปรของการผสมผสานของพวกมันในสายโซ่โพลีเปปไทด์นั้นไม่จำกัดอย่างแท้จริง ซึ่งให้โปรตีนจำนวนมากในเซลล์ของสิ่งมีชีวิต ตัวอย่างเช่น พบโปรตีนมากกว่า 10,000 ชนิดในร่างกายมนุษย์ และพวกมันทั้งหมดถูกสร้างขึ้นจากกรดอะมิโนพื้นฐาน 20 ชนิดเดียวกัน
ในเซลล์ที่มีชีวิต โมเลกุลโปรตีนหรือแต่ละส่วนของพวกมันไม่ใช่สายโซ่ยาว แต่บิดเป็นเกลียวคล้ายกับสปริงที่ขยายออก (นี่คือสิ่งที่เรียกว่า a-helix) หรือพับเป็นชั้นพับ (p-layer) a-helices และ p-layers ดังกล่าวเป็นเรื่องรอง โครงสร้าง.เป็นผลมาจากการก่อตัวของพันธะไฮโดรเจนภายในสายโพลีเปปไทด์หนึ่งสาย (โครงแบบเกลียว) หรือระหว่างสายโพลีเปปไทด์สองสาย (ชั้นพับ)
โปรตีนเคราตินมีลักษณะเป็นเกลียวที่สมบูรณ์ มันเป็นโปรตีนโครงสร้างของเส้นผม เล็บ กรงเล็บ จะงอยปาก ขน และเขา; มันเป็นส่วนหนึ่งของชั้นนอกของผิวหนังของสัตว์มีกระดูกสันหลัง
ในโปรตีนส่วนใหญ่ ส่วนที่เป็นเกลียวและส่วนที่ไม่ใช่เกลียวของสายพอลิเปปไทด์จะถูกพับเป็นรูปทรงกลมสามมิติ ซึ่งเป็นรูปทรงกลม (มีลักษณะเฉพาะของโปรตีนทรงกลม) ลูกกลมของการกำหนดค่าเฉพาะคือ โครงสร้างระดับอุดมศึกษากระรอก. โครงสร้างดังกล่าวทำให้เสถียรโดยพันธะไอออนิก ไฮโดรเจน โควาเลนต์ไดซัลไฟด์ (เกิดขึ้นระหว่างอะตอมของกำมะถันที่เป็นส่วนหนึ่งของซิสเทอีน ซีสทีน และเมจิโอนีน) รวมถึงปฏิกิริยาที่ไม่ชอบน้ำ ปฏิกิริยาที่ไม่ชอบน้ำเป็นสิ่งสำคัญที่สุดในการก่อตัวของโครงสร้างตติยภูมิ ในเวลาเดียวกันโปรตีนจะพับในลักษณะที่โซ่ด้านข้างที่ไม่ชอบน้ำนั้นถูกซ่อนอยู่ภายในโมเลกุลนั่นคือพวกมันได้รับการปกป้องจากการสัมผัสกับน้ำและในทางกลับกันโซ่ด้านที่ชอบน้ำนั้นถูกเปิดเผยสู่ภายนอก
โปรตีนหลายชนิดที่มีโครงสร้างซับซ้อนเป็นพิเศษประกอบด้วยสายโซ่โพลีเปปไทด์หลายสาย (หน่วยย่อย) ก่อตัวขึ้น โครงสร้างสี่ส่วนโมเลกุลโปรตีน โครงสร้างดังกล่าวมีอยู่ ตัวอย่างเช่น ในโปรตีนทรงกลมของเฮโมโกลบิน โมเลกุลของมันประกอบด้วยหน่วยย่อยโพลีเปปไทด์สี่หน่วยแยกกัน (โปรโตเมอร์) ที่อยู่ในโครงสร้างระดับตติยภูมิและส่วนที่ไม่ใช่โปรตีน - heme
เฉพาะในโครงสร้างดังกล่าวเท่านั้นที่เฮโมโกลบินสามารถทำหน้าที่ขนส่งได้
ภายใต้อิทธิพลของปัจจัยทางเคมีและกายภาพต่างๆ (การบำบัดด้วยแอลกอฮอล์, อะซิโตน, กรด, ด่าง, อุณหภูมิสูง, การฉายรังสี, ความดันสูง, ฯลฯ ), โครงสร้างรอง, ตติยภูมิและควอเทอร์นารีของโปรตีนเปลี่ยนแปลงเนื่องจากการแตกของไฮโดรเจนและ พันธะไอออนิก กระบวนการทำลายโครงสร้างดั้งเดิม (ตามธรรมชาติ) ของโปรตีนเรียกว่า การทำให้เสียสภาพในกรณีนี้จะสังเกตเห็นการลดลงของความสามารถในการละลายโปรตีนการเปลี่ยนแปลงในรูปร่างและขนาดของโมเลกุลการสูญเสียกิจกรรมของเอนไซม์ ฯลฯ กระบวนการทำให้เสียสภาพอาจสมบูรณ์หรือบางส่วน ในบางกรณี การเปลี่ยนแปลงไปสู่สภาวะแวดล้อมปกติจะมาพร้อมกับการฟื้นฟูโครงสร้างตามธรรมชาติของโปรตีนที่เกิดขึ้นเองตามธรรมชาติ กระบวนการดังกล่าวเรียกว่า การปรับสภาพใหม่
โปรตีนที่ง่ายและซับซ้อน ตามองค์ประกอบทางเคมี โปรตีนแบ่งออกเป็นแบบง่ายและซับซ้อน ยกโทษให้ฉันโปรตีนประกอบด้วยกรดอะมิโนเท่านั้น ยาก- โปรตีนที่มีส่วนโปรตีนและส่วนที่ไม่ใช่โปรตีน (เทียม) กลุ่มเทียมสามารถเกิดขึ้นได้จากไอออนของโลหะ กรดฟอสฟอริกตกค้าง คาร์โบไฮเดรต ไขมัน ฯลฯ โปรตีนอย่างง่าย ได้แก่ อัลบูมินในเลือด ไฟบริน เอ็นไซม์บางชนิด (ทริปซิน) เป็นต้น โปรตีนเชิงซ้อน ได้แก่ โปรตีโอลิปิดและไกลโคโปรตีนทั้งหมด โปรตีนที่ซับซ้อน เช่น อิมมูโนโกลบูลิน (แอนติบอดี) เฮโมโกลบิน เอนไซม์ส่วนใหญ่ เป็นต้น
หน้าที่ของโปรตีน
- โครงสร้าง.โปรตีนเป็นส่วนหนึ่งของเยื่อหุ้มเซลล์และเมทริกซ์ของออร์แกเนลล์ของเซลล์ ผนังหลอดเลือด กระดูกอ่อน เส้นเอ็น ผม เล็บ กรงเล็บในสัตว์ชั้นสูงประกอบด้วยโปรตีนเป็นหลัก
- ตัวเร่งปฏิกิริยา (เอนไซม์)เอนไซม์โปรตีนกระตุ้นปฏิกิริยาเคมีทั้งหมดในร่างกาย พวกมันให้การสลายสารอาหารในทางเดินอาหาร การตรึงคาร์บอนระหว่างการสังเคราะห์ด้วยแสง ฯลฯ
- ขนส่ง.โปรตีนบางชนิดสามารถเกาะติดและนำพาสารต่างๆ ได้ อัลบูมินในเลือดขนส่งกรดไขมัน โกลบูลิน - ไอออนของโลหะและฮอร์โมน เฮโมโกลบิน - ออกซิเจนและคาร์บอนไดออกไซด์ โมเลกุลของโปรตีนที่ประกอบเป็นเยื่อหุ้มพลาสมามีส่วนเกี่ยวข้องกับการขนส่งสารเข้าสู่เซลล์
- ป้องกันมันดำเนินการโดยอิมมูโนโกลบูลิน (แอนติบอดี) ของเลือดซึ่งให้การป้องกันภูมิคุ้มกันของร่างกาย ไฟบริโนเจนและทรอมบินเกี่ยวข้องกับการแข็งตัวของเลือดและป้องกันเลือดออก
- คอนแทรคไทล์เนื่องจากการเลื่อนของแอกตินและไมโอซินโปรโตไฟบริลที่สัมพันธ์กัน การหดตัวของกล้ามเนื้อจึงเกิดขึ้น รวมถึงการหดตัวภายในเซลล์ที่ไม่ใช่ของกล้ามเนื้อ การเคลื่อนไหวของ cilia และ flagella สัมพันธ์กับการเลื่อนของ microtubules ที่สัมพันธ์กันซึ่งมีลักษณะเป็นโปรตีน
- ระเบียบข้อบังคับฮอร์โมนหลายชนิดเป็นโอลิโกเปปไทด์หรือแย่ (เช่น อินซูลิน กลูคากอน [ตัวต้านอินซูลิน] ฮอร์โมนอะดรีโนคอร์ติโคทรอปิก ฯลฯ)
- ตัวรับโปรตีนบางชนิดที่ฝังอยู่ในเยื่อหุ้มเซลล์สามารถเปลี่ยนโครงสร้างได้ภายใต้อิทธิพลของสภาพแวดล้อมภายนอก นี่คือวิธีรับสัญญาณจากภายนอกและส่งข้อมูลไปยังเซลล์ ตัวอย่างจะเป็น ไฟโตโครเมียม—- โปรตีนไวแสงที่ควบคุมการตอบสนองช่วงแสงของพืชและ ออปซิน -ส่วนประกอบ โรดอปซิน,เม็ดสีที่พบในเซลล์ของเรตินา
- พลังงาน.โปรตีนสามารถทำหน้าที่เป็นแหล่งพลังงานในเซลล์ (หลังจากการไฮโดรไลซิส) โดยปกติ โปรตีนจะใช้สำหรับความต้องการพลังงานในกรณีที่รุนแรง เมื่อปริมาณคาร์โบไฮเดรตและไขมันสำรองหมดลง
เอ็นไซม์ (เอ็นไซม์). เหล่านี้เป็นโปรตีนจำเพาะที่มีอยู่ในสิ่งมีชีวิตทั้งหมดและมีบทบาทเป็นตัวเร่งปฏิกิริยาทางชีวภาพ
ปฏิกิริยาเคมีในเซลล์ที่มีชีวิตดำเนินไปที่อุณหภูมิหนึ่ง ความดันปกติ และความเป็นกรดที่สอดคล้องกันของตัวกลาง ภายใต้สภาวะดังกล่าว ปฏิกิริยาของการสังเคราะห์หรือการสลายตัวของสารจะดำเนินไปอย่างช้าๆ ในเซลล์ หากไม่ได้สัมผัสกับเอนไซม์ เอนไซม์เร่งปฏิกิริยาโดยไม่เปลี่ยนผลลัพธ์โดยรวมโดยการลด พลังงานกระตุ้น,กล่าวคือ ในการปรากฏตัวของพวกมัน พลังงานน้อยกว่ามากจะต้องใช้ในการแสดงปฏิกิริยาต่อโมเลกุลที่ทำปฏิกิริยา หรือปฏิกิริยาดำเนินไปตามเส้นทางที่แตกต่างกันโดยมีอุปสรรคด้านพลังงานที่ต่ำกว่า
กระบวนการทั้งหมดในสิ่งมีชีวิตนั้นดำเนินการโดยตรงหรือโดยอ้อมด้วยการมีส่วนร่วมของเอนไซม์ ตัวอย่างเช่น ภายใต้การกระทำของพวกเขา ส่วนประกอบที่เป็นส่วนประกอบของอาหาร (โปรตีน คาร์โบไฮเดรต ไขมัน ฯลฯ) ถูกแบ่งออกเป็นสารประกอบที่ง่ายกว่า จากนั้นจึงสังเคราะห์ลักษณะโมเลกุลขนาดใหญ่ใหม่ของประเภทนี้ ดังนั้นการละเมิดการก่อตัวและกิจกรรมของเอนไซม์จึงมักนำไปสู่การเกิดโรคร้ายแรง
ตามโครงสร้างเชิงพื้นที่ เอ็นไซม์ประกอบด้วยสายโพลีและเปปไทด์หลายสาย และมักจะมีโครงสร้างสี่ส่วน นอกจากนี้ เอ็นไซม์ยังสามารถรวมโครงสร้างที่ไม่ใช่โปรตีน ส่วนโปรตีนสวมใส่ ชื่ออะพอนไซม์,และไม่ใช่โปรตีน ปัจจัยร่วม(ถ้าเป็นไอออนบวกหรือแอนไอออนของสารอนินทรีย์ เช่น Zn 2- Mn 2+ เป็นต้น) หรือ โคเอ็นไซม์ (โคเอ็นไซม์)(ถ้าเป็นสารอินทรีย์น้ำหนักโมเลกุลต่ำ)
สารตั้งต้นหรือส่วนประกอบของโคเอ็นไซม์หลายชนิดคือวิตามิน ดังนั้นกรด pantothenic จึงเป็นส่วนประกอบสำคัญของโคเอ็นไซม์ A กรดนิโคตินิก (วิตามิน PP) เป็นสารตั้งต้นของ NAD และ NADP เป็นต้น
ตัวเร่งปฏิกิริยาด้วยเอนไซม์เป็นไปตามกฎหมายเดียวกับตัวเร่งปฏิกิริยาที่ไม่ใช่เอนไซม์ในอุตสาหกรรมเคมี แต่แตกต่างจากตัวเร่งปฏิกิริยาตรงที่มีลักษณะผิดปกติ ความจำเพาะสูง(เอ็นไซม์เร่งปฏิกิริยาเพียงปฏิกิริยาเดียวหรือทำหน้าที่เพียงพันธะประเภทเดียวเท่านั้น) สิ่งนี้ทำให้มั่นใจถึงการควบคุมที่ดีของกระบวนการที่สำคัญทั้งหมด (การหายใจ การย่อยอาหาร การสังเคราะห์ด้วยแสง ฯลฯ) ที่เกิดขึ้นในเซลล์และร่างกาย ตัวอย่างเช่น เอนไซม์ยูเรียเร่งการสลายตัวของสารเพียงชนิดเดียว - ยูเรีย (H 2 N-CO-NH 2 + H 2 O -> -» 2NH 3 + CO 2) โดยไม่มีผลต่อตัวเร่งปฏิกิริยาของสารประกอบที่เกี่ยวข้องกับโครงสร้าง
เพื่อให้เข้าใจกลไกการออกฤทธิ์ของเอ็นไซม์ที่มีความจำเพาะสูงเป็นอย่างมาก ทฤษฎีศูนย์แอคทีฟมีความสำคัญตามที่เธอ, ในโมเลกุล ทุกคนเอนไซม์ มีหนึ่งไซต์หรือมากกว่าซึ่งการเร่งปฏิกิริยาเกิดขึ้นเนื่องจากการสัมผัสอย่างใกล้ชิด (หลายจุด) ระหว่างโมเลกุลของเอนไซม์กับสารเฉพาะ (สารตั้งต้น) ศูนย์แอคทีฟคือกลุ่มการทำงาน (เช่น กลุ่มซีรีน OH) หรือกรดอะมิโนเดี่ยว โดยปกติ สำหรับการเร่งปฏิกิริยา จำเป็นต้องมีการรวมกันของสารตกค้างของกรดอะมิโนหลายตัว (โดยเฉลี่ยตั้งแต่ 3 ถึง 12) ตัวที่จัดเรียงในลำดับที่แน่นอน ศูนย์แอคทีฟยังเกิดขึ้นจากไอออนของโลหะที่เกี่ยวข้องกับเอนไซม์ วิตามิน และสารประกอบที่ไม่ใช่โปรตีนอื่นๆ เช่น โคเอ็นไซม์ หรือโคแฟคเตอร์ นอกจากนี้รูปร่างและโครงสร้างทางเคมีของศูนย์แอคทีฟก็เป็นเช่นนั้น กับมีเพียงวัสดุพิมพ์บางประเภทเท่านั้นที่สามารถผูกติดกับมันได้เนื่องจากการสอดคล้องกันในอุดมคติ บทบาทของส่วนที่เหลือของกรดอะมิโนในโมเลกุลของเอนไซม์ขนาดใหญ่คือการจัดเตรียมโมเลกุลที่มีรูปร่างเป็นทรงกลมที่เหมาะสม ซึ่งจำเป็นสำหรับการทำงานอย่างมีประสิทธิภาพของศูนย์แอคทีฟ นอกจากนี้ สนามไฟฟ้าแรงยังเกิดขึ้นรอบโมเลกุลของเอนไซม์ขนาดใหญ่ ในสาขาดังกล่าว การวางแนวของโมเลกุลของสารตั้งต้นและการได้มาซึ่งรูปร่างอสมมาตรเป็นไปได้ สิ่งนี้นำไปสู่การอ่อนตัวของพันธะเคมี และปฏิกิริยาเร่งปฏิกิริยาเกิดขึ้นกับการใช้พลังงานเริ่มต้นที่ต่ำกว่า ดังนั้นจึงมีอัตราที่สูงกว่ามาก ตัวอย่างเช่น หนึ่งโมเลกุลของเอนไซม์คาตาเลสสามารถสลายไฮโดรเจนเปอร์ออกไซด์ได้มากกว่า 5 ล้านโมเลกุล (H 2 0 2) ใน 1 นาที ซึ่งเกิดขึ้นเมื่อสารประกอบต่างๆ ถูกออกซิไดซ์ในร่างกาย
ในเอนไซม์บางตัว เมื่อมีซับสเตรต การกำหนดค่าของศูนย์แอคทีฟจะมีการเปลี่ยนแปลง กล่าวคือ เอ็นไซม์จะปรับกลุ่มการทำงานของมันในลักษณะที่จะให้กิจกรรมเร่งปฏิกิริยาที่ยิ่งใหญ่ที่สุด
ในขั้นตอนสุดท้ายของปฏิกิริยาเคมี สารตั้งต้นของเอนไซม์-สารตั้งต้นจะถูกแยกออกด้วยการก่อตัวของผลิตภัณฑ์สุดท้ายและเอนไซม์อิสระ ศูนย์แอคทีฟที่ปล่อยออกมาในกรณีนี้สามารถรับโมเลกุลของสารตั้งต้นใหม่ได้
อัตราการเกิดปฏิกิริยาของเอนไซม์ขึ้นอยู่กับปัจจัยหลายประการ: ธรรมชาติและความเข้มข้นของเอนไซม์และสารตั้งต้น อุณหภูมิ ความดัน ความเป็นกรดของตัวกลาง การมีอยู่ของสารยับยั้ง ฯลฯ ตัวอย่างเช่น ที่อุณหภูมิใกล้ศูนย์ อัตราการเกิดปฏิกิริยาทางชีวเคมีจะช้าลงจนเหลือน้อยที่สุด . คุณสมบัตินี้ใช้กันอย่างแพร่หลายในภาคต่างๆ ของเศรษฐกิจ โดยเฉพาะอย่างยิ่ง ในด้านการเกษตรและการแพทย์ โดยเฉพาะการอนุรักษ์ของอวัยวะต่างๆ (ไต, หัวใจ, ม้าม, ตับ) ก่อนการปลูกถ่ายให้กับผู้ป่วยในช่วงที่เย็นลง เพื่อลดความรุนแรงของปฏิกิริยาทางชีวเคมีและยืดอายุของอวัยวะ การแช่แข็งอาหารอย่างรวดเร็วช่วยป้องกันการเจริญเติบโตและการเพิ่มจำนวนของจุลินทรีย์ (แบคทีเรีย เชื้อรา ฯลฯ) และยังหยุดการทำงานของเอนไซม์ย่อยอาหาร เพื่อไม่ให้เกิดการย่อยสลายของอาหารอีกต่อไป
แหล่งที่มา : บน. Lemeza L.V. กมลยุกต์ Lisov "คู่มือชีววิทยาสำหรับผู้สมัครเข้ามหาวิทยาลัย"
กระรอก- สารประกอบอินทรีย์โมเลกุลสูง ประกอบด้วยกรด α-amino ตกค้าง
