คุณสมบัติทางสรีรวิทยาของศักยภาพการทำงานของเนื้อเยื่อที่กระตุ้นได้ ความตื่นเต้นง่ายคืออะไร? เวลาที่เป็นประโยชน์คืออะไร chronaxy คืออะไร
การขนส่งแบบพาสซีฟ การแพร่และการออสโมซิส การแพร่คือการเคลื่อนที่ของโมเลกุลหรือไอออนจากบริเวณที่มีความเข้มข้นสูงไปยังบริเวณที่มีความเข้มข้นต่ำ การแพร่จะหยุดลงเมื่อความเข้มข้นของโมเลกุลหรือไอออนทั้งสองด้านของเมมเบรนเท่ากัน การขนส่งแบบพาสซีฟไม่ต้องการพลังงาน 1. ความเข้มของการแพร่กระจายผ่านเมมเบรนขึ้นอยู่กับความแตกต่างของความเข้มข้นของสารทั้งสองด้านของเมมเบรน (บนการไล่ระดับความเข้มข้น) กับการซึมผ่านของพลาสมาเมมเบรนของเซลล์สำหรับการแพร่กระจายของโมเลกุล 2. อัตราการแพร่ผ่านเมมเบรนเป็นสัดส่วนโดยตรงกับพื้นที่ผิวของเมมเบรนและขึ้นอยู่กับอุณหภูมิของสารละลาย 3. Simple diffusion เป็นการขนส่งแบบพาสซีฟซึ่งโมเลกุลขนาดเล็กและไอออนอนินทรีย์สามารถผ่านพลาสมาเมมเบรนของเซลล์ได้อย่างอิสระ 4. ไอออนอนินทรีย์ - เช่น Na + และ K + ผ่านช่องเฉพาะ (เลือกหรือเลือก) ที่อยู่ในเยื่อหุ้มเซลล์ 5. สเตอรอยด์ฮอร์โมนหรือสารประกอบไขมันอื่น ๆ สามารถผ่านชั้นฟอสโฟลิพิดของเมมเบรนได้โดยตรงโดยการแพร่กระจายอย่างง่าย 6. ออสโมซิสคือการแพร่กระจายอย่างง่ายของโมเลกุลของน้ำผ่านเยื่อหุ้มเซลล์ 7. โมเลกุลของน้ำจะเคลื่อนที่จากสารละลายที่มีความเข้มข้นน้อยกว่า (นั่นคือ มีปริมาณน้ำสูง) ไปยังสารละลายที่มีความเข้มข้นสูงกว่า (นั่นคือ มีปริมาณน้ำน้อยกว่า) ขนาดของออสโมซิสขึ้นอยู่กับความแตกต่างของความเข้มข้นของสารละลาย แต่ไม่ได้ขึ้นอยู่กับองค์ประกอบทางเคมี 8. ความเข้มข้นของสารที่ละลายทั้งหมด (เป็นโมลต่อน้ำหนึ่งลิตร) วัดเป็นหน่วยของออสโมลาลิตี้ สารละลายที่มีออสโมลิตีสูงกว่าจะมีแรงดันออสโมติกสูงกว่า 9. น้ำเคลื่อนที่โดยการออสโมซิสจากสารละลายที่มีออสโมติกต่ำและแรงดันออสโมติกต่ำไปยังสารละลายที่มีออสโมติกสูงและแรงดันออสโมติกสูง พลาสมาเมมเบรนของเซลล์ที่กระตุ้นได้ เยื่อหุ้มพื้นผิวทางชีวภาพสร้างเปลือกนอกของเซลล์ที่กระตุ้นทั้งหมด ปัจจุบันมีการใช้แบบจำลองโมเสกของไหลเพื่ออธิบายโครงสร้างของเยื่อหุ้มเซลล์ผิว 1. เยื่อหุ้มเซลล์ประกอบด้วยโมเลกุลฟอสโฟลิพิดสองชั้น โดยที่ปลายของโมเลกุลที่ไม่ชอบน้ำจะหันเข้าหาด้านในของ bilayer และปลายที่ชอบน้ำจะหันเข้าหาเฟสที่เป็นน้ำ โมเลกุลของโปรตีนอยู่ใน bilayer: พื้นผิวเป็นตัวรับและส่วนประกอบสำคัญคือช่องไอออนและปั๊มไอออน 2. คุณสมบัติทางไฟฟ้าของเมมเบรน: 1. ความจุเป็นหน้าที่ของ Bilayer ฟอสโฟลิพิด ซึ่งจะแยกและสะสมประจุที่ทั้งสองด้านของเมมเบรน 2. การนำไฟฟ้าคือส่วนกลับของความต้านทานไฟฟ้าของเมมเบรนที่กระตุ้นได้ 3. การนำไฟฟ้าของเยื่อชีวภาพเป็นหน้าที่ของช่องไอออน การนำไฟฟ้าขึ้นอยู่กับ: 1) ความแตกต่างของความเข้มข้นของไอออนทั้งสองด้านของเมมเบรน 2) ความชุ่มชื้นและเส้นผ่านศูนย์กลางของไอออน 3) การเคลื่อนที่ของไอออน และ 4) ความหนาของเมมเบรน 4. ช่องไอออนแบ่งออกเป็นแบบเลือก (นำไอออนเพียงตัวเดียว - Na2+, K+, Ca2+ หรือ Cl-) และแบบไม่เลือก ตามกลไกการเปิดใช้งาน พวกมันถูกแบ่งออกเป็น 1) กระตุ้นด้วยไฟฟ้าหรือขึ้นกับแรงดันไฟฟ้า (เปิดเพื่อตอบสนองต่อการกระตุ้นด้วยไฟฟ้า), 2) กระตุ้นด้วยคีโมหรือควบคุมตัวรับ (ขึ้นอยู่กับลิแกนด์ สำหรับการเปิดใช้งานจำเป็นต้องผูกมัด ตัวรับซึ่งอยู่ภายในช่องทางโดยมีผู้ไกล่เกลี่ยทางเคมี - ผู้ไกล่เกลี่ย ) และ 3) กลไกที่กระตุ้นได้ (ยืด - ช่องทางกระตุ้นเฉพาะสำหรับการเปิดใช้งานของพวกเขากำลังยืดออก) การขนส่งด้วยผู้ให้บริการ 1. การขนส่งกลูโคส กรดอะมิโน และโมเลกุลมีขั้วอื่นๆ ผ่านพลาสมาเมมเบรนนั้นอาศัยสื่อกลางโดยโปรตีนพาหะที่อยู่ในเยื่อหุ้มเซลล์ และเรียกว่าการแพร่แบบอำนวยความสะดวก 2. Transporter-mediated transport มีความเฉพาะเจาะจง การขนส่งถึงขีดสุดเมื่อผู้ให้บริการถึงสถานะอิ่มตัว 3. เช่นเดียวกับการแพร่กระจายอย่างง่าย การแพร่แบบอำนวยความสะดวกคือการขนส่งแบบพาสซีฟที่ไม่ต้องการการใช้พลังงานของเซลล์ การขนส่งที่ใช้งานอยู่ 1. การขนส่งโมเลกุลและอิออนทั่วเยื่อหุ้มเซลล์ต้องใช้พลังงานเซลล์ (ATP) 2. ในกระบวนการขนส่งแบบแอคทีฟ โมเลกุลพาหะจะถ่ายโอนโมเลกุลและไอออนจากบริเวณที่มีความเข้มข้นต่ำไปยังบริเวณที่มีความเข้มข้นสูง 3. ตัวอย่างที่รู้จักกันดีที่สุดของการขนส่งแบบแอกทีฟหลักคือปั๊ม Na+/K+ ความเข้มข้นของโซเดียมไอออนจะมากกว่าในสภาพแวดล้อมนอกเซลล์ - ที่ด้านนอกของเมมเบรน ในขณะที่โพแทสเซียมไอออนจะมากกว่าภายในเซลล์ การทำงานของปั๊ม Na+/K+ ช่วยรักษาเกรเดียนต์ของความเข้มข้นนี้โดยการขนส่งไอออน Na+ ออกไปด้านนอกและไอออน K+ เข้าไปในเซลล์เทียบกับเกรเดียนต์ของความเข้มข้น 4. การขนส่งที่ใช้งานทุติยภูมิ - การขนส่งสารโดยมีส่วนร่วมของโปรตีนพาหะตามการไล่ระดับสีซึ่งสร้างขึ้นโดยการทำงานของปั๊ม Na + / K + 5. ปั๊ม Ca2+ มีอยู่ในเซลล์ส่วนใหญ่: ชนิด PMCA อยู่ที่พลาสมาเมมเบรน และชนิด SERCA อยู่ที่เยื่อหุ้มเซลล์ซาร์โคพลาสมิกเรติคูลัม 6. บนเยื่อหุ้มยอดของเซลล์ข้างขม่อมของเยื่อบุกระเพาะอาหารในเยื่อบุผิวของไตและเยื่อบุลำไส้มี H + -K + -pump 7. เยื่อหุ้มของออร์แกเนลล์ภายในเซลล์ประกอบด้วยปั๊ม H + (ประเภทสุญญากาศ) ศักยภาพของเยื่อพัก (RRP) 1. ไซโตพลาสซึมของเซลล์ที่อยู่ด้านในของเยื่อหุ้มมีประจุลบเนื่องจากประจุลบ (อนินทรีย์และอินทรีย์) ซึ่งไม่สามารถออกจากเซลล์ได้ และมีประจุบวกอยู่ด้านนอกเนื่องจากไอออนบวก 2. ไอออนบวก K+ อนินทรีย์สามารถผ่านเยื่อหุ้มเซลล์ได้อย่างอิสระเมื่อพัก มันเคลื่อนจากบริเวณที่มีความเข้มข้นสูง (จากภายในเซลล์) ไปยังบริเวณที่มีความเข้มข้นต่ำ (บนผิวนอกของเยื่อหุ้มเซลล์) ผ่านช่องไอออน นี่คือองค์ประกอบ "เคมี" ของประจุบนเมมเบรน 3. ประจุลบที่เหลืออยู่ภายในเซลล์และไอออนบวกที่สะสมอยู่ภายนอกเยื่อหุ้มเซลล์จะสร้างสนามไฟฟ้า และไอออน K + จะเริ่มเคลื่อนที่ในสนามนี้ ไอออนภายในเซลล์จะดึงดูดไอออน K+ ที่เป็นบวก (องค์ประกอบ "ไฟฟ้า") และตามการไล่ระดับความเข้มข้น ไอออน K+ มักจะออกจากเซลล์ 4. ในขณะที่อิทธิพลของสนามไฟฟ้าถูกชดเชยด้วยแรงดันการแพร่กระจาย (เนื่องจากความแตกต่างของความเข้มข้น) จะเกิดสมดุลไฟฟ้าเคมี ที่จุดสมดุล กระแสไอออน K+ ออก (กระแสออก) และภายในเซลล์ (กระแสเข้า) มีค่าเท่ากัน 5. มีการแยกประจุบนพื้นผิวของเมมเบรน - พื้นผิวด้านในจะกลายเป็นค่าลบมากขึ้นเมื่อเทียบกับด้านนอก 6. ในขณะที่สมดุลภายในเซลล์ คุณสามารถลงทะเบียนประจุลบได้เท่ากับ -90 mV ความต่างศักย์นี้เรียกว่าศักย์สมดุลสำหรับ K+ (Ek) ซึ่งสามารถหาได้โดยใช้สมการ Nernst 7. ศักย์ของเยื่อหุ้มเซลล์หรือศักย์ของเยื่อพักตัวนั้นจริง ๆ แล้วน้อยกว่า Ek เล็กน้อย (ปกติจะอยู่ที่ -65 mV ถึง -80 mV) เนื่องจากไอออน Na + บางส่วนสามารถเข้าสู่เซลล์ขณะพักได้ 8. ความเข้มข้นของ Na + ไอออนที่เหลือภายนอกเซลล์มีมากกว่าภายใน และพื้นผิวภายในเซลล์มีประจุลบ ประจุลบดึงดูดไอออน Na+ 9. ในกรณีที่ไม่มีการกระตุ้น กระแสขาออกเล็กน้อยของ K+ จะมาพร้อมกับกระแสขาเข้าเล็กน้อยของ Na+ กระแสที่เข้ามาของไอออน Na+ นั้นมีปริมาณน้อย เนื่องจากเยื่อหุ้มเซลล์แทบจะไม่ยอมให้ไอออนของ Na+ ผ่านเข้าไปได้ 10. การไล่ระดับความเข้มข้นสำหรับ Na+ และ K+ ไอออน และดังนั้น ศักยภาพของเยื่อพักตัว ได้รับการดูแลโดยการทำงานของปั๊ม Na+ /K+ ซึ่งทำให้แน่ใจได้ว่ามีการปลดปล่อยไอออน Na+ 3 ไอออนออกจากเซลล์และโพแทสเซียมไอออน 2 ไอออนเข้าไปในเซลล์พร้อมกัน เซลล์ ปั๊ม Na+/K+ ต้องการพลังงาน ATP (การลำเลียงแบบแอคทีฟ) 11. การทำงานของเครื่องสูบน้ำ Na+ /K+ ทำหน้าที่แยกตัวของมันเองไปยังศักย์ไฟฟ้าของเมมเบรน เนื่องจากเครื่องสูบน้ำจะกำจัดไอออน Na+ ออกจากเซลล์มากกว่าที่จะดึงไอออน K+ ออกมา เนื่องจากการทำงานของปั๊ม พื้นผิวด้านในของเซลล์จะกลายเป็นลบมากยิ่งขึ้น ดังนั้นปั๊มดังกล่าวจึงถูกเรียกว่าปั๊มไฟฟ้า ศักยภาพในการดำเนินการ (AP) 1. การซึมผ่านของเยื่อหุ้มเซลล์สำหรับไอออนนั้นเกิดจากการมีช่องไอออน - ขึ้นอยู่กับแรงดันไฟฟ้าและขึ้นอยู่กับลิแกนด์ 2. ในการตอบสนองต่อการกระตุ้นด้วยไฟฟ้า - เมมเบรนดีโพลาไรเซชัน ช่อง Na + ที่ขึ้นกับแรงดันไฟฟ้าจะเปิดขึ้น 3. เมื่อเมมเบรนถูกดีโพลาไรซ์ถึงระดับเกณฑ์ - ระดับวิกฤตของดีโพลาไรซ์ (CDL) - ช่องโซเดียมทั้งหมดจะเปิดขึ้น 4. การเปิดช่องที่ขึ้นอยู่กับแรงดันไฟฟ้านำไปสู่การสร้างศักยภาพในการดำเนินการ - AP การแพร่ของไอออน Na+ เข้าไปในเซลล์ทำให้เกิดการสลับขั้วที่มากขึ้นของเยื่อหุ้มเซลล์ และการแพร่กระจายของ Na+ เข้าไปในเซลล์มากขึ้น ซึ่งเป็นการดีโพลาไรเซชันที่เกิดขึ้นเองอย่างยั่งยืน (สร้างใหม่) ตามประเภทของการป้อนกลับเชิงบวก 5. กระแสโซเดียมที่เข้ามานำไปสู่การพลิกกลับของ MPP ระหว่างการสลับขั้ว - จาก -70 mV ถึง + 30 mV ในขณะนี้ ประจุภายในเซลล์จะกลายเป็นบวกเป็นเวลา 1-2 มิลลิวินาที (โอเวอร์ชูต) 6. ถัดไป ช่อง Na+ จะปิดใช้งานและปิด ในเวลาเดียวกัน การแพร่กระจายของไอออน K+ ออกไปภายนอกผ่านช่องโพแทสเซียมแบบเปิดจะทำให้ระดับ MPP กลับคืนสู่ระดับเดิม PD ระยะนี้เรียกว่าการรีโพลาไรเซชัน 7. เมื่อความเข้มข้นของไอออน Na + เพิ่มขึ้นภายในเซลล์ การทำงานของปั๊ม Na + / K + จะทำงาน ซึ่งจะปั๊มโซเดียมไอออนออกและปั๊มไอออนโพแทสเซียมเข้าไปในเซลล์พร้อมกัน ฟื้นฟูระดับ MPP เริ่มต้นอย่างรวดเร็ว . การส่งแรงกระตุ้นผ่านทางแยกประสาทและกล้ามเนื้อ (ไซแนปส์) ไซแนปส์เป็นการติดต่อพิเศษระหว่างสองเซลล์ที่ทำหน้าที่ส่งสิ่งกระตุ้น ในระบบประสาทส่วนกลางมีการสัมผัสระหว่างเซลล์ประสาท 2 เซลล์ ส่วนรอบนอกมีการสัมผัสระหว่างเซลล์ประสาทและกล้ามเนื้อ ในหัวใจและกล้ามเนื้อเรียบมีการสัมผัสระหว่างเซลล์กล้ามเนื้อ 2 เซลล์ 1. ไซแนปส์ประกอบด้วยส่วนพรีซินแนปติก รอยแหว่งไซแนปติก และส่วนหลังไซแนปส์ 2. ตามกลไกการส่งผ่านของการกระตุ้น synapses จะแบ่งออกเป็นไฟฟ้าและเคมี 3. Gap-junction ที่พบในกล้ามเนื้อหัวใจและกล้ามเนื้อเรียบ และใน dendro-dendritic synapses ของสมองบางส่วน เป็นไซแนปส์ไฟฟ้า 4. ศักยะงานในไซแนปส์ไฟฟ้าแพร่กระจายโดยตรงจากพรีนาไปยังเยื่อหุ้มโพสซินแนปติก การนำการกระตุ้นในไซแนปส์ไฟฟ้าเป็นแบบทวิภาคี 5. ใน synapses ทางเคมี การกระตุ้นจะดำเนินการในทิศทางเดียวเท่านั้น (จากส่วน presynaptic ไปยังส่วน postynaptic) 6. ในไซแนปส์ทางเคมี เยื่อพรีไซแนปติกประกอบด้วยสารสื่อประสาท (หรือสื่อกลาง) ที่บรรจุอยู่ในถุงหรือถุงซินแนปติก โมเลกุลของผู้ไกล่เกลี่ยถูกปล่อยออกจากถุงเข้าไปใน synaptic cleft โดย exocytosis 7. ตัวกลางเรียกว่าลิแกนด์ของตัวรับที่สอดคล้องกัน และตัวรับที่อยู่บนเยื่อหุ้มโพสซินแนปติกของไซแนปส์เรียกว่าลิแกนด์ขึ้นอยู่กับ (ขึ้นอยู่กับคีโมหรือตัวรับที่ควบคุม) 8. การจับตัวกลางกับรีเซพเตอร์เมมเบรนโพสซินแนปติกนำไปสู่การเปิดของช่องไอออนที่อยู่ภายในรีเซพเตอร์ (ตัวรับไอโอโนโทรปิก) หรือผ่านการเปิดใช้งานจีโปรตีน ช่องไอออนที่อยู่ถัดจากรีเซพเตอร์จะเปิดขึ้น ( เมตาบอโทรปิกรีเซพเตอร์) 9. ในไซแนปส์ประสาทและกล้ามเนื้อ ผู้ไกล่เกลี่ยคือ acetylcholine (ACh) 10. มีตัวรับ cholinergic สองประเภท - นิโคตินิกและมัสคารินิก ตัวรับ cholinergic ชนิด Nicotinic อยู่ที่เยื่อหุ้มเซลล์แบบโพสต์ซินแนปติกของกล้ามเนื้อโครงร่าง 11. เมื่อ 2 โมเลกุล ACh จับกับตำแหน่งพิเศษบนตัวรับ nicotinic cholinergic ช่องไอออนจะเปิดขึ้นสำหรับ Na + ไอออน ไอออน Na+ เข้าสู่เซลล์ตามการไล่ระดับความเข้มข้น เกิดเป็นกระแสโซเดียมที่เข้ามา 12. สิ่งนี้นำไปสู่การสลับขั้วเล็กน้อยของเมมเบรนแบบโพสต์ซินแนปติกและการเกิดขึ้นของการตอบสนองเฉพาะที่ - ศักย์แผ่นท้าย (EPP) การสลับโพลาไรเซชันขนาดเล็กดังกล่าวเกิดจากความจริงที่ว่าช่องไอออนของกล้ามเนื้อโครงร่าง (หรือแผ่นท้าย) ไม่มีการเลือก 13. เมื่อแอมพลิจูดของการตอบสนองในท้องถิ่นถึงระดับเกณฑ์ ช่องโซเดียมแบบเลือกได้เร็วจะเปิดขึ้นในบริเวณใกล้กับซินแนปติก ส่งผลให้เกิดการสร้าง AP 14. หลังจากเปิดใช้งานตัวรับ cholinergic แล้ว ACh จะถูกแยกออกโดยเอนไซม์ acetylcholinesterase (AChE) ให้เป็นโคลีนและกรดอะซิติก โคลีนเข้าสู่ขั้ว presynaptic ผ่านระบบ reuptake สารตกค้างของกรดอะซิติกจะค่อยๆ กระจายเข้าไปในช่องเพอริซินแนปติกและทำให้มันเป็นกรด กล้ามเนื้อโครงร่าง. กล้ามเนื้อโครงร่างประกอบด้วยเส้นใยกล้ามเนื้อ (เซลล์กล้ามเนื้อ) ที่เชื่อมต่อกันด้วยเนื้อเยื่อเกี่ยวพันและยึดติดกับกระดูกด้วยเส้นเอ็น เส้นใยกล้ามเนื้อที่แยกจากกันถูกปกคลุมด้วยเอนโดไมเซียม กลุ่มของเส้นใยกล้ามเนื้อถูกปกคลุมด้วย perimysium และกล้ามเนื้อทั้งหมดถูกปกคลุมด้วย epimysium 1. เส้นใยกล้ามเนื้อโครงร่างถูกเรียกว่า striated เนื่องจากภายใต้กล้องจุลทรรศน์แบบใช้แสงทั่วไป เส้นใยเหล่านี้จะแสดงแสงสลับ (isotropic, I) และแผ่นมืด (anisotropic, A) 2. ตรงกลางของดิสก์ไอโซทรอปิกแต่ละอันจะมีเส้น Z ซึ่งติดอยู่กับเส้นใยแอกติน 3. การหดตัวของเส้นใยกล้ามเนื้อในร่างกายถูกควบคุมโดยเซลล์ประสาทสั่งการของระบบประสาทโซมาติก เซลล์ประสาทสั่งการและเส้นใยกล้ามเนื้อที่แอกซอนของเซลล์ประสาทสั่งการนี้ก่อตัวเป็นหน่วยสั่งการ (MU) ซึ่งเป็นหน่วยการทำงานของกล้ามเนื้อโครงร่าง 4. หนึ่งแอกซอนของเซลล์ประสาทสั่งการ (motoneuron) สามารถสร้างเส้นใยกล้ามเนื้อได้ตั้งแต่ 10 ถึง 1,000 เส้น จำนวนเส้นใยกล้ามเนื้อที่เกิดจากเซลล์ประสาทสั่งการหนึ่งเซลล์จะแตกต่างกันไปขึ้นอยู่กับการทำงานเฉพาะของกล้ามเนื้อนั้นๆ 5. ยิ่งเส้นใยกล้ามเนื้อเข้าสู่ชุดมอเตอร์น้อยลง หรือเซลล์ประสาทสั่งการทำหน้าที่กล้ามเนื้อนี้มากขึ้น การเคลื่อนไหวที่แม่นยำก็สามารถทำได้มากขึ้น (เช่น กล้ามเนื้อนิ้วมือ) และในทางกลับกัน ใยกล้ามเนื้อยิ่งเข้าสู่ชุดมอเตอร์มากขึ้นเท่านั้น การเคลื่อนไหวของกล้ามเนื้อจะแตกต่างกันน้อยลง (เช่น rectus abdominis) 6. การหดตัวต่อเนื่องเป็นเวลานานเกิดจากการกระตุ้นแบบอะซิงโครนัสของชุดมอเตอร์ต่างๆ กลไกการหดตัวของกล้ามเนื้อ 1. ซาร์โคเมียร์เป็นหน่วยการทำงานของเครื่องมือหดตัวของไมโอไฟบริล ซึ่งรวมถึงไมโอฟิลาเมนท์ (บางและหนา) และถูกจำกัดโดยเส้น Z สองเส้นที่อยู่ติดกัน 2. เส้นใยหนา (ด้าย) ประกอบด้วย myosin, เส้นใยบาง - จาก F-actin เส้นใยแอกตินติดอยู่กับเส้น Z H-band ส่วนกลางของซาร์โคเมียร์ประกอบด้วยเส้นใยไมโอซิน 3. เส้นใยบาง ๆ เป็นด้ายคู่ที่บิดเป็นก้นหอย บนเกลียวของ F-actin เป็นเกลียวของโมเลกุลโปรตีน tropomyosin ด้วยขั้นตอน 40 นาโนเมตร โมเลกุลโปรตีนโทรโปนินจะติดอยู่กับโมเลกุลโทรโปไมโอซิน 4. ในขณะพัก tropomyosin จะป้องกันไม่ให้หัวของ myosin จับกับบริเวณที่จับบนเส้นใยแอกติน 5. เส้นใยไมโอซินหนากว่าเส้นใยแอกติน เนื่องจากมีน้ำหนักโมเลกุลสูงกว่า ที่ด้านข้างของเส้นใย myosin มีส่วนยื่นออกมา - สะพานตามขวาง สะพานขวางประกอบด้วยส่วนหัวและส่วนที่เคลื่อนย้ายได้ (ส่วนบานพับ) - คอ 6. ทฤษฎีของเส้นใยเลื่อนอธิบายกลไกการก่อตัวของสะพานขวางระหว่างแอกตินและไมโอซินและกระบวนการเลื่อนของเส้นใยไมโอซินที่สัมพันธ์กัน 7. วัฏจักรของการก่อตัวของสะพานข้ามเริ่มต้นด้วยสถานะคงที่ซึ่งหัว myosin