โครงสร้าง การจำแนก และคุณสมบัติเชิงหน้าที่ของไซแนปส์ ลักษณะทางสัณฐานวิทยาและการทำงานของไซแนปส์ไฟฟ้าและเคมี
เชอร์ริงตันในปี 1897
แปลจากภาษากรีกแปลว่า - ปิด
ไซแนปส์คือ
โครงสร้าง,
ซึ่งผ่าน
มั่นใจ
การส่งข้อมูล
ระหว่างประหม่า
เซลล์ประสาทและ
มีกล้ามเนื้อ
เซลล์.
3 การจำแนกประเภทของไซแนปส์
1. ตามสถานที่:ก.) ส่วนกลาง (สมองและไขสันหลัง)
- axosomatic, axoaxonal, axodendretic;
- เดนโดรโซมาติก, เดนโดรเดนเดรติก
b.) อุปกรณ์ต่อพ่วง (ประสาทและกล้ามเนื้อ, ระบบประสาท).
2. โดยธรรมชาติของการกระทำ:
ก.) น่าตื่นเต้น
ข.) เบรค
3.) ตามวิธีการส่งสัญญาณ:
ก.) ไฟฟ้า;
ข.) สารเคมี;
ค.) ผสม.
4.) โดยการพัฒนาในการเกิด:
ก.) เสถียร (ซินเซสของส่วนโค้งของรีเฟล็กซ์ที่ไม่มีเงื่อนไข);
b.) พลวัต (ปรากฏในกระบวนการพัฒนาของแต่ละบุคคล).
4 การแปลประเภทต่างๆของ synapses
6 ไซแนปส์
เคมีไฟฟ้า
6
7 โครงสร้างของไซแนปส์เคมี:
1. พรีไซแนปติกเมมเบรน;
2. โพสต์ซินแนปติก
เมมเบรน;
3. ไซแนปติกแหว่ง
หลักการเดล:
เซลล์ประสาทหนึ่งตัวเปล่งออกมา
คนกลางคนหนึ่ง
ตอนนี้
แก้ไข
8 โครงสร้างของไซแนปส์เคมี
พรีไซแนปติกพังผืด
เกิดจากแอกซอน
สิ้นสุดซึ่งสูญเสีย myelin sheath ในที่นี้
มันมีถุงซินแนปติกที่มีเส้นผ่านศูนย์กลาง 30-50 นาโนเมตรและ
ไมโตคอนเดรียจำนวนมาก Synaptic vesicles ประกอบด้วย
ผู้ไกล่เกลี่ยและ ATP (ประกอบด้วยควอนตัมของผู้ไกล่เกลี่ย) มี
ประจุลบและ
ขับไล่โดย presynaptic
เยื่อหุ้มถุงจะกระจุกตัวอยู่ใน "โซนแอคทีฟ"
แต่ละตุ่มประกอบด้วยโมเลกุลตัวกลางหลายพันตัว (ตัวอย่างเช่น
acetylcholine) และโมเลกุล ATP.