ที่ องค์ประกอบโปรตีนได้แก่ คาร์บอน ไฮโดรเจน ไนโตรเจน ออกซิเจน กำมะถัน โปรตีนบางชนิดสร้างสารเชิงซ้อนร่วมกับโมเลกุลอื่นๆ ที่มีฟอสฟอรัส เหล็ก สังกะสีและทองแดง
โปรตีนมีน้ำหนักโมเลกุลมาก: ไข่อัลบูมิน - 36,000, เฮโมโกลบิน - 152,000, myosin - 500,000 สำหรับการเปรียบเทียบ: น้ำหนักโมเลกุลของแอลกอฮอล์คือ 46, กรดอะซิติก - 60, เบนซิน - 78
องค์ประกอบของกรดอะมิโนของโปรตีน
กระรอก- โพลีเมอร์ที่ไม่เป็นคาบ โดยมีโมโนเมอร์เป็น กรดอะมิโน. โดยปกติ กรด α-amino 20 ชนิดจะเรียกว่าโปรตีนโมโนเมอร์ แม้ว่าจะพบมากกว่า 170 ชนิดในเซลล์และเนื้อเยื่อ
ขึ้นอยู่กับว่ากรดอะมิโนสามารถสังเคราะห์ในร่างกายของมนุษย์และสัตว์อื่น ๆ ได้หรือไม่ มี: กรดอะมิโนที่ไม่จำเป็น- สามารถสังเคราะห์ได้ กรดอะมิโนที่จำเป็น- ไม่สามารถสังเคราะห์ได้ กรดอะมิโนที่จำเป็นจะต้องกินเข้าไปพร้อมกับอาหาร พืชสังเคราะห์กรดอะมิโนทุกชนิด
ขึ้นอยู่กับองค์ประกอบของกรดอะมิโน โปรตีนคือ: สมบูรณ์- ประกอบด้วยกรดอะมิโนทั้งชุด ชำรุด- กรดอะมิโนบางชนิดไม่มีอยู่ในองค์ประกอบ ถ้าโปรตีนประกอบด้วยกรดอะมิโนเท่านั้นจะเรียกว่า เรียบง่าย. หากโปรตีนประกอบด้วยส่วนประกอบที่ไม่ใช่กรดอะมิโน (กลุ่มเทียม) นอกจากกรดอะมิโนแล้ว จะถูกเรียกว่า ซับซ้อน. กลุ่มเทียมสามารถแสดงด้วยโลหะ (metalloproteins), คาร์โบไฮเดรต (glycoproteins), ไขมัน (lipoproteins), กรดนิวคลีอิก (nucleoproteins)
ทั้งหมด กรดอะมิโนประกอบด้วย: 1) หมู่คาร์บอกซิล (-COOH), 2) หมู่อะมิโน (-NH 2), 3) หมู่เรดิคัลหรืออาร์ (ส่วนที่เหลือของโมเลกุล). โครงสร้างของอนุมูลอิสระในกรดอะมิโนชนิดต่างๆ จะแตกต่างกัน ขึ้นอยู่กับจำนวนของหมู่อะมิโนและหมู่คาร์บอกซิลที่ประกอบเป็นกรดอะมิโน มี: กรดอะมิโนที่เป็นกลางมีหมู่คาร์บอกซิลหนึ่งหมู่และหมู่อะมิโนหนึ่งหมู่ กรดอะมิโนพื้นฐานมีหมู่อะมิโนมากกว่าหนึ่งหมู่ กรดอะมิโนที่เป็นกรดมีหมู่คาร์บอกซิลมากกว่าหนึ่งกลุ่ม
กรดอะมิโนคือ สารประกอบแอมโฟเทอริกเนื่องจากในสารละลายสามารถทำหน้าที่เป็นทั้งกรดและเบสได้ ในสารละลายที่เป็นน้ำ กรดอะมิโนมีอยู่ในรูปแบบไอออนิกที่แตกต่างกัน
พันธะเปปไทด์
เปปไทด์- สารอินทรีย์ประกอบด้วยกรดอะมิโนตกค้างที่เชื่อมต่อกันด้วยพันธะเปปไทด์
การก่อตัวของเปปไทด์เกิดขึ้นจากปฏิกิริยาควบแน่นของกรดอะมิโน เมื่อหมู่อะมิโนของกรดอะมิโนหนึ่งมีปฏิสัมพันธ์กับหมู่คาร์บอกซิลของอีกกลุ่มหนึ่ง จะเกิดพันธะโควาเลนต์ไนโตรเจน-คาร์บอนระหว่างพวกมันซึ่งเรียกว่า เปปไทด์. ขึ้นอยู่กับจำนวนกรดอะมิโนที่ตกค้างเป็นเปปไทด์มี ไดเปปไทด์ ไตรเปปไทด์ เตตราเปปไทด์เป็นต้น การก่อตัวของพันธะเปปไทด์สามารถทำซ้ำได้หลายครั้ง สิ่งนี้นำไปสู่การก่อตัว โพลีเปปไทด์. ที่ปลายด้านหนึ่งของเปปไทด์จะมีหมู่อะมิโนอิสระ (เรียกว่าปลาย N) และปลายอีกด้านหนึ่งมีหมู่คาร์บอกซิลอิสระ (เรียกว่าปลาย C)

การจัดระเบียบเชิงพื้นที่ของโมเลกุลโปรตีน
ประสิทธิภาพของการทำงานเฉพาะบางอย่างของโปรตีนขึ้นอยู่กับการกำหนดค่าเชิงพื้นที่ของโมเลกุลของพวกมัน นอกจากนี้ยังไม่เอื้ออำนวยต่อเซลล์ที่จะเก็บโปรตีนในรูปแบบที่ขยายตัวในรูปแบบของสายโซ่ดังนั้นโซ่โพลีเปปไทด์จึงถูกพับเก็บ โครงสร้างสามมิติบางอย่างหรือรูปแบบ จัดสรร 4 ระดับ การจัดระเบียบเชิงพื้นที่ของโปรตีน.
โครงสร้างหลักของโปรตีน- ลำดับของกรดอะมิโนตกค้างในสายโซ่โพลีเปปไทด์ที่ประกอบเป็นโมเลกุลโปรตีน พันธะระหว่างกรดอะมิโนคือเปปไทด์

หากโมเลกุลโปรตีนประกอบด้วยกรดอะมิโนตกค้างเพียง 10 ตัว จำนวนตัวแปรที่เป็นไปได้ทางทฤษฎีของโมเลกุลโปรตีนที่แตกต่างกันตามลำดับการสลับของกรดอะมิโนคือ 10 20 . ด้วยกรดอะมิโน 20 ชนิด คุณสามารถสร้างส่วนผสมที่หลากหลายยิ่งขึ้น พบโปรตีนประมาณหนึ่งหมื่นชนิดในร่างกายมนุษย์ ซึ่งแตกต่างจากโปรตีนของสิ่งมีชีวิตอื่นๆ
เป็นโครงสร้างหลักของโมเลกุลโปรตีนที่กำหนดคุณสมบัติของโมเลกุลโปรตีนและการกำหนดค่าเชิงพื้นที่ การเปลี่ยนกรดอะมิโนเพียงตัวเดียวไปเป็นอีกตัวหนึ่งในสายโซ่โพลีเปปไทด์ทำให้เกิดการเปลี่ยนแปลงในคุณสมบัติและหน้าที่ของโปรตีน ตัวอย่างเช่นการแทนที่กรดอะมิโนกลูตามีนที่หกในβ-subunit ของเฮโมโกลบินด้วยวาลีนนำไปสู่ความจริงที่ว่าโมเลกุลเฮโมโกลบินโดยรวมไม่สามารถทำหน้าที่หลักได้ - การขนส่งออกซิเจน ในกรณีเช่นนี้บุคคลจะเป็นโรค - โรคโลหิตจางชนิดเคียว
โครงสร้างรอง- สั่งพับโซ่โพลีเปปไทด์เป็นเกลียว (ดูเหมือนสปริงยืด) ขดลวดของเกลียวนั้นเสริมความแข็งแกร่งด้วยพันธะไฮโดรเจนระหว่างหมู่คาร์บอกซิลและหมู่อะมิโน กลุ่ม CO และ NH เกือบทั้งหมดมีส่วนร่วมในการก่อตัวของพันธะไฮโดรเจน พวกมันอ่อนแอกว่าเปปไทด์ แต่การทำซ้ำหลายครั้งพวกมันให้ความเสถียรและความแข็งแกร่งแก่การกำหนดค่านี้ ที่ระดับของโครงสร้างทุติยภูมิ มีโปรตีน: ไฟโบรอิน (ไหม, ใยแมงมุม), เคราติน (ผม, เล็บ), คอลลาเจน (เอ็น)



โครงสร้างตติยภูมิ- การบรรจุสายโพลีเปปไทด์ให้เป็นลูกกลม อันเป็นผลมาจากการเกิดพันธะเคมี (ไฮโดรเจน ไอออนิก ไดซัลไฟด์) และการสร้างปฏิกิริยาที่ไม่ชอบน้ำระหว่างอนุมูลของสารตกค้างของกรดอะมิโน บทบาทหลักในการก่อตัวของโครงสร้างตติยภูมิเล่นโดยปฏิสัมพันธ์ที่ชอบน้ำและไม่ชอบน้ำ ในสารละลายที่เป็นน้ำ อนุมูลที่ไม่ชอบน้ำมักจะซ่อนตัวจากน้ำ โดยจัดกลุ่มอยู่ภายในทรงกลม ในขณะที่อนุมูลที่ชอบน้ำมักจะปรากฏบนพื้นผิวของโมเลกุลอันเป็นผลมาจากความชุ่มชื้น (ปฏิกิริยากับไดโพลน้ำ) ในโปรตีนบางชนิด โครงสร้างระดับตติยภูมิจะมีความเสถียรโดยพันธะโควาเลนต์ไดซัลไฟด์ซึ่งเกิดขึ้นระหว่างอะตอมของกำมะถันของซิสเทอีนตกค้างทั้งสอง ในระดับโครงสร้างตติยภูมิมีเอ็นไซม์ แอนติบอดี ฮอร์โมนบางชนิด

โครงสร้างควอเทอร์นารีลักษณะของโปรตีนที่ซับซ้อนซึ่งเป็นโมเลกุลที่ก่อตัวขึ้นจากสองทรงกลมขึ้นไป หน่วยย่อยถูกเก็บไว้ในโมเลกุลโดยปฏิกิริยาระหว่างไอออนิก ไม่ชอบน้ำ และไฟฟ้าสถิต บางครั้ง ระหว่างการก่อตัวของโครงสร้างควอเทอร์นารี พันธะไดซัลไฟด์เกิดขึ้นระหว่างหน่วยย่อย โปรตีนที่มีการศึกษามากที่สุดที่มีโครงสร้างควอเทอร์นารีคือ เฮโมโกลบิน. มันถูกสร้างขึ้นโดยหน่วยย่อย α สองตัว (เรซิดิวกรดอะมิโน 141 ตัว) และหน่วยย่อย β สองหน่วย (เรซิดิวของกรดอะมิโน 146 ตัว) แต่ละหน่วยย่อยมีความเกี่ยวข้องกับโมเลกุลของฮีมที่มีธาตุเหล็ก
หากโครงสร้างเชิงพื้นที่ของโปรตีนเบี่ยงเบนไปจากปกติด้วยเหตุผลบางประการ โปรตีนจะไม่สามารถทำหน้าที่ของมันได้ ตัวอย่างเช่น สาเหตุของ "โรควัวบ้า" (spongiform encephalopathy) คือโครงสร้างที่ผิดปกติของพรีออน ซึ่งเป็นโปรตีนที่ผิวเซลล์ประสาท
คุณสมบัติของโปรตีน
องค์ประกอบของกรดอะมิโน โครงสร้างของโมเลกุลโปรตีนเป็นตัวกำหนด คุณสมบัติ. โปรตีนรวมคุณสมบัติพื้นฐานและความเป็นกรดที่กำหนดโดยอนุมูลของกรดอะมิโน ยิ่งกรดอะมิโนที่เป็นกรดในโปรตีนมากเท่าใด คุณสมบัติที่เป็นกรดก็จะยิ่งเด่นชัดมากขึ้นเท่านั้น ความสามารถในการให้และแนบ H + กำหนด คุณสมบัติบัฟเฟอร์ของโปรตีน; หนึ่งในบัฟเฟอร์ที่ทรงพลังที่สุดคือเฮโมโกลบินในเม็ดเลือดแดงซึ่งรักษาค่า pH ของเลือดให้อยู่ในระดับคงที่ มีโปรตีนที่ละลายน้ำได้ (ไฟบริโนเจน) มีโปรตีนที่ไม่ละลายน้ำที่ทำหน้าที่เชิงกล (ไฟโบรอิน เคราติน คอลลาเจน) มีโปรตีนที่ออกฤทธิ์ทางเคมี (เอ็นไซม์) มีปฏิกิริยาทางเคมี ทนทานต่อสภาวะแวดล้อมต่างๆ และไม่เสถียรอย่างยิ่ง
ปัจจัยภายนอก (ความร้อน รังสีอัลตราไวโอเลต โลหะหนักและเกลือ การเปลี่ยนแปลงค่า pH การแผ่รังสี การคายน้ำ)
อาจทำให้เกิดการละเมิดโครงสร้างโครงสร้างของโมเลกุลโปรตีน กระบวนการสูญเสียโครงสร้างสามมิติที่มีอยู่ในโมเลกุลโปรตีนที่กำหนดเรียกว่า การทำให้เสียสภาพ. สาเหตุของการเสียสภาพคือการทำลายพันธะที่ทำให้โครงสร้างโปรตีนบางตัวมีเสถียรภาพ เริ่มแรก สายสัมพันธ์ที่อ่อนแอที่สุดจะขาดหายไป และเมื่อเงื่อนไขรุนแรงขึ้น ความสัมพันธ์ที่แน่นแฟ้นยิ่งขึ้น ดังนั้นขั้นแรกคือสี่ส่วนจากนั้นโครงสร้างระดับตติยภูมิและทุติยภูมิจะหายไป การเปลี่ยนแปลงโครงสร้างเชิงพื้นที่นำไปสู่การเปลี่ยนแปลงคุณสมบัติของโปรตีน และทำให้เป็นไปไม่ได้ที่โปรตีนจะทำหน้าที่ทางชีวภาพ หากการเสียสภาพไม่ได้มาพร้อมกับการทำลายโครงสร้างหลักก็สามารถทำได้ ย้อนกลับได้ในกรณีนี้จะเกิดการรักษาตัวเองของลักษณะโครงสร้างของโปรตีน การเปลี่ยนสภาพดังกล่าวขึ้นอยู่กับโปรตีนตัวรับเมมเบรน ตัวอย่างเช่น กระบวนการฟื้นฟูโครงสร้างของโปรตีนหลังจากการเสียสภาพเรียกว่า การปรับสภาพใหม่. หากการคืนค่าการกำหนดค่าเชิงพื้นที่ของโปรตีนเป็นไปไม่ได้ การเสียสภาพจะเรียกว่า กลับไม่ได้.
หน้าที่ของโปรตีน
| การทำงาน | ตัวอย่างและคำอธิบาย |
|---|---|
| การก่อสร้าง | โปรตีนมีส่วนร่วมในการก่อตัวของโครงสร้างเซลล์และโครงสร้างภายนอกเซลล์: เป็นส่วนหนึ่งของเยื่อหุ้มเซลล์ (ไลโปโปรตีน, ไกลโคโปรตีน), ผม (เคราติน), เส้นเอ็น (คอลลาเจน) เป็นต้น |
| ขนส่ง | โปรตีนเฮโมโกลบินในเลือดจับออกซิเจนและขนส่งจากปอดไปยังเนื้อเยื่อและอวัยวะทั้งหมด จากนั้นคาร์บอนไดออกไซด์จะถ่ายโอนไปยังปอด องค์ประกอบของเยื่อหุ้มเซลล์ประกอบด้วยโปรตีนพิเศษที่ให้การถ่ายโอนสารและไอออนบางอย่างจากเซลล์ไปยังสภาพแวดล้อมภายนอกและในทางกลับกัน |
| ระเบียบข้อบังคับ | ฮอร์โมนโปรตีนมีส่วนเกี่ยวข้องกับการควบคุมกระบวนการเผาผลาญอาหาร ตัวอย่างเช่น ฮอร์โมนอินซูลินควบคุมระดับน้ำตาลในเลือด ส่งเสริมการสังเคราะห์ไกลโคเจน และเพิ่มการก่อตัวของไขมันจากคาร์โบไฮเดรต |
| ป้องกัน | ในการตอบสนองต่อการแทรกซึมของโปรตีนหรือจุลินทรีย์จากต่างประเทศ (แอนติเจน) เข้าสู่ร่างกาย โปรตีนพิเศษจะถูกสร้างขึ้น - แอนติบอดีที่สามารถจับและทำให้เป็นกลางได้ ไฟบรินที่เกิดจากไฟบริโนเจนช่วยหยุดเลือด |
| เครื่องยนต์ | โปรตีนที่หดตัวแอกตินและไมโอซินทำให้กล้ามเนื้อหดตัวในสัตว์หลายเซลล์ |
| สัญญาณ | โมเลกุลของโปรตีนถูกฝังอยู่ในเยื่อหุ้มเซลล์ผิว ซึ่งสามารถเปลี่ยนแปลงโครงสร้างระดับอุดมศึกษาเพื่อตอบสนองต่อการกระทำของปัจจัยแวดล้อม จึงรับสัญญาณจากสภาพแวดล้อมภายนอกและส่งคำสั่งไปยังเซลล์ |
| จอง | ในร่างกายของสัตว์โปรตีนจะไม่ถูกเก็บไว้ยกเว้นไข่อัลบูมินเคซีนนม แต่ต้องขอบคุณโปรตีนในร่างกาย สารบางชนิดสามารถเก็บไว้สำรองได้ ตัวอย่างเช่น ในระหว่างการสลายของฮีโมโกลบิน ธาตุเหล็กจะไม่ถูกขับออกจากร่างกาย แต่ถูกเก็บไว้ ก่อตัวเป็นโปรตีนที่ซับซ้อนด้วยโปรตีนเฟอร์ริติน |
| พลังงาน | ด้วยการสลายโปรตีน 1 กรัมไปยังผลิตภัณฑ์ขั้นสุดท้าย 17.6 kJ จะถูกปล่อยออกมา อย่างแรก โปรตีนแตกตัวเป็นกรดอะมิโน และจากนั้นไปยังผลิตภัณฑ์สุดท้าย - น้ำ คาร์บอนไดออกไซด์ และแอมโมเนีย อย่างไรก็ตาม โปรตีนถูกใช้เป็นแหล่งพลังงานก็ต่อเมื่อมีการใช้แหล่งอื่น (คาร์โบไฮเดรตและไขมัน) หมด |
| ตัวเร่งปฏิกิริยา | หนึ่งในหน้าที่ที่สำคัญที่สุดของโปรตีน ให้โปรตีน - เอ็นไซม์ที่เร่งปฏิกิริยาทางชีวเคมีที่เกิดขึ้นในเซลล์ ตัวอย่างเช่น ไรบูโลสไบฟอสเฟตคาร์บอกซิเลสกระตุ้นการตรึง CO2 ระหว่างการสังเคราะห์ด้วยแสง |
เอนไซม์
เอนไซม์, หรือ เอนไซม์เป็นโปรตีนประเภทพิเศษที่เป็นตัวเร่งปฏิกิริยาทางชีวภาพ ต้องขอบคุณเอนไซม์ ปฏิกิริยาทางชีวเคมีดำเนินไปอย่างรวดเร็วมาก อัตราการเกิดปฏิกิริยาของเอนไซม์สูงกว่าอัตราของปฏิกิริยาที่เกี่ยวข้องกับตัวเร่งปฏิกิริยาอนินทรีย์หลายหมื่นเท่า (และบางครั้งก็เป็นล้าน) สารที่เอนไซม์ทำหน้าที่เรียกว่า พื้นผิว.