เชื่อมต่อกับตำแหน่งที่ใช้งานบนเส้นใยแอกตินที่มุม 45o 8. โมเลกุล ATP จับกับหัว myosin ซึ่งนำไปสู่การแยกของหัว myosin ออกจากเส้นใยแอกติน กิจกรรม ATP-ase ของหัว myosin เพิ่มขึ้น ATP จะถูกไฮโดรไลซ์เป็น ADP และอนินทรีย์ฟอสเฟต หัว myosin จะเปิดขึ้น บริเวณบานพับและยึดติดกับจุดศูนย์กลางที่ใช้งานถัดไปบนเส้นใยแอกตินที่มุม 90o 9. เมื่อฟอสเฟตอนินทรีย์หลุดออกจากส่วนหัวของไมโอซิน จะเกิดการเคลื่อนไหวแบบกวาดซึ่งเคลื่อนเส้นใยแอกตินไปยังศูนย์กลางของซาร์โคเมียร์ 11 นาโนเมตร หลังจากนั้นให้ตั้งหัวไมโอซินที่มุม 45o จากนั้นโมเลกุล ADP จะถูกแยกออกและระบบจะกลับสู่สถานะเดิมที่เสถียร 10. หลังจากการเติมโมเลกุล ATP ถัดไป หัวไมโอซินจะแยกออกจากเส้นใยแอกติน และวงจรใหม่ของการสร้างสะพานข้ามและการกรรเชียงจะเริ่มขึ้น 11. ปริมาณ ATP ที่ลดลงและความเป็นไปไม่ได้ที่จะแยกหัว myosin ออกจากเส้นใยแอกตินทำให้เกิดความแข็งแกร่งของกล้ามเนื้อหลังการตาย 12. อินเทอร์เฟซระบบเครื่องกลไฟฟ้า ขณะพัก ความเข้มข้นของไอออน Ca2+ ในซาร์โคพลาสซึมจะต่ำ ดังนั้นหัวไมโอซินจึงไม่สามารถเกาะกับแอกตินได้ Ca2+ ไอออนถูกปั๊มเข้าไปใน sarcoplasmic reticulum (SR) โดย Ca2+-ATPase (Ca2+pump) 13. ส่วนปลายของเซลล์ประสาทสั่งการจะปล่อยอะซิติลโคลีน ซึ่งนำไปสู่การสร้าง PKP และ PD 14. ศักย์ไฟฟ้าแพร่กระจายไปตามเมมเบรน T-tubule ไปถึงเมมเบรน SPR และสลับขั้ว ช่อง Ca2+ ที่อยู่บนเยื่อหุ้มเซลล์ SPR เปิดออก ซึ่งนำไปสู่การแพร่ของ Ca2+ จำนวนมากเข้าไปในซาร์โคพลาสซึม ความเข้มข้นของ Ca2+ ในซาร์โคพลาสซึมเพิ่มขึ้นจาก 1x10-7 M - ขณะหยุดนิ่ง เป็น 1x10-4 M - ระหว่างการกระตุ้น 15. จากนั้น ไอออน Ca2+ จับกับโทรโปนิน C การเปลี่ยนแปลงโครงสร้างในโทรโปไมโอซินเกิดขึ้น วงจรของการเคลื่อนไหวของสะพานขวางเริ่มต้นขึ้น และการหดตัวของซาร์โคเมียร์ - กล้ามเนื้อหดตัว 16. ในสภาวะผ่อนคลาย การทำงานของปั๊ม Ca2+ จะทำงาน ซึ่งจะปั๊มไอออน Ca2+ จากซาร์โคพลาสซึมกลับเข้าไปใน SPR ประเภทของการหดตัวของกล้ามเนื้อโครงร่าง 1. การหดตัวอย่างรวดเร็วและการคลายตัวของกล้ามเนื้อโครงร่างในหลอดทดลองเพื่อตอบสนองต่อการกระตุ้นเรียกว่าการหดตัวของกล้ามเนื้อเดี่ยว (SMC) การรวมตัวของ OMS ที่มีความถี่ของการระคายเคืองเพิ่มขึ้นจะนำไปสู่การหดตัวของบาดทะยัก 2. การหดตัวของเส้นใยกล้ามเนื้อแต่ละเส้นเป็นไปตามกฎทั้งหมดหรือไม่มีเลย 3. ความกว้างของการหดตัวของกล้ามเนื้อทั้งหมดไม่เป็นไปตามกฎของ "ทั้งหมดหรือไม่มีเลย" และอาจเพิ่มขึ้นตามความถี่และความแรงของการกระตุ้นที่เพิ่มขึ้น 4. ความกว้างของการหดตัวยังเพิ่มขึ้นตามจำนวนเส้นใยกล้ามเนื้อที่เกี่ยวข้องกับการหดตัวที่เพิ่มขึ้น ด้วยการหดตัวของเส้นใยกล้ามเนื้อทั้งหมด แอมพลิจูดของการหดตัวจะสูงสุด (เหมาะสมที่สุด) เมื่อเพิ่มความถี่และความแรงของการกระตุ้นมากขึ้น แอมพลิจูดของการหดตัวจะลดลง - ในแง่ลบ 5. ด้วยความถี่ในการกระตุ้นต่ำ การกระตุ้นแต่ละครั้งจะเข้าสู่ระยะผ่อนคลาย OMS ดังนั้นผลรวมของ OMS จะไม่สมบูรณ์ - บาดทะยักหยัก ที่ความถี่การกระตุ้นที่สูงขึ้น การกระตุ้นแต่ละครั้งจะเข้าสู่ระยะหดตัวของ OMS กล้ามเนื้อไม่มีเวลาผ่อนคลาย - บาดทะยักเรียบ 6. การหดตัวเรียกว่าภาพสามมิติหากความตึงเครียดของกล้ามเนื้อเพิ่มขึ้น แต่ไม่เกิดการหดตัว หากกล้ามเนื้อสั้นลงโดยมีความตึงคงที่ การหดตัวดังกล่าวเรียกว่าไอโซโทนิก กล้ามเนื้อเรียบ. กล้ามเนื้อเรียบเป็นเซลล์กล้ามเนื้อนิวเคลียร์เดี่ยวที่มีรูปร่างเป็นแกนหมุน ซึ่งประกอบกันเป็นผนังของอวัยวะภายในและหลอดเลือด และถูกกระตุ้นโดยระบบประสาทอัตโนมัติ 1. แตกต่างจากกล้ามเนื้อโครงร่าง กล้ามเนื้อเรียบไม่มีการเรียงตัวของเส้นใยแอกตินและไมโอซินอย่างสม่ำเสมอ ดังนั้นจึงไม่มีเส้นขวางขวาง และซาร์โคพลาสมิกเรติคูลัมค่อนข้างแสดงออกอย่างอ่อน 2. ไม่มีโทรโปนินในกล้ามเนื้อเรียบ เส้นใยแอกตินติดอยู่กับเนื้อแน่น แทนที่จะเป็น T-tubules บนเยื่อหุ้มเซลล์จะมีการบุกรุกที่เรียกว่า Caveolae 3. เซลล์กล้ามเนื้อเรียบอยู่ติดกันอย่างใกล้ชิดและเชื่อมต่อกันด้วยจุดเชื่อมต่อที่แน่นหนา (nexuses) ซึ่งมีความต้านทานไฟฟ้าต่ำ 4. กล้ามเนื้อเรียบหดตัวช้ากว่ากล้ามเนื้อโครงร่าง ใช้พลังงานน้อยกว่า และสามารถคงสภาพการหดตัวได้นานโดยไม่เมื่อยล้า 5. ในระหว่างการหดตัว ไอออนของ Ca2+ จะถูกปลดปล่อยออกจาก sarcoplasmic reticulum ผ่านช่องทางของตัวรับอิโนซิทอล-3-ฟอสเฟต จับกับสาร Calmodulin และเปิดใช้งาน myosin light chain kinase (MLCK) ซึ่ง phosphorylates ของ myosin light chain ในเวลาเดียวกันกิจกรรมของ myosin ATPase จะเพิ่มขึ้นซึ่งนำไปสู่การเริ่มต้นของวงจรการพายเรือของสะพานขวาง 6. ในระหว่างการคลายตัว ไอออนของ Ca2+ จะถูกกำจัดออกจากซาร์โคพลาสซึมผ่านพลาสมาเมมเบรน หรือเก็บสะสมไว้ในเรติคูลัมของซาร์โคพลาสมิกอีกครั้ง และสายโซ่เบาของไมโอซินจะถูกลดฟอสโฟรีเลตโดยไมโอซินฟอสฟาเทส 7. ในเซลล์กล้ามเนื้อเรียบส่วนใหญ่ ศักยภาพของเยื่อหุ้มเซลล์ไม่คงที่ ซึ่งนำไปสู่การปรากฏของคลื่นสลับโพลาไรซ์หรือศักยภาพของเครื่องกระตุ้นหัวใจอย่างช้าๆ เมมเบรนดีโพลาไรเซชันขึ้นอยู่กับการเพิ่มขึ้นของการซึมผ่านของไอออน Ca2+ 8. ในกล้ามเนื้อเรียบพร้อมกับกระบวนการกระตุ้นและการหดตัวของระบบเครื่องกลไฟฟ้า (ไอออน Ca2+ ทะลุผ่านเซลล์ผ่านช่อง Ca2+ ที่ขึ้นอยู่กับแรงดันไฟฟ้า) มีการมีเพศสัมพันธ์ทางเภสัชกลศาสตร์ - การปล่อยไอออน Ca2+ จากซาร์โคพลาสมิกเรติคูลัมและการหดตัวที่ตามมาโดยไม่มี การเปลี่ยนแปลงอย่างมีนัยสำคัญในศักยภาพของเมมเบรน 9. กล้ามเนื้อเรียบถูกควบคุมโดยแผนกซิมพาเทติกและพาราซิมพาเทติกของระบบประสาทอัตโนมัติ อิทธิพลของอารมณ์ขันก็มีบทบาทสำคัญเช่นกัน - ฮอร์โมน, ปัจจัยด้านกฎระเบียบในท้องถิ่น 10. เซลล์ประสาทที่ทำให้กล้ามเนื้อเรียบสามารถสัมผัสกับซินแนปติกได้หลายครั้ง - เส้นเลือดขอดซึ่งมีคุณสมบัติทั้งหมดของเยื่อหุ้มเซลล์พรีซินแนปติกของปลายประสาท สารสื่อประสาท (acetylcholine หรือ norepinephrine) จะถูกปล่อยออกมาในบริเวณเส้นเลือดขอดทั่วแอกซอน 11. ตัวรับสำหรับสารสื่อประสาทบนเยื่อโพสต์ซินแนปติกจะอยู่ทั้งในบริเวณเส้นเลือดขอดและอยู่ไกลออกไป นอกจากสารสื่อประสาทแล้ว สารสื่อกลางร่วม (ATP, สาร P ฯลฯ) ยังสามารถถูกปล่อยออกมาจากเส้นเลือดขอด ซึ่งจะปรับการตอบสนองของเซลล์กล้ามเนื้อเรียบต่อการทำงานของสื่อกลาง การจำแนกประเภทของกล้ามเนื้อเรียบ 12. กล้ามเนื้อเรียบเดี่ยว (อวัยวะภายใน) มีการสัมผัสระหว่างเซลล์อย่างใกล้ชิด - เน็กซัสที่ให้ปฏิสัมพันธ์ทางไฟฟ้าระหว่างเซลล์ข้างเคียง 13. เซลล์ประเภทนี้บางเซลล์มีคุณสมบัติเป็นอัตโนมัติหรือเครื่องกระตุ้นหัวใจ (ความสามารถในการสร้างศักยภาพในการดำเนินการโดยอิสระ) ดังนั้นเมื่อเซลล์เหล่านี้รู้สึกตื่นเต้น เซลล์กล้ามเนื้อเรียบจำนวนมากสามารถหดตัวพร้อมกันได้ 14. โดยปกติแล้วที่ตำแหน่งของเซลล์เครื่องกระตุ้นหัวใจจะมีเส้นเลือดขอดของเซลล์ประสาทอัตโนมัติ (ในอัตราส่วน 1 เส้นใยประสาท / เส้นใยกล้ามเนื้อ 10-50 เส้น) และเซลล์กล้ามเนื้อเรียบเดียวกันสามารถรับการกระตุ้นพร้อมกันได้ทั้งจากซิมพาเทติกและ เซลล์ประสาทพาราซิมพาเทติก ออกฤทธิ์เป็นปฏิปักษ์ 15. กลุ่มของกล้ามเนื้อเดี่ยวประกอบด้วยกล้ามเนื้อเรียบของผนังหลอดเลือดขนาดเล็ก ระบบทางเดินอาหาร และระบบทางเดินปัสสาวะและอวัยวะสืบพันธุ์ 16. กล้ามเนื้อเรียบหลายหน่วยมีการปกคลุมด้วยเส้นที่ค่อนข้างหนาแน่น (ในอัตราส่วน 1 เส้นใยประสาทต่อ 1 เส้นใยกล้ามเนื้อ) และกระบวนการหดตัวอยู่ภายใต้การควบคุมโดยตรงของระบบประสาทอัตโนมัติ เซลล์กล้ามเนื้อเรียบประเภทนี้แต่ละเซลล์สามารถถูกกระตุ้นและหดตัวโดยไม่ขึ้นกับเซลล์กล้ามเนื้อเรียบที่อยู่รอบๆ เนื่องจาก ปฏิสัมพันธ์ทางไฟฟ้าระหว่างเซลล์มีการพัฒนาไม่ดี 17. ประเภทหลายหน่วยรวมถึงกล้ามเนื้อเรียบของหลอดลมและหลอดเลือดขนาดใหญ่, กล้ามเนื้อของม่านตาและกล้ามเนื้อปรับเลนส์ของตา,
ปฏิกิริยาทางชีวภาพ. สิ่งมีชีวิตและเซลล์ทั้งหมดของพวกมันมีความระคายเคือง เช่น ความสามารถในการตอบสนองต่ออิทธิพลของสิ่งแวดล้อมหรือการรบกวนในสถานะของพวกเขาโดยการเปลี่ยนแปลงโครงสร้างหรือหน้าที่ของมัน ซึ่งเชื่อมโยงความสัมพันธุ์ไม่ได้กับการเปลี่ยนแปลงเชิงปริมาณและคุณภาพในเมแทบอลิซึมและพลังงาน การเปลี่ยนแปลงในโครงสร้างและหน้าที่ของร่างกายและเซลล์ของร่างกายเพื่อตอบสนองต่ออิทธิพลต่างๆ เรียกว่า ปฏิกิริยาทางชีวภาพและอิทธิพลที่ทำให้พวกเขา - สิ่งเร้าหรือสิ่งกระตุ้น.
แนวคิดของการตอบสนองทางชีวภาพรวมถึงกิจกรรมการตอบสนองทุกประเภทของร่างกาย เซลล์และอวัยวะของร่างกายต่ออิทธิพลต่างๆ ปฏิกิริยาของเซลล์แสดงให้เห็นในการเปลี่ยนแปลงรูปร่าง โครงสร้าง การเจริญเติบโตและกระบวนการแบ่งตัว การก่อตัวของสารประกอบทางเคมีต่างๆ ในเซลล์ การแปลงพลังงานศักย์เป็นพลังงานจลน์ (ไฟฟ้า เครื่องกล ความร้อน แสง) ประสิทธิภาพ ของงานอย่างใดอย่างหนึ่ง (การเคลื่อนที่ในอวกาศ การปลดปล่อยสารบางอย่าง การทำงานของการรวมอิเล็กโทรไลต์บางชนิดในเซลล์ เป็นต้น) ปฏิกิริยาของสิ่งมีชีวิตทั้งหมดมีความหลากหลายมากยิ่งขึ้นโดยเฉพาะรูปแบบพฤติกรรมที่ซับซ้อน ในกระบวนการของการดำเนินการกิจกรรมของอวัยวะต่าง ๆ และเซลล์นับไม่ถ้วนจะเปลี่ยนไปเนื่องจากร่างกายมักตอบสนองต่ออิทธิพลต่าง ๆ โดยรวมเป็นระบบเดียวที่ซับซ้อน
สารระคายเคือง. สิ่งระคายเคืองต่อเซลล์ที่มีชีวิตหรือสิ่งมีชีวิตโดยรวมสามารถเป็นการเปลี่ยนแปลงใดๆ ในสภาพแวดล้อมภายนอกหรือสภาวะภายในของสิ่งมีชีวิต ถ้ามีการเปลี่ยนแปลงมากเพียงพอ เกิดขึ้นเร็วพอ และคงอยู่ได้นานพอ
สิ่งเร้าที่เป็นไปได้ที่หลากหลายไม่สิ้นสุดสามารถแบ่งออกเป็น 3 กลุ่ม: ทางกายภาพ, เคมีฟิสิกส์และเคมี สิ่งเร้าทางกายภาพ ได้แก่ อุณหภูมิ กลไก (การกระแทก การทิ่ม ความดัน การเคลื่อนไหว ความเร่ง ฯลฯ) ไฟฟ้า แสง สิ่งเร้าทางกายภาพและเคมีถูกแสดงโดยการเปลี่ยนแปลงของแรงดันออสโมติก, ปฏิกิริยาที่ใช้งานของตัวกลาง, องค์ประกอบของอิเล็กโทรไลต์, สถานะคอลลอยด์ สารระคายเคืองทางเคมีประกอบด้วยสารหลายชนิดที่มีองค์ประกอบและคุณสมบัติต่างกัน และสามารถเปลี่ยนเมแทบอลิซึมของเซลล์ (สารอาหาร ยา สารพิษ ฮอร์โมน เอนไซม์ สารเมแทบอไลต์ ฯลฯ)
สารระคายเคืองต่อเซลล์ที่ก่อให้เกิดกิจกรรมซึ่งมีความสำคัญเป็นพิเศษในกระบวนการชีวิตคือแรงกระตุ้นของเส้นประสาท เป็นธรรมชาติเช่น เกิดขึ้นในร่างกายเอง, สิ่งเร้าทางเคมีไฟฟ้าของเซลล์, แรงกระตุ้นของเส้นประสาท, ทำหน้าที่ไปตามเส้นใยประสาทจากปลายประสาทในระบบประสาทส่วนกลางหรือจากมันไปยังอวัยวะส่วนปลาย, ทำให้เกิดการเปลี่ยนแปลงโดยตรงในสถานะและกิจกรรมของมัน.
สิ่งเร้าทั้งหมดตามสถานที่ที่เกิดขึ้นจะถูกแบ่งออกเป็นสิ่งเร้าภายนอก (ภายนอก-) และภายใน (อินเตอร์-) และตามความสำคัญทางสรีรวิทยา - เพียงพอและไม่เพียงพอ สิ่งเร้าที่เพียงพอคือสิ่งเร้าที่กระทำต่อโครงสร้างทางชีววิทยาที่กำหนดภายใต้สภาพธรรมชาติ ต่อการรับรู้ซึ่งถูกดัดแปลงเป็นพิเศษโดยวิวัฒนาการ และความไวต่อสิ่งเร้าที่มักจะสูงมาก (ตา-แสง หู-เสียง ฯลฯ) สิ่งเร้าเหล่านี้ไม่เพียงพอสำหรับการรับรู้ซึ่งเซลล์หรืออวัยวะนี้ไม่ได้ถูกดัดแปลงเป็นพิเศษ แต่ภายใต้เงื่อนไขบางประการสามารถทำให้เกิดการเปลี่ยนแปลงในโครงสร้างหรือหน้าที่ (กล้ามเนื้อ - สามารถหดตัวเมื่อถูกกระทบ ความร้อนอย่างรวดเร็ว การสัมผัสกับกระแสไฟฟ้า การยืดอย่างกะทันหัน , การกระทำของกรด ฯลฯ .)
ความตื่นเต้นง่ายเซลล์ของเนื้อเยื่อประสาท กล้ามเนื้อ และต่อมได้รับการดัดแปลงเป็นพิเศษเพื่อให้ตอบสนองต่อการระคายเคืองอย่างรวดเร็ว (ตื่นเต้น) เซลล์ของเนื้อเยื่อเหล่านี้เรียกว่าความตื่นเต้นง่ายและความสามารถในการตอบสนองต่อสิ่งเร้าต่าง ๆ ที่มีการกระตุ้นเรียกว่าความตื่นเต้นง่าย ความตื่นเต้นง่าย- นี่คือคุณสมบัติของเยื่อหุ้มเซลล์เพื่อตอบสนองต่อการกระทำของปัจจัยที่ระคายเคือง (น่าตื่นเต้น) โดยการเปลี่ยนการซึมผ่านและสถานะทางไฟฟ้า ปรากฏการณ์นี้เรียกว่าการกระตุ้น การกระตุ้นเป็นปฏิกิริยาทางชีววิทยาที่ซับซ้อนซึ่งแสดงออกมาในการเปลี่ยนแปลงทางกายภาพ เคมีกายภาพ และการทำงานทั้งหมด สัญญาณบังคับของการกระตุ้นคือการเปลี่ยนแปลงสถานะทางไฟฟ้าของเยื่อหุ้มเซลล์ผิว (การเปลี่ยนแปลงของศักย์เยื่อหุ้มเซลล์, MP และการสร้างศักยะงานการแพร่กระจาย, AP) เกิดขึ้นที่เซลล์เดียวหรือส่วนหนึ่ง การกระตุ้นจะแพร่กระจายไปยังส่วนอื่น ๆ ของเซลล์เดียวกันหรือไปยังเซลล์อื่น
การตอบสนองของเซลล์ที่มีชีวิตต่อสิ่งเร้า ไม่ว่าจะในรูปของการกระตุ้นและปฏิกิริยาทางไฟฟ้าที่เกี่ยวข้อง หรือในรูปแบบของการหดตัวหรือการหลั่ง มักจะเกิดขึ้นหลังจากช่วงแฝงหรือช่วงแฝง นี่คือชื่อของช่วงเวลาระหว่างการโจมตีของการกระตุ้นและปฏิกิริยาของเนื้อเยื่อต่อการกระทำของมัน ในช่วงระยะเวลาแฝงต้องผ่านการเปลี่ยนแปลงสถานะของเนื้อเยื่อซึ่งจำเป็นสำหรับปฏิกิริยาที่จะแสดงออกมา ระยะเวลาแฝงของเนื้อเยื่อที่กระตุ้นได้นั้นสั้นกว่าของเนื้อเยื่อที่กระตุ้นไม่ได้ และระยะเวลาแฝงของปฏิกิริยาไฟฟ้าของเนื้อเยื่อนั้นสั้นกว่าการหดตัวของกล้ามเนื้อ และยิ่งกว่านั้นของปฏิกิริยาการหลั่ง
ประวัติการค้นพบปรากฏการณ์ทางไฟฟ้าในเนื้อเยื่อ.
ในปี ค.ศ. 1786 กัลวานี แพทย์และนักสรีรวิทยาชาวอิตาลีซึ่งแขวนขากบไว้ที่ระเบียงเพื่อให้แห้ง สังเกตเห็นว่าเมื่อขาที่แกว่งไปตามลมสัมผัสกับตะแกรงโลหะของระเบียง การหดตัวจะเกิดขึ้น กัลวานีสรุปว่าหากมีการสร้างวงจรระหว่างเส้นประสาทและกล้ามเนื้อโดยใช้ตัวนำโลหะ และในขณะเดียวกันกล้ามเนื้อก็หดตัว นี่เป็นหลักฐานของการปรากฏตัวของ "ไฟฟ้าของสัตว์" เขาเชื่อว่าเส้นประสาทและกล้ามเนื้อมีประจุตรงข้ามกัน
อย่างไรก็ตาม นักฟิสิกส์โวลตาได้แสดงข้อสรุปที่ผิดพลาดของกัลวานีโดยทำการทดลองดังกล่าว เขาสังเกตเห็นว่าราวระเบียงเป็นทองแดง และตะขอที่อุ้งเท้าแขวนเป็นเหล็ก หลังจากพยายามติดแหนบที่เท้า ขาข้างหนึ่งทำด้วยทองแดง ส่วนอีกข้างหนึ่งเป็นสังกะสีหรือเหล็ก โวลตามีอาการกล้ามเนื้อหดตัว ดังนั้นเขาจึงสรุปว่ากล้ามเนื้อหดตัวไม่ใช่เพราะ "ไฟฟ้าของสัตว์" ปล่อยออกมา แต่เป็นเพราะกระแสไฟฟ้าไหลระหว่างโลหะสองชนิดที่สัมผัสกับอิเล็กโทรไลต์ซึ่งทำให้เส้นประสาทที่ขาของกบระคายเคือง
ไม่เห็นด้วยกับ Volta กัลวานีจัดการทดลองครั้งที่สอง ประกอบด้วยการหดตัวของกล้ามเนื้อที่ไม่ใช่โลหะ การหดตัวทำได้โดยการพันเส้นประสาทเหนือกล้ามเนื้อที่เตรียมไว้โดยใช้เครื่องแก้ว อย่างไรก็ตาม ปรากฎว่าการหดตัวจะเกิดขึ้นได้ก็ต่อเมื่อกล้ามเนื้อได้รับความเสียหาย และหากผ่ากล้ามเนื้ออย่างระมัดระวังโดยไม่ทำลายพื้นผิว ก็จะไม่มีการหดตัวเกิดขึ้นในระหว่างการทดลองดังกล่าว ต่อมา เฮอร์แมน นักสรีรวิทยาชาวเยอรมันได้แสดงให้เห็นว่าหากนำอิเล็กโทรดของกัลวาโนมิเตอร์ไปใช้กับกล้ามเนื้อที่ไม่บุบสลาย ก็จะไม่เห็นความแตกต่างที่อาจเกิดขึ้นได้ แต่ถ้ากล้ามเนื้อหรือเส้นประสาทได้รับความเสียหาย ถูกตัด และหนึ่งในขั้วไฟฟ้าถูกแช่อยู่ในบาดแผลนี้ เข็มแกลวาโนมิเตอร์จะเบี่ยงเบน ซึ่งแสดงว่ามีกระแสไฟฟ้าเกิดขึ้นระหว่างส่วนที่เสียหายและไม่เสียหายของกล้ามเนื้อที่มีชีวิตกับส่วนที่เสียหาย มีประจุเป็นลบ เรียกกระแสนี้ว่า กระแสไฟฟ้าขัดข้องหรือกระแสไฟฟ้านิ่ง .