Synaptic vesicles มีหลายส่วน -
สระสำรองและหมุนเวียน
ได้รับการจัดสรรเป็นส่วน ๆ -
ควอนตัม
ความกว้างของรอยแยก synaptic คือ 20-50 นาโนเมตร เธอ
เต็มไปด้วยของเหลวระหว่างเซลล์และมีโครงสร้าง
องค์ประกอบ: เมมเบรนชั้นใต้ดินประกอบด้วยเส้นใยไฟเบอร์
ที่เชื่อมระหว่างก่อนและหลังซินแน็ปทิก
เยื่อ นอกจากนี้ยังมีเอนไซม์ที่สลายโมเลกุล
คนกลาง
9
เมมเบรนโพสซินแนปติก (หรือแผ่นท้าย) มีมากมาย
เท่า,
เพิ่มขึ้น
สี่เหลี่ยม
ของเธอ
ปฏิสัมพันธ์กับคนกลาง ไม่มีแรงดันไฟฟ้าขึ้นอยู่กับเมมเบรน
ช่องไอออน แต่ความหนาแน่นของช่องรับ-gated สูง (การเลือกไอออนต่ำ)
จำนวนตัวรับบนพื้นผิวของเมมเบรนโพสซินแนปติก
อาจแตกต่างกันไป ดังนั้นด้วยการจัดสรรขนาดใหญ่
ปริมาณของผู้ไกล่เกลี่ย - การลดความไวของตัวรับเกิดขึ้น ใน
โดยเฉพาะจำนวนตัวรับบน
เมมเบรนโพสต์ซินแนปติก (กำจัดตัวรับ)
ยกเว้น
สิ่งนี้จะลดความไวต่อผู้ไกล่เกลี่ย
ตรงกันข้าม ในระหว่างการประณาม เมื่อผู้ไกล่เกลี่ยปล่อยตัวอย่างรุนแรง
ลดลง จำนวนตัวรับสามารถเพิ่มขึ้นอย่างรวดเร็ว
ดังนั้น ไซแนปส์จึงเป็นโครงสร้างที่มีไดนามิกสูง
ซึ่งเป็นตัวกำหนดความเป็นพลาสติกของมัน
10. 10 ไซแนปส์พลาสติก
การเปลี่ยนแปลงเกิดขึ้นในทุกระดับ มันคือการเปลี่ยนแปลงจำนวนตัวรับสารสื่อประสาทในโพสต์ไซแนปส์
การเปลี่ยนแปลง
วี
ของพวกเขา
การทำงาน
สามารถ
และ
การแก้ไขหลังการแปล
การศึกษาที่ดีที่สุดคือฟอสโฟรีเลชั่น
นี่เป็นกระบวนการของการเปลี่ยนแปลงอย่างรวดเร็วในโครงสร้างของตัวรับ
ซึ่งเอ็นไซม์ที่เรียกว่าไคเนส
แนบสารตกค้างของกรดฟอสฟอริกเข้ากับหนึ่งในนั้น
กรดอะมิโนในสายโพลีเปปไทด์ของตัวรับ มันนำไปสู่การ
ต่อการเปลี่ยนแปลงอย่างมากในโครงสร้างตัวรับและ
อาจส่งผลต่อประสิทธิภาพการทำงานอย่างจริงจัง
ยกเว้น
ไป,
ฟอสโฟรีเลชั่น
ถูกเปิดเผย
เป้าหมายระดับโมเลกุลอื่น ๆ อีกมากมายที่พบใน
โพสต์ไซแนปส์ มีการเปลี่ยนแปลงในโครงร่างโครงร่างการสังเคราะห์
โปรตีนเพิ่มเติมทั้งโดยทั่วไปในเซลล์และภายใน
กระดูกสันหลัง.
11. 11 องค์ประกอบของประสาทและกล้ามเนื้อไซแนปส์
12.
12สุดยอด
โครงสร้าง
ประหม่า-
มีกล้ามเนื้อ
ไทย
ไซแนปส์
13. การปลดปล่อยคนกลางในไซแนปส์เกิดขึ้นเป็นส่วนๆ (ควอนตา) ควอนตัมเครื่องส่งสัญญาณตั้งอยู่ใน synaptic vesicle และถูกปล่อยออกมาจาก
13 ทฤษฎีควอนตัม-vesicular.การปลดปล่อยคนกลางในไซแนปส์เกิดขึ้นเป็นส่วนๆ
(ควอนตัม).
ควอนตัมเครื่องส่งสัญญาณตั้งอยู่ในถุงซินแนปติกและ
ออกจากปลายประสาทโดยเอกโซไซโทซิส
ในปี 1954 เดล กัสติโยและแคตซ์
อธิบายรายละเอียดเกี่ยวกับ PEP และ MEPP
ที่จุดเชื่อมต่อประสาทและกล้ามเนื้อ
พวกเขาแนะนำว่าคนกลาง
ปลดปล่อย
แน่ใจ
ส่วน - ควอนตัม
ในปี พ.ศ. 2498 ภาษาบาลี
พาลา,
เดอ
Robertis และ Bennett ค้นพบ
ไซแนปติก
ถุง
กับ
โดยใช้
อิเล็กทรอนิกส์
กล้องจุลทรรศน์.