เอ็นไซม์เป็นโปรตีนทรงกลม ลักษณะโครงสร้างเอนไซม์สามารถแบ่งออกเป็นสองกลุ่ม: ง่ายและซับซ้อน เอนไซม์อย่างง่ายเป็นโปรตีนอย่างง่าย กล่าวคือ ประกอบด้วยกรดอะมิโนเท่านั้น เอนไซม์ที่ซับซ้อนเป็นโปรตีนเชิงซ้อน กล่าวคือ นอกจากส่วนของโปรตีนแล้ว ยังรวมถึงกลุ่มของธรรมชาติที่ไม่ใช่โปรตีนด้วย - ปัจจัยร่วม. สำหรับเอนไซม์บางชนิด วิตามินจะทำหน้าที่เป็นปัจจัยร่วม ในโมเลกุลของเอนไซม์จะแยกส่วนพิเศษที่เรียกว่าศูนย์แอคทีฟ แอคทีฟเซ็นเตอร์- ส่วนเล็ก ๆ ของเอนไซม์ (จากสามถึงสิบสองของกรดอะมิโนตกค้าง) ซึ่งการจับกันของสารตั้งต้นหรือสารตั้งต้นเกิดขึ้นกับการก่อตัวของสารตั้งต้นของเอนไซม์ - สารตั้งต้น เมื่อเสร็จสิ้นปฏิกิริยา สารเชิงซ้อนของเอนไซม์-สารตั้งต้นจะสลายตัวเป็นเอนไซม์และผลิตภัณฑ์จากปฏิกิริยา เอนไซม์บางชนิดมี (นอกเหนือจากแอกทีฟ) ศูนย์อัลลอสเตอร์- ไซต์ที่แนบตัวควบคุมอัตราการทำงานของเอนไซม์ ( เอนไซม์อัลโลสเตอริก).

ปฏิกิริยาเร่งปฏิกิริยาด้วยเอนไซม์มีลักษณะดังนี้: 1) ประสิทธิภาพสูง 2) หัวกะทิและทิศทางของการกระทำที่เข้มงวด 3) ความจำเพาะของพื้นผิว 4) การควบคุมที่ละเอียดและแม่นยำ สารตั้งต้นและความจำเพาะของปฏิกิริยาของปฏิกิริยาเร่งปฏิกิริยาด้วยเอนไซม์อธิบายโดยสมมติฐานของ E. Fischer (1890) และ D. Koshland (1959)
E. ฟิชเชอร์ (สมมติฐานกุญแจล็อค)แนะนำว่าการกำหนดค่าเชิงพื้นที่ของไซต์แอคทีฟของเอนไซม์และสารตั้งต้นควรสอดคล้องกันทุกประการ พื้นผิวถูกเปรียบเทียบกับ "กุญแจ" เอนไซม์ - กับ "ล็อค"
D. Koshland (สมมติฐาน "ถุงมือมือ")เสนอว่าการโต้ตอบเชิงพื้นที่ระหว่างโครงสร้างของสารตั้งต้นและจุดศูนย์กลางของเอนไซม์นั้นถูกสร้างขึ้นในขณะที่มีปฏิสัมพันธ์ซึ่งกันและกันเท่านั้น สมมติฐานนี้เรียกอีกอย่างว่า สมมุติฐานที่เหนี่ยวนำให้เกิดความพอดี.
อัตราการเกิดปฏิกิริยาของเอนไซม์ขึ้นอยู่กับ: 1) อุณหภูมิ 2) ความเข้มข้นของเอนไซม์ 3) ความเข้มข้นของสารตั้งต้น 4) pH ควรเน้นว่าเนื่องจากเอนไซม์เป็นโปรตีน กิจกรรมของเอนไซม์จึงสูงที่สุดภายใต้สภาวะปกติทางสรีรวิทยา
เอนไซม์ส่วนใหญ่สามารถทำงานได้ที่อุณหภูมิระหว่าง 0 ถึง 40°C เท่านั้น ภายในขีดจำกัดเหล่านี้ อัตราการเกิดปฏิกิริยาจะเพิ่มขึ้นประมาณ 2 เท่าสำหรับทุกๆ 10 °C ที่เพิ่มขึ้นของอุณหภูมิ ที่อุณหภูมิสูงกว่า 40 °C โปรตีนจะผ่านการเปลี่ยนสภาพและการทำงานของเอนไซม์จะลดลง ที่อุณหภูมิใกล้เคียงกับจุดเยือกแข็ง เอ็นไซม์จะหยุดทำงาน
เมื่อปริมาณของสารตั้งต้นเพิ่มขึ้น อัตราของปฏิกิริยาของเอนไซม์จะเพิ่มขึ้นจนกว่าจำนวนโมเลกุลของสารตั้งต้นจะเท่ากับจำนวนโมเลกุลของเอนไซม์ เมื่อปริมาณสารตั้งต้นเพิ่มขึ้นอีก อัตราจะไม่เพิ่มขึ้น เนื่องจากบริเวณที่ทำงานของเอนไซม์อิ่มตัว การเพิ่มขึ้นของความเข้มข้นของเอนไซม์ทำให้กิจกรรมเร่งปฏิกิริยาเพิ่มขึ้น เนื่องจากโมเลกุลของสารตั้งต้นจำนวนมากขึ้นได้รับการแปลงสภาพต่อหน่วยเวลา

สำหรับเอนไซม์แต่ละตัว มีค่า pH ที่เหมาะสมที่สุดซึ่งแสดงกิจกรรมสูงสุด (เปปซิน - 2.0, อะไมเลสน้ำลาย - 6.8, ไลเปสตับอ่อน - 9.0) ที่ค่า pH สูงหรือต่ำ กิจกรรมของเอนไซม์จะลดลง ด้วยการเปลี่ยนแปลงอย่างรวดเร็วของค่า pH เอนไซม์จะเสื่อมสภาพ
ความเร็วของเอนไซม์ allosteric ถูกควบคุมโดยสารที่ยึดติดกับศูนย์ allosteric ถ้าสารเหล่านี้เร่งปฏิกิริยาจะเรียกว่า ตัวกระตุ้นถ้าพวกเขาช้าลง - สารยับยั้ง.
การจำแนกเอนไซม์
ตามประเภทของการเปลี่ยนแปลงทางเคมีที่เร่งปฏิกิริยา เอ็นไซม์แบ่งออกเป็น 6 คลาส:
- ออกซิโดเรดักเตส(การถ่ายโอนอะตอมไฮโดรเจนออกซิเจนหรืออิเล็กตรอนจากสารหนึ่งไปยังอีกสารหนึ่ง - ดีไฮโดรจีเนส)
- โอน(การถ่ายโอนกลุ่มเมธิล, เอซิล, ฟอสเฟตหรืออะมิโนจากสารหนึ่งไปยังอีกสารหนึ่ง - ทรานสอะมิเนส)
- ไฮโดรเลส(ปฏิกิริยาไฮโดรไลซิสซึ่งผลิตภัณฑ์สองชนิดเกิดขึ้นจากสารตั้งต้น - อะไมเลส, ไลเปส)
- lyases(นอกเหนือจากไฮโดรไลติกนอกเหนือจากสารตั้งต้นหรือการกำจัดกลุ่มอะตอมจากนั้นในขณะที่พันธะ C-C, C-N, C-O, C-S สามารถแตกได้ - decarboxylase)
- ไอโซเมอเรส(การจัดเรียงภายในโมเลกุล - ไอโซเมอเรส),
- ligases(การเชื่อมต่อของสองโมเลกุลอันเป็นผลมาจากการก่อตัวของพันธะ C-C, C-N, C-O, C-S - synthetase)
คลาสจะถูกแบ่งออกเป็นคลาสย่อยและคลาสย่อย ในการจำแนกระหว่างประเทศในปัจจุบัน เอนไซม์แต่ละตัวมีรหัสเฉพาะ ซึ่งประกอบด้วยตัวเลขสี่ตัวคั่นด้วยจุด หมายเลขแรกคือคลาส หมายเลขที่สองคือคลาสย่อย หมายเลขที่สามคือคลาสย่อย หมายเลขที่สี่คือหมายเลขซีเรียลของเอนไซม์ในคลาสย่อยนี้ ตัวอย่างเช่น รหัสอาร์จิเนสคือ 3.5.3.1
ไปที่ การบรรยายครั้งที่ 2"โครงสร้างและหน้าที่ของคาร์โบไฮเดรตและไขมัน"
ไปที่ การบรรยาย№4"โครงสร้างและหน้าที่ของกรดนิวคลีอิกเอทีพี"
ส่งงานที่ดีของคุณในฐานความรู้เป็นเรื่องง่าย ใช้แบบฟอร์มด้านล่าง
นักศึกษา นักศึกษาระดับบัณฑิตศึกษา นักวิทยาศาสตร์รุ่นเยาว์ที่ใช้ฐานความรู้ในการศึกษาและการทำงานจะขอบคุณอย่างยิ่ง
โพสต์เมื่อ http://www.allbest.ru
บทที่ 1 บทนำ
รายงานการปฏิวัติทางชีววิทยาตอนนี้ค่อนข้างซ้ำซากจำเจ นอกจากนี้ยังถือว่าเถียงไม่ได้ว่าการเปลี่ยนแปลงเชิงปฏิวัติเหล่านี้เกี่ยวข้องกับการก่อตัวของวิทยาศาสตร์ที่ซับซ้อนที่จุดตัดของชีววิทยาและเคมี ซึ่งในจำนวนนี้อณูชีววิทยาและเคมีชีวภาพถูกครอบครองและยังคงครองตำแหน่งศูนย์กลางต่อไป
“อณูชีววิทยาเป็นวิทยาศาสตร์ที่มีจุดมุ่งหมายเพื่อทำความเข้าใจธรรมชาติของปรากฏการณ์ชีวิตโดยการศึกษาวัตถุและระบบทางชีววิทยาในระดับที่เข้าใกล้โมเลกุลหนึ่ง ... ลักษณะที่ปรากฏของชีวิต ... เกิดจากโครงสร้างคุณสมบัติและปฏิสัมพันธ์ของโมเลกุลของ สารที่มีความสำคัญทางชีววิทยาเป็นหลัก โปรตีนและกรดนิวคลีอิก ”
“เคมีชีวภาพเป็นวิทยาศาสตร์ที่ศึกษาสารที่อยู่ภายใต้กระบวนการของชีวิต ... วัตถุหลักของเคมีชีวภาพคือไบโอโพลีเมอร์ (โปรตีนและเปปไทด์ กรดนิวคลีอิกและนิวคลีโอไทด์ ลิปิด โพลีแซคคาไรด์ ฯลฯ)
จากการเปรียบเทียบนี้ จะเห็นได้ชัดว่าการศึกษาโปรตีนมีความสำคัญต่อการพัฒนาชีววิทยาสมัยใหม่อย่างไร
ชีววิทยา โปรตีน ชีวเคมี
บทที่ 2 ประวัติการวิจัยโปรตีน
2.1 ระยะเริ่มต้นของเคมีโปรตีน
โปรตีนเป็นหนึ่งในเป้าหมายของการวิจัยทางเคมีเมื่อ 250 ปีที่แล้ว ในปี ค.ศ. 1728 นักวิทยาศาสตร์ชาวอิตาลี Jacopo Bartolomeo Beccari ได้รับการเตรียมโปรตีนชนิดแรกคือกลูเตนจากแป้งสาลี เขานำกลูเตนไปกลั่นแบบแห้งและทำให้แน่ใจว่าผลิตภัณฑ์ของการกลั่นนี้เป็นด่าง นี่เป็นข้อพิสูจน์ครั้งแรกของความเป็นหนึ่งเดียวกันของธรรมชาติของสสารของอาณาจักรพืชและสัตว์ เขาตีพิมพ์ผลงานของเขาในปี ค.ศ. 1745 และนี่เป็นเอกสารฉบับแรกเกี่ยวกับโปรตีน
ใน XVIII - ต้นศตวรรษที่ XIX มีการอธิบายซ้ำ ๆ เกี่ยวกับสารโปรตีนจากพืชและสัตว์ คุณลักษณะของคำอธิบายดังกล่าวคือการบรรจบกันของสารเหล่านี้และการเปรียบเทียบกับสารอนินทรีย์
เป็นสิ่งสำคัญที่จะต้องทราบว่าในขณะนั้น แม้กระทั่งก่อนการมาถึงของการวิเคราะห์ธาตุ มีความคิดว่าโปรตีนจากแหล่งต่างๆ เป็นกลุ่มของสารแต่ละชนิดที่มีคุณสมบัติใกล้เคียงกัน
ในปี ค.ศ. 1810 J. Gay-Lussac และ L. Tenard ได้กำหนดองค์ประกอบพื้นฐานของสารโปรตีน ในปี 1833 J. Gay-Lussac ได้พิสูจน์ว่าจำเป็นต้องมีไนโตรเจนในโปรตีน และในไม่ช้าก็พบว่าปริมาณไนโตรเจนในโปรตีนที่แตกต่างกันนั้นใกล้เคียงกัน ในเวลาเดียวกัน นักเคมีชาวอังกฤษ ดี. ดาลตัน พยายามอธิบายสูตรแรกของสารโปรตีน เขาแสดงพวกมันว่าเป็นสารที่ค่อนข้างง่าย แต่เพื่อเน้นความแตกต่างของแต่ละคนด้วยองค์ประกอบเดียวกัน เขาจึงใช้วิธีวาดภาพโมเลกุลที่ตอนนี้เรียกว่าไอโซเมอร์ อย่างไรก็ตาม แนวคิดเรื่อง isomerism ยังไม่มีอยู่ในสมัยของดาลตัน
สูตรโปรตีนโดย D. Dalton
ได้มาจากสูตรเชิงประจักษ์สูตรแรกของโปรตีนและมีการเสนอสมมติฐานแรกเกี่ยวกับความสม่ำเสมอขององค์ประกอบ ดังนั้น N. Lieberkün เชื่อว่าอัลบูมินอธิบายโดยสูตร C 72 H 112 N 18 SO 22 และ A. Danilevsky เชื่อว่าโมเลกุลของโปรตีนนี้อย่างน้อยต้องมีขนาดใหญ่กว่า: C 726 H 1171 N 194 S 3 โอ 214
นักเคมีชาวเยอรมัน J. Liebig ในปี 1841 แนะนำว่าโปรตีนจากสัตว์มีความคล้ายคลึงกันระหว่างโปรตีนจากพืช: การดูดซึมโปรตีนจากพืชตระกูลถั่วในร่างกายของสัตว์ตาม Liebig นำไปสู่การสะสมของโปรตีนที่คล้ายกัน - เคซีน หนึ่งในทฤษฎีที่แพร่หลายที่สุดเกี่ยวกับเคมีอินทรีย์ก่อนโครงสร้างคือทฤษฎีของอนุมูลซึ่งเป็นส่วนประกอบที่ไม่เปลี่ยนแปลงของสารที่เกี่ยวข้อง ในปี ค.ศ. 1836 ชาวดัตช์ จี. มัลเดอร์ เสนอว่าโปรตีนทั้งหมดมีอนุมูลเหมือนกัน ซึ่งเขาเรียกว่า โปรตีน (จากคำภาษากรีก "ฉันเป็นผู้นำ", "ฉันเป็นผู้นำ") โปรตีนตาม Mulder มีองค์ประกอบ Pr = C 40 H 62 N 10 O 12 ในปี ค.ศ. 1838 G. Mulder ได้เผยแพร่สูตรโปรตีนตามทฤษฎีโปรตีน สิ่งเหล่านี้คือสิ่งที่เรียกว่า สูตร dualistic โดยที่โปรตีน Radical ทำหน้าที่เป็นกลุ่มเชิงบวก และอะตอมของกำมะถันหรือฟอสฟอรัสเป็นค่าลบ พวกเขาช่วยกันสร้างโมเลกุลที่เป็นกลางทางไฟฟ้า: โปรตีนในซีรัมในเลือด Pr 10 S 2 P, ไฟบริน Pr 10 SP อย่างไรก็ตาม การตรวจสอบวิเคราะห์ข้อมูลของ G. Mulder ดำเนินการโดย Lyaskovskii นักเคมีชาวรัสเซีย และ Yu. Liebig พบว่า "อนุมูลโปรตีน" ไม่มีอยู่จริง
ในปี ค.ศ. 1833 นักวิทยาศาสตร์ชาวเยอรมัน เอฟ. โรส ค้นพบปฏิกิริยาไบยูเรตสำหรับโปรตีน ซึ่งเป็นหนึ่งในปฏิกิริยาสีหลักสำหรับสารโปรตีนและอนุพันธ์ของโปรตีนในปัจจุบัน (เพิ่มเติมเกี่ยวกับปฏิกิริยาสีในหน้า 53) นอกจากนี้ยังสรุปได้ว่านี่เป็นปฏิกิริยาที่ละเอียดอ่อนที่สุดสำหรับโปรตีน ดังนั้นจึงดึงดูดความสนใจมากที่สุดจากนักเคมีในขณะนั้น
ในช่วงกลางของศตวรรษที่ 19 มีการพัฒนาวิธีการมากมายในการสกัดโปรตีน ทำให้บริสุทธิ์ และแยกโปรตีนออกจากสารละลายของเกลือที่เป็นกลาง ในปี ค.ศ. 1847 K. Reichert ได้ค้นพบความสามารถของโปรตีนในการสร้างผลึก ในปี พ.ศ. 2379 ที. ชวานน์ได้ค้นพบเปปซิน ซึ่งเป็นเอนไซม์ที่สลายโปรตีน ในปี ค.ศ. 1856 แอล. คอร์วิซาร์ได้ค้นพบเอนไซม์ที่คล้ายคลึงกันอีกชนิดหนึ่งคือทริปซิน โดยการศึกษาการทำงานของเอนไซม์เหล่านี้กับโปรตีน นักชีวเคมีพยายามไขปริศนาของการย่อยอาหาร อย่างไรก็ตาม สารที่เกิดจากการกระทำของเอ็นไซม์โปรตีเอส (โปรตีเอสซึ่งรวมถึงเอ็นไซม์ข้างต้น) บนโปรตีนได้รับความสนใจมากที่สุด: บางส่วนเป็นชิ้นส่วนของโมเลกุลโปรตีนดั้งเดิม (เรียกว่า เปปโตน ) ในขณะที่คนอื่นไม่ได้อยู่ภายใต้การแตกแยกเพิ่มเติมโดยโปรตีเอสและอยู่ในกลุ่มของสารประกอบที่รู้จักกันตั้งแต่ต้นศตวรรษ - กรดอะมิโน (อนุพันธ์ของกรดอะมิโนตัวแรก, แอสพาราจีนเอไมด์, ถูกค้นพบในปี 1806 และกรดอะมิโนตัวแรก ซีสทีนในปี พ.ศ. 2353) กรดอะมิโนในองค์ประกอบของโปรตีนถูกค้นพบครั้งแรกในปี พ.ศ. 2363 โดยนักเคมีชาวฝรั่งเศส A. Braconno เขาใช้กรดไฮโดรไลซิสของโปรตีนและพบสารที่มีรสหวานในไฮโดรไลเสต ซึ่งเขาเรียกว่าไกลซีน ในปีพ.ศ. 2382 การมีอยู่ของลิวซีนในโปรตีนได้รับการพิสูจน์แล้ว และในปี พ.ศ. 2392 เอฟ. บอปป์ ได้แยกกรดอะมิโนอีกชนิดหนึ่งออกจากโปรตีน - ไทโรซีน (ดูภาคผนวก II สำหรับรายการวันที่ค้นพบกรดอะมิโนในโปรตีนทั้งหมด)