ในปี พ.ศ. 2380 มัตเตอุจิแสดงให้เห็นว่ากระแสพักตัวของกล้ามเนื้อโครงร่างลดลงระหว่างการหดตัว Matteuchi ทำการทดลองอีกครั้งหนึ่ง เขาเตรียมประสาทและกล้ามเนื้อ 2 อย่าง และโยนเส้นประสาทเส้นที่ 2 ไปเหนือกล้ามเนื้อเส้นที่ 1 ในขณะเดียวกันก็ระคายเคืองเส้นประสาทของการเตรียมการครั้งที่ 1 ทำให้กล้ามเนื้อหดตัว ปรากฎว่ากล้ามเนื้อที่ 2 ก็เริ่มหดตัวเช่นกัน สิ่งนี้ไม่สามารถอธิบายได้ด้วยอิทธิพลของกระแสพักบนเส้นประสาท เนื่องจากการหดตัวของกล้ามเนื้อที่สองเกิดขึ้นเมื่อกล้ามเนื้อมัดแรกตื่นเต้นเท่านั้น ประสบการณ์นี้จะยิ่งชัดเจนมากขึ้นหากแทนที่จะเป็นกล้ามเนื้อส่วนแรกที่เราใช้หัวใจการทำงานของกบ เมื่อเส้นประสาทถูกโยนด้วยตะขอแก้วของการเตรียมประสาทและกล้ามเนื้อที่หัวใจของกบ กล้ามเนื้อขาจะเริ่มหดตัวตามจังหวะการเต้นของหัวใจ สาเหตุของปรากฏการณ์นี้ถูกค้นพบในภายหลัง
ในปี พ.ศ. 2393 Dubois-Reymond นักวิจัยชาวฝรั่งเศสผู้มีชื่อเสียงได้ค้นพบว่าหลังจากเกิดการระคายเคือง คลื่นไฟฟ้าจะไหลไปตามเส้นประสาท ในปี พ.ศ. 2411 เฮอร์แมนได้แสดงให้เห็นว่าสาเหตุของเรื่องนี้ก็คือ กระแสไฟฟ้าที่เกิดขึ้นระหว่างการกระตุ้นจะไปถึงส่วนที่อยู่ใกล้เคียง กระตุ้นมัน จากนั้นจึงไปถึงส่วนถัดไป และผ่านการสัมผัสดังกล่าว คลื่นกระตุ้นจะวิ่งไปตามเส้นประสาท เหมือนกับไฟตาม สายฟิคฟอร์ด.
ถ้าส่วนของเส้นประสาทถูกทำให้ระคายเคืองด้วยไฟฟ้ากระแสตรงเพียงจังหวะเดียว และกระแสถูกเบี่ยงเบนจากส่วนถัดไปด้วยอิเล็กโทรดสองตัวไปยังกัลวาโนมิเตอร์หรือไปยังหลอดของออสซิลโลสโคปแคโทด ในตอนแรก ในช่วงเวลาที่มีการระคายเคือง จะไม่มีการเบี่ยงเบน มีการบันทึก เนื่องจากภายใต้อิเล็กโทรดดิสชาร์จทั้งสองมีศักยภาพเท่ากัน หลังจากนั้นไม่นานก็แพร่กระจาย การกระตุ้นไปถึงอิเล็กโทรดดิสชาร์จตัวแรก จากนั้นกัลวาโนมิเตอร์จะบันทึกความต่างศักย์ในรูปแบบของการสั่นเชิงลบ - ลูกศรเบี่ยงเบนไปทางซ้าย (ลงบนออสซิลโลสโคป) เมื่อคลื่นกระตุ้น - อยู่ระหว่างขั้วไฟฟ้า ลูกศรจะกลับสู่ตำแหน่งเดิม จากนั้นคลื่นกระตุ้นจะไปถึงขั้วไฟฟ้าที่สอง - ลูกศรจะเบี่ยงเบนไปทางขวา (ลำแสงขึ้น) เมื่อคลื่นกระตุ้นไปไกลกว่านั้น ทั้งลำแสงของออสซิลโลสโคปและเข็มกัลวาโนมิเตอร์จะกลับสู่ตำแหน่งเดิม
จากข้อเท็จจริงเหล่านี้สามารถสรุปได้ดังต่อไปนี้:
1. ขณะพัก ความต่างศักย์จะเกิดขึ้นระหว่างบริเวณเนื้อเยื่อที่ไม่บุบสลายและเนื้อเยื่อที่เสียหายเท่านั้น (กระแสความเสียหายหรือกระแสที่เหลือ)
2. เมื่อการกระตุ้นผ่านเส้นประสาทกระแสการกระทำจะเกิดขึ้น
3. กระแสของการกระทำนี้ไม่ได้อยู่กับที่ แต่กระจายออกไป
4. กระแสการกระทำคือความผันผวนที่อาจเกิดขึ้นในเชิงลบ
การศึกษาที่แม่นยำยิ่งขึ้นเกี่ยวกับกลไกของการเปลี่ยนแปลงทางไฟฟ้าในเนื้อเยื่อขณะพักและระหว่างการกระตุ้นเป็นไปได้ด้วยความก้าวหน้าของเทคโนโลยีการวัดทางไฟฟ้าและไมโครอิเล็กโทรด ตอนนี้เรามาพิจารณาข้อมูลสมัยใหม่เกี่ยวกับกระบวนการทางไฟฟ้าในเนื้อเยื่อ
ศักยภาพในการพักผ่อน. ปรากฎว่าระหว่างพื้นผิวด้านนอกของเซลล์และโปรโตพลาสซึมในสถานะพักมีความต่างศักย์ของคำสั่ง 60-90 mV และพื้นผิวเซลล์จะถูกประจุไฟฟ้าด้วยความเคารพต่อโปรโตพลาสซึม ความต่างศักย์นี้เรียกว่า ศักย์เมมเบรน หรือศักย์พักตัว การวัดที่แม่นยำทำได้โดยใช้ไมโครอิเล็กโทรดภายในเซลล์เท่านั้น
ตามทฤษฎีเมมเบรนไอออนของ Hodgkin-Huxley ศักย์ไฟฟ้าชีวภาพเกิดจากความเข้มข้นที่ไม่เท่ากันของ K +, Na +, Cl- ไอออนภายในและภายนอกเซลล์ และความสามารถในการซึมผ่านที่แตกต่างกันของเยื่อหุ้มพื้นผิวสำหรับพวกมัน
จากข้อมูลของกล้องจุลทรรศน์อิเล็กตรอน การวิเคราะห์ทางเคมี และการศึกษาทางสรีรวิทยาทางไฟฟ้า มีข้อสันนิษฐานว่าเมมเบรนประกอบด้วยโมเลกุลฟอสโฟลิพิดสองชั้น ซึ่งปกคลุมจากด้านในด้วยชั้นของโมเลกุลโปรตีน และจากด้านนอกด้วยชั้นของโมเลกุลโปรตีน และมิวโคโพลีแซคคาไรด์ สันนิษฐานว่าในเยื่อหุ้มเซลล์มีช่องที่บางที่สุด (รูพรุน) ที่มีเส้นผ่านศูนย์กลางหลายอังสตรอม ผ่านช่องทางเหล่านี้ โมเลกุลของน้ำและสารอื่น ๆ รวมถึงไอออนที่มีเส้นผ่านศูนย์กลางที่สอดคล้องกับขนาดของรูขุมขนจะเข้าสู่เซลล์และปล่อยทิ้งไว้ กลุ่มที่มีประจุต่าง ๆ ได้รับการแก้ไขในองค์ประกอบโครงสร้างของเมมเบรนซึ่งทำให้ผนังของช่องมีประจุหนึ่งหรืออีกอันหนึ่ง ดังนั้น การมีหมู่ฟอสเฟตและคาร์บอกซิลที่แยกตัวออกจากกันในเยื่อหุ้มเส้นใยประสาทจึงเป็นเหตุผลว่าทำไม (เยื่อหุ้ม) จึงสามารถซึมผ่านไปยังแอนไอออนได้น้อยกว่าไอออนบวก
ความสามารถในการซึมผ่านของเมมเบรนสำหรับไอออนบวกที่แตกต่างกันนั้นไม่เหมือนกันและเปลี่ยนแปลงตามธรรมชาติภายใต้สถานะการทำงานที่แตกต่างกันของเนื้อเยื่อ ในขณะพัก เยื่อของเส้นใยประสาทจะซึมผ่าน K ไอออนได้ประมาณ 25 เท่ามากกว่า Na ไอออน และในระหว่างการกระตุ้น โซเดียมจะซึมผ่านได้สูงกว่าโพแทสเซียมประมาณ 20 เท่า
นอกจากการซึมผ่านแล้ว การไล่ระดับความเข้มข้นของไอออนทั้งสองด้านของเมมเบรนยังมีความสำคัญอย่างยิ่งต่อการเกิดศักย์ของเมมเบรน มีการแสดงให้เห็นว่าไซโตพลาสซึมของเซลล์ประสาทและกล้ามเนื้อมีไอออน K+ มากกว่า 30-59 เท่า (500 เมกกะวัตต์/ลิตร เทียบกับ 10 เมกกะวัตต์/ลิตร) แต่มีไอออนของ Na+ น้อยกว่า 8-10 เท่า (35 เมกกะไบต์/ลิตร เทียบกับ 350 เมกกะวัตต์/ลิตร) ) และมี Cl-ion น้อยกว่าของเหลวนอกเซลล์ถึง 50 เท่า (ดูตาราง) ค่าของศักยภาพการพักตัวของเส้นใยประสาทและเซลล์ถูกกำหนดโดยอัตราส่วนของไอออน K+ ที่มีประจุบวกซึ่งแพร่กระจายต่อหน่วยเวลาจากเซลล์ออกไปตามเกรเดียนต์ของความเข้มข้น และไอออน Na+ ที่มีประจุบวกซึ่งกระจายไปตามเกรเดียนต์ของความเข้มข้นในทิศทางตรงกันข้าม ดังนั้น ในการทดลองแบบจำลองบนแอกซอนปลาหมึก ด้วยความเข้มข้นของ K+ ที่เกิดขึ้นในใยประสาท ค่าของกระแส K+ คือ -120 mV หากจำลองเฉพาะการไล่ระดับสีโซเดียมในการทดลองดังกล่าว กระแส Na+ จะเท่ากับ +30 mV ศักยภาพของเมมเบรนที่วัดได้จริงของเส้นประสาทนั้นเท่ากับผลรวมของกระแสที่พุ่งตรงตรงข้ามทั้งสองนี้ นั่นคือ -90mV
แม้ว่าอัตราการแพร่กระจายของ Na+ และ K+ ไอออนผ่านเมมเบรนที่เหลือจะต่ำ แต่ความแตกต่างระหว่างความเข้มข้นของไอออนภายนอกเซลล์และภายในเซลล์จะต้องปรับให้เท่ากันในที่สุดหากไม่มีกลไกพิเศษในเซลล์ที่ช่วยให้มั่นใจได้ ปล่อย ("สูบน้ำออก") จากโปรโตพลาสซึมของ Na + ไอออนที่เจาะเข้าไปและการแนะนำ ("การฉีด") ของ K + ไอออน กลไกนี้เรียกโดยนัยว่าปั๊มโซเดียม-โพแทสเซียม
เพื่อรักษาความไม่สมดุลของไอออน ปั๊ม Na-K จะต้องทำงานบางอย่างเพื่อต่อต้านการไล่ระดับความเข้มข้นของไอออน แหล่งพลังงานโดยตรงสำหรับการทำงานของปั๊มคือการแยก ATP ซึ่งเกิดขึ้นภายใต้อิทธิพลของ ATPase ที่แปลเป็นภาษาท้องถิ่นในเมมเบรนและเปิดใช้งานโดยไอออน Na + และ K + (เรียกว่า ATPase ที่ขึ้นกับ Na-K) การยับยั้งการทำงานของเอนไซม์นี้ทำให้ปั๊มหยุดชะงัก เป็นผลให้โปรโตพลาสซึมอุดมด้วย Na+ และสูญเสีย K+ ผลที่ตามมาโดยตรงคือการลดลงหรือแม้แต่การหายไปอย่างสมบูรณ์ของ MP (ศักยภาพในการพักตัวหรือศักยภาพของเยื่อหุ้มเซลล์)
การสลับโพลาไรเซชันของเมมเบรนเกิดขึ้นเนื่องจากการไล่ระดับความเข้มข้น K + ออกมา แต่เนื่องจาก CL- ไอออนซึ่งไม่สามารถผ่านเมมเบรนได้จะเก็บไอออนบวกด้วยไฟฟ้าสถิต K + ส่วนเกินจะถูกสร้างขึ้นในขอบเขต และระหว่างพื้นผิวด้านนอกและด้านในของเมมเบรน ซึ่งมีประจุบวกและลบตามลำดับ มีความต่างศักย์ประมาณ -90 mV เมมเบรนที่เหลือจะถูกดีโพลาไรซ์อย่างต่อเนื่อง เนื่องจากผลของการทำงานของปั๊ม Na-K ทำให้การไล่ระดับความเข้มข้นของไอออนที่จำเป็นสำหรับสิ่งนี้ยังคงอยู่
ศักยภาพในการดำเนินการ. หากส่วนของเส้นประสาทหรือเส้นใยกล้ามเนื้อสัมผัสกับสิ่งกระตุ้นที่รุนแรงเพียงพอ (เช่น ไฟฟ้าช็อต) จะเกิดการกระตุ้นในบริเวณนี้ ซึ่งอาการที่สำคัญที่สุดอย่างหนึ่งคือความผันผวนอย่างรวดเร็วของ MP ซึ่งเรียกว่าการกระทำ ศักยภาพ (AP)
ด้วยการบันทึกภายในเซลล์ จะพบว่าพื้นผิวของบริเวณที่ตื่นเต้นในช่วงเวลาสั้นๆ มาก ซึ่งวัดเป็นหน่วยหนึ่งในพันของวินาที จะกลายเป็นประจุไฟฟ้าลบเมื่อเทียบกับพื้นที่พักผ่อนที่อยู่ติดกัน เช่น เมื่อตื่นเต้นที่เรียกว่า. เติมเมมเบรน การวัดที่แม่นยำแสดงให้เห็นว่าแอมพลิจูด AP สูงกว่าค่า MF 30–50 mV เหตุผลของเรื่องนี้คือเมื่อมีการกระตุ้น PP ไม่เพียงหายไปเท่านั้น แต่ยังมีความต่างศักย์ของเครื่องหมายตรงกันข้ามซึ่งเป็นผลมาจากการที่ผิวด้านนอกของเมมเบรนกลายเป็นประจุลบเมื่อเทียบกับด้านใน
ใน PD เป็นเรื่องปกติที่จะแยกความแตกต่างระหว่างจุดสูงสุด (ที่เรียกว่าขัดขวาง) และศักยภาพในการติดตาม จุดสูงสุดของ AP มีเฟสขึ้นและลง ก่อนขึ้นเฟสจะเรียกว่าเด่นชัดมากหรือน้อย ศักยภาพของท้องถิ่นหรือการตอบสนองของท้องถิ่น เนื่องจากโพลาไรเซชันเริ่มต้นของเมมเบรนจะหายไปในช่วงขาขึ้น จึงเรียกว่าระยะดีโพลาไรเซชัน ดังนั้นเฟสจากมากไปน้อยในระหว่างที่โพลาไรเซชันของเมมเบรนกลับสู่ระดับเดิมเรียกว่าเฟสรีโพลาไรเซชัน ระยะเวลาของจุดสูงสุดของ AP ในเส้นประสาทและเส้นใยกล้ามเนื้อโครงร่างจะแตกต่างกันไปภายใน 0.4-5.0 มิลลิวินาที ในกรณีนี้ ระยะรีโพลาไรเซชันจะยาวกว่าเสมอ
นอกจากจุดสูงสุดแล้ว ศักยภาพการติดตามสองแบบยังมีความแตกต่างกันใน PD - การสลับโพลาไรเซชันการติดตาม (การติดตามศักยภาพเชิงลบ) และการติดตามไฮเปอร์โพลาไรเซชัน (การติดตามศักยภาพในเชิงบวก ความกว้างของศักยภาพเหล่านี้ไม่เกินหลายมิลลิโวลต์ และระยะเวลาแตกต่างกันไปตั้งแต่หลายสิบถึงหลายร้อยของ มิลลิวินาที ศักยภาพในการติดตามนั้นเกี่ยวข้องกับกระบวนการกู้คืนการพัฒนาในกล้ามเนื้อและเส้นประสาทหลังจากสิ้นสุดการกระตุ้น
สาเหตุของ PD คือการเปลี่ยนแปลงการซึมผ่านของไอออนของเมมเบรน ส่วนที่เหลือดังที่ได้กล่าวไปแล้วการซึมผ่านของเมมเบรนสำหรับ K + นั้นเกินความสามารถในการซึมผ่านของโซเดียม เป็นผลให้การไหลของไอออนที่มีประจุบวกออกจากโปรโตพลาสซึมมีมากกว่าการไหลของ Na+ ที่ตรงกันข้าม ดังนั้นเมมเบรนที่เหลือจึงมีประจุบวกอยู่ด้านนอก
เมื่อเซลล์สัมผัสกับสารระคายเคืองการซึมผ่านของ Na + ไอออนของเมมเบรนจะเพิ่มขึ้นอย่างรวดเร็วและในที่สุดก็จะมากกว่าการซึมผ่านของ K + ประมาณ 20 เท่า ดังนั้นการไหลของไอออน Na + เข้าสู่เซลล์จึงเริ่มเกินอย่างมีนัยสำคัญ การไหลออกของ K+ กระแส Na+ ถึง +150 mV ในเวลาเดียวกัน เอาต์พุตของ K + จากเซลล์จะลดลงบ้าง ทั้งหมดนี้นำไปสู่การบิดเบือน (การพลิกกลับ) ของ MF และพื้นผิวด้านนอกของเมมเบรนจะกลายเป็นประจุไฟฟ้าลบเมื่อเทียบกับพื้นผิวด้านใน การเปลี่ยนแปลงนี้ถูกบันทึกเป็นสาขาจากน้อยไปมากของจุดสูงสุดของ AP (ระยะการสลับขั้ว)
การเพิ่มขึ้นของการซึมผ่านของเมมเบรนสำหรับไอออนของ Na+ ยังคงดำเนินต่อไปในเซลล์ประสาทในช่วงเวลาสั้นๆ มันเชื่อมต่อกับการเปิดระยะสั้นของสิ่งที่เรียกว่า Na+-channels (ให้แม่นยำยิ่งขึ้น ชัตเตอร์ M ในช่องเหล่านี้) ซึ่งจากนั้นจะถูกแทนที่ด้วยการปิดรูขุมขน Na+ อย่างเร่งด่วนด้วยความช่วยเหลือของสิ่งที่เรียกว่า H-ประตู กระบวนการนี้เรียกว่าการยับยั้งโซเดียม เป็นผลให้การไหลของ Na เข้าสู่เซลล์หยุดลง
การมี Na- และ K-channel พิเศษและกลไกที่ซับซ้อนสำหรับการล็อคและเปิดประตูได้รับการศึกษาโดยนักชีวฟิสิกส์ค่อนข้างดี แสดงให้เห็นว่ามีกลไกการเลือกที่ควบคุมบางช่อง ตัวอย่างเช่น ยาพิษ tetrodotoxin จะบล็อกเฉพาะ Na-pore และ tetraethylammonium จะบล็อก K-pores เท่านั้น แสดงให้เห็นว่าในบางเซลล์การกระตุ้นเกิดขึ้นเกี่ยวข้องกับการเปลี่ยนแปลงของการซึมผ่านของเยื่อหุ้มเซลล์สำหรับ Ca ++ ในเซลล์อื่น - สำหรับ Mg + การวิจัยเกี่ยวกับกลไกการเปลี่ยนแปลงการซึมผ่านของเมมเบรนยังคงดำเนินต่อไป
อันเป็นผลมาจากการยับยั้ง Na และการเพิ่มขึ้นของ K-permeability พร้อมกัน มีการปลดปล่อย K+ ไอออนบวกจากโปรโตพลาสซึมไปยังสารละลายภายนอกที่เพิ่มขึ้น อันเป็นผลมาจากกระบวนการทั้งสองนี้ สถานะโพลาไรซ์ของเมมเบรนจะกลับคืนมา (การรีโพลาไรเซชัน) และพื้นผิวด้านนอกของมันจะได้รับประจุบวกอีกครั้ง ในอนาคต กระบวนการคืนค่าองค์ประกอบไอออนปกติของเซลล์และการไล่ระดับความเข้มข้นของไอออนที่จำเป็นเกิดขึ้นเนื่องจากการเปิดใช้งานปั๊ม Na-K
ดังนั้น ในเซลล์ที่มีชีวิต จึงมีการเคลื่อนที่ของไอออนผ่านเยื่อหุ้มเซลล์ที่แตกต่างกันสองแบบ หนึ่งในนั้นดำเนินการตามการไล่ระดับความเข้มข้นของไอออนและไม่ต้องการพลังงาน ดังนั้นจึงเรียกว่าการขนส่งแบบพาสซีฟ มีหน้าที่รับผิดชอบในการเกิดขึ้นของ MP และ PD และท้ายที่สุดจะนำไปสู่การทำให้ความเข้มข้นของไอออนเท่ากันทั้งสองด้านของเยื่อหุ้มเซลล์ ประเภทที่สองของการเคลื่อนที่ของไอออนผ่านเยื่อหุ้มเซลล์ ซึ่งกระทำโดยเทียบกับความเข้มข้นของเกรเดียนต์ ประกอบด้วยการ "สูบ" ไอออน Na+ ออกจากโปรโตพลาสซึม และ "บังคับ" ไอออน K+ เข้าไปในเซลล์ การขนส่งไอออนประเภทนี้เป็นไปได้ก็ต่อเมื่อมีการใช้พลังงาน - นี่คือการขนส่งแบบแอคทีฟ เป็นผลมาจากการทำงานของระบบเอนไซม์พิเศษ (ที่เรียกว่าปั๊ม) และด้วยเหตุนี้ ความแตกต่างเริ่มต้นของความเข้มข้นที่จำเป็นต่อการรักษา MP จึงได้รับการฟื้นฟู
เงื่อนไขสำหรับการตื่นตัว. สำหรับการเกิดขึ้นของ AP มีความจำเป็นที่ภายใต้อิทธิพลของสิ่งเร้าบางอย่าง การเพิ่มขึ้นของการซึมผ่านของไอออนของเยื่อหุ้มเซลล์ที่กระตุ้นได้เกิดขึ้น อย่างไรก็ตาม การกระตุ้นเป็นไปได้ก็ต่อเมื่อสารที่กระทำต่อเมมเบรนมีค่าต่ำสุด (เกณฑ์) ที่แน่นอน ซึ่งสามารถเปลี่ยนศักยภาพของเมมเบรน (MP หรือ Eo) ไปสู่ระดับวิกฤต (Ek ระดับวิกฤตของการดีโพลาไรเซชัน) สิ่งเร้าซึ่งมีความแข็งแรงต่ำกว่าค่าเกณฑ์เรียกว่าเกณฑ์ย่อยซึ่งสูงกว่า - ค่าเหนือเกณฑ์ แสดงให้เห็นว่าแรงเกณฑ์ที่จำเป็นสำหรับการกระตุ้นด้วยไมโครอิเล็กโทรดภายในเซลล์คือ 10 -7 - 10 -9 A
ดังนั้น, เงื่อนไขหลักสำหรับการเกิด PD มีดังต่อไปนี้: ศักยภาพของเมมเบรนจะต้องเท่ากับหรือน้อยกว่าระดับวิกฤตของการดีโพลาไรเซชัน (Eo<= Eк)
เหตุผลของปรากฏการณ์นี้จะชัดเจนสำหรับเราในภายหลังหลังจากมีการชี้แจงกลไกบางอย่างของการกระทำของกระแสไฟฟ้าคงที่บนเนื้อเยื่อที่กระตุ้นได้
ในสภาวะของห้องปฏิบัติการและระหว่างการศึกษาทางคลินิกบางอย่าง เครื่องกระตุ้นไฟฟ้าถูกใช้เพื่อทำให้เส้นประสาทและกล้ามเนื้อระคายเคือง ซึ่งจ่ายได้ง่ายทั้งในแอมพลิจูดและระยะเวลา และในรูป โดยเลียนแบบแรงกระตุ้นของเส้นประสาทตามธรรมชาติ กลไกของผลกระทบที่ระคายเคืองของกระแสต่อเนื้อเยื่อโดยหลักการแล้วจะเหมือนกันสำหรับสิ่งเร้าทุกประเภทโดยใกล้เคียงกับกลไกการออกฤทธิ์ของแรงกระตุ้นของเส้นประสาทมากที่สุด อย่างไรก็ตาม กลไกเหล่านี้จะถูกเปิดเผยในรูปแบบที่แตกต่างกันมากที่สุด เมื่อใช้ไฟฟ้ากระแสตรง
ความหงุดหงิด
ความหงุดหงิดคือความสามารถของระบบชีวิตภายใต้อิทธิพลของสิ่งเร้าที่จะย้ายจากสถานะพักผ่อนทางสรีรวิทยาไปสู่สถานะของกิจกรรม รูปแบบของการแสดงออกของความหงุดหงิดนั้นมีความหลากหลายมาก ในบางเซลล์ รูปร่าง โครงสร้าง การเจริญเติบโตและกระบวนการแบ่งเซลล์จะเปลี่ยนไป ในที่อื่น ๆ มีการสร้างสารประกอบทางเคมีต่าง ๆ งานนี้หรือการเคลื่อนไหวจะดำเนินการ
สารระคายเคืองต่อเซลล์ที่มีชีวิตหรือสิ่งมีชีวิตโดยรวมสามารถทำให้เกิดการเปลี่ยนแปลงใดๆ ในสภาพแวดล้อมภายนอกหรือสภาวะภายในของสิ่งมีชีวิต หากถึงค่าที่กำหนด
ในบรรดาสิ่งเร้าคือ ทางกายภาพ(อุณหภูมิ ทิ่มแทง ความดัน แสง เสียง สิ่งเร้าทางไฟฟ้า) ทางกายภาพและทางเคมี (การเปลี่ยนแปลงของแรงดันออสโมติก, ปฏิกิริยาแอคทีฟของตัวกลาง, องค์ประกอบของอิเล็กโทรไลต์, สถานะคอลลอยด์) และ เคมี(อาหารเคมี, สารประกอบทางเคมีที่เกิดขึ้นในร่างกาย - ฮอร์โมน, ผลิตภัณฑ์เมตาบอลิซึม ฯลฯ)
ข้าว. 17.การทดลองครั้งแรก (I) และครั้งที่สอง (II)
สิ่งกระตุ้นตามธรรมชาติของเซลล์ที่ทำให้เกิดกิจกรรมคือแรงกระตุ้นของเส้นประสาท
ความตื่นเต้นง่าย
เซลล์ของเนื้อเยื่อประสาทและกล้ามเนื้อได้รับการปรับให้เข้ากับปฏิกิริยาตอบสนองต่อการระคายเคืองอย่างรวดเร็ว เซลล์ของเนื้อเยื่อเหล่านี้เรียกว่า excitable และเรียกความสามารถในการตอบสนองต่อการระคายเคืองด้วยการกระตุ้น ความตื่นเต้นง่าย
การกระตุ้นสามารถย้ายจากที่หนึ่งไปยังอีกเซลล์หนึ่งจากเซลล์หนึ่งไปยังอีกเซลล์หนึ่งได้
 ข้าว. 18.ขั้นตอนตามลำดับในการเตรียมประสาทและกล้ามเนื้อ
ข้าว. 18.ขั้นตอนตามลำดับในการเตรียมประสาทและกล้ามเนื้อ
การกระตุ้นนั้นมีลักษณะที่ซับซ้อนของปรากฏการณ์ทางเคมี, การทำงาน, เคมีฟิสิกส์, ปรากฏการณ์ทางไฟฟ้า สัญญาณบังคับของการกระตุ้นคือการเปลี่ยนแปลงสถานะทางไฟฟ้าของเยื่อหุ้มเซลล์ผิว เป็นปรากฏการณ์ทางไฟฟ้าที่ช่วยให้เกิดการกระตุ้นในเนื้อเยื่อที่กระตุ้นได้
ปรากฏการณ์ไบโออิเล็กทริก
การเกิดขึ้นและการแพร่กระจายของการกระตุ้นนั้นสัมพันธ์กับการเปลี่ยนแปลงของประจุไฟฟ้าของเนื้อเยื่อที่มีชีวิต ซึ่งเรียกว่าปรากฏการณ์ไฟฟ้าชีวภาพ
ปรากฏการณ์ทางไฟฟ้าในสัตว์เป็นที่ทราบกันมานานแล้ว นอกจากนี้ในพ.ศ. 2319 อธิบายไว้ในภาพรังสีไฟฟ้า การทดลองของแพทย์ชาวอิตาลี Luigi (1791) ควรถือเป็นจุดเริ่มต้นของการศึกษาทดลองเกี่ยวกับปรากฏการณ์ทางไฟฟ้าในเนื้อเยื่อของสัตว์ ในการทดลอง เขาใช้ส่วนขาหลังของกบที่เชื่อมต่อกับกระดูกสันหลัง เมื่อแขวนอุปกรณ์เหล่านี้ไว้กับตะขอทองแดงจากราวเหล็กของระเบียง เขาสังเกตเห็นว่าเมื่อแขนขาของกบแกว่งไปตามแรงลม กล้ามเนื้อของกบจะหดตัวเมื่อสัมผัสราวบันไดแต่ละครั้ง จากข้อมูลนี้ ฉันได้ข้อสรุปว่าการกระตุกของขาเกิดจาก "ไฟฟ้าของสัตว์" ซึ่งมีต้นกำเนิดในไขสันหลังของกบและส่งผ่านตัวนำโลหะ (ตะขอและราวระเบียง) ไปยังกล้ามเนื้อของกบ ยา.