14. 14 ศักย์จานท้าย
มีศักยภาพในการโพสต์ซินแน็ปติคกระตุ้น (EPSP)เฉพาะที่บนเมมเบรนโพสซินแนปติก ค่าของมัน
กำหนดโดยจำนวนของควอนตัมสื่อกลางที่ปล่อยออกมา ในการเชื่อมต่อกับ
นี้:
1) EPSP ซึ่งแตกต่างจาก PD คือไม่อยู่ภายใต้กฎหมาย "All or Nothing" แต่
ปฏิบัติตามกฎการรวม:
ยิ่งมีการเผยแพร่ตัวกลางมากเท่าไหร่ มูลค่าของ EPSP ก็จะยิ่งมากขึ้นเท่านั้น
2) ข้อแตกต่างประการที่สองระหว่าง EPSP และ PD คืออิเล็กโทรโทนิก
การกระจายเช่น การสลายตัวที่อาจเกิดขึ้นกับระยะห่างจากขั้ว
จาน
ไม่ตื่นเต้น - บนจานท้ายจะถูกบันทึกไว้
จิ๋ว
ศักยภาพ
เทอร์มินัล
บันทึก
(เอ็ม.พี.เค.พี),
ซึ่งเป็นคลื่นของการดีโพลาไรเซชันขนาดเล็ก 0.5
เอ็มวี ต้นกำเนิดของพวกเขาเกี่ยวข้องกับการปลดปล่อยควอนตัมที่เกิดขึ้นเอง
คนกลาง
จาก
พรีไซแนปติก
เยื่อ,
เนื่องจาก
การยึดเกาะโดยธรรมชาติของถุงซินแนปติกกับเยื่อหุ้มเซลล์ (~1
ควอนตัมต่อวินาที)
สำหรับการเกิดขึ้นของ EPSP การเปิดตัวพร้อมกันของ
ควอนตัมคนกลางหลายร้อยคน
15. 15
16. 16
ศักยภาพและกระแสขั้ว
บันทึกเมื่อ
แตกต่าง
ระยะทางจาก
ของเธอ
17. 17
หากไซแนปส์ถูกกระตุ้นก็จะเพิ่มขึ้นการซึมผ่านของเมมเบรนโพสซินแนปติก
โซเดียมและโพแทสเซียม EPSP เกิดขึ้น เขามีอยู่
เฉพาะที่บนเมมเบรนโพสซินแนปติกเท่านั้น แต่
ถ้าขนาดของโพลาไรเซชันแบบโพสต์ซินแนปติก
เมมเบรนถึงระดับวิกฤตแล้ว EPSP
แปลงเป็นศักยภาพในการดำเนินการ
เซลล์ที่ออกมา
หากไซแนปส์ถูกยับยั้ง สารสื่อประสาทที่ถูกปล่อยออกมา
เพิ่มการซึมผ่านของโพสต์ซินแนปติก
เมมเบรนสำหรับโพแทสเซียมและคลอรีน กำลังพัฒนา
ไฮเปอร์โพลาไรเซชัน (TPSP) ขยายไปถึง
เยื่อหุ้มเซลล์ออกจากกันเพิ่มขีด จำกัด
ความตื่นเต้นและลดความตื่นเต้นง่าย
18. 18 ศักยภาพหลังการซิงโครไนซ์
19. 19 กลไกการเปลี่ยน vPKP/EPSP เป็นเซลล์ PD
19กลไกของการเปลี่ยนแปลง
VPKP/EPSP ในเซลล์ PD
หลังจากเริ่มมีอาการของ VPKD ระหว่างการสลับขั้ว
เมมเบรนแผ่นท้ายและที่เหลือ
ส่วนหนึ่งของเยื่อหุ้มเซลล์ที่กระตุ้นด้วยไฟฟ้าของเส้นใยกล้ามเนื้อ
ติดกับแผ่นท้าย - มีในท้องถิ่น
ปัจจุบัน. กระแสนี้เกิดจากการแจกจ่าย Na+ ไอออน
เข้ามา
ผ่าน
เคมี
ช่อง
- ระหว่าง