ในช่วงปลายยุค 80 ในศตวรรษที่ 19 กรดอะมิโน 19 ตัวถูกแยกออกจากโปรตีนไฮโดรไลเสต และความคิดเห็นเริ่มแข็งแกร่งขึ้นอย่างช้าๆ ว่าข้อมูลเกี่ยวกับผลิตภัณฑ์ของการไฮโดรไลซิสของโปรตีนมีข้อมูลสำคัญเกี่ยวกับโครงสร้างของโมเลกุลโปรตีน อย่างไรก็ตาม กรดอะมิโนถือว่าจำเป็น แต่ไม่ใช่ส่วนประกอบหลักของโปรตีน
ในการเชื่อมต่อกับการค้นพบกรดอะมิโนในองค์ประกอบของโปรตีน นักวิทยาศาสตร์ชาวฝรั่งเศส P. Schutzenberger ในยุค 70 ศตวรรษที่ XIX เสนอสิ่งที่เรียกว่า ทฤษฎียูไรด์ โครงสร้างโปรตีน ตามที่โมเลกุลโปรตีนประกอบด้วยแกนกลางซึ่งมีบทบาทโดยโมเลกุลไทโรซีนและกลุ่มที่ซับซ้อนที่ติดอยู่ (ด้วยการแทนที่ของอะตอมไฮโดรเจน 4 อะตอม) เรียกว่า Schutzenberger ลิวซีน . อย่างไรก็ตาม สมมติฐานนี้ได้รับการสนับสนุนเพียงเล็กน้อยจากการทดลอง และการวิจัยเพิ่มเติมได้รับการพิสูจน์ว่าไม่สอดคล้องกัน
2.2 ทฤษฎี “สารประกอบเชิงซ้อนคาร์บอน-ไนโตรเจน” อ.ย. Danilevsky
ทฤษฎีดั้งเดิมเกี่ยวกับโครงสร้างของโปรตีนถูกแสดงออกมาในยุค 80 นักชีวเคมีชาวรัสเซียในศตวรรษที่ XIX A. Ya. Danilevsky เขาเป็นนักเคมีคนแรกที่ให้ความสนใจต่อลักษณะพอลิเมอร์ที่เป็นไปได้ของโครงสร้างของโมเลกุลโปรตีน ในช่วงต้นยุค 70 เขาเขียนถึง A.M. Butlerov ว่า "อนุภาคอัลบูมินเป็นพอลิเมอร์ผสม" ซึ่งสำหรับคำจำกัดความของโปรตีน เขาไม่พบ "คำที่เหมาะสมกว่าคำว่าพอลิเมอร์ในความหมายกว้าง" จากการศึกษาปฏิกิริยาไบยูเรต เขาแนะนำว่าปฏิกิริยานี้เกี่ยวข้องกับโครงสร้างของอะตอมของคาร์บอนและไนโตรเจนที่ไม่ต่อเนื่อง - N - C - N - C - N - ซึ่งรวมอยู่ในสิ่งที่เรียกว่า คาร์โบนาโซ t ซับซ้อน R "- NH - CO - NH - CO - R" จากสูตรนี้ Danilevsky เชื่อว่าโมเลกุลโปรตีนประกอบด้วยสารประกอบเชิงซ้อนของคาร์บอน - ไนโตรเจนจำนวน 40 ชนิด สารประกอบเชิงซ้อนของกรดอะมิโนคาร์บอน - ไนโตรเจนตาม Danilevsky มีลักษณะดังนี้:
จากข้อมูลของ Danilevsky สารประกอบเชิงซ้อนของคาร์บอน - ไนโตรเจนสามารถเชื่อมต่อกันด้วยพันธะอีเทอร์หรือเอไมด์เพื่อสร้างโครงสร้างโมเลกุลสูง
2.3 ทฤษฎี “คิริน” ก. Kossel
นักสรีรวิทยาและนักชีวเคมีชาวเยอรมัน A. Kossel ศึกษาโปรตามีนและฮิสโตน ซึ่งเป็นโปรตีนที่ค่อนข้างง่าย พบว่าอาร์จินีนจำนวนมากก่อตัวขึ้นในระหว่างการไฮโดรไลซิส นอกจากนี้ เขายังค้นพบในองค์ประกอบของไฮโดรไลเสตกรดอะมิโนที่ไม่รู้จักในขณะนั้น - ฮิสติดีน จากสิ่งนี้ Kossel แนะนำว่าสารโปรตีนเหล่านี้ถือได้ว่าเป็นแบบจำลองที่ง่ายที่สุดของโปรตีนที่ซับซ้อนมากขึ้นซึ่งสร้างขึ้นในความเห็นของเขาตามหลักการต่อไปนี้: อาร์จินีนและฮิสทิดีนก่อตัวเป็นแกนกลาง ("แกนโปรตามีน") ซึ่งล้อมรอบด้วยสารเชิงซ้อนของกรดอะมิโนอื่นๆ
ทฤษฎีของ Kossel เป็นตัวอย่างที่สมบูรณ์แบบที่สุดสำหรับการพัฒนาสมมติฐานของโครงสร้างที่กระจัดกระจายของโปรตีน (เสนอครั้งแรกตามที่กล่าวไว้ข้างต้นโดย G. Mulder) สมมติฐานนี้ถูกใช้โดยนักเคมีชาวเยอรมัน M. Siegfried เมื่อต้นศตวรรษที่ 20 เขาเชื่อว่าโปรตีนถูกสร้างขึ้นจากสารเชิงซ้อนของกรดอะมิโน (อาร์จินีน + ไลซีน + กรดกลูตามีน) ซึ่งเขาเรียกว่า คิรินามิ (จากภาษากรีก "kyrios" พื้นฐาน) อย่างไรก็ตาม สมมติฐานนี้ถูกหยิบยกขึ้นในปี 1903 เมื่อ E. Fisher กำลังพัฒนาของเขาอย่างแข็งขัน ทฤษฎีเปปไทด์ ซึ่งเป็นกุญแจไขความลึกลับของโครงสร้างของโปรตีน
2.4 ทฤษฎีเปปไทด์ E. ฟิชเชอร์
นักเคมีชาวเยอรมัน Emil Fischer ซึ่งโด่งดังไปทั่วโลกในด้านการศึกษาสารประกอบ purine (อัลคาลอยด์ของกลุ่มคาเฟอีน) และถอดรหัสโครงสร้างของน้ำตาลสร้างทฤษฎีเปปไทด์ซึ่งส่วนใหญ่ได้รับการยืนยันในทางปฏิบัติและได้รับการยอมรับในระดับสากลในช่วงชีวิตของเขา ซึ่งเขาได้รับรางวัลโนเบลที่สองในประวัติศาสตร์เคมี รางวัล (รางวัลแรกได้รับโดย Ya.G. Van't Hoff)
เป็นสิ่งสำคัญที่ฟิชเชอร์สร้างแผนการวิจัยที่แตกต่างจากที่เคยทำมาก่อนอย่างมาก แต่คำนึงถึงข้อเท็จจริงทั้งหมดที่ทราบในขณะนั้น ประการแรก เขายอมรับว่าเป็นสมมติฐานที่เป็นไปได้มากที่สุดว่าโปรตีนถูกสร้างขึ้นจากกรดอะมิโนที่เชื่อมต่อกันด้วยพันธะเอไมด์:
ฟิชเชอร์เรียกพันธะประเภทนี้ (โดยการเปรียบเทียบกับเปปโตน) เปปไทด์ . เขาแนะนำว่าโปรตีนคือ โพลีเมอร์ของกรดอะมิโนที่เชื่อมโยงกันด้วยพันธะเปปไทด์ . Danilevsky และ Hert แสดงออกถึงความคิดเกี่ยวกับธรรมชาติของพอลิเมอร์ของโครงสร้างของโปรตีน แต่พวกเขาเชื่อว่า "โมโนเมอร์" เป็นรูปแบบที่ซับซ้อนมาก - เปปโทนหรือ "สารประกอบเชิงซ้อนของคาร์บอน - ไนโตรเจน"
พิสูจน์ชนิดเปปไทด์ของสารประกอบของกรดอะมิโนตกค้าง E. ฟิชเชอร์ดำเนินการจากการสังเกตต่อไปนี้ อย่างแรก ทั้งในระหว่างการไฮโดรไลซิสของโปรตีนและระหว่างการสลายตัวของเอนไซม์ กรดอะมิโนต่างๆ ได้ถูกสร้างขึ้น สารประกอบอื่น ๆ นั้นอธิบายได้ยากมากและยิ่งหาได้ยากยิ่ง นอกจากนี้ ฟิสเชอร์รู้ว่าโปรตีนไม่ได้มีคุณสมบัติเด่นของคุณสมบัติที่เป็นกรดหรือพื้นฐานซึ่งหมายความว่าเขาโต้แย้งกลุ่มอะมิโนและคาร์บอกซิลในองค์ประกอบของกรดอะมิโนในโมเลกุลโปรตีนถูกปิดและเหมือนเดิม ( amphotericity ของโปรตีน อย่างที่พวกเขาพูดตอนนี้ )
ฟิชเชอร์ได้แบ่งการแก้ปัญหาของโครงสร้างโปรตีนออกเป็นบทบัญญัติดังต่อไปนี้:
การกำหนดคุณภาพและเชิงปริมาณของผลิตภัณฑ์จากการไฮโดรไลซิสที่สมบูรณ์ของโปรตีน
การสร้างโครงสร้างของผลิตภัณฑ์ขั้นสุดท้ายเหล่านี้
การสังเคราะห์พอลิเมอร์กรดอะมิโนด้วยสารประกอบประเภทเอไมด์ (เปปไทด์)
การเปรียบเทียบสารประกอบที่ได้จากโปรตีนธรรมชาติ
จากแผนนี้ จะเห็นได้ว่าฟิชเชอร์ใช้วิธีการใหม่นี้เป็นครั้งแรก - การสังเคราะห์สารประกอบแบบจำลอง เป็นวิธีพิสูจน์โดยการเปรียบเทียบ
2.5 การพัฒนาวิธีการสังเคราะห์กรดอะมิโน
เพื่อดำเนินการสังเคราะห์อนุพันธ์ของกรดอะมิโนที่เชื่อมต่อกันด้วยพันธะเปปไทด์ ฟิสเชอร์ได้ทำงานอย่างมากในการศึกษาโครงสร้างและการสังเคราะห์กรดอะมิโน
ก่อนฟิสเชอร์ วิธีการทั่วไปในการสังเคราะห์กรดอะมิโนคือการสังเคราะห์ไซยาโนไฮดรินของ A. Strecker:
ตามปฏิกิริยาของ Strecker เป็นไปได้ที่จะสังเคราะห์อะลานีน ซีรีน และกรดอะมิโนอื่น ๆ และจากการดัดแปลง (ปฏิกิริยา Zelinsky-Stadnikov) ทั้งกรดอะมิโนและกรดแทน N ของพวกมัน
อย่างไรก็ตาม ฟิสเชอร์เองก็พยายามพัฒนาวิธีการสังเคราะห์กรดอะมิโนที่รู้จักกันทั้งหมดในขณะนั้น เขาถือว่าวิธีการของ Strecker นั้นไม่เป็นสากลเพียงพอ ดังนั้น อี. ฟิสเชอร์จึงต้องมองหาวิธีการทั่วไปในการสังเคราะห์กรดอะมิโน ซึ่งรวมถึงกรดอะมิโนที่มีสารอนุมูลอิสระเชิงซ้อน
เขาเสนอให้แอมิเนตกรดคาร์บอกซิลิกที่ถูกแทนที่โบรโมในตำแหน่ง - เพื่อให้ได้อนุพันธ์โบรโมเขาใช้ตัวอย่างเช่นในการสังเคราะห์กรดลิวซีนกรดอะริเลตหรืออัลคิลเลตมาโลนิก:
แต่อี. ฟิชเชอร์ล้มเหลวในการสร้างวิธีการที่เป็นสากลอย่างแท้จริง นอกจากนี้ยังมีการพัฒนาปฏิกิริยาที่น่าเชื่อถือมากขึ้น ตัวอย่างเช่น นักศึกษาของฟิชเชอร์ G. Lakes เสนอการปรับเปลี่ยนต่อไปนี้เพื่อให้ได้ซีรีน:
ฟิชเชอร์ยังพิสูจน์ด้วยว่าโปรตีนประกอบด้วยเรซิดิวกรดอะมิโนที่ออกฤทธิ์ทางแสง (ดูหน้า 11) สิ่งนี้ทำให้เขาต้องพัฒนาระบบการตั้งชื่อใหม่ของสารประกอบเชิงแสง วิธีการแยกและการสังเคราะห์ออปติคัลไอโซเมอร์ของกรดอะมิโน ฟิชเชอร์ยังได้ข้อสรุปว่าโปรตีนประกอบด้วยสารตกค้างในรูปแบบ L ของกรดอะมิโนที่ออกฤทธิ์ทางแสง และเขาได้พิสูจน์สิ่งนี้โดยใช้หลักการไดแอสเตอรีโอไอโซเมอร์ริซึมก่อน หลักการนี้มีดังต่อไปนี้: แอลคาลอยด์ที่ออกฤทธิ์ทางสายตา (บรูซีน สตริกนิน ซินโชนีน ควินิดีน และควินิน) ถูกเติมลงในอนุพันธ์ของ N-acyl ของกรดอะมิโนราซิมิก เป็นผลให้เกิดเกลือในรูปแบบสเตอริโอไอโซเมอร์สองรูปแบบที่มีความสามารถในการละลายต่างกัน หลังจากแยกไดแอสเทอรีโอไอโซเมอร์เหล่านี้ออก อัลคาลอยด์ก็ถูกนำกลับคืนมาและกลุ่มอะซิลถูกกำจัดออกโดยการไฮโดรไลซิส
ฟิสเชอร์สามารถพัฒนาวิธีการตรวจวัดกรดอะมิโนอย่างสมบูรณ์ในผลิตภัณฑ์ของการไฮโดรไลซิสของโปรตีน: เขาเปลี่ยนเอสเทอร์ไฮโดรคลอไรด์ของกรดอะมิโนโดยการบำบัดด้วยด่างเข้มข้นในความเย็นให้เป็นเอสเทอร์อิสระ ซึ่งไม่สามารถประเมินค่าได้ว่าเป็นซาโปนิฟิเคชัน จากนั้นของผสมของอีเทอร์เหล่านี้ถูกกลั่นแบบเศษส่วนและกรดอะมิโนแต่ละตัวถูกแยกจากเศษส่วนที่เป็นผลลัพธ์โดยการตกผลึกแบบเศษส่วน
วิธีการวิเคราะห์แบบใหม่นี้ไม่เพียงแต่ยืนยันว่าโปรตีนประกอบด้วยสารตกค้างของกรดอะมิโนเท่านั้น แต่ยังทำให้สามารถกลั่นและเสริมรายการกรดอะมิโนที่พบในโปรตีนได้อีกด้วย แต่ถึงกระนั้น การวิเคราะห์เชิงปริมาณก็ไม่สามารถตอบคำถามหลักได้ว่า หลักการของโครงสร้างของโมเลกุลโปรตีนคืออะไร และอี. ฟิชเชอร์ได้กำหนดหนึ่งในภารกิจหลักในการศึกษาโครงสร้างและคุณสมบัติของโปรตีน: การพัฒนา ทดลอง mอีวิธีการสังเคราะห์สารประกอบที่มีส่วนประกอบหลักคือกรดอะมิโนเกี่ยวกับคุณเชื่อมต่อด้วยพันธะเปปไทด์
ดังนั้น ฟิชเชอร์จึงกำหนดภารกิจที่ไม่สำคัญ - เพื่อสังเคราะห์สารประกอบประเภทใหม่เพื่อสร้างหลักการของโครงสร้าง
ฟิชเชอร์แก้ไขปัญหานี้ และนักเคมีได้รับหลักฐานที่น่าเชื่อถือว่าโปรตีนเป็นโพลีเมอร์ของกรดอะมิโนที่เชื่อมต่อกันด้วยพันธะเปปไทด์:
CO - CHR" - NH - CO - CHR"" - NH - CO CHR""" - NH -
ตำแหน่งนี้ได้รับการสนับสนุนจากหลักฐานทางชีวเคมี ระหว่างทางปรากฏว่าโปรตีเอสไม่ได้ไฮโดรไลซ์พันธะทั้งหมดระหว่างกรดอะมิโนในอัตราเดียวกัน ความสามารถในการแยกพันธะเปปไทด์ได้รับผลกระทบจากการกำหนดค่าทางแสงของกรดอะมิโน หมู่แทนที่ที่ไนโตรเจนของหมู่อะมิโน ความยาวของสายเปปไทด์ และชุดของเรซิดิวที่รวมอยู่ในนั้น
หลักฐานหลักของทฤษฎีเปปไทด์คือการสังเคราะห์โมเดลเปปไทด์และการเปรียบเทียบกับเปปไทด์ของโปรตีนไฮโดรไลเสต ผลการวิจัยพบว่าเปปไทด์ที่เหมือนกันกับที่สังเคราะห์ได้นั้นแยกได้จากโปรตีนไฮโดรไลเสต