 ข้าว. 19.โครงการหดตัวทุติยภูมิ
ข้าว. 19.โครงการหดตัวทุติยภูมิ
ตำแหน่งของกัลวานีเกี่ยวกับ "กระแสไฟฟ้าของสัตว์" นี้ถูกต่อต้านโดยนักฟิสิกส์อเล็กซานเดอร์ ในปี พ.ศ. 2335 เขาได้ทดลองซ้ำอีกครั้งของกัลวานีและยืนยันว่าปรากฏการณ์ที่กัลวานีอธิบายนั้นไม่ถือเป็น "ไฟฟ้าจากสัตว์" ในการทดลองของกัลวานี แหล่งที่มาของกระแสไม่ใช่ไขสันหลังของกบ แต่เป็นวงจรที่สร้างจากโลหะต่างชนิดกัน - ทองแดงและเหล็ก
ถูกต้อง ประสบการณ์ครั้งแรกของกัลวานีไม่ได้พิสูจน์ว่ามี "ไฟฟ้าจากสัตว์" แต่การศึกษาเหล่านี้ดึงดูดความสนใจของนักวิทยาศาสตร์ให้ศึกษาปรากฏการณ์ทางไฟฟ้าในการก่อตัวของสิ่งมีชีวิต
เพื่อตอบสนองต่อคำคัดค้านของโวลตา กัลวานีทำการทดลองครั้งที่สอง ครั้งนี้ไม่มีโลหะเข้ามาเกี่ยวข้อง ในขณะเดียวกันก็สังเกตเห็นการหดตัวของขาของกบด้วย
สัมผัสกับกัลวานี เมื่อต้องการทำเช่นนี้ ให้ตรึงกบไว้และตัดมันตามแนวขวางของกระดูกสันหลังส่วนบนของทรวงอก ใช้ผ้าเช็ดปากจับกระดูกสันหลังส่วนที่เหลือ เอาผิวหนังออกจากขาหลัง แล้วใช้แหนบดึงเศษเนื้อด้านในออก ลำต้นประสาทของช่องท้องศักดิ์สิทธิ์ซึ่งอยู่ทั้งสองด้านของกระดูกสันหลังเป็นมัดจะมองเห็นได้ชัดเจน นำแหนบของกัลวานีแผ่นหนึ่งมาไว้ใต้มัดใยประสาททั้งสองมัด แล้วใช้แหนบอีกแผ่นแตะเส้นประสาทด้านบน ในขณะเดียวกันกล้ามเนื้อขาก็หดตัว (รูปที่ 17, I) แหนบของกัลวานีประกอบด้วยแผ่นสังกะสีและทองแดง อธิบายว่าทำไมกล้ามเนื้อขาจึงหดตัวในการทดลองของกัลวานี
 ข้าว. 20.
ข้าว. 20.
ก- ช่องทางเดียว ข- สองช่อง; ทางด้านขวา - ปลายอิเล็กโทรดใกล้กับร่างกายเซลล์ประสาท
ตอนนี้เตรียมการเตรียมกบประสาทและกล้ามเนื้อ
ขั้นตอนหลักในการเตรียมการเตรียมประสาทและกล้ามเนื้อแสดงในรูปที่ 18
ทำให้กบเคลื่อนที่ไม่ได้ ใช้สะโพกของเธอด้วยมือซ้าย (ในตำแหน่งนี้กระดูกสันหลังจะโดดเด่น) แล้วตัดกระดูกสันหลัง 1-1.5 ซม. เหนือตำแหน่งที่เกิดกระดูกเชิงกราน (รูปที่ 18, 1) ถอดส่วนที่ห้อยด้านหน้าของลำตัวและด้านในออก ยึดกระดูกสันหลังส่วนที่เหลือให้แน่นด้วยแหนบหรือมือซ้าย. ใช้แหนบอีกอันจับผิวหนังบริเวณกระดูกสันหลังแล้วดึงลงเพื่อบิดออกจากแขนขา (รูปที่ 18 2 ). วางแขนขาบนจานที่สะอาดแล้วเติมสารละลาย Ringer's ล้างมือให้สะอาดหรือเช็ดให้แห้งจากเมือกที่เกาะอยู่บนผิวหนังของกบ จับชิ้นส่วนของกระดูกสันหลังด้วยแหนบหรือมือของคุณแล้วงอลงเพื่อให้แขนขาห้อยเป็นมุมกับกระดูกสันหลังและกระดูกก้นกบจะโดดเด่น (รูปที่ 18.3)
ตัดกระดูกก้นกบออกอย่างระมัดระวัง ในขณะเดียวกัน ให้กรรไกรอยู่ใกล้กระดูกมากที่สุด เพื่อไม่ให้เส้นประสาทที่วิ่งขนานกันทั้งสองข้างเสียหาย หลังจากตัดก้นกบแล้วให้เตรียมใส่จานแล้วแบ่งออกเป็นสองซีก ในการทำเช่นนี้ ขั้นแรกให้ตัดส่วนที่เหลือของกระดูกสันหลังตามยาว แล้วจึงค่อยประกบที่หัวหน่าว (รูปที่ 18, 4)
 ข้าว. 21.กใช้ไมโครอิเล็กโทรด:
ข้าว. 21.กใช้ไมโครอิเล็กโทรด:
ม- ไมโครอิเล็กโทรด และ- อิเล็กโทรดที่ไม่แยแส ลำแสงบนหน้าจอออสซิลโลสโคปขแสดงให้เห็นว่าก่อนการเจาะของไมโครอิเล็กโทรบ้าน ความต่างศักย์ของเมมเบรนระหว่างดู่ มและ และมีค่าเท่ากับศูนย์ ในขณะที่เจาะ (แสดงด้วยลูกศร) จะตรวจพบความต่างศักย์ ซึ่งบ่งชี้ว่าด้านในของเมมเบรนมีประจุไฟฟ้าเป็นลบเมื่อเทียบกับพื้นผิวด้านนอก
เก็บแขนขาข้างหนึ่งไว้เป็นอะไหล่โดยเก็บไว้ในสารละลายของ Ringer วางอีกอันไว้ที่ด้านหลังแล้วแยกกระดูกเชิงกรานด้วยกรรไกร ใช้แหนบจับชิ้นส่วนของกระดูกสันหลัง เคลื่อนเส้นประสาทไซอาติกออกไปด้านข้าง แล้วเอากระดูกเชิงกรานออก ใช้แหนบ 2 อัน กระจายกล้ามเนื้อบนพื้นผิวด้านหลังของต้นขาตามแนวกึ่งกลาง (รูปที่ 18, 5) หลีกเลี่ยงการสัมผัสเส้นประสาทด้วยกรรไกรและแหนบแยกออกจากเนื้อเยื่อรอบ ๆ ตามต้นขาถึงหัวเข่า (ควรใช้ตะขอแก้วจะดีกว่า) นำเส้นประสาทไปด้านข้างและปล่อยโคนขาออกจากกล้ามเนื้อ(รูปที่ 18, ข). แยกน่องออกจากกระดูกที่ขาท่อนล่าง กล้ามเนื้อ,ตัดเอ็นร้อยหวายแล้วมัดด้าย ตัดขาส่วนล่างและเท้าใต้เข่าออก (รูปที่ 18, 7) . ใส่ยาลงในแก้วที่มีสารละลายของ Ringer
ทำการทดสอบกัลวานีครั้งที่สอง (การลดลงโดยไม่ใช้โลหะ) ในการทำเช่นนี้ ให้เตรียมประสาทและกล้ามเนื้อไว้บนกระดาน ตัดชิ้นส่วนของกล้ามเนื้อออกและใช้ตะขอแก้วโยนเส้นประสาทของยาไปยังส่วนที่บาดเจ็บของกล้ามเนื้ออย่างรวดเร็วเพื่อให้สัมผัสกับพื้นผิวของกล้ามเนื้อที่เสียหายและไม่เสียหายพร้อมกัน (รูปที่ 17, II) จากนั้นกล้ามเนื้อจะหดตัว อธิบายว่าเหตุใดสิ่งนี้จึงเกิดขึ้น
อย่างไรก็ตาม Galvani กลายเป็นเรื่องถูกต้องในคำกล่าวของเขาเกี่ยวกับการมีอยู่ของ "ไฟฟ้าจากสัตว์" ซึ่งต่อมาได้รับการยืนยันจากการศึกษาของนักวิทยาศาสตร์คนอื่นๆ
ในแง่นี้ การทดลองของ Matteuciu ซึ่งเรียกว่าการหดตัวทุติยภูมิเป็นที่สนใจ
โยนเส้นประสาทของการเตรียมประสาทและกล้ามเนื้ออีกอันหนึ่งไปเหนือกล้ามเนื้อของการเตรียมประสาทและกล้ามเนื้อหนึ่ง (รูปที่ 19) และทำให้เส้นประสาทของการเตรียมครั้งแรกระคายเคืองด้วยกระแสไฟฟ้า คุณสังเกตการหดตัวของกล้ามเนื้อและการแก้ไขครั้งที่สอง สิ่งนี้อธิบายได้จากความจริงที่ว่าเมื่อกล้ามเนื้อของยาตัวแรกตื่นเต้น กระแสการกระทำจะเกิดขึ้นซึ่งทำให้เกิดการกระตุ้นของยาประสาทและกล้ามเนื้อตัวที่สอง
ต่อมานักวิทยาศาสตร์ชาวรัสเซียได้มีส่วนร่วมที่สำคัญมากในการศึกษาปรากฏการณ์ไฟฟ้าชีวภาพ ได้แก่ I. M. Sechenov ผู้ค้นพบปรากฏการณ์ทางไฟฟ้าในสมองด้วยความช่วยเหลือของเครื่องวัดกระแสไฟฟ้า N. E. Vvedensky, A. F. Samoilov และคนอื่น ๆ
ในปัจจุบัน มีอุปกรณ์ที่ล้ำสมัยและมีความไวสูง (หลอดรังสีแคโทดพร้อมเครื่องขยายสัญญาณอิเล็กทรอนิกส์) ที่ทำให้สามารถบันทึกปรากฏการณ์ทางไฟฟ้าในเนื้อเยื่อและอวัยวะได้
ศักยภาพในการพักผ่อนและศักยภาพในการเคลื่อนไหวของกล้ามเนื้อ
ระหว่างผิวนอกของเซลล์กับไซโตพลาสซึมที่อยู่นิ่ง มีความต่างศักย์ประมาณ 60-90 mV และผิวเซลล์จะถูกประจุไฟฟ้าในทางบวกเมื่อเทียบกับไซโตพลาสซึม ความต่างศักย์นี้เรียกว่า ศักยภาพในการพักผ่อน หรือ ศักยภาพของเมมเบรน การวัดศักยภาพการพักอย่างแม่นยำทำได้ด้วยความช่วยเหลือของไมโครอิเล็กโทรดที่ออกแบบมาสำหรับการผันกระแสภายในเซลล์ แอมพลิฟายเออร์ที่ทรงพลังมาก และอุปกรณ์บันทึกที่ละเอียดอ่อน - ออสซิลโลสโคป
 ข้าว. 22.ศักย์ไฟฟ้าของแอกซอนปลาหมึก ถ่ายด้วยอิเล็กโทรดภายในเซลล์
ข้าว. 22.ศักย์ไฟฟ้าของแอกซอนปลาหมึก ถ่ายด้วยอิเล็กโทรดภายในเซลล์
ไมโครอิเล็กโทรด (รูปที่ 20) เป็นเส้นเลือดฝอยแก้วบาง ๆ ซึ่งส่วนปลายมีเส้นผ่านศูนย์กลางประมาณหนึ่งไมครอน เส้นเลือดฝอยนี้เต็มไปด้วยน้ำเกลือ อิเล็กโทรดโลหะแช่อยู่ในนั้นและเชื่อมต่อกับเครื่องขยายเสียงและออสซิลโลสโคป (รูปที่ 21) ทันทีที่ไมโครอิเล็กโทรดเจาะเยื่อหุ้มเซลล์ ลำแสงออสซิลโลสโคปจะเบี่ยงเบนลงด้านล่างจากตำแหน่งเดิมและถูกตั้งค่าเป็นตำแหน่งใหม่ระดับ. สิ่งนี้บ่งชี้ว่ามีความแตกต่างที่อาจเกิดขึ้นระหว่างพื้นผิวด้านนอกและด้านในของเยื่อหุ้มเซลล์
คำอธิบายที่สมบูรณ์ที่สุดของที่มาของศักยภาพที่เหลือคือสิ่งที่เรียกว่า เยื่อ แต่ไอออนิกทฤษฎี. ตามทฤษฎีนี้ เซลล์ทั้งหมดถูกปกคลุมด้วยเมมเบรนที่มีการซึมผ่านของไอออนที่แตกต่างกันไม่เท่ากัน ในแง่นี้ ภายในเซลล์ในไซโตพลาสซึมมีโพแทสเซียมไอออนมากกว่า 30-50 เท่า โซเดียมไอออนน้อยกว่า 8-10 เท่า และคลอไรด์ไอออนน้อยกว่า 50 เท่าบนผิวเซลล์ ขณะพัก เยื่อหุ้มเซลล์จะซึมผ่านโพแทสเซียมไอออนได้ดีกว่าโซเดียมไอออน
การแพร่กระจายของไอออนโพแทสเซียมที่มีประจุบวกจากไซโตพลาสซึมไปยังผิวเซลล์ทำให้มีประจุบวกที่ผิวด้านนอกของเยื่อหุ้มเซลล์
ดังนั้น พื้นผิวของเซลล์ที่อยู่นิ่งจึงมีประจุบวก ในขณะที่ด้านในของเมมเบรนมีประจุลบเนื่องจากคลอไรด์ไอออน กรดอะมิโน และแอนไอออนอินทรีย์ขนาดใหญ่อื่นๆ ซึ่งแทบไม่ทะลุผ่านเยื่อหุ้มเซลล์
หากส่วนของเส้นประสาทหรือเส้นใยกล้ามเนื้ออยู่ภายใต้การกระตุ้นที่แรงเพียงพอ การกระตุ้นจะเกิดขึ้นในบริเวณนี้ ซึ่งแสดงออกมาในความผันผวนอย่างรวดเร็วของศักย์เยื่อหุ้มเซลล์และเรียกว่า ศักยภาพในการดำเนินการ
ศักยภาพในการดำเนินการสามารถบันทึกได้โดยใช้อิเล็กโทรดที่ติดบนผิวด้านนอกของเส้นใย (ตะกั่วนอกเซลล์) หรือไมโครอิเล็กโทรดที่ใส่เข้าไปในไซโตพลาสซึม (ตะกั่วในเซลล์) (รูปที่ 22)
เมื่อนำไมโครอิเล็กโทรดเข้าไปในเส้นใยประสาท ศักยภาพประมาณ 60 มิลลิโวลต์จะถูกบันทึกเป็นอันดับแรก นี่คือศักยภาพของเมมเบรน
ศักยภาพในการดำเนินการคือการเพิ่มขึ้นอย่างฉับพลันในศักยภาพของเมมเบรน ในกรณีนี้ ไม่เพียงแต่การเปลี่ยนแปลงของศักยภาพการพักเชิงลบจากค่า - 60 mV เป็นศูนย์เท่านั้น แต่ยังเปลี่ยนผ่านเส้นศูนย์ไปหลายสิบมิลลิโวลต์ด้วย ดังนั้นแอมพลิจูดของศักย์ไฟฟ้าจึงเกินค่าพักตัว ศักย์ไฟฟ้าและมีค่าประมาณ 90 mV
ในศักยภาพของการกระทำนั้นมีความโดดเด่น จุดสูงสุด(ขัดขวางตามศัพท์ของผู้แต่งภาษาอังกฤษ) และ ติดตามศักยภาพ ในช่วงพีค ศักย์เมมเบรนจะเปลี่ยนสัญญาณ
สาเหตุของศักยภาพในการดำเนินการคือการเปลี่ยนแปลงการซึมผ่านของไอออนของเมมเบรน เมื่อเกิดการระคายเคือง การซึมผ่านของโซเดียมไอออนของเยื่อหุ้มเซลล์จะเพิ่มขึ้น มีแนวโน้มที่จะเข้าสู่เซลล์ เนื่องจาก ประการแรก มันมีประจุบวกและแรงไฟฟ้าสถิตดึงเข้าไปข้างใน และประการที่สอง ความเข้มข้นภายในเซลล์ต่ำ ส่วนที่เหลือ เยื่อหุ้มเซลล์ไม่สามารถซึมผ่านไปยังไอออนนี้ได้ การระคายเคืองเปลี่ยนการซึมผ่านของเมมเบรน และการไหลของไอออนโซเดียมที่มีประจุบวกจากสภาพแวดล้อมภายนอกของเซลล์ไปยังไซโตพลาสซึมมีมากกว่าการไหลของโพแทสเซียมไอออนจากเซลล์สู่ภายนอกอย่างมีนัยสำคัญ เป็นผลให้มีการเบี่ยงเบนของศักยภาพของเมมเบรน (ระยะการสลับขั้ว) พื้นผิวด้านในของเมมเบรนกลายเป็นประจุบวก และพื้นผิวด้านนอกเนื่องจากการสูญเสียโซเดียมไอออนที่มีประจุบวกจึงกลายเป็นประจุลบ ณ จุดนี้ จุดสูงสุดของศักยภาพในการดำเนินการจะถูกบันทึกไว้
การเพิ่มขึ้นของการซึมผ่านของเมมเบรนไปยังโซเดียมไอออนนั้นใช้เวลาสั้นมาก หลังจากนี้กระบวนการกู้คืนเกิดขึ้นในเซลล์ซึ่งนำไปสู่ความจริงที่ว่าการซึมผ่านของเมมเบรนสำหรับโซเดียมไอออนลดลงอีกครั้งและสำหรับโพแทสเซียมไอออนจะเพิ่มขึ้น เนื่องจากโพแทสเซียมไอออนยังมีประจุบวก ออกจากเซลล์ พวกมันจึงคืนความสัมพันธ์เดิมทั้งภายนอกและภายในเซลล์ (ระยะรีโพลาไรเซชัน)
ไม่เกิดการสะสมของโซเดียมภายในเซลล์ด้วยการกระตุ้นซ้ำๆ เนื่องจากโซเดียมจะถูกขับออกมาอย่างต่อเนื่องเนื่องจากกลไกทางชีวเคมีพิเศษที่เรียกว่า "ปั๊มโซเดียม" นอกจากนี้ยังมีข้อมูลเกี่ยวกับการขนส่งโพแทสเซียมที่ใช้งานอยู่โดยใช้ "ปั๊มโซเดียมโพแทสเซียม"
ดังนั้น ตามทฤษฎีเมมเบรน การเลือกซึมผ่านของเยื่อหุ้มเซลล์จึงมีความสำคัญอย่างยิ่งในการกำเนิดปรากฏการณ์ไฟฟ้าชีวภาพ ซึ่งกำหนดองค์ประกอบไอออนิกที่แตกต่างกันบนพื้นผิวและภายในเซลล์ และด้วยเหตุนี้ ประจุไฟฟ้าที่แตกต่างกันของพื้นผิวเหล่านี้ . ควรสังเกตว่าข้อกำหนดหลายข้อของทฤษฎีเมมเบรนยังคงเป็นที่ถกเถียงกันอยู่และต้องการการพัฒนาเชิงลึก
ดังนั้นคุณสมบัติทางไฟฟ้าชีวภาพของเซลล์เนื่องจากการกระจายตัวของไอออนแร่ที่ไม่สม่ำเสมอจึงมีบทบาทสำคัญในกระบวนการกระตุ้นเซลล์ การเกิดขึ้นของศักยภาพในการดำเนินการเป็นตัวบ่งชี้ลักษณะเฉพาะของการกระตุ้นในเซลล์ เนื้อเยื่อ และอวัยวะต่างๆ ดังนั้นการศึกษาทางสรีรวิทยาทางไฟฟ้าจึงถูกนำมาใช้กันอย่างแพร่หลายในห้องปฏิบัติการทดลองและคลินิก
ความสำคัญของการลงทะเบียนปรากฏการณ์ไฟฟ้าชีวภาพ การลงทะเบียนศักย์ไฟฟ้าในสิ่งมีชีวิตทำให้สามารถตัดสินการเปลี่ยนแปลงในการทำงานทางสรีรวิทยาหลายอย่างได้ เนื่องจากกิจกรรมทางไฟฟ้าของอวัยวะต่างๆ ของมนุษย์มักมีลักษณะทั่วไปและคงที่ วิธีการทางสรีรวิทยาทางไฟฟ้าจึงถูกนำมาใช้กันอย่างแพร่หลายในการวินิจฉัยโรคในเวชปฏิบัติ มีความก้าวหน้าอย่างมากในการจำแนกโรคของหัวใจ ระบบประสาท และกล้ามเนื้อ วิธีการทางสรีรวิทยาทางไฟฟ้ามีบทบาทสำคัญในการแก้ปัญหาต่างๆ ของสรีรวิทยาอวกาศ ด้วยความช่วยเหลือของวิธีการ telemetry มันเป็นไปได้ที่จะส่งข้อมูลเกี่ยวกับสถานะของกล้ามเนื้อหัวใจ, กิจกรรมของสมอง, กล้ามเนื้อโครงร่างและอวัยวะอื่น ๆ ในสภาวะไร้น้ำหนัก, เกินพิกัด
การศึกษากิจกรรมทางไฟฟ้าของระบบประสาทส่วนกลางเป็นพื้นฐานสำหรับการออกแบบคอมพิวเตอร์อิเล็กทรอนิกส์ที่ซับซ้อน
Electroencephalography เป็นการบันทึกปรากฏการณ์ไฟฟ้าชีวภาพที่เกิดขึ้นในสมอง ส่วนใหญ่อยู่ในเปลือกสมอง
การออกแบบอิเล็กโทรดต่างๆ ใช้เพื่อเบี่ยงเบนกระแสชีวภาพจากโครงสร้างสมองต่างๆ ในการทดลองกับสัตว์ สามารถเสียบอิเล็กโทรดผ่านกระดูกของกะโหลกศีรษะโดยตรงไปยังบริเวณที่ต้องการของสมอง อิเล็กโทรด "ฝัง" ดังกล่าวถูกยึดไว้ในสมองเป็นเวลานานโดยตัวยึดพิเศษและทำให้สามารถศึกษากิจกรรมทางไฟฟ้าของสมองบางส่วนภายใต้สภาวะต่างๆ ของสัตว์ได้
 ข้าว. 23.
ข้าว. 23.
ในปัจจุบัน การบันทึกปรากฏการณ์ทางไฟฟ้าในระยะยาวสามารถทำได้แม้ในเซลล์สมองแต่ละเซลล์ด้วยความช่วยเหลือของไมโครอิเล็กโทรด
เมื่อบันทึกกระแสชีวภาพในสมองมนุษย์ - อิเล็กโทรเอนฟาโลแกรม - พวกเขามักจะใช้อิเล็กโทรดสีเงินซึ่งดูเหมือนจานขนาดเท่าเหรียญสองโกเปค อิเล็กโทรดบนหัวของวัตถุถูกยึดด้วยหมวกตาข่าย หมวกกันน็อคทำจากแถบยางยืดซึ่งควบคุมความตึง หมวกกันน็อคซึ่งสวมเข้ากับศีรษะของวัตถุอย่างแน่นหนา ยึดอิเล็กโทรดไว้อย่างแน่นหนา (รูปที่ 23)
กระแสชีวภาพของสมองถูกบันทึกไว้ในอุปกรณ์ - อิเล็กโทรกราฟี, มีการออกแบบที่แตกต่างกันและรวมถึงแอมพลิฟายเออร์กระแสชีวภาพ ออสซิลโลสโคป และแผงควบคุมที่ซับซ้อนสำหรับพวกมัน
ปัจจุบันมีการผลิตอุปกรณ์ที่สามารถบันทึกกิจกรรมทางไฟฟ้าได้พร้อมกันตั้งแต่ 2 ถึง 32 จุดของสมองและอีกมากมาย
สำหรับการวิเคราะห์ปรากฏการณ์ทางไฟฟ้าที่เกิดขึ้นในสมองและอวัยวะอื่น ๆ อย่างแม่นยำจะใช้คอมพิวเตอร์อิเล็กทรอนิกส์
กิจกรรมทางไฟฟ้าของสมองมนุษย์เป็นจังหวะ อิเล็กโทรดที่อยู่บนพื้นผิวของศีรษะทำให้สามารถลงทะเบียนกระแสชีวภาพจากเซลล์สมองจำนวนมากที่อยู่ใต้พวกมันได้ในคราวเดียว ดังนั้นลักษณะทั่วไปของเอนฟาโลแกรมจึงซับซ้อนมาก ในขณะเดียวกันก็เป็นไปได้ที่จะพิสูจน์ว่ามีจังหวะความผันผวนของกิจกรรมทางไฟฟ้าที่เด่นชัดและเกิดขึ้นบ่อยครั้งน้อยที่สุด ชื่อของจังหวะเหล่านี้แสดงตามอัตภาพด้วยตัวอักษรกรีก α, β, δ , ϑ (รูปที่ 25 ตารางที่ 4)
ตารางที่ 4
จังหวะไฟฟ้าสมอง
 ข้าว. 25.