ในการศึกษาเหล่านี้ E.Fischer และนักเรียนของเขา E.Abdergalden ได้พัฒนาวิธีการกำหนดลำดับกรดอะมิโนในโปรตีนเป็นครั้งแรก สาระสำคัญของมันคือการสร้างธรรมชาติของกรดอะมิโนตกค้างของพอลิเปปไทด์ที่มีหมู่อะมิโนอิสระ (กรดอะมิโนปลาย N) ในการทำเช่นนี้ พวกเขาเสนอให้ปิดกั้นปลายอะมิโนในเปปไทด์ด้วยกลุ่มแนฟทาลีน-ซัลโฟนิล ซึ่งไม่ถูกแยกออกจากกันในระหว่างการไฮโดรไลซิส การแยกกรดอะมิโนที่ติดฉลากด้วยหมู่ดังกล่าวออกจากไฮโดรไลเสต ทำให้สามารถระบุได้ว่ากรดอะมิโนใดเป็นปลาย N
หลังจากการวิจัยของอี. ฟิชเชอร์ เห็นได้ชัดว่าโปรตีนเป็นโพลีเปปไทด์ นี่เป็นความสำเร็จที่สำคัญ ซึ่งรวมถึงงานการสังเคราะห์โปรตีน: มันชัดเจนว่าต้องสังเคราะห์อะไรกันแน่หลังจากงานเหล่านี้ปัญหาของการสังเคราะห์โปรตีนได้รับทิศทางที่แน่นอนและความเข้มงวดที่จำเป็น
เมื่อพูดถึงงานของฟิชเชอร์โดยรวมแล้ว ควรสังเกตว่าแนวทางการวิจัยนั้นค่อนข้างจะเป็นเรื่องปกติของศตวรรษที่ 20 ที่จะถึงนี้ ซึ่งดำเนินการด้วยตำแหน่งทางทฤษฎีและเทคนิควิธีการที่หลากหลาย การสังเคราะห์ของเขาดูน้อยลงเรื่อย ๆ เหมือนศิลปะตามสัญชาตญาณมากกว่าความรู้ที่แน่นอนและเข้าใกล้การสร้างชุดของอุปกรณ์เทคโนโลยีที่แม่นยำและเกือบจะ
2. 6 วิกฤตการณ์ของทฤษฎีเปปไทด์
ในการเชื่อมต่อกับการใช้วิธีการวิจัยทางกายภาพและทางเคมีกายภาพใหม่ในช่วงต้นทศวรรษที่ 20 ศตวรรษที่ 20 มีข้อสงสัยว่าโมเลกุลโปรตีนเป็นสายโซ่โพลีเปปไทด์ที่ยาว สมมติฐานเกี่ยวกับความเป็นไปได้ของการบรรจุแบบกะทัดรัดของสายโซ่เปปไทด์ได้รับการปฏิบัติด้วยความสงสัย ทั้งหมดนี้จำเป็นต้องมีการแก้ไขทฤษฎีเปปไทด์ของอี. ฟิชเชอร์
ในยุค 20-30 ทฤษฎี diketopiperazine ได้รับการยอมรับอย่างกว้างขวาง ตามข้อมูลดังกล่าว วงแหวนไดคีโทปิเปอเรสซึ่งเกิดขึ้นในระหว่างการหมุนเวียนของกรดอะมิโนสองตัวที่ตกค้าง มีบทบาทสำคัญในการสร้างโครงสร้างโปรตีน นอกจากนี้ยังสันนิษฐานว่าโครงสร้างเหล่านี้เป็นแกนกลางของโมเลกุลซึ่งมีเปปไทด์สั้นหรือกรดอะมิโนติดอยู่ (“สารตัวเติม” ของโครงกระดูกวัฏจักรของโครงสร้างหลัก) แผนการที่น่าเชื่อถือที่สุดสำหรับการมีส่วนร่วมของ diketopiperazines ในการสร้างโครงสร้างโปรตีนถูกนำเสนอโดยนักเรียนของ N.D. Zelinsky และ E. Fisher
อย่างไรก็ตาม ความพยายามที่จะสังเคราะห์สารประกอบแบบจำลองที่มีไดคีโทปิเพอราซีนนั้นมีผลเพียงเล็กน้อยสำหรับเคมีโปรตีน ต่อมา ทฤษฎีเปปไทด์ได้รับชัยชนะ แต่งานเหล่านี้มีผลกระตุ้นต่อเคมีของไพเพอราซีนโดยทั่วไป
หลังจากทฤษฎีเปปไทด์และไดคีโทปิเปอเรส ความพยายามยังคงพิสูจน์การมีอยู่ของโครงสร้างเปปไทด์เพียงอย่างเดียวในโมเลกุลโปรตีน ในเวลาเดียวกัน พวกเขาพยายามจินตนาการไม่เพียงแต่ประเภทของโมเลกุลเท่านั้น แต่ยังรวมถึงโครงร่างทั่วไปด้วย
สมมติฐานดั้งเดิมแสดงโดย D.L. Talmud นักเคมีชาวโซเวียต เขาแนะนำว่าโซ่เปปไทด์ในองค์ประกอบของโมเลกุลโปรตีนจะถูกพับเป็นวงแหวนขนาดใหญ่ ซึ่งจะเป็นขั้นตอนหนึ่งในการสร้างความคิดของเขาเกี่ยวกับโปรตีนทรงกลม
ในเวลาเดียวกัน ข้อมูลปรากฏขึ้นซึ่งบ่งชี้ชุดของกรดอะมิโนที่แตกต่างกันในโปรตีนที่ต่างกัน แต่รูปแบบที่ควบคุมลำดับของกรดอะมิโนในโครงสร้างโปรตีนนั้นไม่ชัดเจน
M. Bergman และ K. Niemann เป็นคนแรกที่พยายามตอบคำถามนี้ในสมมติฐานของ "ความถี่ที่ไม่ต่อเนื่อง" ตามนั้น ลำดับของกรดอะมิโนตกค้างในโมเลกุลโปรตีนเป็นไปตามรูปแบบตัวเลข ซึ่งเป็นรากฐานมาจากหลักการของโครงสร้างของโมเลกุลโปรตีนไหมไฟโบรอิน แต่ตัวเลือกนี้ไม่ประสบความสำเร็จเพราะ โปรตีนนี้เป็นไฟบริลลาร์ ในขณะที่โครงสร้างของโปรตีนทรงกลมเป็นไปตามรูปแบบที่แตกต่างกันโดยสิ้นเชิง
ตามคำกล่าวของ M. Bergman และ K. Nieman กรดอะมิโนแต่ละชนิดเกิดขึ้นในสายโซ่โพลีเปปไทด์ในช่วงเวลาหนึ่งหรือตามที่เอ็ม. เบิร์กแมนกล่าวว่ามี “ความเป็นช่วง” ที่แน่นอน ระยะนี้ถูกกำหนดโดยธรรมชาติของกรดอะมิโนตกค้าง
พวกเขาจินตนาการถึงโมเลกุลไหมไฟโบรอินดังนี้:
GlyAlaGlyTyr GlyAlaGlyอาร์ก GlyAlaGlyx GlyAlaGlyx
(GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyx) 12
GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyอาร์ก
(GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyx) 13
สมมติฐาน Bergman-Niemann มีผลกระทบอย่างมีนัยสำคัญต่อการพัฒนาเคมีของกรดอะมิโน มีงานจำนวนมากที่ทุ่มเทให้กับการตรวจสอบ
บทสรุปของบทนี้ควรสังเกตว่าในช่วงกลางศตวรรษที่ XX มีการสะสมหลักฐานเพียงพอของความถูกต้องของทฤษฎีเปปไทด์ บทบัญญัติหลักได้รับการเสริมและกลั่นกรอง ดังนั้นศูนย์วิจัยโปรตีนในศตวรรษที่ 20 ได้วางขอบเขตการวิจัยและค้นหาวิธีการสังเคราะห์โปรตีนด้วยวิธีเทียมแล้ว ปัญหานี้แก้ไขได้สำเร็จ มีการพัฒนาวิธีการที่เชื่อถือได้ในการกำหนดโครงสร้างหลักของโปรตีน - ลำดับของกรดอะมิโนในสายเปปไทด์ วิธีการสำหรับการสังเคราะห์ทางเคมี (abiogenic) ของพอลิเปปไทด์ที่ผิดปกติได้รับการพัฒนา (วิธีการเหล่านี้จะกล่าวถึงในรายละเอียดเพิ่มเติม ในบทที่ 8 หน้า 36) รวมถึงวิธีการสังเคราะห์พอลิเปปไทด์โดยอัตโนมัติ สิ่งนี้ทำให้เป็นไปได้ในปี 1962 สำหรับนักเคมีชาวอังกฤษที่ใหญ่ที่สุด F. Senger เพื่อถอดรหัสโครงสร้างและสังเคราะห์ฮอร์โมนอินซูลินเทียมซึ่งเป็นยุคใหม่ในการสังเคราะห์โปรตีนโพลีเปปไทด์ที่ใช้งานได้
บทที่ 3 องค์ประกอบทางเคมีของโปรตีน
3.1 พันธะเปปไทด์
โปรตีนเป็นโพลีเมอร์ที่ไม่ปกติซึ่งสร้างขึ้นจากสารตกค้างของกรด α-amino ซึ่งสูตรทั่วไปในสารละลายที่เป็นน้ำที่ค่า pH ใกล้เคียงกับค่าเป็นกลางสามารถเขียนเป็น NH 3 + CHRCOO - . เรซิดิวกรดอะมิโนในโปรตีนถูกเชื่อมโยงเข้าด้วยกันโดยพันธะเอไมด์ระหว่างหมู่ α-อะมิโนและα-คาร์บอกซิล พันธะเปปไทด์ระหว่าง สอง- สารตกค้างของกรดอะมิโนมักถูกเรียกว่า พันธะเปปไทด์ และพอลิเมอร์ที่สร้างขึ้นจากสารตกค้างของกรด α-amino ที่เชื่อมต่อกันด้วยพันธะเปปไทด์เรียกว่า โพลีเปปไทด์ โปรตีนในฐานะโครงสร้างที่มีนัยสำคัญทางชีววิทยาสามารถเป็นได้ทั้งโพลีเปปไทด์เดี่ยวหรือโพลีเปปไทด์หลายตัวที่ก่อรูปสารเชิงซ้อนเดียวอันเป็นผลมาจากการกระทำระหว่างกันที่ไม่ใช่โควาเลนต์
3.2 องค์ประกอบองค์ประกอบของโปรตีน
จากการศึกษาองค์ประกอบทางเคมีของโปรตีนจำเป็นต้องค้นหาก่อนอื่นว่าองค์ประกอบทางเคมีประกอบด้วยอะไรและประการที่สองโครงสร้างของโมโนเมอร์ เพื่อตอบคำถามแรก กำหนดองค์ประกอบเชิงปริมาณและคุณภาพขององค์ประกอบทางเคมีของโปรตีน การวิเคราะห์ทางเคมีแสดงให้เห็น มีอยู่ในโปรตีนทั้งหมด คาร์บอน (50-55%), ออกซิเจน (21-23%), ไนโตรเจน (15-17%), ไฮโดรเจน (6-7%), กำมะถัน (0.3-2.5%) นอกจากนี้ยังพบฟอสฟอรัส ไอโอดีน เหล็ก ทองแดง และมาโครและองค์ประกอบขนาดเล็กอื่นๆ ในองค์ประกอบของโปรตีนแต่ละชนิดในปริมาณที่น้อยมาก
เนื้อหาขององค์ประกอบทางเคมีหลักในโปรตีนสามารถเปลี่ยนแปลงได้ ยกเว้นไนโตรเจน ความเข้มข้นที่มีลักษณะเฉพาะคือความคงตัวมากที่สุดและเฉลี่ย 16% นอกจากนี้ ปริมาณไนโตรเจนในสารอินทรีย์อื่นๆ ยังต่ำอีกด้วย ตามนี้ เสนอให้กำหนดปริมาณโปรตีนโดยไนโตรเจนที่เป็นส่วนประกอบ เมื่อรู้ว่าไนโตรเจน 1 กรัมบรรจุอยู่ในโปรตีน 6.25 กรัม ปริมาณไนโตรเจนที่พบจะถูกคูณด้วยค่า 6.25 และได้รับปริมาณโปรตีน
เพื่อตรวจสอบลักษณะทางเคมีของโปรตีนโมโนเมอร์ จำเป็นต้องแก้ปัญหาสองประการ: แยกโปรตีนออกเป็นโมโนเมอร์และค้นหาองค์ประกอบทางเคมีของโปรตีน การสลายโปรตีนออกเป็นส่วนๆ ทำได้โดยการไฮโดรไลซิส - การเดือดของโปรตีนเป็นเวลานานด้วยกรดแร่ที่แรง (กรดไฮโดรไลซิส)หรือบริเวณ (ไฮโดรไลซิสอัลคาไลน์). โดยทั่วไปจะใช้การต้มที่อุณหภูมิ 110 องศาเซลเซียสกับ HCl เป็นเวลา 24 ชั่วโมง ในขั้นตอนต่อไป สารที่ประกอบเป็นไฮโดรไลเสตจะถูกแยกออก เพื่อจุดประสงค์นี้มีการใช้วิธีการต่าง ๆ บ่อยที่สุด - โครมาโตกราฟี (สำหรับรายละเอียดเพิ่มเติมโปรดดูบทที่ "วิธีการวิจัย ... ") กรดอะมิโนเป็นส่วนหลักของไฮโดรไลเสตที่แยกจากกัน
3.3. กรดอะมิโน
ปัจจุบันพบกรดอะมิโนที่แตกต่างกันถึง 200 ชนิดในวัตถุต่างๆ ของสัตว์ป่า ตัวอย่างเช่นในร่างกายมนุษย์มีประมาณ 60 ตัว อย่างไรก็ตามโปรตีนมีกรดอะมิโนเพียง 20 ชนิดซึ่งบางครั้งเรียกว่ากรดจากธรรมชาติ
กรดอะมิโนเป็นกรดอินทรีย์ที่อะตอมของไฮโดรเจน - อะตอมของคาร์บอนถูกแทนที่ด้วยหมู่อะมิโน - NH 2 ดังนั้นโดยธรรมชาติทางเคมี กรดอะมิโนเหล่านี้จึงมีสูตรทั่วไปคือ
จากสูตรนี้จะเห็นได้ว่าองค์ประกอบของกรดอะมิโนทั้งหมดประกอบด้วยหมู่ทั่วไปดังต่อไปนี้: - CH 2 - NH 2 - COOH. โซ่ด้านข้าง (อนุมูล - R) กรดอะมิโนต่างกัน ดังที่เห็นได้จากภาคผนวก 1 ลักษณะทางเคมีของอนุมูลมีความหลากหลาย ตั้งแต่อะตอมไฮโดรเจนไปจนถึงสารประกอบไซคลิก เป็นอนุมูลที่กำหนดคุณสมบัติโครงสร้างและหน้าที่ของกรดอะมิโน
กรดอะมิโนทั้งหมด ยกเว้น glycine อะมิโนอะซิติกที่ง่ายที่สุดสำหรับคุณ (NH 3 + CH 2 COO) มีไครัลอะตอม C และสามารถมีอยู่ในรูปของอีแนนทิโอเมอร์สองตัว (ไอโซเมอร์ออปติคัล):
โปรตีนที่ศึกษาทั้งหมดในปัจจุบันมีเพียงกรดอะมิโนของซีรีย์ L ซึ่งหากเราพิจารณาอะตอม chiral จากด้านข้างของอะตอม H กลุ่ม NH 3 + , COO และ R Radical จะเรียงตามเข็มนาฬิกา ความจำเป็นในการสร้างโมเลกุลพอลิเมอร์ที่มีนัยสำคัญทางชีวภาพจากอีแนนชิโอเมอร์ที่กำหนดไว้อย่างเข้มงวดนั้นชัดเจน - จากส่วนผสมของราซีมิกของอีแนนชิโอเมอร์สองตัว จะได้ส่วนผสมที่ซับซ้อนอย่างคาดไม่ถึงของไดแอสเทอริโอไอโซเมอร์ คำถามที่ว่าทำไมสิ่งมีชีวิตบนโลกขึ้นอยู่กับโปรตีนที่สร้างขึ้นอย่างแม่นยำจาก L- และไม่ใช่กรด D-amino ยังคงเป็นปริศนาที่น่าสนใจ ควรสังเกตว่ากรด D-amino นั้นค่อนข้างแพร่หลายในธรรมชาติและยิ่งกว่านั้นยังเป็นส่วนหนึ่งของ oligopeptides ที่มีความสำคัญทางชีวภาพ
โปรตีนถูกสร้างขึ้นจากกรด α-amino พื้นฐาน 20 ชนิด แต่ส่วนที่เหลือ กรดอะมิโนที่ค่อนข้างหลากหลาย เกิดขึ้นจากกรดอะมิโน 20 ชนิดที่มีอยู่แล้วในองค์ประกอบของโมเลกุลโปรตีน ในบรรดาการเปลี่ยนแปลงเหล่านี้ อันดับแรกควรสังเกตการก่อตัว สะพานซัลไฟด์ ในระหว่างการออกซิเดชันของซิสเทอีนสองตัวที่ตกค้างในองค์ประกอบของโซ่เปปไทด์ที่เกิดขึ้นแล้ว เป็นผลให้สารตกค้างของกรดไดอะมิโนไดคาร์บอกซิลิกเกิดขึ้นจากซิสเทอีนเรซิดิวสองตัว ซีสทีน (ดูภาคผนวก I) ในกรณีนี้ การเชื่อมขวางเกิดขึ้นภายในสายโพลีเปปไทด์หนึ่งสายหรือระหว่างสายโซ่ที่ต่างกันสองสาย ในฐานะที่เป็นโปรตีนขนาดเล็กที่มีสายพอลิเปปไทด์สองสายเชื่อมต่อกันด้วยไดซัลไฟด์บริดจ์ เช่นเดียวกับการเชื่อมขวางภายในหนึ่งในสายโซ่โพลีเปปไทด์:
ตัวอย่างที่สำคัญของการดัดแปลงเรซิดิวกรดอะมิโนคือการแปลงโพรลีนเรซิดิวไปเป็นเรซิดิว ไฮดรอกซีโพรลีน :
การเปลี่ยนแปลงนี้เกิดขึ้นและในระดับที่มีนัยสำคัญ ระหว่างการก่อตัวขององค์ประกอบโปรตีนที่สำคัญของเนื้อเยื่อเกี่ยวพัน - คอลลาเจน .
การดัดแปลงโปรตีนที่สำคัญมากอีกประเภทหนึ่งคือ ฟอสโฟรีเลชั่นของกลุ่มไฮดรอกโซของสารตกค้างของซีรีน ทรีโอนีน และไทโรซีน ตัวอย่างเช่น
กรดอะมิโนในสารละลายในน้ำอยู่ในสถานะแตกตัวเป็นไอออนเนื่องจากการแตกตัวของหมู่อะมิโนและคาร์บอกซิลที่ประกอบขึ้นเป็นอนุมูล กล่าวอีกนัยหนึ่ง พวกมันเป็นสารประกอบแอมโฟเทอริกและสามารถมีอยู่ได้ทั้งในรูปกรด (ผู้ให้โปรตอน) หรือในฐานะที่เป็นเบส (ตัวรับผู้บริจาค)
กรดอะมิโนทั้งหมดขึ้นอยู่กับโครงสร้างแบ่งออกเป็นหลายกลุ่ม:
อะไซคลิก. กรดอะมิโนโมโนอะมิโนโมโนคาร์บอกซิลิกมีเอมีนหนึ่งกลุ่มและกลุ่มคาร์บอกซิลหนึ่งกลุ่มในสารละลายที่เป็นน้ำพวกมันเป็นกลาง บางส่วนมีลักษณะโครงสร้างทั่วไปซึ่งช่วยให้พิจารณาร่วมกันได้:
ไกลซีนและอะลานีน Glycine (glycocol หรือ aminoacetic acid) ไม่ใช้งานทางสายตา - เป็นกรดอะมิโนเพียงชนิดเดียวที่ไม่มีอีแนนทิโอเมอร์ Glycine มีส่วนร่วมในการก่อตัวของนิวคลีอิกและน้ำดีถึง - t, heme เป็นสิ่งจำเป็นสำหรับการวางตัวเป็นกลางของผลิตภัณฑ์ที่เป็นพิษในตับ ร่างกายใช้อะลานีนในกระบวนการเผาผลาญคาร์โบไฮเดรตและพลังงานต่างๆ ไอโซเมอร์ - อะลานีนเป็นส่วนสำคัญของวิตามินแพนโทธีนิกสำหรับคุณ โคเอ็นไซม์ A (CoA) สารสกัดของกล้ามเนื้อ
ซีรีนและทรีโอนีนพวกมันอยู่ในกลุ่มของกรดไฮดรอกซีเพราะ มีหมู่ไฮดรอกซิล ซีรีนเป็นส่วนหนึ่งของเอ็นไซม์ต่าง ๆ ซึ่งเป็นโปรตีนหลักของนม - เคซีน และยังเป็นส่วนหนึ่งของไลโปโปรตีนหลายชนิด ธรีโอนีนมีส่วนร่วมในการสังเคราะห์โปรตีน ซึ่งเป็นกรดอะมิโนที่จำเป็น
ซิสเทอีนและเมไทโอนีนกรดอะมิโนที่มีอะตอมกำมะถัน ค่าของ cysteine ถูกกำหนดโดยการปรากฏตัวของกลุ่ม sulfhydryl (-SH) ในองค์ประกอบของมันซึ่งทำให้สามารถออกซิไดซ์ได้ง่ายและปกป้องร่างกายจากสารที่มีความสามารถในการออกซิไดซ์สูง (ในกรณีที่ได้รับบาดเจ็บจากรังสีฟอสฟอรัส พิษ) เมไทโอนีนมีลักษณะเด่นคือมีหมู่เมทิลเคลื่อนที่ได้ง่าย ซึ่งใช้สำหรับการสังเคราะห์สารประกอบสำคัญในร่างกาย (โคลีน ครีเอทีน ไทมีน อะดรีนาลีน เป็นต้น)
วาลีน ลิวซีน และไอโซลิวซีนพวกมันเป็นกรดอะมิโนที่แตกแขนงซึ่งเกี่ยวข้องอย่างแข็งขันในการเผาผลาญและไม่ได้สังเคราะห์ในร่างกาย
กรดอะมิโนโมโนอะมิโนไดคาร์บอกซิลิกมีหมู่อะมิโนหนึ่งหมู่และหมู่คาร์บอกซิลสองหมู่ และให้ปฏิกิริยาที่เป็นกรดในสารละลายที่เป็นน้ำ เหล่านี้รวมถึงแอสปาร์ติกและกลูตามีนสำหรับคุณ แอสพาราจีนและกลูตามีน พวกมันเป็นส่วนหนึ่งของตัวกลางไกล่เกลี่ยของระบบประสาท
กรดอะมิโนไดอะมิโนโมโนคาร์บอกซิลิกในสารละลายที่เป็นน้ำจะมีปฏิกิริยาเป็นด่างเนื่องจากการมีอยู่ของหมู่เอมีนสองกลุ่ม ที่เกี่ยวข้องกับพวกเขา ไลซีนเป็นสิ่งจำเป็นสำหรับการสังเคราะห์ฮิสโตนและในเอนไซม์จำนวนหนึ่ง อาร์จินีนมีส่วนร่วมในการสังเคราะห์ยูเรียครีเอทีน
วัฏจักร. กรดอะมิโนเหล่านี้มีนิวเคลียสอะโรมาติกหรือเฮเทอโรไซคลิกในองค์ประกอบและตามกฎแล้วจะไม่ถูกสังเคราะห์ในร่างกายมนุษย์และต้องได้รับอาหาร พวกเขามีส่วนร่วมอย่างแข็งขันในกระบวนการเผาผลาญที่หลากหลาย ดังนั้น ฟีนิล-อะลานีนจึงทำหน้าที่เป็นแหล่งหลักในการสังเคราะห์ไทโรซีน ซึ่งเป็นสารตั้งต้นของสารสำคัญทางชีววิทยาจำนวนหนึ่ง ได้แก่ ฮอร์โมน (ไทรอกซีน อะดรีนาลีน) เม็ดสีบางชนิด ทริปโตเฟนนอกจากจะมีส่วนร่วมในการสังเคราะห์โปรตีนแล้วยังเป็นส่วนประกอบของวิตามินพีพี เซโรโทนิน ทริปตามีน และเม็ดสีอีกจำนวนหนึ่ง ฮิสทิดีนจำเป็นสำหรับการสังเคราะห์โปรตีน ซึ่งเป็นสารตั้งต้นของฮีสตามีน ซึ่งส่งผลต่อความดันโลหิตและการหลั่งน้ำย่อย
บทที่ 4 โครงสร้าง
เมื่อศึกษาองค์ประกอบของโปรตีน พบว่า โปรตีนทั้งหมดถูกสร้างขึ้นตามหลักการเดียวและมีการจัดองค์กรสี่ระดับ: ประถม, มัธยม, ตติยภูมิ,และบางส่วนของพวกเขา ควอเตอร์นารีโครงสร้าง
4.1 โครงสร้างหลัก
เป็นสายโซ่เชิงเส้นของกรดอะมิโนที่จัดเรียงเป็นลำดับที่แน่นอนและเชื่อมต่อกันด้วยพันธะเปปไทด์ พันธะเปปไทด์ เกิดขึ้นจากหมู่ -คาร์บอกซิลของกรดอะมิโนหนึ่งและหมู่ -เอมีนของอีกกลุ่มหนึ่ง:
พันธะเปปไทด์อันเนื่องมาจากพันธะ p, -คอนจูเกชั่น -พันธะของกลุ่มคาร์บอนิลและ p-orbital ของอะตอม N ซึ่งจัดวางคู่อิเล็กตรอนที่ไม่แบ่งแยกไว้ ไม่สามารถถือเป็นพันธะเดี่ยวและแทบไม่มีการหมุนรอบ มัน. ด้วยเหตุผลเดียวกัน ไครัลอะตอม C และคาร์บอนิลอะตอม Ck ของเรซิดิวกรดอะมิโนตัวที่ i ใดๆ ของสายเปปไทด์และอะตอม N และ C ของเรซิดิวที่ (i+1)-อยู่ในระนาบเดียวกัน อะตอมของคาร์บอนิล O และอะตอมของเอไมด์ H อยู่ในระนาบเดียวกัน (อย่างไรก็ตาม วัสดุที่สะสมในการศึกษาโครงสร้างของโปรตีนแสดงให้เห็นว่าข้อความนี้ไม่ได้เข้มงวดทั้งหมด: อะตอมที่เกี่ยวข้องกับอะตอมของเปปไทด์ไนโตรเจนไม่ได้อยู่ใน ระนาบเดียวกันกับมัน แต่สร้างพีระมิดสามส่วนที่มีมุมระหว่างพันธะใกล้กับ 120 ดังนั้นระหว่างระนาบที่เกิดจากอะตอม C ผม , C ผม , O ผม และ N ผม +1 , H ผม +1 , C ผม + 1 มีบางมุมที่แตกต่างจาก 0 แต่ตามกฎแล้วไม่เกิน 1 และไม่มีบทบาทพิเศษ) ดังนั้น ในเชิงเรขาคณิต โซ่โพลีเปปไทด์ถือได้ว่าก่อตัวขึ้นโดยชิ้นส่วนแบนๆ ดังกล่าวที่แต่ละอะตอมมีหกอะตอม การจัดเรียงชิ้นส่วนเหล่านี้ร่วมกัน เช่นเดียวกับการจัดเรียงระนาบสองระนาบร่วมกัน จะต้องกำหนดโดยมุมสองมุม ด้วยเหตุนี้ จึงเป็นเรื่องปกติที่จะใช้มุมบิดซึ่งกำหนดลักษณะการหมุนรอบพันธะ N C และ C C k
เรขาคณิตของโมเลกุลใด ๆ ถูกกำหนดโดยลักษณะทางเรขาคณิตสามกลุ่มของพันธะเคมี - ความยาวของพันธะ มุมพันธะ และมุมบิดระหว่างพันธะที่อยู่ติดกับอะตอมข้างเคียง สองกลุ่มแรกถูกกำหนดในระดับเด็ดขาดโดยธรรมชาติของอะตอมที่มีส่วนร่วมและพันธะที่เกิดขึ้น ดังนั้นโครงสร้างเชิงพื้นที่ของโพลีเมอร์จึงถูกกำหนดโดยส่วนใหญ่โดยมุมบิดระหว่างการเชื่อมโยงของกระดูกสันหลังของพอลิเมอร์ของโมเลกุลเช่น โครงสร้างโซ่โพลีเมอร์ ที่ R มุมไอออน , เช่น. มุมการหมุนของการเชื่อมต่อ A-B รอบการเชื่อมต่อ B-C ที่สัมพันธ์กับการเชื่อมต่อ C-ดีถูกกำหนดให้เป็นมุมระหว่างระนาบที่มีอะตอม A, B, C และอะตอมบี, ค, ดี.
ในระบบดังกล่าว อาจเป็นไปได้ว่าพันธะ A-B และ C-D จะขนานกันและอยู่ด้านเดียวกันของพันธะ B-C หากเราพิจารณาระบบนี้ควบคู่ไปกับฉันzi B-C จากนั้นการเชื่อมต่อ A-B เหมือนเดิมก็บดบังการเชื่อมต่อค- ดีจึงเรียกรูปแบบนี้ว่าสวาทยาบดบัง ตามคำแนะนำของสหภาพเคมีระหว่างประเทศ IUPAC (International Union of Pure and Applied Chemistry) และ IUB (International Union of Biochemistry) มุมระหว่างระนาบ ABC และ BCD ถือเป็นค่าบวก ถ้าเพื่อนำรูปแบบไปเป็น บดบังสภาพโดยการหมุนผ่านมุมไม่เกิน 180 ใกล้จุดเชื่อมต่อของผู้สังเกตมากที่สุดจะต้องหมุนตามเข็มนาฬิกา หากพันธะนี้ต้องหมุนทวนเข็มนาฬิกาเพื่อให้ได้รูปร่างที่บดบัง มุมจะถือเป็นค่าลบ จะเห็นได้ว่าคำจำกัดความนี้ไม่ได้ขึ้นอยู่กับพันธะที่ใกล้ชิดกับผู้สังเกต
ในกรณีนี้ ดังที่เห็นได้จากรูป ทิศทางของชิ้นส่วนที่มีอะตอม C i -1 และ C i [(i-1)-th fragment] และชิ้นส่วนที่มีอะตอม C i และ C i + 1 (ส่วน i-th) ถูกกำหนดโดยมุมบิดที่สอดคล้องกับการหมุนรอบพันธะ N ผม C ผม และพันธะ C ผม C ผม k . มุมเหล่านี้มักจะแสดงเป็น และ ในกรณีที่กำหนด ตามลำดับ ผม และ ผม ค่าของมันสำหรับหน่วยโมโนเมอร์ทั้งหมดของสายโซ่โพลีเปปไทด์ส่วนใหญ่จะกำหนดรูปทรงเรขาคณิตของสายโซ่นี้ ไม่มีค่าที่ชัดเจนสำหรับค่าของแต่ละมุมเหล่านี้หรือสำหรับการรวมกันของพวกเขาแม้ว่าจะมีการกำหนดข้อ จำกัด สำหรับทั้งคู่ซึ่งพิจารณาจากคุณสมบัติของชิ้นส่วนเปปไทด์เองและโดยธรรมชาติของอนุมูลด้านข้างเช่น ลักษณะของกรดอะมิโนตกค้าง
จนถึงปัจจุบัน มีการสร้างลำดับกรดอะมิโนสำหรับโปรตีนต่างๆ หลายพันชนิด การบันทึกโครงสร้างของโปรตีนในรูปแบบของสูตรโครงสร้างโดยละเอียดนั้นยุ่งยากและมองไม่เห็น ดังนั้นจึงใช้รูปแบบการเขียนย่อ - สามตัวอักษรหรือหนึ่งตัวอักษร (โมเลกุล vasopressin):
เมื่อเขียนลำดับกรดอะมิโนในสายพอลิเปปไทด์หรือโอลิโกเปปไทด์โดยใช้สัญลักษณ์ย่อ จะถูกสันนิษฐาน เว้นแต่จะระบุไว้เป็นอย่างอื่นว่าหมู่ α-อะมิโนอยู่ทางซ้ายและหมู่ α-คาร์บอกซิลอยู่ทางขวา ส่วนที่สอดคล้องกันของสายพอลิเปปไทด์เรียกว่าปลาย N (ปลายเอมีน) และปลาย C (ปลายคาร์บอกซิล) และส่วนที่เหลือของกรดอะมิโนเรียกว่าเรซิดิวของปลาย N และปลาย C ตามลำดับ
4.2 โครงสร้างรอง
ชิ้นส่วนของโครงสร้างเชิงพื้นที่ของพอลิเมอร์ชีวภาพที่มีโครงสร้างเป็นระยะของแกนหลักโพลีเมอร์ถือเป็นองค์ประกอบของโครงสร้างทุติยภูมิ
หากมุมของประเภทเดียวกันที่กล่าวถึงในหน้า 15 ใกล้เคียงกันในบางส่วนของโซ่ โครงสร้างของสายโพลีเปปไทด์จะมีคุณลักษณะเป็นระยะ โครงสร้างดังกล่าวมีสองประเภท - เกลียวและยืด (แบนหรือพับ)
เกลียวพิจารณาโครงสร้างที่อะตอมทั้งหมดประเภทเดียวกันอยู่บนเกลียวเดียวกัน ในกรณีนี้ เกลียวจะถือว่าถูกต้อง ถ้าเมื่อสังเกตตามแนวแกนของเกลียว เกลียวจะเคลื่อนออกจากผู้สังเกตตามเข็มนาฬิกา และไปทางซ้าย - ถ้ามันเคลื่อนออกทวนเข็มนาฬิกา สายโซ่โพลีเปปไทด์มีโครงสร้างเป็นเกลียวหากอะตอม C ทั้งหมดอยู่บนเกลียวเดียว อะตอมคาร์บอนิลทั้งหมด C k - อยู่อีกด้านหนึ่ง อะตอม N ทั้งหมด - ในส่วนที่สาม และระยะพิทช์ของเกลียวสำหรับอะตอมทั้งสามกลุ่มควรเหมือนกัน จำนวนอะตอมต่อเกลียวหนึ่งรอบก็ควรเท่ากัน ไม่ว่าเราจะพูดถึงอะตอม C k , C หรือ N หรือไม่ก็ตาม ระยะห่างจากเกลียวทั่วไปสำหรับอะตอมทั้งสามประเภทนี้จะต่างกัน
องค์ประกอบหลักของโครงสร้างรองของโปรตีนคือ -helices และ -folds
โครงสร้างโปรตีนเฮลิคอล เกลียวหลายชนิดเป็นที่รู้จักสำหรับสายโพลีเปปไทด์ ในหมู่พวกเขาเกลียวขวาเป็นเรื่องธรรมดาที่สุด เกลียวในอุดมคติมีระยะพิทช์ 0.54 นาโนเมตร และจำนวนอะตอมของชนิดเดียวกันต่อเทิร์นของเกลียวคือ 3.6 ซึ่งหมายถึงคาบที่สมบูรณ์ในห้ารอบของเกลียวทุกๆ 18 กรดอะมิโนตกค้าง ค่ามุมบิดสำหรับ α-helix ในอุดมคติ = - 57 = - 47 , และระยะทางจากอะตอมที่สร้างสายโซ่โพลีเปปไทด์ไปยังแกนของเกลียวคือ 0.15 นาโนเมตรสำหรับ N, 0.23 นาโนเมตรสำหรับ C และ 0.17 นาโนเมตร สำหรับซีเค โครงสร้างใด ๆ ที่มีอยู่โดยมีปัจจัยที่ทำให้เสถียร ในกรณีของเกลียว ปัจจัยดังกล่าวคือพันธะไฮโดรเจนที่เกิดจากอะตอมคาร์บอนิลแต่ละอะตอมของชิ้นส่วนที่ (i + 4) ปัจจัยสำคัญในการรักษาเสถียรภาพของα-helix ก็คือการวางแนวขนานของโมเมนต์ไดโพลของพันธะเปปไทด์
โครงสร้างโปรตีนแบบพับ หนึ่งในตัวอย่างทั่วไปของโครงสร้างเป็นระยะพับของโปรตีนคือสิ่งที่เรียกว่า -พับประกอบด้วยสองชิ้นส่วน แต่ละชิ้นมีโพลีเปปไทด์แทน
การพับยังถูกทำให้เสถียรโดยพันธะไฮโดรเจนระหว่างอะตอมไฮโดรเจนของกลุ่มเอมีนของชิ้นส่วนหนึ่งกับอะตอมออกซิเจนของกลุ่มคาร์บอกซิลของอีกชิ้นส่วนหนึ่ง ในกรณีนี้ แฟรกเมนต์สามารถมีการวางแนวขนานและขนานกันที่สัมพันธ์กัน