ข้าว. 25.
1 - จังหวะอัลฟา 2 - จังหวะเบต้า; 3 - จังหวะเดลต้า; 4 - จังหวะทีต้า; 5 - แกนหมุน; ขึ้น y - การประทับเวลา
ด้วยแรงกระตุ้นสู่ศูนย์กลางที่ จำกัด ไปยังพื้นที่ของสมองภายใต้การศึกษามักจะสังเกตคลื่นช้าที่มีช่วงการสั่นขนาดใหญ่ หากแรงกระตุ้นจำนวนมากเข้าสู่เยื่อหุ้มสมอง เซลล์ในบริเวณนี้อาจอยู่ในขั้นตอนต่างๆ ของการกระตุ้น และกิจกรรมทางไฟฟ้าทั้งหมดในบริเวณนี้มีลักษณะผันผวนบ่อยครั้งโดยมีแอมพลิจูดเล็กน้อยของประเภท จังหวะเบต้ารูปที่ 26 แสดงอิเล็กโทรเอนฟาโลแกรมจากบริเวณท้ายทอยของเยื่อหุ้มสมอง (โซนการมองเห็นของเปลือกสมองอยู่ที่นั่น) มันแสดงให้เห็นการเปลี่ยนแปลงอย่างชัดเจน จังหวะอัลฟ่าเข้าสู่จังหวะเบต้าและย้อนกลับเมื่อเปิดและปิดตา กล่าวคือ มีการเพิ่มและลดลงของการไหลของแรงกระตุ้นจากศูนย์กลางไปยังคอร์เทกซ์การมองเห็น
เทคโนโลยีสมัยใหม่ทำให้สามารถลงทะเบียนเอนฟาโลแกรมในบุคคลในระยะไกลและแม้แต่ในสภาพการบินในอวกาศ
วิธีการศึกษากิจกรรมไฟฟ้าของกล้ามเนื้อก็แพร่หลายเช่นกัน - การตรวจด้วยคลื่นไฟฟ้า
 อิเล็กโทรดโลหะผิวหนังที่มีเส้นผ่านศูนย์กลาง 10 มม. ใช้เพื่อเบี่ยงเบนศักยภาพทางชีวภาพของกล้ามเนื้อมนุษย์ อิเล็กโทรดได้รับการแก้ไขบนกล้ามเนื้อภายใต้การศึกษาด้วยผ้าพันแขนยืดหยุ่น ระหว่างพวกเขากับผิวหนังมักจะวางพิเศษที่ปรับปรุงการสัมผัสกับร่างกายและการนำไฟฟ้า การสั่นของศักยภาพทางชีวภาพของกล้ามเนื้อยังมีลักษณะที่เป็นจังหวะ เฉพาะความถี่และแอมพลิจูดเท่านั้นที่มากกว่าเมื่อบันทึกอิเล็กโทรเอนฟาโลแกรม การเพิ่มขึ้นของกิจกรรมของกล้ามเนื้อนั้นมาพร้อมกับการเพิ่มขึ้นของแอมพลิจูดและความผันผวนของอิเล็กโทรไมโอแกรม ด้วยความเหนื่อยล้าของกล้ามเนื้อความถี่ของการสั่นตามกฎจะตก
อิเล็กโทรดโลหะผิวหนังที่มีเส้นผ่านศูนย์กลาง 10 มม. ใช้เพื่อเบี่ยงเบนศักยภาพทางชีวภาพของกล้ามเนื้อมนุษย์ อิเล็กโทรดได้รับการแก้ไขบนกล้ามเนื้อภายใต้การศึกษาด้วยผ้าพันแขนยืดหยุ่น ระหว่างพวกเขากับผิวหนังมักจะวางพิเศษที่ปรับปรุงการสัมผัสกับร่างกายและการนำไฟฟ้า การสั่นของศักยภาพทางชีวภาพของกล้ามเนื้อยังมีลักษณะที่เป็นจังหวะ เฉพาะความถี่และแอมพลิจูดเท่านั้นที่มากกว่าเมื่อบันทึกอิเล็กโทรเอนฟาโลแกรม การเพิ่มขึ้นของกิจกรรมของกล้ามเนื้อนั้นมาพร้อมกับการเพิ่มขึ้นของแอมพลิจูดและความผันผวนของอิเล็กโทรไมโอแกรม ด้วยความเหนื่อยล้าของกล้ามเนื้อความถี่ของการสั่นตามกฎจะตก
ข้าว.26. การเปลี่ยนแปลงของคลื่นไฟฟ้าสมองในคอร์เทกซ์ท้ายทอยแสดงการเปลี่ยนจากอัลฟ่าเป็นเบต้าเมื่อลืมตา (ลูกศรขึ้น) และการฟื้นตัวของอัลฟ่าเมื่อหลับตา (ลูกศรลง)
รูปที่ 27 แสดงอิเล็กโทรไมโอแกรมของกล้ามเนื้อ triceps และ biceps ของไหล่เมื่อทำงานกับไฟล์ พวกเขาแสดงการสั่นที่เพิ่มขึ้นอย่างชัดเจนเมื่อกล้ามเนื้อตื่นเต้น Electromyography ช่วยให้คุณกำหนดระดับของการมีส่วนร่วมของกล้ามเนื้อบางส่วนในการเคลื่อนไหวที่ทำ วิธีนี้มีความสำคัญอย่างยิ่งสำหรับการตรวจหาภาวะกล้ามเนื้อเป็นอัมพาตในโรคบางชนิด ดังนั้นด้วยความช่วยเหลือของ electromyography ทำให้สามารถตรวจพบอัมพาตของกล้ามเนื้อทางเดินหายใจในโปลิโอก่อนที่จะหยุดลง และนี่เป็นสิ่งสำคัญมากสำหรับการดำเนินมาตรการที่จำเป็น (ถ่ายโอนไปยังการควบคุมโดยใช้อุปกรณ์พิเศษ) เพื่อช่วยชีวิตบุคคล
 ข้าว. 27. Electromyogram triceps (1)
และสองหัว (2)
กล้ามเนื้อไหล่ของคนงานเมื่อยื่น
ข้าว. 27. Electromyogram triceps (1)
และสองหัว (2)
กล้ามเนื้อไหล่ของคนงานเมื่อยื่น
ข้อมูล Electromyography ถูกใช้เพื่อสร้างอวัยวะเทียมที่ใช้งานอยู่และหุ่นยนต์ควบคุม
"มือประดิษฐ์" เป็นหนึ่งในอุปกรณ์ชิ้นแรกที่ควบคุมโดยกระแสชีวภาพของกล้ามเนื้อ นี่คืออวัยวะเทียมแบบแอคทีฟที่สร้างการเคลื่อนไหวของมือตามปกติ อวัยวะเทียมที่ใช้งานอยู่นี้มีสร้อยข้อมือสำหรับเปลี่ยนกระแสชีวภาพของกล้ามเนื้อของตอไม้ที่เก็บรักษาไว้ สร้อยข้อมือมีถ้วยโลหะที่บรรจุสารนำไฟฟ้า กระแสของกล้ามเนื้อที่ถูกดึงออกด้วยความช่วยเหลือของสร้อยข้อมือจะถูกขยายและป้อนเข้าสู่หน่วยพิเศษบนอวัยวะเทียมซึ่งจะมีการสร้างสัญญาณควบคุม สัญญาณจะเปิดใช้งานมอเตอร์ขนาดเล็กที่ขับเคลื่อนอวัยวะเทียม อวัยวะเทียมทำจากวัสดุพลาสติกที่มีรูปร่าง ขนาด และสีใกล้เคียงกับมือปกติ
การส่งแรงกระตุ้นในประสาทสัมผัส
การกระตุ้นจากเซลล์ประสาทหนึ่งไปยังอีกเซลล์หนึ่ง หรือจากใยประสาทไปยังกล้ามเนื้อหรือเซลล์ต่อมถูกส่งด้วยความช่วยเหลือของ ไซแนปส์ นกฮูก
แอกซอนของเซลล์ประสาทส่วนใหญ่ที่เข้าใกล้เซลล์ประสาทอื่นๆ จะแตกแขนงและก่อตัวเป็นปลายจำนวนมากบนร่างกายของเซลล์เหล่านี้และเดนดรีนของพวกมัน (รูปที่ . 28). จุดติดต่อเหล่านี้เรียกว่าไซแนปส์
 ข้าว. 28.ตำแหน่งของไซแนปส์ในร่างกายของเซลล์ประสาทและเดนไดรต์ของมัน (อ้างอิงจาก Haach และ Barr)
ข้าว. 28.ตำแหน่งของไซแนปส์ในร่างกายของเซลล์ประสาทและเดนไดรต์ของมัน (อ้างอิงจาก Haach และ Barr)
จำนวนของไซแนปส์ในร่างกายของเซลล์ประสาทหนึ่งเซลล์ถึง 100 หรือมากกว่านั้น และในเดนไดรต์ของเซลล์ประสาทหนึ่งเซลล์ - หลายพัน เส้นใยประสาทหนึ่งเส้นสามารถสร้างได้ถึง 10,000 ไซแนปส์ในเซลล์ประสาทจำนวนมาก
ไซแนปส์มีโครงสร้างที่ซับซ้อน (รูปที่ 29) มันเกิดจากเยื่อหุ้มสองอัน - พรีไซแนปติก และ โพสต์ ออปติก, ซึ่งระหว่างนั้นมี ช่องว่างสรุป presynaptic membrane อยู่ที่ปลายประสาท ปลายประสาทในระบบประสาทส่วนกลางมีลักษณะเหมือนปุ่ม วงแหวน หรือแผ่นโลหะ บนร่างกายของเซลล์พีระมิดมีปลายประสาทหลายพันเส้น ปุ่มซินแนปติกแต่ละปุ่มถูกหุ้มด้วยเมมเบรนพรีซินแนปติก เมมเบรนโพสซินแนปติกตั้งอยู่บนร่างกายหรือเดนไดรต์ของเซลล์ประสาทที่ส่งกระแสประสาท ในบริเวณ presynaptic มักพบการสะสมของไมโตคอนเดรียจำนวนมาก
การกระตุ้นผ่านประสาทจะถูกส่งทางเคมีด้วยความช่วยเหลือของผู้ไกล่เกลี่ยพิเศษหรือคนกลางอยู่ในแผ่นซินแนปติก ใน synaptic plaque ตัวส่งสัญญาณจะอยู่ใน synaptic vesicles ซึ่งแต่ละตัวมีโมเลกุลตัวส่งสัญญาณมากถึง 2,000 ตัว ไซแนปส์ที่ต่างกันจะสร้างสารสื่อประสาทที่ต่างกัน ส่วนใหญ่ acetylcholine, adrenaline หรือ noradrenaline ทำหน้าที่เป็นสื่อกลางใน synapses บางชนิด - กรดกลูตามิก
กลไกการส่งผ่านของการกระตุ้นในไซแนปส์คืออะไร? การมาถึงของกระแสประสาทที่จุดสิ้นสุดของพรีซินแนปติกจะมาพร้อมกับการปลดปล่อยสารสื่อประสาทแบบซิงโครนัสเข้าสู่รอยแหว่งไซแนปติกจากถุงซินแนปติกที่อยู่ในบริเวณใกล้เคียง ขนาดของช่องว่างนี้มีขนาดเล็กมาก (ประมาณ 200 Å) และสารสื่อประสาทซึ่งเข้าถึงเยื่อหุ้มโพสต์ซินแนปติกอย่างรวดเร็วจะทำปฏิกิริยากับสารของมัน อันเป็นผลมาจากการโต้ตอบนี้ โครงสร้างของเมมเบรนแบบโพสต์ซินแนปติกเปลี่ยนแปลงชั่วคราว ความสามารถในการซึมผ่านของโซเดียมไอออนเพิ่มขึ้น ซึ่งนำไปสู่การเคลื่อนที่ของไอออน และเป็นผลให้ลักษณะที่ปรากฏ ศักยภาพของโพสซินแนปติกกระตุ้น เมื่อศักยภาพนี้ถึงค่าหนึ่ง การกระตุ้นการแพร่กระจายจะเกิดขึ้น - ศักยภาพในการดำเนินการ
 ข้าว. 29.
ข้าว. 29.
1 - เมมเบรนพรีซินแนปติก 2 - เมมเบรนโพสต์ซินแน็ปติก; 3 - ซินแนปติกช่างเป็นช่องว่าง 4 - ถุง synaptic; 5 - นิวโรโปรโตไฟบริล; 6 - ไมโตคอนเดรีย
หลังจากเวลาสั้น ๆ (หลายมิลลิวินาที) ผู้ไกล่เกลี่ยจะถูกทำลายโดยเอนไซม์พิเศษ
ในปัจจุบัน นักประสาทสรีรวิทยาส่วนใหญ่ยอมรับการมีอยู่ของประสาทไขสันหลังและในส่วนต่างๆ ของสมองของซินแนปส์สองประเภทที่มีคุณภาพแตกต่างกัน - กระตุ้นและยับยั้ง เป็นที่เชื่อกันว่าในเซลล์ประสาทยับยั้งพิเศษที่ปลายประสาทของซอนมีการผลิตสื่อกลางพิเศษที่มีผลยับยั้งเซลล์ประสาทที่ตามมา ลักษณะของผู้ไกล่เกลี่ยนี้ยังไม่เป็นที่แน่ชัด ในเปลือกสมองผู้เขียนบางคนคิดว่ากรดแกมมาอะมิโนบิวทีริกเป็นสื่อกลาง
ภายใต้อิทธิพลของแรงกระตุ้นที่มาตามแอกซอนของเซลล์ประสาทแบบยับยั้ง ตัวไกล่เกลี่ยจะถูกปล่อยเข้าไปในรอยแหว่งไซแนปติก ซึ่งทำให้เกิดการเปลี่ยนแปลงเฉพาะในเยื่อหุ้มโพสซินแนปติก สาระสำคัญของการเปลี่ยนแปลงเหล่านี้คือตัวกลางที่ยับยั้งซึ่งมีปฏิสัมพันธ์กับสารของเมมเบรนแบบโพสต์ซินแนปติกเพิ่มการซึมผ่านของไอออนโพแทสเซียมและคลอไรด์ จำนวนแอนไอออนเพิ่มขึ้นภายในเซลล์ เป็นผลให้ไม่มีการลดลงของประจุภายในของเมมเบรน (เช่นเดียวกับที่เกิดขึ้นในไซแนปส์ excitatory) แต่การเพิ่มขึ้นของประจุภายในของเมมเบรนโพสซินแนปติก สิ่งนี้นำไปสู่การเกิดขึ้น หน้าแรก กายวิภาคของมนุษย์ กระแสนิ่ง สามารถตรวจพบความต่างศักย์ได้หากอิเล็กโทรดออสซิลโลสโคปถูกนำไปใช้กับกล้ามเนื้อที่เสียหายในลักษณะที่หนึ่งในนั้น ... ประวัติการศึกษาของมนุษย์ กายวิภาคศาสตร์ สรีรวิทยาของมนุษย์ สถานที่ของมนุษย์ในธรรมชาติ เซลล์ เมแทบอลิซึมของเซลล์ ดิวิชั่น ... เมแทบอลิซึม การควบคุมการทำงานของร่างกาย ระบบประสาท รีเฟล็กซ์และรีเฟล็กซ์อาร์ค รีเฟล็กซ์แบบไม่มีเงื่อนไขและแบบมีเงื่อนไข เลือด พลาสมาเลือด ปฏิกิริยาของเลือด ...
1. ความหงุดหงิด (ปฏิกิริยา) ของเซลล์คือความสามารถ (คุณสมบัติ) ของพวกเขาในการตอบสนองต่ออิทธิพลภายนอกอย่างแข็งขันจากกิจกรรมรูปแบบใดรูปแบบหนึ่ง ตัวอย่างเช่น โดยการเพิ่มเมแทบอลิซึมและการเจริญเติบโต เร่งการแบ่งตัว การหลั่งสาร การเคลื่อนไหว แรงกระตุ้นไฟฟ้า
ความตื่นเต้นง่าย - ความสามารถของเซลล์ในการตอบสนองต่อการกระตุ้นด้วยการกระตุ้น (เช่นการสร้างศักยภาพในการดำเนินการ) เนื้อเยื่อที่กระตุ้น ได้แก่ ประสาทและกล้ามเนื้อ
การกระตุ้นคือการตอบสนองของเซลล์ที่กระตุ้นต่อการกระทำของสิ่งเร้า (ศักยะงาน)
เซลล์ที่สามารถกระตุ้น - กล้ามเนื้อ, เส้นประสาท, ต่อม - เรียกว่าตื่นเต้น เซลล์ที่กระตุ้นได้คือเซลล์ที่มีความตื่นเต้นง่ายรวมถึงองค์ประกอบของตัวรับความรู้สึก - ปลายประสาทและเซลล์ตัวรับพิเศษ ความตื่นเต้นง่ายของเซลล์เหล่านี้ทำให้มั่นใจได้ถึงปฏิกิริยาของจุลินทรีย์ ความตื่นเต้นง่ายยังพบได้ในสิ่งมีชีวิตเซลล์เดียว
IRRITANT-stimulus ผลกระทบใด ๆ ที่สามารถทำให้เกิด biol ปฏิกิริยาของเนื้อเยื่อสิ่งมีชีวิต การเปลี่ยนแปลงโครงสร้างและหน้าที่ ปฏิกิริยาของเนื้อเยื่อต่อ R. naz ระคายเคือง
เนื่องจากอิทธิพลภายนอกทำให้เกิดความตื่นเต้น สารระคายเคืองสามารถทำหน้าที่: สิ่งจูงใจ อิทธิพลใดๆ ที่สามารถทำให้เกิด biol ปฏิกิริยาของเนื้อเยื่อสิ่งมีชีวิต การเปลี่ยนแปลงโครงสร้างและหน้าที่ 1) เคมี ไฟฟ้า เครื่องกล ฯลฯ (2) ขีดจำกัด ขีดจำกัดสูงสุด ขีดจำกัดย่อย (3) เพียงพอและไม่เพียงพอ
การกระตุ้นที่เพียงพอสอดคล้องกับเซลล์ประเภทนี้ มันทำให้เกิดการกระตุ้นแม้ในระดับพลังงาน (ปริมาณ) ที่ต่ำมาก เช่น แสง - สำหรับตัวรับแสง, เสียง - สำหรับตัวรับการได้ยิน, สารที่มีกลิ่น - สำหรับตัวรับกลิ่น ฯลฯ สำหรับเซลล์ประสาทและกล้ามเนื้อของสิ่งมีชีวิตหลายเซลล์ กระแสไฟฟ้าและสารเคมีบางชนิดที่ผลิตโดยเซลล์อื่น ๆ เป็นสิ่งเร้าที่เพียงพอ สิ่งเร้าอื่น ๆ ทั้งหมดเรียกว่าไม่เพียงพอ
พลังงานขั้นต่ำ (กำลัง) ของสิ่งเร้าที่จำเป็นต่อการกระตุ้นเซลล์เรียกว่าเกณฑ์ (เกณฑ์) ในกรณีของสิ่งเร้าที่ไม่เพียงพอ (เช่น การกระทำเชิงกลต่อเซลล์รับแสงหรือใยประสาท) สิ่งเร้านั้นจะมีค่าพลังงานเกินเกณฑ์สำหรับสิ่งเร้าที่เพียงพอโดยลำดับความสำคัญหลายลำดับ ความตื่นเต้นง่ายที่เกี่ยวข้องกับสิ่งเร้านั้นวัดจากระดับการระคายเคือง ความตื่นเต้นง่ายแปรผกผันกับค่าเกณฑ์
การกระตุ้นที่เกิดขึ้นที่จุดกระตุ้นของการกระทำในหลาย ๆ กรณีสามารถแพร่กระจายได้ครอบคลุมทั้งเซลล์ สิ่งนี้เกี่ยวข้องอย่างใกล้ชิดกับการตอบสนองทางไฟฟ้าและมีความสำคัญอย่างยิ่งต่อกิจกรรมของระบบประสาทและโดยเฉพาะอย่างยิ่งวิถีของมัน (การส่งสัญญาณประสาท)
เซลล์ที่กระตุ้นได้ในกรณีที่ไม่มีสิ่งเร้าเพียงพอมีลักษณะของการพักผ่อนทางสรีรวิทยาซึ่งแน่นอนว่าไม่เท่ากับการไม่มีการใช้งานอย่างสมบูรณ์เพราะมันเกี่ยวข้องกับการเผาผลาญอย่างต่อเนื่อง
อิทธิพลภายนอกบางอย่างสามารถทำให้เกิดปฏิกิริยากับสัญญาณเชิงลบในเซลล์ (การเผาผลาญอาหารลดลง, การเจริญเติบโต, ความตื่นเต้นง่ายลดลงเมื่อเทียบกับสิ่งเร้า) ปฏิกิริยาดังกล่าวเรียกว่าการยับยั้ง การยับยั้งอาจเกิดจากอิทธิพลจากสภาพแวดล้อมภายนอกและอิทธิพลจากเซลล์อื่นของร่างกาย
2. เยื่อชีวภาพทั้งหมดมีลักษณะโครงสร้างและคุณสมบัติทั่วไป ในปัจจุบัน แบบจำลองของเหลว-โมเสกของโครงสร้างเมมเบรนเป็นที่ยอมรับโดยทั่วไป
เมมเบรนประกอบด้วยชั้นไขมันเกิดจากฟอสโฟลิปิดเป็นส่วนใหญ่ ฟอสโฟลิปิดคือไตรกลีเซอไรด์ซึ่งกรดไขมันหนึ่งตัวถูกแทนที่ด้วยกรดฟอสฟอริก ส่วนของโมเลกุลซึ่งมีกรดฟอสฟอริกตกค้างอยู่เรียกว่าส่วนหัวที่ชอบน้ำ ส่วนที่มีกรดไขมันตกค้างอยู่เรียกว่าส่วนหางที่ไม่ชอบน้ำ ในเยื่อหุ้มเซลล์ ฟอสโฟลิพิดถูกจัดเรียงอย่างเป็นระเบียบ: หางของโมเลกุลที่ไม่ชอบน้ำจะหันเข้าหากัน และส่วนหัวที่ชอบน้ำจะหันออกด้านนอกเข้าหาน้ำ
นอกจากไขมันแล้ว เยื่อหุ้มประกอบด้วยโปรตีน(เฉลี่ย ≈ 60%) พวกเขากำหนดหน้าที่เฉพาะส่วนใหญ่ของเมมเบรน (การขนส่งโมเลกุลบางชนิด การเร่งปฏิกิริยาของปฏิกิริยา การรับและการแปลงสัญญาณจากสิ่งแวดล้อม ฯลฯ) มี: 1) โปรตีนส่วนปลาย (อยู่ที่ผิวด้านนอกหรือด้านในของชั้นไขมัน), 2) โปรตีนกึ่งอินทิกรัล (แช่อยู่ในชั้นไขมันที่ระดับความลึกต่างกัน), 3) อินทิกรัลหรือทรานส์เมมเบรน, โปรตีน (ซึมผ่านเยื่อหุ้มเซลล์ ผ่านในขณะที่สัมผัสกับภายนอกและกับสภาพแวดล้อมภายในเซลล์) โปรตีนอินทิกรัลในบางกรณีเรียกว่าการก่อตัวเป็นช่องหรือช่อง เนื่องจากสามารถพิจารณาได้ว่าเป็นช่องทางที่ชอบน้ำซึ่งโมเลกุลมีขั้วผ่านเข้าไปในเซลล์ (ส่วนประกอบที่เป็นไขมันของเยื่อหุ้มเซลล์จะไม่ยอมให้ผ่าน)
โครงสร้างเมมเบรน: A - หัวที่ชอบน้ำของฟอสโฟลิปิด B - หางที่ไม่ชอบน้ำของฟอสโฟลิปิด 1 - บริเวณที่ไม่ชอบน้ำของโปรตีน E และ F; 2 - บริเวณที่ชอบน้ำของโปรตีน F; 3 - สายโซ่โอลิโกแซ็กคาไรด์ที่แตกแขนงติดอยู่กับไขมันในโมเลกุลไกลโคลิพิด (ไกลโคลิปิดพบได้น้อยกว่าไกลโคโปรตีน) 4 - สายโซ่โอลิโกแซ็กคาไรด์ที่แตกแขนงติดอยู่กับโปรตีนในโมเลกุลไกลโคโปรตีน 5 - ช่องที่ชอบน้ำ (ทำหน้าที่เป็นรูพรุนซึ่งไอออนและโมเลกุลมีขั้วสามารถผ่านได้)
เมมเบรนอาจมีคาร์โบไฮเดรต (ถึง 10%) ส่วนประกอบคาร์โบไฮเดรตของเมมเบรนแสดงด้วยสายโซ่โอลิโกแซ็กคาไรด์หรือโพลีแซ็กคาไรด์ที่เกี่ยวข้องกับโมเลกุลโปรตีน (ไกลโคโปรตีน) หรือไขมัน (ไกลโคลิพิด) โดยพื้นฐานแล้ว คาร์โบไฮเดรตจะอยู่ที่ผิวด้านนอกของเมมเบรน คาร์โบไฮเดรตทำหน้าที่รับของเมมเบรน ในเซลล์สัตว์ ไกลโคโปรตีนก่อตัวเป็น epimembrane complex ซึ่งก็คือ glycocalyx ซึ่งมีความหนาหลายสิบนาโนเมตร ตัวรับเซลล์จำนวนมากอยู่ในนั้นโดยช่วยให้เกิดการยึดเกาะของเซลล์
โมเลกุลของโปรตีน คาร์โบไฮเดรต และลิพิด เคลื่อนที่ได้ สามารถเคลื่อนที่ในระนาบของเยื่อหุ้มเซลล์ได้ ความหนาของพลาสมาเมมเบรนอยู่ที่ประมาณ 7.5 นาโนเมตร
หน้าที่ของเมมเบรน: 1. การแยกเนื้อหาของเซลล์ออกจากสภาพแวดล้อมภายนอก 2. การควบคุมเมแทบอลิซึมระหว่างเซลล์กับสิ่งแวดล้อม 3. การแบ่งเซลล์ออกเป็นอพาร์ทเมนท์ (“ช่อง”) 4. ตำแหน่งของ “ท่อลำเลียงเอนไซม์” 5. การตรวจสอบ การสื่อสารระหว่างเซลล์ในเนื้อเยื่อของสิ่งมีชีวิตหลายเซลล์ (การยึดเกาะ) 6.การรู้จำสัญญาณ
ที่สำคัญที่สุด คุณสมบัติเมมเบรน- การซึมผ่านที่เลือกได้เช่น เมมเบรนสามารถซึมผ่านได้ดีกับสารหรือโมเลกุลบางชนิดและซึมผ่านได้ไม่ดี (หรือผ่านไม่ได้ทั้งหมด) ไปยังสารอื่น คุณสมบัตินี้รองรับการทำงานด้านกฎระเบียบของเมมเบรน ซึ่งช่วยให้เกิดการแลกเปลี่ยนสารระหว่างเซลล์กับสิ่งแวดล้อมภายนอก
3. กระบวนการผ่านของสารผ่านเยื่อหุ้มเซลล์เรียกว่าการขนส่งสาร แยกแยะ: 1) การขนส่งแบบพาสซีฟ - กระบวนการส่งผ่านสารโดยไม่ใช้พลังงาน 2) การขนส่งแบบแอคทีฟ - กระบวนการส่งผ่านสารโดยมีค่าใช้จ่ายด้านพลังงาน 3) การขนส่งโดยมีการเปลี่ยนแปลงโครงสร้างของเยื่อหุ้มเซลล์ (exocytosis, endocytosis) หรือไม่มีการเปลี่ยนแปลงโครงสร้างของเยื่อหุ้มเซลล์ (รูปแบบการขนส่งอื่น ๆ ทั้งหมด) 4) นี่คือการขนส่งที่เกี่ยวข้องกับการถ่ายโอนสารสองชนิด (cotransport) ซึ่งสามารถดำเนินการตามประเภทของ symport (สารสองชนิดไปในทิศทางของน้ำ - เช่น Na + กลูโคส) หรือตามประเภทของ antiport ( สารหนึ่งเข้าไปในเซลล์สารตัวที่สองออกจากเซลล์หรือในทางกลับกัน - Na K) แอนติโพดของการขนส่งร่วมคือการขนส่งธรรมดาหรือยูนิพอร์ต กล่าวคือ เมื่อสารหนึ่งถูกถ่ายโอน ตัวอย่างเช่น โมเลกุลของกลูโคส
1-ด้วยการขนส่งแบบพาสซีฟ สารจะเคลื่อนที่จากบริเวณที่มีความเข้มข้นสูงกว่าไปยังบริเวณที่มีความเข้มข้นต่ำกว่า เช่น ตามระดับความเข้มข้น ในสารละลายใดๆ มีโมเลกุลของตัวทำละลายและตัวถูกละลาย กระบวนการเคลื่อนที่ของโมเลกุลของตัวถูกละลายเรียกว่า การแพร่ การเคลื่อนที่ของโมเลกุลของตัวทำละลายเรียกว่า ออสโมซิส หากโมเลกุลมีประจุ การขนส่งจะได้รับผลกระทบจากการไล่ระดับสีทางไฟฟ้า ดังนั้น เรามักจะพูดถึงการไล่ระดับสีทางเคมีไฟฟ้า ซึ่งรวมการไล่ระดับสีทั้งสองเข้าด้วยกัน ความเร็วของการขนส่งขึ้นอยู่กับขนาดของการไล่ระดับสี
สามารถแยกแยะประเภทของการขนส่งแบบพาสซีฟต่อไปนี้: 1) การแพร่กระจายอย่างง่าย - การขนส่งสารโดยตรงผ่านชั้นไขมัน (ออกซิเจน, คาร์บอนไดออกไซด์); 2) การแพร่กระจายผ่านช่องเมมเบรน - ขนส่งผ่านโปรตีนที่สร้างช่อง (Na+, K+, Ca2+, Cl-); 3) อำนวยความสะดวกในการแพร่กระจาย - การขนส่งสารโดยใช้โปรตีนขนส่งพิเศษซึ่งแต่ละชนิดมีหน้าที่ในการเคลื่อนที่ของโมเลกุลหรือกลุ่มของโมเลกุลที่เกี่ยวข้อง (กลูโคส, กรดอะมิโน, นิวคลีโอไทด์); 4) ออสโมซิส - การขนส่งโมเลกุลของน้ำ (ในระบบชีวภาพทั้งหมด น้ำเป็นตัวทำละลาย)
2- ความจำเป็นในการขนส่งแบบแอคทีฟเกิดขึ้นเมื่อจำเป็นเพื่อให้แน่ใจว่ามีการถ่ายโอนโมเลกุลข้ามเมมเบรนกับการไล่ระดับสีด้วยไฟฟ้าเคมี การขนส่งนี้ดำเนินการโดยโปรตีนพาหะพิเศษซึ่งเป็นกิจกรรมที่ต้องใช้พลังงาน แหล่งพลังงานคือโมเลกุล ATP
Na+/K+-pump (ปั๊มโซเดียม-โพแทสเซียม)การทำงานของปั๊ม Na+/K+ สำหรับการทำงานปกติ เซลล์จะต้องรักษาอัตราส่วนของไอออน K + และ Na + ในไซโตพลาสซึมและในสภาพแวดล้อมภายนอก ความเข้มข้นของ K+ ภายในเซลล์ควรสูงกว่าภายนอกอย่างมีนัยสำคัญ และ Na+ - ในทางกลับกัน ควรสังเกตว่า Na+ และ K+ สามารถแพร่ผ่านเยื่อเมมเบรนได้อย่างอิสระ ปั๊ม Na+/K+ จะต่อต้านการทำให้ความเข้มข้นของไอออนเท่ากัน และปั๊ม Na+ ออกจากเซลล์และ K+ เข้าไปในเซลล์อย่างแข็งขัน เครื่องสูบน้ำ Na+/K+ เป็นโปรตีนเมมเบรนที่สามารถเปลี่ยนแปลงโครงสร้างได้ จึงสามารถติดทั้ง K+ และ Na+ ได้ วงจรการทำงานของปั๊ม Na+/K+ สามารถแบ่งออกเป็นระยะต่างๆ ต่อไปนี้: 1) การเกาะของ Na+ จากด้านในของเมมเบรน 2) ฟอสโฟรีเลชั่นของปั๊มโปรตีน 3) การปล่อย Na+ ในพื้นที่นอกเซลล์ 4) การยึดติดของ K+ จากภายนอกเมมเบรน 5) โปรตีน dephosphorylation pump 6) การปลดปล่อย K+ ในพื้นที่ภายในเซลล์ ปั๊มโซเดียมโพแทสเซียมใช้พลังงานเกือบหนึ่งในสามของพลังงานทั้งหมดที่จำเป็นสำหรับชีวิตของเซลล์ ในการทำงานหนึ่งรอบ ปั๊มจะสูบจ่าย 3Na+ ออกจากเซลล์และปั๊มออกเป็น 2K+
3-เอนโดไซโทซิส- กระบวนการดูดซึมโดยเซลล์ของอนุภาคขนาดใหญ่และโมเลกุลขนาดใหญ่ endocytosis มีสองประเภท: 1) phagocytosis - การจับและการดูดซึมของอนุภาคขนาดใหญ่ (เซลล์, ส่วนของเซลล์, โมเลกุลขนาดใหญ่) และ 2) pinocytosis - การจับและการดูดซึมของของเหลว (สารละลาย, สารละลายคอลลอยด์, สารแขวนลอย) ปรากฏการณ์ฟาโกไซโทซิสถูกค้นพบโดย I.I. เมชนิคอฟในปี พ.ศ. 2425 ในระหว่างการสร้างเอนโดไซโทซิส พลาสมาเมมเบรนจะสร้างการบุกรุก ขอบของมันผสานกัน และโครงสร้างที่แยกออกจากไซโตพลาสซึมโดยเมมเบรนเดี่ยวจะถูกเชื่อมเข้าไปในไซโตพลาสซึม โปรโตซัวจำนวนมากและเซลล์เม็ดเลือดขาวบางชนิดมีความสามารถในการทำลายเซลล์ พบ Pinocytosis ในเซลล์เยื่อบุผิวของลำไส้ใน endothelium ของเส้นเลือดฝอย
เอ็กโซไซโทซิส- กระบวนการย้อนกลับของ endocytosis: การกำจัดสารต่าง ๆ ออกจากเซลล์ ในระหว่างการเอ็กโซไซโทซิส เยื่อหุ้มตุ่มจะหลอมรวมกับเยื่อหุ้มไซโตพลาสซึมด้านนอก เนื้อหาของตุ่มจะถูกนำออกไปนอกเซลล์ และเยื่อหุ้มของมันรวมอยู่ในเยื่อหุ้มไซโตพลาสซึมด้านนอก ด้วยวิธีนี้ฮอร์โมนจะถูกขับออกจากเซลล์ของต่อมไร้ท่อ และในโปรโตซัว อาหารที่ไม่ย่อยจะยังคงอยู่
ช่องไอออนเป็นโปรตีนเมมเบรนที่ทำหน้าที่เป็นอนุภาคขนส่งสำหรับไอออนที่เกี่ยวข้อง เนื่องจากอนุภาคมีประจุภายในช่องไอออน ช่องไอออนแต่ละช่องมีปาก ตัวกรองแบบเลือก ประตู และกลไกควบคุมประตู บางช่องถูกควบคุมโดยความต่างศักย์ทั่วเมมเบรน ค่าของศักย์เมมเบรนจะเปิดประตูของช่องหรือปิดไว้ ตัวแปรที่สองของช่องไอออนคือช่องที่มีรีเซพเตอร์เกท: ในกรณีนี้ ประตูช่องจะถูกควบคุมโดยรีเซพเตอร์ที่อยู่บนพื้นผิวเมมเบรน: เมื่อสื่อกลางโต้ตอบกับรีเซพเตอร์นี้ ช่องไอออนสามารถเปิดได้ ช่องโซเดียมมีปาก ตัวกรองแบบเลือก และกลไกประตู พวกเขามีประตูสองประเภท - การเปิดใช้งาน (m-gates) และการปิดใช้งาน (p-gates) เมื่อ MP ลดลง (เช่น ถึง 60 mV) ประตูกระตุ้นจะเปิดขึ้นและปล่อยให้โซเดียมไอออนเข้าไปในเซลล์ แต่ในไม่ช้า ประตูปิดการใช้งานจะเริ่มปิด (การหยุดทำงานของช่องโซเดียมจะเกิดขึ้น) ในเวลาต่อมา ประตูการเปิดใช้งานจะปิด ประตูการปิดใช้งานจะเปิดขึ้น และช่องพร้อมสำหรับรอบใหม่ ช่องทางถูกปิดกั้นโดย tetrodotoxin, ยาชาเฉพาะที่ (โนโวเคน, สารอื่นๆ) นี้ใช้ในทางการแพทย์ ช่องโพแทสเซียมก็ค่อนข้างเลือกเช่นกัน - ส่วนใหญ่ปล่อยให้โพแทสเซียมไอออนผ่าน พวกมันถูกบล็อกโดยเตตระเอทิลแอมโมเนียม กระบวนการปิดใช้งานของพวกเขาแสดงออกอย่างอ่อน ช่องแคลเซียม - มีคุณสมบัติทั้งหมดของช่องไอออน (ปาก, กลไกประตู, ตัวกรอง) พวกมันถูกบล็อกโดยไอออนของแมงกานีส, นิกเกิล, แคดเมียม (ไดวาเลนต์ไอออน) รวมถึงสารทางยาเช่น verapamil, nifedipine, diltiazem ซึ่งใช้ในการปฏิบัติทางคลินิก
4. เนื้อเยื่อที่น่าตื่นเต้นและคุณสมบัติทั่วไป
เนื้อเยื่อกระตุ้นเป็นโครงสร้างประสาท กล้ามเนื้อ และต่อมที่สามารถตื่นเต้นได้เองหรือตอบสนองต่อการกระทำของสารระคายเคือง การกระตุ้นคือการสร้างศักยภาพในการดำเนินการ (AP) + การแพร่กระจายของ AP + การตอบสนองของเนื้อเยื่อเฉพาะต่อศักยภาพนี้ ตัวอย่างเช่น การหดตัว การปลดปล่อยความลับ การปลดปล่อยควอนตัมสื่อกลาง
คุณสมบัติของเนื้อเยื่อและตัวบ่งชี้ที่น่าตื่นเต้น:
คุณสมบัติ
1. Excitability - ความสามารถในการตื่นเต้น
2. การนำไฟฟ้า - ความสามารถในการกระตุ้นเช่น ดำเนินการ PD
3. การหดตัว - ความสามารถในการพัฒนาแรงหรือความตึงเครียดเมื่อตื่นเต้น
4. Lability - หรือ functional mobility - ความสามารถในการทำกิจกรรมเข้าจังหวะ
5. ความสามารถในการหลั่งความลับ (กิจกรรมหลั่ง) ผู้ไกล่เกลี่ย
ตัวบ่งชี้
1. เกณฑ์การระคายเคือง, rheobase, chronaxy, ระยะเวลาของระยะการทนไฟสัมบูรณ์, อัตราที่พัก
2. ความเร็วของ PD ตัวอย่างเช่นในเส้นประสาทสามารถไปถึง 120 m / s (ประมาณ 600 km / h)
3. ค่าสูงสุดของแรง (แรงดัน) ที่พัฒนาขึ้นระหว่างการกระตุ้น
4. จำนวนการกระตุ้นสูงสุดต่อหน่วยเวลา ตัวอย่างเช่น เส้นประสาทสามารถสร้าง AP ได้ 1,000 จุดใน 1 วินาที
5. ปรากฏการณ์ทางไฟฟ้าในเนื้อเยื่อที่กระตุ้น
กฎหมายระยะเวลาบังคับ:ผลกระทบที่น่ารำคาญของกระแสตรงไม่เพียงขึ้นอยู่กับขนาดของมันเท่านั้น แต่ยังขึ้นอยู่กับเวลาที่มันทำหน้าที่ด้วย ยิ่งกระแสมากเท่าไหร่ เวลาที่ต้องกระตุ้นก็จะยิ่งน้อยลงเท่านั้น
การศึกษาการพึ่งพาระยะเวลาของแรงแสดงให้เห็นว่าสิ่งหลังมีลักษณะเกินความจริง จากนี้ไปกระแสที่ต่ำกว่าค่าต่ำสุดที่แน่นอนจะไม่ทำให้เกิดการกระตุ้นไม่ว่าจะทำงานนานแค่ไหน และยิ่งพัลส์ปัจจุบันสั้นลง ก็ยิ่งระคายเคืองน้อยลง เหตุผลสำหรับ "การพึ่งพา" นี้คือความจุของเมมเบรน กระแส "สั้น" มากเพียงแค่ไม่มีเวลาปล่อยความจุนี้ไปยังระดับวิกฤตของการดีโพลาไรเซชัน , และทำให้เกิดความตื่นตัวเรียกว่าเวลาที่มีประโยชน์
เนื่องจากคำจำกัดความของเวลานี้เป็นเรื่องยากจึงนำเสนอแนวคิด ลำดับเหตุการณ์- เวลาต่ำสุดที่กระแสไฟฟ้าเท่ากับสองรีโอเบสต้องทำหน้าที่บนเนื้อเยื่อเพื่อให้เกิดการตอบสนอง คำจำกัดความของ chronaxia - chronaximetry - ใช้ในคลินิก กระแสไฟฟ้าที่ใช้กับกล้ามเนื้อจะผ่านทั้งกล้ามเนื้อและเส้นใยประสาทและปลายของพวกมันจะอยู่ในกล้ามเนื้อนี้ เนื่องจาก chronaxy ของเส้นใยประสาทนั้นน้อยกว่า chronaxy ของเส้นใยกล้ามเนื้อ เมื่อตรวจสอบ chronaxy กล้ามเนื้อจะได้รับ chronaxy ของเส้นใยประสาท หากเส้นประสาทได้รับความเสียหายหรือ motoneurons ที่สอดคล้องกันของไขสันหลังตาย (สิ่งนี้เกิดขึ้นกับโรคโปลิโอและโรคอื่น ๆ ) เส้นใยประสาทจะเสื่อมลงและจากนั้นจะมีการกำหนดลำดับของเส้นใยกล้ามเนื้อซึ่งมากกว่าเส้นใยประสาท .
กฎแห่งความแข็งแกร่ง. เพื่อให้เกิดความตื่นตัว สิ่งกระตุ้นต้องแรงพอ - เกณฑ์หรือสูงกว่าเกณฑ์ โดยปกติแล้ว คำว่า "เกณฑ์" หมายถึงความแรงขั้นต่ำของสิ่งเร้าที่สามารถทำให้เกิดการกระตุ้นได้ ตัวอย่างเช่น ในการกระตุ้นให้เซลล์ประสาทกระตุ้นที่ MP = -70 mV และ FAC = -50 mV แรงเกณฑ์ควรเท่ากับ -20 mV แรงกระตุ้นที่ลดลงจะไม่ทำให้เกิดการตอบสนอง
ผลที่สำคัญอย่างหนึ่งของกฎหมายนี้คือการแนะนำแนวคิดของ "เกณฑ์กระตุ้น" (ระดับต่ำสุดของการกระตุ้นที่สามารถทำให้เกิดการตื่นตัว) การกำหนดตัวบ่งชี้นี้
กฎหมายทั้งหมดหรือไม่มีเลย:ภายใต้สิ่งเร้าที่เป็นเกณฑ์ไม่ทำให้เกิดการตอบสนอง ("ไม่มีอะไร") ในสิ่งเร้าที่เป็นเกณฑ์มีการตอบสนองสูงสุด ("ทุกอย่าง") ตามกฎ "ทั้งหมดหรือไม่มีเลย" กล้ามเนื้อหัวใจและเส้นใยกล้ามเนื้อเส้นเดียวจะหดตัว กฎหมายทั้งหมดหรือไม่มีเลยนั้นไม่สมบูรณ์แบบ ประการแรก ไม่มีการตอบสนองที่มองเห็นได้ต่อสิ่งเร้าที่มีระดับต่ำกว่าเกณฑ์ แต่การเปลี่ยนแปลงของศักยภาพเยื่อหุ้มเซลล์ที่พักตัวเกิดขึ้นในเนื้อเยื่อในรูปแบบของการกระตุ้นเฉพาะที่ (การตอบสนองเฉพาะที่) ประการที่สอง กล้ามเนื้อหัวใจที่ยืดออกด้วยเลือดเมื่อเติมเต็มห้องของหัวใจจะทำปฏิกิริยาตามกฎทั้งหมดหรือไม่มีอะไรเลย แต่ความกว้างของการหดตัวจะมากกว่าเมื่อเทียบกับการหดตัวของกล้ามเนื้อหัวใจที่ไม่ได้ยืดออกด้วย เลือด.
5. ศักยภาพของเมมเบรนและแหล่งกำเนิด
MP หรือค่าศักย์ไฟฟ้าขณะพัก คือค่าความต่างศักย์ระหว่างพื้นผิวด้านนอกและด้านในของเมมเบรนที่เหลือ โดยเฉลี่ยแล้ว ในเซลล์ของเนื้อเยื่อที่กระตุ้นได้ จะมีค่าถึง 50–80 mV โดยมีเครื่องหมาย “–” อยู่ภายในเซลล์ สาเหตุหลักมาจากโพแทสเซียมไอออน อย่างที่ทราบกันดีว่าในเซลล์ของเนื้อเยื่อที่กระตุ้นได้ความเข้มข้นของโพแทสเซียมไอออนถึง 150 มิลลิโมล / ลิตรในสิ่งแวดล้อม - 4-5 มิลลิโมล (ไอออนโพแทสเซียมมีอยู่ในเซลล์มากกว่าในสิ่งแวดล้อม) ดังนั้นตามระดับความเข้มข้นโพแทสเซียมสามารถออกจากเซลล์ได้และสิ่งนี้เกิดขึ้นกับการมีส่วนร่วมของช่องโพแทสเซียมซึ่งบางช่องเปิดอยู่ เป็นผลให้เนื่องจากความจริงที่ว่าเมมเบรนไม่สามารถผ่านเข้าไปในเซลล์แอนไอออน (กลูตาเมต, แอสปาร์เตต, ฟอสเฟตอินทรีย์) อนุภาคที่มีประจุลบส่วนเกินจะเกิดขึ้นที่พื้นผิวด้านในของเซลล์และเกิดอนุภาคที่มีประจุบวกมากเกินไป บนพื้นผิวด้านนอก มีความต่างศักย์ ยิ่งความเข้มข้นของโพแทสเซียมในตัวกลางสูงเท่าใดอัตราส่วนนี้ก็จะยิ่งต่ำลงเท่านั้น ค่าของศักย์เมมเบรนก็จะยิ่งต่ำลงเท่านั้น อย่างไรก็ตาม ค่าที่คำนวณได้มักจะต่ำกว่าค่าจริง ตัวอย่างเช่น ตามการคำนวณ MP ควรเป็น -90 mV แต่ในความเป็นจริง -70 mV ความแตกต่างนี้เกิดจากการที่ไอออนของโซเดียมและคลอไรด์มีส่วนในการสร้างสนามแม่เหล็ก โดยเฉพาะอย่างยิ่ง เป็นที่ทราบกันดีว่ามีโซเดียมในตัวกลางมากกว่า (140 มิลลิโมล/ลิตร เทียบกับ 14 มิลลิโมล/ลิตรในเซลล์) โซเดียมจึงสามารถเข้าสู่เซลล์ได้ แต่ช่องโซเดียมส่วนใหญ่จะปิดเมื่อพัก ดังนั้นโซเดียมไอออนเพียงส่วนน้อยจึงเข้าสู่เซลล์ แต่ถึงอย่างนั้นก็เพียงพอที่จะชดเชยประจุลบส่วนเกินอย่างน้อยบางส่วน ในทางตรงกันข้ามคลอรีนไอออนจะเข้าสู่เซลล์ (บางส่วน) และนำประจุลบเข้ามา เป็นผลให้ค่าของศักยภาพเมมเบรนถูกกำหนดโดยโพแทสเซียมเป็นหลัก เช่นเดียวกับโซเดียมและคลอรีน
เพื่อรักษาสนามแม่เหล็กให้อยู่ในระดับคงที่จำเป็นต้องรักษาความไม่สมดุลของไอออนิก - ความไม่สมมาตรของไอออนิก โดยเฉพาะอย่างยิ่งสำหรับจุดประสงค์นี้จะใช้ปั๊มโพแทสเซียมโซเดียม (และคลอไรด์) ซึ่งจะคืนค่าความไม่สมดุลของไอออนิกโดยเฉพาะอย่างยิ่งหลังจากการกระตุ้น ข้อพิสูจน์ถึงธรรมชาติของโพแทสเซียมของสนามแม่เหล็กคือการพึ่งพา: ยิ่งความเข้มข้นของโพแทสเซียมในตัวกลางสูง ค่าของสนามแม่เหล็กก็จะยิ่งต่ำลง สำหรับการนำเสนอเพิ่มเติม แนวคิดมีความสำคัญ: ดีโพลาไรเซชัน (การลดลงของสนามแม่เหล็ก เช่น จากลบ 90 mV เป็นลบ 70 mV) และไฮเปอร์โพลาไรเซชันเป็นปรากฏการณ์ที่ตรงกันข้าม
6. ศักยภาพในการดำเนินการ- นี่คือการเปลี่ยนแปลงระยะสั้นในความต่างศักย์ระหว่างพื้นผิวด้านนอกและด้านในของเมมเบรน (หรือระหว่างจุดสองจุดในเนื้อเยื่อ) ซึ่งเกิดขึ้นในขณะที่มีการกระตุ้น ศักยภาพในการดำเนินการอาจแตกต่างกันในพารามิเตอร์ ขึ้นอยู่กับชนิดของเซลล์และแม้แต่ในส่วนต่างๆ ของเยื่อหุ้มเซลล์เดียวกัน ตัวอย่างที่โดดเด่นที่สุดของความแตกต่างคือศักยภาพในการออกฤทธิ์ของกล้ามเนื้อหัวใจและศักยภาพในการออกฤทธิ์ของเซลล์ประสาทส่วนใหญ่ อย่างไรก็ตาม ปรากฏการณ์ต่อไปนี้สนับสนุนศักยภาพของการกระทำใดๆ:
1 เมมเบรนของเซลล์ที่มีชีวิตถูกโพลาไรซ์ - พื้นผิวด้านในมีประจุลบเมื่อเทียบกับด้านนอกเนื่องจากในสารละลายใกล้กับพื้นผิวด้านนอกมีอนุภาคที่มีประจุบวก (ไอออนบวก) มากกว่าและใกล้กับพื้นผิวด้านใน - อนุภาคที่มีประจุลบ (แอนไอออน) มากขึ้น
เมมเบรน 2 ชนิดมีความสามารถในการซึมผ่านแบบเลือกได้ - ความสามารถในการซึมผ่านของอนุภาคต่างๆ (อะตอมหรือโมเลกุล) ขึ้นอยู่กับขนาด ประจุไฟฟ้า และคุณสมบัติทางเคมี
เมมเบรน 3 ชั้นของเซลล์ที่กระตุ้นได้สามารถเปลี่ยนการซึมผ่านของไอออนบวกบางประเภทได้อย่างรวดเร็ว ทำให้ประจุบวกส่งผ่านจากภายนอกสู่ภายใน
คุณสมบัติสองประการแรกเป็นลักษณะของเซลล์ที่มีชีวิตทั้งหมด ประการที่สามคือคุณสมบัติของเซลล์ของเนื้อเยื่อที่กระตุ้นได้และสาเหตุที่เยื่อหุ้มของพวกมันสามารถสร้างและนำศักยภาพในการดำเนินการได้
ขั้นตอนศักยภาพในการดำเนินการ
1. Prespike - กระบวนการของการสลับขั้วอย่างช้าๆของเมมเบรนจนถึงระดับวิกฤตของการสลับขั้ว (การกระตุ้นในท้องถิ่น, การตอบสนองในท้องถิ่น)
2. ศักย์ไฟฟ้าสูงสุดหรือสไปค์ ประกอบด้วยส่วนที่เพิ่มขึ้น (การสลับโพลาไรเซชันของเมมเบรน) และส่วนจากมากไปน้อย (การรีโพลาไรเซชันของเมมเบรน) มีขั้นตอนหรือองค์ประกอบดังต่อไปนี้: การตอบสนองในพื้นที่เป็นขั้นตอนเริ่มต้นของการสลับขั้ว ข. ระยะดีโพลาไรเซชันคือการลดลงอย่างรวดเร็วของศักย์เมมเบรนเป็นศูนย์และการเติมประจุของเมมเบรน (การพลิกกลับหรือการโอเวอร์ชูต) วี. ขั้นตอนการรีโพลาไรเซชัน - การฟื้นฟูระดับเริ่มต้นของศักยภาพของเมมเบรน
ในนั้นเฟสของโพลาไรเซชันอย่างรวดเร็วและเฟสของโพลาไรเซชันช้านั้นแตกต่างกัน ในทางกลับกันเฟสของโพลาไรเซชันช้าจะแสดงโดยกระบวนการติดตาม (ศักยภาพ): การปฏิเสธการติดตาม (การติดตามการสลับขั้ว) และการติดตามเชิงบวก (การติดตามไฮเปอร์โพลาไรเซชัน)
3. ศักยภาพการติดตามเชิงลบ - จากระดับวิกฤตของการสลับขั้วไปจนถึงระดับเริ่มต้นของโพลาไรเซชันของเมมเบรน (การติดตามการสลับขั้ว)
4. ศักยภาพในการติดตามที่เป็นบวก - การเพิ่มขึ้นของศักยภาพของเยื่อหุ้มเซลล์และการกลับสู่ค่าเดิมอย่างค่อยเป็นค่อยไป (การติดตามไฮเปอร์โพลาไรเซชัน)
รูปร่างของศักยภาพในการดำเนินการ (ระหว่างการบันทึกภายในเซลล์) ขึ้นอยู่กับชนิดของเนื้อเยื่อที่กระตุ้นได้: ในแอกซอนของเซลล์ประสาท, กล้ามเนื้อโครงร่าง - ศักยภาพคล้ายจุดสูงสุด, ในกล้ามเนื้อเรียบในบางกรณีคล้ายจุดสูงสุด, อื่น ๆ - คล้ายที่ราบสูง (ตัวอย่างเช่น ศักยภาพในการดำเนินการของกล้ามเนื้อเรียบของมดลูกของหญิงตั้งครรภ์มีรูปร่างที่ราบสูง และระยะเวลาของมันคือเกือบ 1 นาที) ในกล้ามเนื้อหัวใจ ศักยภาพในการดำเนินการมีรูปร่างที่ราบสูง
7. ความตื่นเต้นง่าย - ความสามารถของสิ่งมีชีวิตในการรับรู้การเปลี่ยนแปลงในสภาพแวดล้อมภายนอกและตอบสนองต่อการเปลี่ยนแปลงเหล่านี้ (การระคายเคือง) ด้วยปฏิกิริยากระตุ้น การกระตุ้นเกี่ยวข้องกับการดำรงอยู่ในเยื่อหุ้มเซลล์ของโครงสร้างโมเลกุลพิเศษที่มีความไวเฉพาะต่อการกระทำของสิ่งเร้าบางอย่าง: กระแสไฟฟ้า, เคมี, กล, ความร้อนและอิทธิพลอื่น ๆ ระดับของการกระตุ้นเซลล์ขึ้นอยู่กับระยะ AP ในขั้นตอนการตอบสนองในท้องถิ่น ความตื่นเต้นง่ายจะเพิ่มขึ้น ขั้นตอนของความตื่นเต้นง่ายนี้เรียกว่าการเติมแฝง
ในขั้นตอนของ AP repolarization เมื่อช่องโซเดียมทั้งหมดเปิดออกและโซเดียมไอออนพุ่งเข้าสู่เซลล์ราวกับหิมะถล่ม ไม่มีแม้แต่ตัวกระตุ้นที่แรงมากก็สามารถกระตุ้นกระบวนการนี้ได้ ดังนั้นเฟสของการดีโพลาไรเซชันจึงสอดคล้องกับเฟสของการไม่ปลุกปั่นโดยสมบูรณ์หรือการหักเหของแสงสัมบูรณ์
ในช่วงรีโพลาไรเซชัน ช่องโซเดียมจะปิดมากขึ้นเรื่อยๆ อย่างไรก็ตาม พวกมันสามารถเปิดได้อีกครั้งภายใต้การกระทำของสิ่งเร้าเหนือขีดจำกัด เหล่านั้น. ความตื่นเต้นเริ่มเพิ่มขึ้นอีกครั้ง สิ่งนี้สอดคล้องกับระยะของการไม่ปลุกระดมสัมพัทธ์หรือการหักเหสัมพัทธ์
ในระหว่างการสลับโพลาไรเซชันการติดตาม MP จะอยู่ในระดับวิกฤต ดังนั้นแม้แต่สิ่งเร้าก่อนเกณฑ์ก็อาจทำให้เกิดการกระตุ้นเซลล์ได้ ดังนั้นในเวลานี้ความตื่นเต้นของเธอจึงเพิ่มขึ้น ขั้นตอนนี้เรียกว่าขั้นตอนของความสูงส่งหรือความตื่นเต้นเหนือปกติ
ในช่วงเวลาที่เกิดไฮเปอร์โพลาไรเซชันการติดตาม MP จะสูงกว่าระดับเริ่มต้น เช่น KUD เพิ่มเติมและความตื่นเต้นง่ายจะลดลง เธออยู่ในช่วงของความตื่นเต้นที่ต่ำกว่าปกติ ควรสังเกตว่าปรากฏการณ์ที่พักยังเกี่ยวข้องกับการเปลี่ยนแปลงการนำไฟฟ้าของช่องไอออน หากกระแสดีโพลาไรซ์เพิ่มขึ้นอย่างช้าๆ สิ่งนี้จะนำไปสู่การปิดใช้งานโซเดียมบางส่วนและเปิดใช้งานช่องโพแทสเซียม ดังนั้นการพัฒนาของ PD จึงไม่เกิดขึ้น
8. ใยประสาท- กระบวนการของเซลล์ประสาทที่ปกคลุมด้วยเยื่อหุ้มเกลีย
ในส่วนต่าง ๆ ของระบบประสาท เปลือกหุ้มของเส้นใยประสาทแตกต่างกันอย่างมีนัยสำคัญในโครงสร้างของมัน ซึ่งรองรับการแบ่งเส้นใยทั้งหมดออกเป็นไมอีลินและไม่มีไมอีลิน ทั้งสองประกอบด้วยกระบวนการของเซลล์ประสาทที่อยู่ตรงกลางของเส้นใย จึงเรียกว่าแกนทรงกระบอก (แอกซอน) และเยื่อเกลียที่อยู่ล้อมรอบ
ขึ้นอยู่กับความเข้มของภาระการทำงาน เซลล์ประสาทสร้างเส้นใยชนิดหนึ่งหรือชนิดอื่น สำหรับส่วนร่างกายของระบบประสาทซึ่งทำให้กล้ามเนื้อโครงร่างมีระดับการทำงานสูงเส้นใยประสาทชนิดไมอีลินมีลักษณะเฉพาะและสำหรับส่วนที่เป็นพืชซึ่งทำให้อวัยวะภายในเป็นชนิดที่ไม่ใช่ไมอีลิน .