โครงสร้างที่เกิดจากปฏิสัมพันธ์ดังกล่าวเป็นโครงสร้างลูกฟูก สิ่งนี้ส่งผลต่อค่าของมุมบิดและ หากอยู่ในโครงสร้างที่แบนและยืดเต็มที่ควรเป็น 180 ดังนั้นในชั้น β จริงจะมีค่า = - 119 และ = + 113 เพื่อให้สองส่วนของสายโซ่โพลีเปปไทด์อยู่ในแนวที่ชอบ การก่อตัวของ α-folds ซึ่งเป็นส่วนที่มีโครงสร้างที่แตกต่างอย่างมากจากส่วนเป็นระยะ
4.2.1 ปัจจัยที่มีผลต่อการสร้างโครงสร้างทุติยภูมิ
โครงสร้างของบางส่วนของสายโซ่โพลีเปปไทด์ขึ้นอยู่กับโครงสร้างของโมเลกุลโดยรวมเป็นหลัก ปัจจัยที่มีอิทธิพลต่อการก่อตัวของพื้นที่ที่มีโครงสร้างทุติยภูมินั้นมีความหลากหลายมากและไม่ได้ระบุอย่างครบถ้วนในทุกกรณี เป็นที่ทราบกันว่ามีกรดอะมิโนตกค้างจำนวนหนึ่งเกิดขึ้นเป็นพิเศษในชิ้นส่วนเกลียว α ส่วนอีกจำนวนหนึ่ง - ใน α-เท่า กรดอะมิโนบางตัว - ส่วนใหญ่อยู่ในบริเวณที่ไม่มีโครงสร้างเป็นระยะ โครงสร้างรองถูกกำหนดโดยโครงสร้างหลักเป็นส่วนใหญ่ ในบางกรณี ความหมายทางกายภาพของการพึ่งพาอาศัยกันดังกล่าวสามารถเข้าใจได้จากการวิเคราะห์ทางสเตอริโอเคมีของโครงสร้างเชิงพื้นที่ ตัวอย่างเช่น ดังที่เห็นได้จากรูป ไม่เพียงแต่เรดิคัลข้างเคียงของกรดอะมิโนที่ตกค้างตามสายโซ่เท่านั้นที่ถูกนำมารวมกันใน -helix แต่ยังมีสารตกค้างบางคู่ที่อยู่บนเกลียวที่อยู่ติดกันด้วย อย่างแรกเลย แต่ละอัน (i + 1) เรซิดิวที่ (i + 4) -th และ (i+5)-th ดังนั้นในตำแหน่ง (i + 1) และ (i + 2), (i + 1) และ (i + 4), (i + 1) และ (i + 5) -helices สองอนุมูลขนาดใหญ่ไม่ค่อยเกิดขึ้นพร้อมกันเช่น ตัวอย่างเช่น , เป็นอนุมูลด้านข้างของไทโรซีน, ทริปโตเฟน, ไอโซลิวซีน เข้ากันได้น้อยกว่ากับโครงสร้างเกลียวคือการมีอยู่พร้อมกันของเรซิดิวขนาดใหญ่สามตัวในตำแหน่ง (i+1), (i+2) และ (i+5) หรือ (i+1), (i+4) และ (i+ 5). ดังนั้น การรวมกรดอะมิโนดังกล่าวในชิ้นส่วนเฮลิคัลเป็นข้อยกเว้นที่หาได้ยาก
4.3 โครงสร้างระดับตติยภูมิ
คำนี้หมายถึงการพับสมบูรณ์ในช่องว่างของสายพอลิเปปไทด์ทั้งหมด รวมถึงการพับของเรดิคัลด้านข้าง ภาพที่สมบูรณ์ของโครงสร้างระดับอุดมศึกษาได้มาจากพิกัดของอะตอมทั้งหมดของโปรตีน ด้วยความสำเร็จอย่างมหาศาลของการวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ ข้อมูลดังกล่าว ยกเว้นพิกัดของอะตอมไฮโดรเจน ได้รับสำหรับโปรตีนจำนวนมาก ข้อมูลเหล่านี้เป็นข้อมูลจำนวนมหาศาลที่จัดเก็บไว้ในคลังข้อมูลพิเศษบนสื่อที่เครื่องอ่านได้ และการประมวลผลนั้นคิดไม่ถึงโดยไม่ต้องใช้คอมพิวเตอร์ความเร็วสูง พิกัดอะตอมที่ได้รับจากคอมพิวเตอร์ให้ข้อมูลที่ครบถ้วนเกี่ยวกับเรขาคณิตของสายโซ่โพลีเปปไทด์ รวมถึงค่าของมุมบิด ซึ่งทำให้สามารถเปิดเผยโครงสร้างเป็นเกลียว รอยพับ หรือชิ้นส่วนที่ไม่ปกติได้ ตัวอย่างของแนวทางการวิจัยดังกล่าวคือแบบจำลองเชิงพื้นที่ของโครงสร้างของเอนไซม์ไคเนสฟอสกลีเซอเรต:
โครงร่างทั่วไปของโครงสร้างของไคเนสฟอสโฟกลีเซอเรต เพื่อความชัดเจน ส่วนที่เป็นเกลียว α จะแสดงเป็นทรงกระบอก และส่วนพับ α จะแสดงเป็นริบบิ้นพร้อมลูกศรระบุทิศทางของโซ่จากปลาย N ถึงปลาย C เส้นเป็นส่วนที่ผิดปกติซึ่งเชื่อมต่อชิ้นส่วนที่มีโครงสร้าง
ภาพของโครงสร้างที่สมบูรณ์ของแม้แต่โมเลกุลโปรตีนขนาดเล็กบนระนาบ ไม่ว่าจะเป็นหน้าหนังสือหรือหน้าจอแสดงผล ไม่ได้ให้ข้อมูลมากนักเนื่องจากโครงสร้างที่ซับซ้อนอย่างยิ่งของวัตถุ เพื่อให้ผู้วิจัยสามารถเห็นภาพโครงสร้างเชิงพื้นที่ของโมเลกุลของสารที่ซับซ้อนได้ พวกเขาใช้วิธีการของคอมพิวเตอร์กราฟิกสามมิติ ซึ่งช่วยให้สามารถแสดงแต่ละส่วนของโมเลกุลและจัดการพวกมัน โดยเฉพาะอย่างยิ่ง การเปลี่ยนพวกมันใน มุมขวา.
โครงสร้างระดับตติยภูมิเกิดขึ้นจากปฏิกิริยาที่ไม่ใช่โควาเลนต์ (ไฟฟ้าสถิต ไอออนิก แรงแวนเดอร์วาลส์ ฯลฯ) ของอนุมูลด้านข้างที่จัดกรอบ α-helices และส่วนพับ และชิ้นส่วนที่ไม่เป็นระยะๆ ของสายพอลิเปปไทด์ ในบรรดาพันธบัตรที่ถือโครงสร้างตติยภูมิควรสังเกต:
ก) สะพานไดซัลไฟด์ (- S - S -)
b) สะพานเอสเทอร์ (ระหว่างหมู่คาร์บอกซิลและหมู่ไฮดรอกซิล)
ค) สะพานเกลือ (ระหว่างหมู่คาร์บอกซิลและหมู่อะมิโน)
ง) พันธะไฮโดรเจน
ตามรูปร่างของโมเลกุลโปรตีนเนื่องจากโครงสร้างระดับตติยภูมิกลุ่มโปรตีนต่อไปนี้มีความโดดเด่น:
โปรตีนทรงกลม โครงสร้างเชิงพื้นที่ของโปรตีนเหล่านี้ในการประมาณคร่าวๆ สามารถแสดงเป็นลูกบอลหรือทรงรีที่ไม่ยาวเกินไป - globที่ลี่. ตามกฎแล้ว ส่วนสำคัญของสายโซ่โพลีเปปไทด์ของโปรตีนดังกล่าวจะสร้าง β-helices และ β-folds อัตราส่วนระหว่างพวกเขาอาจแตกต่างกันมาก ตัวอย่างเช่น ที่ myoglobin(เพิ่มเติมเกี่ยวกับเรื่องนี้ในหน้า 28) มี 5 ส่วนที่เป็นเกลียวและไม่พับเดียว ในอิมมูโนโกลบูลิน (รายละเอียดเพิ่มเติมเกี่ยวกับหน้า 42) ในทางตรงกันข้ามองค์ประกอบหลักของโครงสร้างทุติยภูมิคือ -folds และ -helices หายไปทั้งหมด ในโครงสร้างข้างต้นของ phosphoglycerate kinase โครงสร้างทั้งสองประเภทจะแสดงเหมือนกันโดยประมาณ ในบางกรณี ดังที่เห็นได้ในตัวอย่างของ phosphoglycerate kinase สองส่วนหรือมากกว่าแยกจากกันในอวกาศ (แต่แน่นอนว่าเชื่อมต่อกันด้วยสะพานเปปไทด์) มองเห็นได้ชัดเจน - โดเมนบ่อยครั้ง บริเวณการทำงานที่แตกต่างกันของโปรตีนถูกแยกออกเป็นโดเมนที่ต่างกัน
โปรตีนไฟบริล โปรตีนเหล่านี้มีรูปร่างเป็นเส้นใยยาวและทำหน้าที่โครงสร้างในร่างกาย ในโครงสร้างหลัก พวกมันมีส่วนที่ทำซ้ำและสร้างโครงสร้างรองที่ค่อนข้างสม่ำเสมอสำหรับสายพอลิเปปไทด์ทั้งหมด ดังนั้นโปรตีน - ครีเอทีน (ส่วนประกอบโปรตีนหลักของเล็บ, ผม, ผิวหนัง) จึงถูกสร้างขึ้นจากเกลียวขยาย ไหมไฟโบรอินประกอบด้วยชิ้นส่วนที่ทำซ้ำเป็นระยะ Gly - Ala - Gly - Ser ทำให้เกิดรอยพับ มีองค์ประกอบทั่วไปน้อยกว่าของโครงสร้างทุติยภูมิ เช่น สายคอลลาเจนโพลีเปปไทด์ที่สร้าง เกลียวซ้ายด้วยพารามิเตอร์ที่แตกต่างจาก -helices อย่างมาก ในเส้นใยคอลลาเจน สายโซ่โพลีเปปไทด์แบบเฮลิคัลสามสายถูกบิดเป็นซุปเปอร์คอยล์ด้านขวาตัวเดียว:
4.4 โครงสร้างควอเทอร์นารี
ในกรณีส่วนใหญ่ สำหรับการทำงานของโปรตีน จำเป็นต้องมีพอลิเมอร์หลายสายรวมกันเป็นคอมเพล็กซ์เดียว คอมเพล็กซ์ดังกล่าวยังถือเป็นโปรตีนที่ประกอบด้วยหลายตัว หน่วยย่อย. โครงสร้างหน่วยย่อยมักปรากฏในวรรณคดีทางวิทยาศาสตร์เป็นโครงสร้างสี่ส่วน
โปรตีนที่ประกอบด้วยหน่วยย่อยหลายหน่วยมีการกระจายอย่างกว้างขวางในธรรมชาติ ตัวอย่างคลาสสิกคือโครงสร้างควอเทอร์นารีของเฮโมโกลบิน (รายละเอียดเพิ่มเติม - หน้า 26) หน่วยย่อยมักจะเขียนแทนด้วยตัวอักษรกรีก เฮโมโกลบินมีหน่วยย่อยสองและสองหน่วย การมีหน่วยย่อยหลายหน่วยมีความสำคัญต่อการใช้งาน - เพิ่มระดับความอิ่มตัวของออกซิเจน โครงสร้างควอเทอร์นารีของเฮโมโกลบินถูกกำหนดเป็น 2 2 .
โครงสร้างหน่วยย่อยเป็นลักษณะเฉพาะของเอ็นไซม์หลายชนิด โดยเฉพาะอย่างยิ่งเอ็นไซม์ที่ทำหน้าที่ที่ซับซ้อน ตัวอย่างเช่น RNA polymerase จาก อี. โคไลมีโครงสร้างหน่วยย่อย 2 " กล่าวคือ มันถูกสร้างขึ้นจากหน่วยย่อยสี่ประเภทที่แตกต่างกัน และ -หน่วยย่อยถูกทำซ้ำ โปรตีนนี้ทำหน้าที่ที่ซับซ้อนและหลากหลาย - เริ่มต้น DNA จับสารตั้งต้น - ไรโบนิวคลีโอไซด์ไตรฟอสเฟต และยังถ่ายโอนสารตกค้างของนิวคลีโอไทด์ไปสู่การเติบโต สายโซ่โพลีไรโบนิวคลีโอไทด์และหน้าที่อื่นๆ
การทำงานของโปรตีนหลายชนิดขึ้นอยู่กับสิ่งที่เรียกว่า ระเบียบ allosteric- สารประกอบพิเศษ (เอฟเฟคเตอร์) “ปิดสวิตช์” หรือ “เปิดสวิตช์” การทำงานของแอคทีฟเซ็นเตอร์ของเอ็นไซม์ เอนไซม์ดังกล่าวมีไซต์การจดจำเอฟเฟกต์พิเศษ แถมยังมีความพิเศษอีกด้วย หน่วยงานกำกับดูแลซึ่งรวมถึงส่วนที่ระบุ ตัวอย่างคลาสสิกคือเอนไซม์โปรตีนไคเนสที่กระตุ้นการถ่ายโอนกรดฟอสฟอริกที่ตกค้างจากโมเลกุล ATP ไปยังโปรตีนซับสเตรต
บทที่ 5 คุณสมบัติ
โปรตีนมีน้ำหนักโมเลกุลสูง บางชนิดสามารถละลายได้ในน้ำ สามารถบวมได้ มีลักษณะเฉพาะด้วยกิจกรรมทางแสง การเคลื่อนที่ในสนามไฟฟ้า และคุณสมบัติอื่นๆ บางอย่าง
โปรตีนมีส่วนร่วมอย่างแข็งขันในปฏิกิริยาเคมี คุณสมบัตินี้เกิดจากการที่กรดอะมิโนที่ประกอบเป็นโปรตีนมีกลุ่มการทำงานที่แตกต่างกันซึ่งสามารถทำปฏิกิริยากับสารอื่นได้ เป็นสิ่งสำคัญที่อันตรกิริยาดังกล่าวจะเกิดขึ้นภายในโมเลกุลโปรตีน ส่งผลให้เกิดการก่อตัวของเปปไทด์ ไฮโดรเจนซัลไฟด์ และพันธะประเภทอื่นๆ สารประกอบและอิออนต่างๆ สามารถเกาะกับอนุมูลของกรดอะมิโนได้ และด้วยเหตุนี้โปรตีนจึงช่วยให้ขนส่งผ่านเลือดได้
โปรตีนเป็นสารประกอบโมเลกุลขนาดใหญ่ เหล่านี้เป็นพอลิเมอร์ที่ประกอบด้วยโมโนเมอร์นับร้อยนับพันของกรดอะมิโน ตามนั้น และ มวลโมเลกุลโปรตีนอยู่ในช่วง 10,000 - 1,000,000 ดังนั้น ribonuclease (เอ็นไซม์ที่ทำลาย RNA) มีกรดอะมิโนตกค้าง 124 ตัว และมีน้ำหนักโมเลกุลประมาณ 14,000 Myoglobin (โปรตีนของกล้ามเนื้อ) ประกอบด้วย 153 กรดอะมิโนตกค้าง มีโมเลกุล น้ำหนัก 17,000 และเฮโมโกลบิน - 64,500 (574 กรดอะมิโนตกค้าง) น้ำหนักโมเลกุลของโปรตีนอื่น ๆ สูงกว่า: -โกลบูลิน (แบบฟอร์มแอนติบอดี) ประกอบด้วยกรดอะมิโน 1250 และมีน้ำหนักโมเลกุลประมาณ 150,000 และน้ำหนักโมเลกุลของเอนไซม์กลูตาเมตดีไฮโดรจีเนสเกิน 1,000,000
การกำหนดน้ำหนักโมเลกุลดำเนินการด้วยวิธีการต่างๆ เช่น ออสโมเมตริก การกรองเจล ออปติคัล ฯลฯ อย่างไรก็ตาม วิธีการตกตะกอนที่เสนอโดย T. Svedberg ที่แม่นยำที่สุดคือ โดยอาศัยข้อเท็จจริงที่ว่าในระหว่างการปั่นแยกเหวี่ยงด้วยความเข้มข้นสูงด้วยความเร่งสูงถึง 900,000 กรัม อัตราการตกตะกอนของโปรตีนจะขึ้นอยู่กับน้ำหนักโมเลกุลของโปรตีน
คุณสมบัติที่สำคัญที่สุดของโปรตีนคือความสามารถในการแสดงทั้งความเป็นกรดและเบส กล่าวคือทำหน้าที่เป็น แอมโฟเทอริกอิเล็กโทรไลต์ สิ่งนี้ทำให้มั่นใจได้โดยกลุ่มที่แยกตัวออกจากกันที่ประกอบขึ้นเป็นอนุมูลของกรดอะมิโน ตัวอย่างเช่น คุณสมบัติที่เป็นกรดของโปรตีนนั้นมาจากกลุ่มคาร์บอกซิลของกรดอะมิโนแอสปาร์ติกกลูตามิก และคุณสมบัติที่เป็นด่างนั้นมาจากอนุมูลของอาร์จินีน ไลซีน และฮิสทิดีน ยิ่งโปรตีนมีกรดอะมิโนไดคาร์บอกซิลิกมากเท่าไหร่ คุณสมบัติที่เป็นกรดของกรดก็จะยิ่งแรงขึ้นและในทางกลับกัน
กลุ่มเหล่านี้ยังมีประจุไฟฟ้าที่สร้างประจุโดยรวมของโมเลกุลโปรตีน ในโปรตีนที่กรดอะมิโนแอสปาร์ติกและกลูตามีนมีอิทธิพลเหนือ ประจุของโปรตีนจะเป็นลบ กรดอะมิโนพื้นฐานที่มากเกินไปจะให้ประจุบวกกับโมเลกุลโปรตีน เป็นผลให้ในสนามไฟฟ้า โปรตีนจะเคลื่อนที่ไปทางแคโทดหรือขั้วบวก ขึ้นอยู่กับขนาดของประจุทั้งหมด ดังนั้นในสภาพแวดล้อมที่เป็นด่าง (pH 7 - 14) โปรตีนจะบริจาคโปรตอนและกลายเป็นประจุลบ ในขณะที่ในสภาพแวดล้อมที่เป็นกรด (pH 1 - 7) การแตกตัวของกลุ่มกรดจะถูกระงับและโปรตีนจะกลายเป็นไอออนบวก
ดังนั้น ปัจจัยที่กำหนดพฤติกรรมของโปรตีนเป็นไอออนบวกหรือประจุลบคือปฏิกิริยาของตัวกลาง ซึ่งพิจารณาจากความเข้มข้นของไฮโดรเจนไอออนและแสดงโดยค่า pH อย่างไรก็ตาม ที่ค่า pH บางค่า จำนวนประจุบวกและประจุลบจะเท่ากัน และโมเลกุลจะกลายเป็นกลางทางไฟฟ้า กล่าวคือ มันจะไม่เคลื่อนที่ในสนามไฟฟ้า ค่า pH ของตัวกลางนี้ถูกกำหนดให้เป็นจุดไอโซอิเล็กทริกของโปรตีน ในกรณีนี้ โปรตีนอยู่ในสถานะที่เสถียรน้อยที่สุด และเมื่อ pH ของกรดหรือด่างเปลี่ยนแปลงไปเล็กน้อย โปรตีนก็จะตกตะกอนได้ง่าย สำหรับโปรตีนธรรมชาติส่วนใหญ่ จุดไอโซอิเล็กทริกอยู่ในสภาพแวดล้อมที่เป็นกรดเล็กน้อย (pH 4.8 - 5.4) ซึ่งบ่งชี้ถึงความเด่นของกรดอะมิโนไดคาร์บอกซิลิกในองค์ประกอบ
คุณสมบัติ amphoteric รองรับคุณสมบัติบัฟเฟอร์ของโปรตีนและการมีส่วนร่วมในการควบคุมค่า pH ของเลือด ค่า pH ของเลือดมนุษย์จะคงที่และอยู่ในช่วง 7.36 - 7.4 แม้ว่าจะมีสารต่างๆ ที่มีลักษณะเป็นกรดหรือเป็นด่าง ซึ่งให้อาหารเป็นประจำหรือเกิดขึ้นในกระบวนการเมตาบอลิซึม ดังนั้นจึงมีกลไกพิเศษในการควบคุมกรด-เบส ความสมดุลของสภาพแวดล้อมภายในร่างกาย ระบบดังกล่าวรวมถึงระบบที่พิจารณาในบทที่ ระบบบัฟเฟอร์เฮโมโกลบิน “การจำแนกประเภท” (หน้า 28) การเปลี่ยนแปลงค่า pH ของเลือดมากกว่า 0.07 บ่งชี้ถึงการพัฒนาของกระบวนการทางพยาธิวิทยา การเปลี่ยนแปลงของค่า pH ไปทางด้านกรดเรียกว่าภาวะกรด (acidosis) และด้านที่เป็นด่างเรียกว่าอัลคาโลซิส (alkalosis)
สิ่งที่สำคัญอย่างยิ่งสำหรับร่างกายคือความสามารถของโปรตีนในการดูดซับสารและไอออนบางชนิด (ฮอร์โมน วิตามิน เหล็ก ทองแดง) ซึ่งละลายได้ไม่ดีในน้ำหรือเป็นพิษ (บิลิรูบิน กรดไขมันอิสระ) โปรตีนขนส่งพวกมันผ่านทางเลือดไปยังสถานที่ที่มีการเปลี่ยนแปลงเพิ่มเติมหรือทำให้เป็นกลาง
สารละลายโปรตีนในน้ำมีลักษณะเฉพาะของตัวเอง ประการแรก โปรตีนมีความสัมพันธ์กับน้ำสูง กล่าวคือ พวกเขา ชอบน้ำซึ่งหมายความว่าโมเลกุลโปรตีน เช่น อนุภาคที่มีประจุ จะดึงดูดไดโพลของน้ำ ซึ่งอยู่รอบๆ โมเลกุลโปรตีนและก่อตัวเป็นน้ำหรือเปลือกไฮเดรต เปลือกนี้ปกป้องโมเลกุลโปรตีนจากการเกาะติดกันและตกตะกอน ขนาดของเปลือกน้ำขึ้นอยู่กับโครงสร้างของโปรตีน ตัวอย่างเช่น อัลบูมินจับกับโมเลกุลของน้ำได้ง่ายขึ้นและมีเปลือกน้ำที่ค่อนข้างใหญ่ ในขณะที่โกลบูลิน ไฟบริโนเจนจับกับน้ำได้แย่กว่า และเปลือกไฮเดรชั่นมีขนาดเล็กกว่า ดังนั้น ความคงตัวของสารละลายในน้ำของโปรตีนจึงถูกกำหนดโดยปัจจัยสองประการ: การปรากฏตัวของประจุบนโมเลกุลโปรตีนและเปลือกน้ำที่อยู่รอบๆ เมื่อปัจจัยเหล่านี้ถูกกำจัดออกไป โปรตีนจะตกตะกอน กระบวนการนี้สามารถย้อนกลับและย้อนกลับไม่ได้
...เอกสารที่คล้ายกัน
โปรตีน (โปรตีน) เป็นสารอินทรีย์ธรรมชาติที่มีโมเลกุลสูงและมีไนโตรเจน ซึ่งโมเลกุลเหล่านี้สร้างจากกรดอะมิโน โครงสร้างของโปรตีน การจำแนกประเภทของโปรตีน คุณสมบัติทางกายภาพและเคมีของโปรตีน หน้าที่ทางชีวภาพของโปรตีน เอนไซม์.
บทคัดย่อ เพิ่มเมื่อ 05/15/2007
ลักษณะสำคัญของกระบวนการเมตาบอลิซึม เมแทบอลิซึมและพลังงาน ลักษณะทั่วไป การจำแนกประเภท หน้าที่ องค์ประกอบทางเคมีและสมบัติของโปรตีน บทบาททางชีวภาพของพวกมันในการสร้างสิ่งมีชีวิต โปรตีนที่มีโครงสร้างและซับซ้อน วิธีการตกตะกอน
การนำเสนอ, เพิ่ม 04/24/2013
คุณสมบัติทางกายภาพและเคมี ปฏิกิริยาสีของโปรตีน องค์ประกอบและโครงสร้างหน้าที่ของโปรตีนในเซลล์ ระดับโครงสร้างโปรตีน ไฮโดรไลซิสของโปรตีน การขนส่ง และบทบาทการป้องกัน โปรตีนเป็นวัสดุก่อสร้างของเซลล์ ค่าพลังงานของมัน
บทคัดย่อ เพิ่มเมื่อ 06/18/2010
คุณสมบัติทางกายภาพ ชีวภาพ และเคมีของโปรตีน การสังเคราะห์และวิเคราะห์โปรตีน การกำหนดโครงสร้างหลัก ทุติยภูมิ อุดมศึกษา และควอเทอร์นารีของโปรตีน การเปลี่ยนสภาพ การแยก และการทำให้บริสุทธิ์ของโปรตีน การใช้โปรตีนในอุตสาหกรรมและยา
บทคัดย่อ เพิ่มเมื่อ 06/10/2015
โปรตีน - สารประกอบอินทรีย์น้ำหนักโมเลกุลสูง องค์ประกอบของกรดอะมิโน การกำหนดคุณสมบัติของโปรตีนโดยองค์ประกอบและโครงสร้างของโมเลกุลโปรตีน การกำหนดลักษณะหน้าที่หลักของโปรตีน ออร์แกเนลล์ของเซลล์และหน้าที่ของพวกมัน การหายใจของเซลล์และโครงสร้าง
ทดสอบเพิ่ม 06/24/2012
แนวคิดและโครงสร้างของโปรตีน กรดอะมิโนเป็นโมโนเมอร์ การจำแนกและพันธุ์ของกรดอะมิโน ลักษณะของพันธะเปปไทด์ ระดับการจัดระเบียบของโมเลกุลโปรตีน คุณสมบัติทางเคมีและทางกายภาพของโปรตีน วิธีการวิเคราะห์และหน้าที่ของโปรตีน
การนำเสนอ, เพิ่ม 04/14/2014
บทบาททางชีวภาพของน้ำ หน้าที่ของเกลือแร่ ไขมันที่เรียบง่ายและซับซ้อน ระดับของการจัดระเบียบของโปรตีน โครงสร้าง พลังงาน การจัดเก็บ และหน้าที่ควบคุมของไขมัน โครงสร้าง ตัวเร่งปฏิกิริยา มอเตอร์ ฟังก์ชันการขนส่งของโปรตีน
การนำเสนอ, เพิ่ม 05/21/2015
องค์ประกอบกรดอะมิโนของโปรตีนในสิ่งมีชีวิต บทบาทของรหัสพันธุกรรม ส่วนผสมของกรดอะมิโนมาตรฐาน 20 ชนิด การแยกโปรตีนออกเป็นกลุ่มโมเลกุลทางชีววิทยาที่แยกจากกัน โปรตีนที่ชอบน้ำและไม่ชอบน้ำ หลักการสร้างโปรตีนระดับองค์กร
งานสร้างสรรค์เพิ่มเมื่อ 11/08/2009
องค์ประกอบพื้นฐานและองค์ประกอบทางเคมีของเนื้อเยื่อของกล้ามเนื้อ ประเภทของโปรตีนของซาร์โคพลาสซึมและไมโอไฟบริล เนื้อหาต่อจำนวนโปรตีนทั้งหมด น้ำหนักโมเลกุล การกระจายในองค์ประกอบโครงสร้างของกล้ามเนื้อ หน้าที่และบทบาทของตนในร่างกาย โครงสร้างของโมเลกุลไมโอซิน
การนำเสนอ, เพิ่ม 12/14/2014
โปรตีนเป็นแหล่งอาหารหน้าที่หลัก กรดอะมิโนที่เกี่ยวข้องกับการสร้างโปรตีน โครงสร้างของสายโซ่โพลีเปปไทด์ การเปลี่ยนแปลงของโปรตีนในร่างกาย โปรตีนที่สมบูรณ์และไม่สมบูรณ์ โครงสร้างโปรตีน คุณสมบัติทางเคมี ปฏิกิริยาเชิงคุณภาพ
เป็นที่ทราบกันดีอยู่แล้วว่าสิ่งมีชีวิตขึ้นอยู่กับสารอินทรีย์ เช่น โปรตีน ไขมัน คาร์โบไฮเดรต และกรดนิวคลีอิก แต่ที่สำคัญที่สุดในบรรดาสารเหล่านี้คือโปรตีน
สารส่วนใหญ่ที่วิทยาศาสตร์รู้จักจะเปลี่ยนจากของแข็งเป็นของเหลวเมื่อถูกความร้อน แต่มีสารที่ในทางกลับกันกลายเป็นของแข็งเมื่อถูกความร้อน สารเหล่านี้ถูกรวมเข้าเป็นชั้นที่แยกจากกันโดยนักเคมีชาวฝรั่งเศสชื่อ Pierre Joseph Macke ในปี 1777 โดยการเปรียบเทียบกับไข่ขาวซึ่งจับตัวเป็นก้อนเมื่อถูกความร้อน สารเหล่านี้เรียกว่าโปรตีน โปรตีนเรียกอีกอย่างว่าโปรตีน ในภาษากรีกโปรตีน (proteios) หมายถึง "ในตอนแรก" ชื่อนี้มาจากโปรตีนในปี ค.ศ. 1838 เมื่อนักชีวเคมีชาวดัตช์เจอราร์ด มุลเดอร์เขียนว่าสิ่งมีชีวิตบนโลกจะเป็นไปไม่ได้หากไม่มีสารบางชนิด ซึ่งเป็นสารที่สำคัญที่สุดในบรรดาสารที่นักวิทยาศาสตร์รู้จักและจำเป็นต้องมีอยู่ในพืชทุกชนิดอย่างแน่นอน และสัตว์ Mulder เรียกสารนี้ว่าโปรตีน
โปรตีนเป็นสารที่ซับซ้อนที่สุดในบรรดาสารอาหารทั้งหมด ในทุกเซลล์ของร่างกายมนุษย์ ปฏิกิริยาเคมีเกิดขึ้น ซึ่งโปรตีนมีบทบาทสำคัญมาก
โปรตีนทำมาจากอะไร

โปรตีน ได้แก่ ไนโตรเจน ออกซิเจน ไฮโดรเจน คาร์บอน แต่สารอาหารอื่นๆ ไม่มีไนโตรเจน
โปรตีนเป็นพอลิเมอร์ตามธรรมชาติ โพลีเมอร์คือสารที่โมเลกุลประกอบด้วยอะตอมจำนวนมาก ย้อนกลับไปในศตวรรษที่ 19 นักเคมีชาวรัสเซีย Alexander Mikhailovich Butlerov ได้พิสูจน์ว่าหากโครงสร้างของโมเลกุลเปลี่ยนแปลงไป คุณสมบัติของสารก็จะเปลี่ยนไปด้วย โครงสร้างหลักของโปรตีนคือกรดอะมิโน และในโปรตีนนั้นมีกรดอะมิโนหลายชนิดรวมกัน ดังนั้นในธรรมชาติจึงมีโปรตีนหลากหลายชนิดที่มีคุณสมบัติต่างกัน ด้วยความช่วยเหลือของการวิจัยพบว่ามีกรดอะมิโนประมาณ 20 ชนิดที่เกี่ยวข้องกับการสร้างโปรตีน
กระบวนการสร้างโมเลกุลโปรตีนเป็นอย่างไร

กรดอะมิโนจะเกาะติดกันเป็นลำดับ อันเป็นผลมาจากกระบวนการนี้ทำให้เกิดสายโซ่ซึ่งเรียกว่าโพลีเปปไทด์ ต่อจากนั้น โพลีเปปไทด์อาจม้วนหรือสร้างรูปร่างอื่น คุณสมบัติของโปรตีนขึ้นอยู่กับองค์ประกอบของกรดอะมิโน จำนวนกรดอะมิโนที่เกี่ยวข้องกับการสังเคราะห์ และลำดับที่กรดอะมิโนเหล่านี้เกาะติดกัน ตัวอย่างเช่น การสังเคราะห์โปรตีนสองชนิดเกี่ยวข้องกับกรดอะมิโนจำนวนเท่ากัน ซึ่งมีองค์ประกอบเหมือนกันด้วย แต่ถ้ากรดอะมิโนเหล่านี้อยู่ในลำดับที่ต่างกัน เราก็จะได้โปรตีนสองชนิดที่ต่างกันโดยสิ้นเชิง
หากเปปไทด์มีกรดอะมิโนตกค้างไม่เกิน 15 ชนิด เรียกว่าโอลิโกเปปไทด์ และเปปไทด์ที่มีกรดอะมิโนตกค้างมากถึงหลายหมื่นหรือแม้แต่หลายแสนตัวนั้นเรียกว่าโปรตีน โมเลกุลโปรตีนมีโครงสร้างเชิงพื้นที่ที่กะทัดรัด โครงสร้างนี้อาจอยู่ในรูปของเส้นใย โปรตีนดังกล่าวเรียกว่าไฟบริลลาร์ พวกเขากำลังสร้างโปรตีน หากโมเลกุลโปรตีนมีโครงสร้างเป็นลูกกลม โปรตีนจะเรียกว่าทรงกลม โปรตีนเหล่านี้รวมถึงเอ็นไซม์ แอนติบอดี และฮอร์โมนบางชนิด
ขึ้นอยู่กับกรดอะมิโนที่รวมอยู่ในองค์ประกอบของโปรตีน โปรตีนมีความสมบูรณ์และไม่สมบูรณ์ โปรตีนที่สมบูรณ์ประกอบด้วยกรดอะมิโนครบชุด โปรตีนที่ไม่สมบูรณ์ กรดอะมิโนบางชนิดขาดหายไป
โปรตีนยังแบ่งออกเป็นแบบง่ายและซับซ้อน โปรตีนอย่างง่ายประกอบด้วยกรดอะมิโนเท่านั้น องค์ประกอบของโปรตีนที่ซับซ้อนนอกเหนือจากกรดอะมิโนยังรวมถึงโลหะ, คาร์โบไฮเดรต, ไขมัน, กรดนิวคลีอิก
บทบาทของโปรตีนในร่างกายมนุษย์

โปรตีนทำหน้าที่ต่าง ๆ ในร่างกายมนุษย์
1.โครงสร้าง. โปรตีนเป็นส่วนหนึ่งของเซลล์ของเนื้อเยื่อและอวัยวะทั้งหมด
2. ป้องกัน. โปรตีนอินเตอร์เฟอรอนถูกสังเคราะห์ในร่างกายเพื่อป้องกันไวรัส
3. ดวิกาเทลนาฉัน. โปรตีนไมโอซินเกี่ยวข้องกับกระบวนการหดตัวของกล้ามเนื้อ
4. ขนส่ง.เฮโมโกลบินซึ่งเป็นโปรตีนในองค์ประกอบของเซลล์เม็ดเลือดแดงเกี่ยวข้องกับการถ่ายเทออกซิเจนและคาร์บอนไดออกไซด์
5. พลังงานฉัน. อันเป็นผลมาจากการเกิดออกซิเดชันของโมเลกุลโปรตีน พลังงานที่จำเป็นสำหรับชีวิตของร่างกายจึงถูกปล่อยออกมา
6. ตัวเร่งปฏิกิริยาฉัน. เอนไซม์โปรตีนทำหน้าที่เป็นตัวเร่งปฏิกิริยาทางชีวภาพที่เพิ่มอัตราการเกิดปฏิกิริยาเคมีในเซลล์
7. ระเบียบข้อบังคับฉัน. ฮอร์โมนควบคุมการทำงานของร่างกายต่างๆ ตัวอย่างเช่น อินซูลินควบคุมระดับน้ำตาลในเลือด
ในธรรมชาติมีโปรตีนจำนวนมากที่สามารถทำหน้าที่ได้หลากหลาย แต่หน้าที่ที่สำคัญที่สุดของโปรตีนคือการรักษาชีวิตบนโลกร่วมกับชีวโมเลกุลอื่นๆ
 ลุงวันยา พล็อตเรื่อง “ลุงอีวาน ทัศนคติต่ออาจารย์ของผู้อื่น
ลุงวันยา พล็อตเรื่อง “ลุงอีวาน ทัศนคติต่ออาจารย์ของผู้อื่น Tsakhes น้อยชื่อเล่น Zinnober
Tsakhes น้อยชื่อเล่น Zinnober Maikov, Apollon Nikolaevich - ชีวประวัติสั้น
Maikov, Apollon Nikolaevich - ชีวประวัติสั้น