ใยประสาทถูกจัดประเภท โดย 1.ระยะเวลาที่เป็นไปได้ของการกระทำ 2. โครงสร้าง (เส้นผ่านศูนย์กลาง) ของเส้นใย 3. ความเร็วของการกระตุ้น
เส้นใยประสาทกลุ่มต่อไปนี้มีความโดดเด่น : 1. กลุ่ม A (อัลฟา, เบต้า, แกมมา, เดลต้า) - ศักยภาพในการดำเนินการที่สั้นที่สุด, ปลอกไมอีลินที่หนาที่สุด, อัตราการกระตุ้นสูงสุด 2. กลุ่ม B - ปลอกไมอีลินเด่นชัดน้อยกว่า 3.group C - ไม่มีปลอกไมอีลิน
คุณสมบัติของการกระตุ้นตามเส้นใยประสาท myelinated และ unmyelinated:
เส้นใยไมอีลิน - มีเปลือกหุ้มที่มีคุณสมบัติต้านทานไฟฟ้าสูง เฉพาะในส่วนสกัดกั้นของ Ranvier ภายใต้การกระทำของสิ่งเร้า ความตื่นเต้นจะเกิดขึ้นในการสกัดกั้น Ranvier ที่ใกล้ที่สุด การสกัดกั้นเพื่อนบ้านในสถานะโพลาไรซ์ กระแสที่เกิดขึ้นทำให้เกิดการสลับขั้วของการสกัดกั้นที่อยู่ติดกัน โหนดของ Ranvier มีความหนาแน่นสูงของ Na-channel ดังนั้นในแต่ละโหนดถัดไปจึงมีศักยภาพในการดำเนินการที่ใหญ่ขึ้นเล็กน้อย (ในแอมพลิจูด) ด้วยเหตุนี้การกระตุ้นจึงแพร่กระจายโดยไม่มีการลดลงและสามารถกระโดดข้ามโหนดต่างๆ นี่คือทฤษฎีเกลือของทาซากิ ข้อพิสูจน์ของทฤษฎีคือยาถูกฉีดเข้าไปในเส้นใยประสาทที่ปิดกั้นการสกัดกั้นหลายจุด แต่หลังจากนั้นก็มีการบันทึกการนำการกระตุ้น นี่เป็นวิธีที่เชื่อถือได้และให้ผลกำไรสูง เนื่องจากความเสียหายเล็กน้อยถูกกำจัดออกไป ความเร็วของการกระตุ้นเพิ่มขึ้น และต้นทุนด้านพลังงานลดลง
เส้นใยปราศจากไมอีลิน - พื้นผิวมีคุณสมบัติเป็นไฟฟ้าตลอด ดังนั้นกระแสวงกลมขนาดเล็กจึงเกิดขึ้นที่ระยะไม่กี่ไมโครเมตร ความตื่นเต้นมีรูปแบบของคลื่นที่เดินทางตลอดเวลา
วิธีนี้ให้ผลกำไรน้อยกว่า: ต้นทุนพลังงานสูง (สำหรับการทำงานของปั๊ม Na-K) อัตราการกระตุ้นที่ต่ำกว่า
มีกฎสามข้อของการระคายเคืองตามเส้นใยประสาท
กฎแห่งความสมบูรณ์ทางกายวิภาคและสรีรวิทยา การนำแรงกระตุ้นไปตามเส้นใยประสาทเป็นไปได้ก็ต่อเมื่อไม่มีการละเมิดความสมบูรณ์
กฎของการกระตุ้นแบบแยกตัว มีคุณสมบัติหลายประการของการแพร่กระจายของการกระตุ้นในเส้นใยประสาทส่วนปลาย, เยื่อและไม่ใช่ปอด ในใยประสาทส่วนปลาย การกระตุ้นจะส่งไปตามใยประสาทเท่านั้น แต่จะไม่ส่งไปยังใยประสาทข้างเคียงที่อยู่ในลำประสาทเดียวกัน ในเส้นใยประสาทที่เป็นเยื่อใย เยื่อไมอีลินทำหน้าที่เป็นฉนวน เนื่องจากไมอีลินทำให้ความต้านทานเพิ่มขึ้นและความจุไฟฟ้าของเปลือกลดลง ในเส้นใยประสาทที่ไม่ใช่เนื้อ การกระตุ้นจะถูกส่งแยกกัน
กฎหมายของการกระตุ้นทวิภาคี ใยประสาทนำกระแสประสาทในสองทิศทาง - ศูนย์กลางและแรงเหวี่ยง
9. คุณสมบัติทางสรีรวิทยาของกล้ามเนื้อ
ความตื่นเต้นง่าย - ความสามารถในการเข้าสู่สภาวะตื่นเต้นภายใต้การกระทำของสิ่งเร้า การนำไฟฟ้า - ความสามารถในการกระตุ้น การหดตัวคือความสามารถของกล้ามเนื้อในการเปลี่ยนความยาวหรือความตึงเพื่อตอบสนองต่อสิ่งเร้า Lability - ความแข็งแรงของกล้ามเนื้อคือ 200-300 Hz
ด้วยการกระตุ้นกล้ามเนื้อโดยตรง (การระคายเคืองโดยตรง) หรือโดยอ้อมผ่านเส้นประสาทสั่งการที่ทำให้กล้ามเนื้อกระตุก (การระคายเคืองทางอ้อม) การกระตุ้นเพียงครั้งเดียวจะเกิดขึ้น การหดตัวของกล้ามเนื้อเดี่ยวซึ่งมีสามขั้นตอนที่แตกต่างกัน: ระยะเวลาแฝง - เวลาตั้งแต่เริ่มมีอาการกระตุ้นจนถึงเริ่มมีการตอบสนอง; ระยะหดตัว (ระยะย่อ); ระยะผ่อนคลาย
การหดตัวของกล้ามเนื้อมีสองประเภท. หากปลายทั้งสองของกล้ามเนื้อยึดอยู่กับที่อย่างถาวร การหดตัวแบบไอโซเมตริกจะเกิดขึ้น และที่ความยาวคงที่ ความตึงจะเพิ่มขึ้น หากปลายด้านหนึ่งของกล้ามเนื้อว่าง ในกระบวนการหดตัว ความยาวของกล้ามเนื้อจะลดลงและความตึงเครียดจะไม่เปลี่ยนแปลง - การหดตัวดังกล่าวเรียกว่าไอโซโทนิก ในร่างกาย การหดตัวดังกล่าวมีความสำคัญต่อประสิทธิภาพของการเคลื่อนไหวใดๆ
บาดทะยัก กล้ามเนื้อเกร็งบาดทะยัก- สถานะของการหดตัวเป็นเวลานาน, ความตึงเครียดของกล้ามเนื้ออย่างต่อเนื่อง, ซึ่งเกิดขึ้นเมื่อกระแสประสาทที่มีความถี่สูงเข้ามาทางเซลล์ประสาทสั่งการ. ในเวลาเดียวกัน จะไม่มีการผ่อนคลายระหว่างการหดตัวเดี่ยวที่ต่อเนื่องกันและการรวมเข้าด้วยกัน ซึ่งนำไปสู่การหดตัวของกล้ามเนื้อสูงสุดที่มั่นคง
มีบาดทะยัก dentate และเรียบ ด้วยบาดทะยัก dentate แรงกระตุ้นของเส้นประสาทที่ตามมาแต่ละครั้งจะทำหน้าที่ในกล้ามเนื้อที่เริ่มคลายตัวและเกิดการหดตัวที่ไม่สมบูรณ์ ด้วยบาดทะยักแบบเรียบซึ่งมีแอมพลิจูดที่ใหญ่กว่า ผลกระทบของแรงกระตุ้นจะเกิดขึ้นเมื่อสิ้นสุดระยะเวลาที่สั้นลง ซึ่งนำไปสู่การรวมการหดตัวอย่างสมบูรณ์
10. คุณสมบัติของโครงสร้างและการส่งผ่านของการกระตุ้นในประสาทและกล้ามเนื้อประสาท ทฤษฎีสมัยใหม่ของการหดตัวของกล้ามเนื้อและการผ่อนคลาย
รอยต่อประสาทและกล้ามเนื้อประกอบด้วยโครงสร้างหลัก 3 โครงสร้าง ได้แก่ เยื่อหุ้มเซลล์ก่อนไซแนปติก รอยแหว่งไซแนปติก และเยื่อหุ้มโพสต์ซินแนปติก เยื่อหุ้มเซลล์ก่อนไซแนปติกครอบคลุมปลายประสาท และเยื่อหุ้มเซลล์หลังไซแนปติกหุ้มเซลล์เอฟเฟกเตอร์ ระหว่างพวกเขาคือรอยแยก synaptic เมมเบรนโพสต์ซินแนปติกแตกต่างจากพรีซินแนปติกตรงที่มีตัวรับเคมีโปรตีนที่ไวต่อสารไกล่เกลี่ยและฮอร์โมนเท่านั้น แต่ยังรวมถึงยาและสารที่เป็นพิษด้วย โครงสร้างของไซแนปส์ประสาทและกล้ามเนื้อกำหนดคุณสมบัติทางสรีรวิทยาของมัน:
1) การกระตุ้นข้างเดียว (จาก presynaptic ไปยังเมมเบรน postsynaptic) ในที่ที่มีตัวรับที่ไวต่อผู้ไกล่เกลี่ยเฉพาะในเมมเบรน postsynaptic;
2) ความล่าช้าของ synaptic ในการกระตุ้นซึ่งเกี่ยวข้องกับอัตราการแพร่กระจายต่ำของผู้ไกล่เกลี่ยเมื่อเทียบกับความเร็วของแรงกระตุ้นของเส้นประสาท
3) ความสามารถในการทำงานต่ำและความเมื่อยล้าของไซแนปส์สูง
4) ความไวในการเลือกสูงของไซแนปส์ต่อสารเคมี
การถ่ายโอนความตื่นเต้น
การกระตุ้นจะแพร่กระจายไปตามใยประสาทในรูปของศักยะงาน (แรงกระตุ้นเส้นประสาท) ไปถึงเยื่อหุ้มเซลล์ก่อนไซแนปติก ทำให้เกิดการสลับขั้ว ซึ่งนำไปสู่การเปิดช่องแคลเซียม ไอออน Ca2+ เข้าสู่ปลายประสาทและส่งเสริมการปลดปล่อยสื่อกลางจากถุงไซแนปติกและปล่อยเข้าไปในรอยแหว่งไซแนปติก ผู้ไกล่เกลี่ยกระจายอย่างรวดเร็วผ่านช่องว่างและทำหน้าที่ในเมมเบรนโพสต์ซินแนปติก - โต้ตอบกับตัวรับ (acetylcholine - กับตัวรับ cholinergic, norepinephrine - กับตัวรับ adrenoreceptor ฯลฯ ) เมมเบรนตอบสนองต่อปฏิสัมพันธ์ของผู้ไกล่เกลี่ยกับตัวรับโดยการเปลี่ยนการซึมผ่านของไอออน Na + และ K + ซึ่งนำไปสู่การสลับขั้ว การเกิดขึ้นของศักย์ไฟฟ้า และการสร้างศักย์ไฟฟ้าหลังไซแนปติกกระตุ้น ภายใต้อิทธิพลของศักยภาพนี้ การสลับขั้วของส่วนเมมเบรนที่อยู่ติดกับไซแนปส์จะเกิดขึ้น ดังนั้นศักยภาพในการดำเนินการจึงกระจายไปทั่วอวัยวะ ผู้ไกล่เกลี่ยจะถูกปล่อยเข้าไปใน synaptic cleft ไม่เพียง แต่ในระหว่างการกระตุ้นเท่านั้น แต่ยังรวมถึงช่วงพักด้วย
ไซแนปส์- นี่คือรูปแบบโครงสร้างและหน้าที่ที่รับประกันการเปลี่ยนแปลงของการกระตุ้นหรือการยับยั้งจากปลายเส้นใยประสาทไปยังเซลล์ที่อยู่ภายใน
โครงสร้างไซแนปส์:
1) เมมเบรน presynaptic (เมมเบรนไฟฟ้าในขั้วซอนสร้างไซแนปส์บนเซลล์กล้ามเนื้อ);
2) เมมเบรนโพสต์ซินแนปติก (เมมเบรนไฟฟ้าของเซลล์ที่มีการสร้างไซแนปส์)
3) synaptic cleft (ช่องว่างระหว่างเยื่อหุ้ม presynaptic และ postsynaptic นั้นเต็มไปด้วยของเหลวที่มีลักษณะคล้ายพลาสม่าของเลือดในองค์ประกอบ)
ซินแนปส์มีหลายประเภท
1. โดยการแปล:
1) ประสาทส่วนกลาง
2) ประสาทส่วนปลาย
2. การจำแนกประเภทของไซแนปส์ตามหน้าที่:
1) ไซแนปส์กระตุ้น;
2) การระงับประสาท
3. ตามกลไกของการส่งผ่านการกระตุ้นในประสาท:
1) สารเคมี
2) ไฟฟ้า
(อย่างจำเป็น)
ไซแนปส์ Myoneural (ประสาทและกล้ามเนื้อ) - เกิดจากแอกซอนของเซลล์ประสาทสั่งการและเซลล์กล้ามเนื้อ
แรงกระตุ้นของกระแสประสาทเกิดขึ้นที่บริเวณทริกเกอร์ของเซลล์ประสาท เคลื่อนที่ไปตามแอกซอนไปยังกล้ามเนื้อที่มีเส้นเอ็น ไปถึงปลายแอกซอน และในขณะเดียวกันก็เปลี่ยนโพลาไรซ์ของเยื่อหุ้มพรีซินแนปติก
หลังจากนั้นช่องโซเดียมและแคลเซียมจะเปิดออก และไอออน Ca จากสิ่งแวดล้อมรอบไซแนปส์จะเข้าสู่ปลายแอกซอน ในกระบวนการนี้ การเคลื่อนไหวแบบบราวเนียนของถุงจะถูกสั่งไปยังเยื่อพรีไซแนปติก Ca ไอออนกระตุ้นการเคลื่อนไหวของถุงน้ำ เมื่อไปถึงเยื่อ presynaptic ถุงจะแตกและปล่อย acetylcholine (4 Ca ไอออนจะปล่อย 1 ควอนตัมของ acetylcholine) รอยแยกซินแนปติกเต็มไปด้วยของเหลวที่คล้ายกับพลาสมาของเลือดในองค์ประกอบ การแพร่กระจายของ ACh จากเยื่อหุ้มเซลล์ก่อนไซแนปติกไปยังเยื่อหุ้มเซลล์หลังไซแนปติกเกิดขึ้นผ่านมัน แต่อัตราของมันจะต่ำมาก นอกจากนี้ การแพร่กระจายยังเป็นไปได้ตามเส้นใยเส้นใยที่อยู่ในรอยแหว่งไซแนปติก หลังจากการแพร่ ACh จะเริ่มโต้ตอบกับตัวรับเคมี (ChR) และ cholinesterase (ChE) ที่อยู่บนเยื่อหุ้มเซลล์หลังไซแนปติก
ตัวรับ cholinergic ทำหน้าที่รับและ cholinesterase ทำหน้าที่ของเอนไซม์ บนเมมเบรนโพสต์ซินแน็ปติกจะอยู่ดังนี้:
XP-XE-XP-XE-XP-XE
XP + AX \u003d MECP - ศักยภาพขนาดเล็กของแผ่นท้าย
จากนั้นจึงสรุปผล MECP อันเป็นผลมาจากการสรุป EPSP ถูกสร้างขึ้น - ศักยภาพของโพสต์ซินแนปติกแบบกระตุ้น เยื่อโพสต์ซินแนปติกมีประจุลบเนื่องจาก EPSP และในบริเวณที่ไม่มีไซแนปส์ (เส้นใยกล้ามเนื้อ) ประจุจะเป็นบวก ความต่างศักย์เกิดขึ้น ศักย์ไฟฟ้าจะก่อตัวขึ้น ซึ่งจะเคลื่อนที่ไปตามระบบการนำไฟฟ้าของเส้นใยกล้ามเนื้อ
ChE + ACh = การทำลาย ACh ให้กับโคลีนและกรดอะซิติก
ในสภาวะพักผ่อนทางสรีรวิทยาสัมพัทธ์ ไซแนปส์อยู่ในกิจกรรมไฟฟ้าชีวภาพเบื้องหลัง ความสำคัญของมันอยู่ที่ความจริงที่ว่ามันเพิ่มความพร้อมของไซแนปส์ในการดำเนินการกระตุ้นเส้นประสาท ซึ่งจะช่วยอำนวยความสะดวกอย่างมากในการส่งการกระตุ้นของเส้นประสาทผ่านไซแนปส์ ส่วนที่เหลือ 1-2 ถุงในขั้วแอกซอนอาจเข้าใกล้เยื่อ presynaptic โดยไม่ตั้งใจอันเป็นผลมาจากการที่พวกมันจะสัมผัสกับมัน ถุงน้ำแตกเมื่อสัมผัสกับเยื่อหุ้มเซลล์ก่อนไซแนปติก และสารที่อยู่ในรูปควอนตัม 1 ของ ACh จะเข้าสู่รอยแหว่งไซแนปติก ตกลงบนเยื่อแผ่นหลังไซแนปติก ซึ่ง MPN จะก่อตัวขึ้น
11. คุณสมบัติของโครงสร้างและการทำงานของกล้ามเนื้อเรียบ
กล้ามเนื้อเรียบประกอบด้วยเซลล์รูปแกนหมุน เซลล์ตั้งอยู่ในองค์ประกอบของมัดกล้ามเนื้อและอยู่ติดกันอย่างใกล้ชิด เยื่อหุ้มเซลล์ที่อยู่ติดกันก่อตัวเป็นเน็กซัสซึ่งทำหน้าที่ส่งแรงกระตุ้นจากเซลล์หนึ่งไปยังอีกเซลล์หนึ่ง เซลล์ของกล้ามเนื้อเรียบประกอบด้วย myofilaments ของแอกตินและไมโอซิน ซึ่งอยู่ในลำดับที่น้อยกว่าในเส้นใยกล้ามเนื้อโครงร่าง Sarcoplasmic reticulum ในกล้ามเนื้อเรียบมีการพัฒนาน้อยกว่าในกล้ามเนื้อโครงร่าง
พื้นฐานของปฏิกิริยาการปรับตัวของสิ่งมีชีวิตคือ ความหงุดหงิด- ความสามารถในการตอบสนองต่อผลกระทบโดยการเปลี่ยนแปลงโครงสร้างและหน้าที่ ทุกเซลล์ของสัตว์และพืชมีความหงุดหงิด ในช่วงวิวัฒนาการ ความหงุดหงิดของเนื้อเยื่อบางส่วนถึงการพัฒนาสูงสุดและถูกเปลี่ยนเป็น ความตื่นเต้นง่าย(ความสามารถในการตอบสนองต่อการระคายเคืองด้วยการกระตุ้น) เนื้อเยื่อที่กระตุ้น ได้แก่ เนื้อเยื่อประสาท กล้ามเนื้อ และสารคัดหลั่ง ประเมินความตื่นเต้นโดย เกณฑ์การระคายเคือง(ความแรงขั้นต่ำของสิ่งเร้าที่สามารถทำให้เกิดการตื่นตัวได้) สารระคายเคืองโดยธรรมชาติแบ่งออกเป็นทางกายภาพ เคมี ชีวภาพ (ไวรัส แบคทีเรีย ฯลฯ) เพียงพอและไม่เพียงพอ สิ่งเร้าที่เพียงพอเรียกว่าสิ่งเร้าเพื่อการรับรู้ซึ่งโครงสร้างทางชีววิทยาได้รับการดัดแปลงเป็นพิเศษ ดังนั้นความแรงของเกณฑ์ของสิ่งเร้าที่เพียงพอจึงมีค่าน้อยที่สุด ตัวอย่างเช่น แสงเพียงพอสำหรับเซลล์รับแสง และแรงกระตุ้นของเส้นประสาทก็เพียงพอสำหรับกล้ามเนื้อ สิ่งเร้าที่ไม่เพียงพอเรียกว่าสิ่งเร้าที่กระทำต่อโครงสร้างที่ไม่ได้ปรับให้เข้ากับการรับรู้ ตัวอย่างเช่น กล้ามเนื้อโครงร่างตอบสนองโดยการหดตัวต่อสิ่งเร้าทางไฟฟ้า
ปรากฏการณ์ไฟฟ้าชีวภาพในเนื้อเยื่อที่ตื่นเต้น การกระตุ้นเป็นชุดของกระบวนการอันเป็นผลมาจากการสลับขั้วในระยะสั้นของเยื่อหุ้มเซลล์ไซโตพลาสซึมทำให้เกิดการตอบสนองของเซลล์เฉพาะ (การนำกระแสประสาท การหดตัวของกล้ามเนื้อ ฯลฯ)
Luigi Galvani ดึงความสนใจไปที่การหดตัวของกล้ามเนื้อในการเตรียมพร้อมของขาหลังซึ่งแขวนอยู่บนตะขอทองแดงสัมผัสกับราวเหล็กของระเบียง จากการทดลองนี้ (การทดลองครั้งแรกของกัลวานี) สรุปได้ว่าการหดตัวเกิดจาก "ไฟฟ้า" ที่ส่งไปตามตะขอและราวบันไดจากไขสันหลังไปยังกล้ามเนื้อ อย่างไรก็ตาม นักฟิสิกส์ A. Volta เสนอว่าแหล่งที่มาในปัจจุบันไม่ใช่สมอง แต่เป็นศักยภาพที่จุดสัมผัสของโลหะต่างชนิดกัน ในการตอบสนองต่อสิ่งนี้ L. Galvani ใช้ตะขอแก้วเหวี่ยงเส้นประสาทไซอาติกไปที่กล้ามเนื้อของขาซึ่งทำให้กล้ามเนื้อหดตัว (การทดลองครั้งที่สองหรือการทดลองที่ไม่มีโลหะ) และพิสูจน์การมีอยู่ของ "ไฟฟ้าจากสัตว์" ต่อมาพบว่าเซลล์ที่เหลือภายในมีประจุลบเมื่อเทียบกับพื้นผิว ศักยภาพการพัก (RP) นี้อยู่ระหว่าง 30 ถึง 100 mV
ในช่วงกลางศตวรรษที่ 20 A. Hodgkin, E. Huxley และ B. Katz ได้สร้างทฤษฎีเมมเบรน-อิออน โดยอ้างอิงจาก MT เกิดจากโพแทสเซียม โซเดียม และคลอรีนไอออนที่มีความเข้มข้นต่างกันบนทั้งสองด้านของเยื่อหุ้มเซลล์ เมื่อเปรียบเทียบกับของเหลวนอกเซลล์ ไซโตพลาสซึมของเซลล์ประสาทและกล้ามเนื้อมีโพแทสเซียมไอออนมากกว่า 30-50 เท่า โซเดียมไอออนน้อยกว่า 8-10 เท่า และคลอไรด์ไอออนน้อยกว่า 50 เท่า ความสามารถในการซึมผ่านสูงของเมมเบรนสำหรับโพแทสเซียมนำไปสู่การปลดปล่อยโพแทสเซียมภายในเซลล์บางส่วนออกสู่สิ่งแวดล้อมรอบ ๆ เซลล์และทำให้เกิดประจุบวกที่ผิวด้านนอกของเมมเบรน ไอออนอินทรีย์ซึ่งเมมเบรนไม่สามารถซึมผ่านได้ จะสร้างประจุลบที่ผิวด้านในของเมมเบรน และรักษาระดับความเข้มข้นสูงของโพแทสเซียมในเซลล์และโซเดียมภายนอกปั๊มโซเดียม-โพแทสเซียม
การระคายเคืองของเซลล์ที่ถูกกระตุ้นสามารถทำให้เกิดการตอบสนองเฉพาะที่หรือศักยภาพในการดำเนินการ การตอบสนองในท้องถิ่นเกิดขึ้นกับการระคายเคืองใต้ธรณีประตู มันขึ้นอยู่กับความแรงของสิ่งเร้าโดยตรง, แปลเป็นภาษาท้องถิ่นบนพื้นผิวของเซลล์เฉพาะที่บริเวณที่เกิดการระคายเคืองและเพิ่มความตื่นเต้นง่ายของเซลล์ ศักยภาพในการดำเนินการ(PD) เกิดขึ้นภายใต้อิทธิพลของสิ่งเร้าที่เป็นขีดจำกัดหรือเหนือขีดจำกัด ในเวลาเดียวกัน ความสามารถในการซึมผ่านของเมมเบรนสำหรับโซเดียมเพิ่มขึ้น และเป็นผลจากการแทรกซึมของโซเดียมเข้าไปในเซลล์ เมมเบรนของมันถูกประจุบวกโดยคำนึงถึงสภาพแวดล้อมภายนอก จากนั้นช่องโซเดียมจะปิดและเปิดช่องโพแทสเซียมเพิ่มเติม อันเป็นผลมาจากการปลดปล่อยโพแทสเซียมออกจากเซลล์ การฟื้นฟู MP (การสลับโพลาไรเซชันของเมมเบรน) จึงเริ่มต้นขึ้น
ใน PD มี (รูปที่ 1.):
1. Prespike (การตอบสนองในท้องถิ่น) - การสลับขั้วของเมมเบรนจนถึงระดับวิกฤต
2. Spike - ประกอบด้วยส่วนขึ้น (depolarization) และจากมากไปน้อย (repolarization)
3. ศักยภาพการติดตาม - ประกอบด้วยการสลับโพลาไรเซชันการติดตามและไฮเปอร์โพลาไรเซชัน
ความตื่นเต้นง่ายในช่วง prespike เพิ่มขึ้น (ระยะของความตื่นเต้นง่ายที่เพิ่มขึ้น) และแม้แต่การกระตุ้นเพิ่มเติมที่อ่อนแอก็สามารถทำให้เกิดการก่อตัวของ AP ในช่วงระยะเวลาขัดขวาง เมมเบรนจะไม่ถูกกระตุ้น (การหักเหของแสงสัมบูรณ์) จากนั้นความตื่นเต้นง่ายจะค่อยๆ กลับคืนมา (ความหักเหสัมพัทธ์) ในเวลานี้สำหรับการกระตุ้นใหม่ จำเป็นต้องมีการกระตุ้นเหนือระดับ ด้วยการสลับโพลาไรเซชันการติดตาม ความตื่นเต้นง่ายจะเพิ่มขึ้น (ความสูงส่ง) และด้วยไฮเปอร์โพลาไรเซชัน ความตื่นเต้นจะลดลง (ความตื่นเต้นต่ำกว่าปกติ)
กฎของการระคายเคือง สะท้อนให้เห็นถึงการพึ่งพาการตอบสนองของเนื้อเยื่อที่กระตุ้นต่อความแรงของสิ่งเร้า
กฎหมายทั้งหมดหรือไม่มีเลย: สิ่งเร้าที่เป็นเกณฑ์ย่อยจะไม่ทำให้เกิดการตอบสนอง (“ไม่มีอะไร”) และสิ่งเร้าที่เป็นเกณฑ์ทำให้เกิดการตอบสนองสูงสุด (“ทุกอย่าง”) ตามกฎหมายนี้เส้นใยกล้ามเนื้อเส้นเดียวและหัวใจหดตัว
กฎแห่งความแข็งแกร่ง : ยิ่งระคายเคืองรุนแรงยิ่งตอบสนอง ตามกฎหมายนี้ การทำงานของกล้ามเนื้อโครงร่าง ประกอบด้วยเส้นใยกล้ามเนื้อที่มีความตื่นตัวต่างกัน เส้นใยที่กระตุ้นได้ดีที่สุดจะตอบสนองต่อสิ่งเร้าที่เป็นเกณฑ์ การเพิ่มความแข็งแรงของสิ่งเร้ายังเกี่ยวข้องกับเส้นใยที่มีความตื่นเต้นในการตอบสนองน้อยลง และความกว้างของการหดตัวของกล้ามเนื้อจะเพิ่มขึ้น
กฎการกระตุ้นของ Dubois-Reymond : การกระทำของไฟฟ้ากระแสตรงขึ้นอยู่กับความแรงและอัตราการเพิ่มขึ้น ด้วยการเพิ่มขึ้นอย่างช้าๆ เนื้อเยื่อจะปรับให้เข้ากับสิ่งกระตุ้น (ที่พัก) และการกระตุ้นอาจไม่เกิดขึ้น
กฎแห่งแรง-เวลา สะท้อนให้เห็นถึงการพึ่งพาค่าเกณฑ์ของกระแสตรงในเวลาที่ดำเนินการ ยิ่งพัลส์ปัจจุบันสั้นลงเท่าใด ความน่ารำคาญก็จะยิ่งน้อยลงเท่านั้น เรียกว่าปริมาณกระแสต่ำสุดที่สามารถกระตุ้นด้วยระยะเวลาไม่ จำกัด ของการกระทำ รีโอเบส. ช่วงเวลาที่กระแสเท่ากับรีโอเบสทำให้เกิดการกระตุ้นเรียกว่า ช่วงเวลาที่ดี. โครนาเซีย- เวลาต่ำสุดที่กระแสเท่ากับสองรีโอเบสทำให้เกิดปฏิกิริยา
กฎว่าด้วยไฟฟ้ากระแสตรง : เมื่อกระแสปิด การกระตุ้นจะเกิดขึ้นใต้แคโทด และเมื่อกระแสเปิด ใต้แอโนด
กฎของอิเล็กโทรโทนทางสรีรวิทยา : ในพื้นที่ของแคโทด ความตื่นเต้นง่ายเพิ่มขึ้น (kathelektroton) และที่ขั้วบวกจะลดลง (anelectroton) ด้วยการกระทำของกระแสตรงเป็นเวลานาน ความตื่นเต้นง่ายภายใต้แคโทดจะลดลง (ภาวะซึมเศร้าของแคโทด) และภายใต้ขั้วบวกจะเพิ่มขึ้น (ความสูงส่งของขั้วบวก)
ใยประสาท มีคุณสมบัติ: ความตื่นเต้นง่าย การนำไฟฟ้า และความสามารถ. การกระตุ้นจะแพร่กระจายไปตามเส้นใยประสาทก็ต่อเมื่อมัน ความสมบูรณ์ทางกายวิภาคและสรีรวิทยาไม่ผ่านไปยังใยประสาทข้างเคียง (กฎแห่งการกระทำที่แยกตัวออกมา) ไม่เปลี่ยนแปลงแอมพลิจูด ( กฎของการนำไฟฟ้าที่ไม่ลดทอนหรือไม่ลดลง) และดำเนินการทั้งสองทิศทางจากสถานที่เกิดการระคายเคือง ( กฎหมายการถือครองทวิภาคี).
พื้นที่ตื่นเต้นของพื้นผิวของเส้นใยประสาทจะได้รับประจุลบ เนื่องจากบริเวณที่ไม่มีการกระตุ้นที่อยู่ติดกันมีประจุบวก กระแสไฟฟ้าจะไหลระหว่างกัน สิ่งนี้จะทำให้พื้นที่พักผ่อนตื่นเต้นและเปลี่ยนประจุด้วย ในท้ายที่สุด การกระตุ้นจะกระจายไปทั่วพื้นผิวทั้งหมดของเส้นใยประสาทที่ไม่ใช่ไมอีลิน (ไม่มีไมอีลิน) (รูปที่ 2ก.) ในกระบวนการเยื่อไมอีลิน (เยื่อกระดาษ) ของเซลล์ประสาท การกระตุ้นสามารถเกิดขึ้นได้ที่โหนดของแรนเวียร์เท่านั้น ดังนั้นมันจึงแพร่กระจายโดยการกระโดดจากจุดสกัดหนึ่งไปยังอีกจุดหนึ่ง (รูปที่ 2b.) และเคลื่อนที่ได้เร็วกว่าเส้นใยที่ไม่มีเนื้อมาก
ตามเส้นผ่านศูนย์กลางและความเร็วของการนำการกระตุ้นใยประสาทแบ่งออกเป็นประเภท A, B และ C เส้นใยที่หนาที่สุดของประเภท A (เส้นผ่านศูนย์กลาง 12-22 ไมครอน) ด้วยความเร็วสูงสุด (70-120 m / s) การกระตุ้นจากสมองไปยังกล้ามเนื้อโครงร่างและจากกล้ามเนื้อตัวรับไปยังสมอง จากตัวรับอื่น ๆ เส้นใยชนิด A มาพร้อมกับเส้นผ่านศูนย์กลางที่เล็กกว่าเล็กน้อย (จาก 8 ถึง 1 ไมครอน) และความเร็วในการนำไฟฟ้ากระตุ้น (5-70 m / s) เส้นใยประเภท B รวมถึงเส้นใยพืช preganglionic (เส้นผ่านศูนย์กลาง - 1-3.5 ไมครอน, ความเร็วในการกระตุ้น - 3-18 m / s) เฉพาะเส้นใยประเภท C เท่านั้นที่เป็นอะไมโคติก (เส้นผ่านศูนย์กลาง 0.5–2 ไมโครเมตร ความเร็วกระตุ้นน้อยกว่า 3 เมตร/วินาที) พวกมันเป็นใยใยประสาทหลังปมประสาท และยังมาจากตัวรับความเจ็บปวด ซึ่งเป็นส่วนหนึ่งของตัวรับความร้อนและตัวรับแรงกด
มีเส้นใยประสาท lability (ความคล่องตัวในการทำงาน). วัดจากจำนวนการกระตุ้นสูงสุดที่เส้นใยประสาทสามารถทำซ้ำได้ ในเส้นใยประสาท ความสามารถในการรับรู้จะสูงกว่า (สูงถึง 1,000 Hz) มากกว่าในโครงสร้างที่กระตุ้นได้อื่นๆ หากเส้นประสาทได้รับความเสียหาย (จากสารเคมี ความร้อน ความเย็น หรือกระแสไฟฟ้า) โดยไม่ละเมิดความสมบูรณ์ทางกายวิภาค ก็จะเกิดสภาวะขึ้นในนั้น พาราไบโอซิส . ในกรณีนี้ ขั้นตอนการปรับระดับ ความขัดแย้ง และการยับยั้งจะถูกแทนที่อย่างต่อเนื่อง ใน ปรับระดับเฟส - ด้วยความระคายเคืองที่หายาก แรงกระตุ้นทั้งหมดจะถูกส่งผ่านบริเวณที่เสียหาย และเพียงบางส่วนเท่านั้นที่มีจังหวะสูง ใน ขัดแย้ง -การตอบสนองต่อการระคายเคืองบ่อยครั้งน้อยกว่าการระคายเคืองที่หายาก ใน เบรค- เส้นประสาทไม่กระตุ้นใด ๆ เมื่อออกจากพาราไบโอซิส ระยะเดียวกันจะถูกสังเกต แต่ในลำดับที่กลับกัน
ทั้งหมด กล้ามเนื้อ มีความตื่นเต้นง่าย (ความสามารถในการตื่นเต้นจากการกระทำของสิ่งเร้า) การนำไฟฟ้า (ความสามารถในการกระตุ้น) และการหดตัว (ความสามารถในการเปลี่ยนความยาวหรือความตึงเครียดเมื่อตื่นเต้น) สำหรับ หัวใจและส่วนของเส้นใยกล้ามเนื้อเรียบนอกเหนือจากคุณสมบัติที่ระบุไว้แล้ว ความเป็นอัตโนมัติ (ความสามารถในการกระตุ้นโดยธรรมชาติ) เป็นลักษณะเฉพาะ คุณสมบัติเฉพาะของกล้ามเนื้อเรียบทั้งหมดคือ พลาสติก(ความสามารถในการรักษาความยาวที่กำหนดให้ได้นาน).
ความแข็งแรงของกล้ามเนื้อถูกกำหนดโดยน้ำหนักบรรทุกสูงสุดที่สามารถยกได้ และ งาน- ผลคูณของน้ำหนักที่ยกขึ้นตามความสูงของลิฟต์ ทำงานสูงสุดที่โหลดปานกลาง ที่ การหดตัวของไอโซโทนิกกล้ามเนื้อเปลี่ยนความยาวและความตึงจะคงที่ (นี่คือวิธีที่กล้ามเนื้อหดตัวโดยไม่มีแรงต้านต่อการเปลี่ยนแปลงความยาว) ที่ การหดตัวแบบสามมิติความยาวของกล้ามเนื้อจะคงที่และความตึงเครียดจะเพิ่มขึ้น (เช่น เมื่อพยายามยกของที่มากเกินไป) ตามสภาพธรรมชาติก็มี ตัดแบบผสม(ทั้งความยาวและความตึงของกล้ามเนื้อเปลี่ยนไป)
สิ่งกระตุ้นอย่างหนึ่งทำให้เกิด การหดตัวของกล้ามเนื้อเดี่ยว. มันไฮไลท์: ระยะเวลาแฝง(เวลาตั้งแต่เริ่มกระตุ้นจนถึงเริ่มตอบสนอง) เฟสสั้นลงและ ระยะผ่อนคลาย. หากสิ่งกระตุ้นที่ตามมาแต่ละครั้งมาถึงกล้ามเนื้อโครงร่างในช่วงเวลาที่สั้นลง บาดทะยักเรียบและอยู่ในช่วงผ่อนคลาย - บาดทะยัก. ภายใต้สภาวะตามธรรมชาติ กล้ามเนื้อโครงร่างได้รับแรงกระตุ้นหลายอย่าง ซึ่งกล้ามเนื้อตอบสนองด้วยบาดทะยักเรียบ แอมพลิจูดของมันมักจะสูงกว่าแอมพลิจูดของการหดตัวเพียงครั้งเดียว ไม่. Vvedensky อธิบายสิ่งนี้ด้วยข้อดีและข้อเสีย เหมาะสมที่สุด- ความถี่ที่เกิดการระคายเคืองในช่วงของความตื่นเต้นง่ายที่เพิ่มขึ้น (สูงสุดบาดทะยัก) ในแง่ร้าย- ความถี่ที่การระคายเคืองใหม่ถูกนำมาใช้กับระยะของความตื่นเต้นง่ายที่ลดลง (บาดทะยัก - น้อยที่สุด)
เมื่อศึกษากล้ามเนื้อโครงร่างและกล้ามเนื้อหัวใจในแสงโพลาไรซ์ จะมองเห็นโซนสลับที่มีความหนาแน่นของแสงต่างกัน (รูปที่ 3) สิ่งนี้ทำให้สามารถแบ่งองค์ประกอบที่หดตัวของเส้นใยกล้ามเนื้อโครงร่าง (myofibrils) ออกเป็นหน่วยการทำงาน - ซาร์โคเมเรส(พื้นที่ระหว่างเมมเบรน Z ที่อยู่ติดกัน)

ข้าว. 3.ไมโครกราฟของส่วนของกล้ามเนื้อโครงร่าง
ความหนาแน่นเชิงแสงที่มีลักษณะเฉพาะของบริเวณซาร์โคเมียร์นั้นเกิดจากตำแหน่งของโปรตีนที่หดตัว (แอกตินและไมโอซิน) ในพวกมัน (รูปที่ 4)

ข้าว. 4.เค้าโครงของโปรตีนที่หดตัวในซาร์โคเมียร์ (ในโซน H เส้นใยไมโอซินไม่มีสะพานเชื่อม)
ใกล้ Z-membranes มีแอกติน (บริเวณแสง - ไอโซโทรปิกหรือ ฉัน-ดิสก์). ใกล้กับตรงกลางของซาร์โคเมียร์ มืด ( แอนไอโซโทรปิก) A-disc ที่มี myosin และ actin M-line ผ่านใจกลางของซาร์โคเมียร์ ด้านใดด้านหนึ่งคือ myosin (การตรัสรู้หรือ H-zone) ด้านหนึ่งแอกตินติดอยู่กับเมมเบรน Z และอีกด้านหนึ่งเป็นอิสระและสิ้นสุดระหว่างเส้นใย myosin ในบริเวณ A-disk (ที่ชายแดนกับโซน H) ปลายทั้งสองของเส้นใยไมโอซินเป็นอิสระ
เมื่อหดตัว ความกว้างของดิสก์ไอโซทรอปิกจะลดลง ในกรณีนี้ ใยไมโอซินสามารถเข้าถึงแผ่น Z ได้เมื่อสิ้นสุด และความยาวของซาร์โคเมียร์สามารถสั้นลงได้ 30-50%
กลไกการหดตัวของกล้ามเนื้อ เส้นใย myosin เกือบทั้งหมดมีสะพานด้านข้าง (ขาดเฉพาะใกล้กับ M-line) หลังจากจับกับแอกตินแล้ว พวกมันจะเปลี่ยนมุมเอียง (โดยใช้พลังงานของ ATP) ซึ่งเคลื่อนโปรตีนที่หดตัวสัมพันธ์กัน (ซาร์โคเมียร์สั้นลง) จากนั้นแอกตินจะเชื่อมต่อกับสะพานไมโอซินอีกอันหนึ่งและมีความคืบหน้าต่อไป
ส่วนที่เหลือ การจับตัวกันของแอกตินกับไมโอซินจะถูกขัดขวางโดยโปรตีนโทรโปนินและโทรโปไมโอซิน เมื่อตื่นเต้น พวกมันจะถูก "ทำให้เป็นกลาง" โดยแคลเซียมที่เข้าไปในซาร์โคพลาสซึม (ไซโตพลาสซึมของเซลล์กล้ามเนื้อ) และปฏิสัมพันธ์ของโปรตีนที่หดตัวจะเริ่มขึ้น การหยุดชะงักของการกระตุ้นนำไปสู่การกำจัดแคลเซียมออกจาก sarcoplasm, troponin และ tropomyosin ทำลายคอมเพล็กซ์ของ actin และ myosin - กล้ามเนื้อคลายตัว
กล้ามเนื้อโครงร่างใช้แคลเซียมจากถังภายในเซลล์ของ reticulum sarcoplasmic กล้ามเนื้อเรียบได้รับแคลเซียมจากช่องว่างระหว่างเซลล์เท่านั้น ในขณะที่กล้ามเนื้อหัวใจใช้ทั้งสองแหล่งของไอออนนี้ การใช้แหล่งแคลเซียมภายในเซลล์เพียงอย่างเดียวช่วยให้กล้ามเนื้อโครงร่างหดตัวและคลายตัวด้วยความเร็วสูงสุด และกล้ามเนื้อเรียบจะค่อยๆ เปลี่ยนสี
ไซแนปส์ - การเชื่อมต่อการทำงานระหว่างเซลล์ประสาทกับเซลล์อื่นๆ มีไซแนปส์ไฟฟ้าและเคมี ประสาทไฟฟ้าความต้านทานไฟฟ้าต่ำในลักษณะเฉพาะในพื้นที่สัมผัสของเซลล์และ AP จะถูกถ่ายโอนไปยังเมมเบรนที่อยู่ติดกันอย่างรวดเร็ว ประสาทเคมีประกอบด้วย presynaptic membrane, postsynaptic membrane และ synaptic cleft (รูปที่ 5.) ภายใต้อิทธิพลของแรงกระตุ้นของเส้นประสาทใน synapses สารเคมีสารเคมีจะถูกปล่อยออกมา - ผู้ไกล่เกลี่ย (เช่น acetylcholine, norepinephrine) จากแอกซอนที่หนาขึ้นของ presynaptic เข้าไปในรอยแยก synaptic และการมีปฏิสัมพันธ์กับตัวรับบนเมมเบรน postsynaptic ในซิงก์กระตุ้น (excitatory synapses) สิ่งนี้นำไปสู่การเกิดขึ้นของ excitatory (EPSP) และในการยับยั้ง - ยับยั้ง (IPSP) ศักยภาพหลังซินแน็ปติก หลังจากที่สื่อกลางทำหน้าที่ของมันแล้วเซลล์จะถูกทำลายหรือดูดซึม
|
|
|
ข้าว. 5.แผนผังโครงสร้างของไซแนปส์เคมี |

นอกระบบประสาทส่วนกลาง สารสื่อประสาทที่พบมากที่สุดคืออะเซทิลโคลีน มันอำนวยความสะดวกในการกระตุ้นผ่านปมประสาทพืช เพิ่มการหลั่งของอะดรีนาลีนจากต่อมหมวกไตและกรดไฮโดรคลอริกจากต่อมในกระเพาะอาหาร กดการทำงานของหัวใจ และทำให้กล้ามเนื้อเรียบของอวัยวะภายในและต่อมไร้ท่อบางส่วนหดตัว ในกล้ามเนื้อเรียบของหลอดลม, ลำไส้, กระเพาะปัสสาวะ, มดลูก, กล้ามเนื้อวงกลมและปรับเลนส์ของดวงตา, acetylcholine นำไปสู่หลอดลม, เพิ่มการบีบตัวของลำไส้และกระเพาะอาหาร (ด้วยการผ่อนคลายของกล้ามเนื้อหูรูด), การหดตัวของกระเพาะปัสสาวะและ การหดตัวของรูม่านตา
ตามสถานที่ประสาทถูกแบ่งออกเป็นส่วนปลาย (ประสาทและกล้ามเนื้อ, ตัวรับ - เซลล์ประสาท, ฯลฯ ); ส่วนกลาง (axosomatic, axodendritic ฯลฯ ); โดยเครื่องหมายการกระทำ(กระตุ้นและยับยั้ง) และ โดยคนกลางที่เลือก(cholinergic, adrenergic เป็นต้น)
ประสาทเคมีกระตุ้นการกระตุ้นในทิศทางเดียว ส่งแรงกระตุ้นได้ช้ากว่าไปตามใยประสาท (synaptic delay); มีความอ่อนล้าและไวต่อสารเคมีสูง
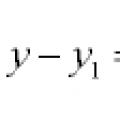 สมการเส้นและเส้นโค้งในระนาบ
สมการเส้นและเส้นโค้งในระนาบ สมการเอกพันธ์ทั่วไป สมการเอกพันธ์ทั่วไปของอันดับสอง
สมการเอกพันธ์ทั่วไป สมการเอกพันธ์ทั่วไปของอันดับสอง ขอบคุณเครื่องหมายจุลภาคเนื่องจากข้อเท็จจริงที่ว่าในพวกเขา
ขอบคุณเครื่องหมายจุลภาคเนื่องจากข้อเท็จจริงที่ว่าในพวกเขา