จานท้ายและซาร์โคเลมมา
หากขนาดของกระแสท้องถิ่นอนุญาตให้มีการสลับขั้ว
เยื่อใยกล้ามเนื้อ
ก่อน
จากนั้นเอกก็เปิดขึ้น
ช่อง Ca 2+ ที่ขึ้นกับแรงดันไฟฟ้าของ sarcolemma, อินพุต
แคลเซียมไอออนเสร็จสิ้นการสลับขั้ว - PD เกิดขึ้น
ซึ่งจะกระจายไปตามเส้นใยกล้ามเนื้อ
ดังนั้น
ทาง
วี.พี.เค.พี
ผลพลอยได้
(หรือ
ถูกเปลี่ยน) เป็น PD ของเส้นใยกล้ามเนื้อ
20. 20 ไซแนปส์ประสาทและกล้ามเนื้อ
21. 21 ตำแหน่งของช่องที่ควบคุมโดยตัวรับและช่องแรงดันไฟฟ้าบนเยื่อหุ้มเซลล์ของกล้ามเนื้อ
Ca ขึ้นอยู่กับศักยภาพช่อง
PP= -80 มิลลิโวลต์
โพสต์ซินแนปติก
พังผืด
-80 มิลลิโวลต์
ตัวรับขับเคลื่อน
ช่อง
Ca ขึ้นอยู่กับศักยภาพ
ช่อง
PP= -80mV
22.
22การส่งสัญญาณกระตุ้นประสาท
- ไซแนปส์ของกล้ามเนื้อ
ทางแยกประสาทและกล้ามเนื้อ
สิ้นสุด presynaptic
เมมเบรนโพสต์ซินแน็ปติก
การผันคำกริยาด้วยไฟฟ้า
ปล่อยอะเซทิลโคลีน
อะเซทิลโคลีนเอสเทอเรส
H - ตัวรับ ACh
สพป
PD ของซาร์โคเลมมา
การลดน้อยลง
กล้ามเนื้อ
23. 23 เมแทบอลิซึมของผู้ไกล่เกลี่ย: อช
24. 24 เมแทบอลิซึมของผู้ไกล่เกลี่ย: เปิด
25. 25 ตามผลกระทบที่กระทำโดยผู้ไกล่เกลี่ยต่อเมมเบรนโพสซินแนปติก เคมีซินเซสแบ่งออกเป็น:
1. ไอโอโนทรอปิก2. เมตาบอโทรปิก
26. 26 การส่งผ่านของการกระตุ้นในไซแนปส์เคมี
1. โมเลกุลสารสื่อประสาทเข้าสู่เมมเบรน
ถุงซินแนปติก,
ตั้งอยู่ที่
ขั้วพรีไซแนปติก
และมุ่งเน้นที่
โซนที่ใช้งานอยู่
เยื่อหุ้มเซลล์ก่อนไซแนปติก
2. AP มาพร้อมแอกซอน
สลับขั้ว
เยื่อหุ้มเซลล์ก่อนไซแนปติก
3. เนื่องจากการสลับขั้ว
เปิด
ขึ้นอยู่กับแรงดันไฟฟ้า
ช่อง Ca2+ และ Ca2+
เข้าสู่เทอร์มินัล
4. เพิ่มขึ้นในเซลล์
[Ca2+] กระตุ้นการหลอมรวม
ถุงซินแนปติกด้วย
เยื่อหุ้มเซลล์ก่อนไซแนปติก
และการปล่อยสารสื่อประสาท
แหว่ง synaptic
(เอกโซไซโทซิส).
27. 27 การส่งผ่านของการกระตุ้นในไซแนปส์เคมี
5. สารสื่อประสาทควอนตัมเข้าสู่ synaptic
ช่องว่าง, กระจายอยู่ในนั้น.
ส่วนหนึ่งของโมเลกุลสารสื่อประสาท
เกี่ยวข้องกับเฉพาะ
ตัวรับสำหรับพวกเขา
เมมเบรนโพสต์ซินแน็ปติก
6. สารสื่อประสาทถูกผูกไว้
ตัวรับถูกเปิดใช้งาน
นำไปสู่การเปลี่ยนแปลง
โพลาไรซ์
เมมเบรนโพสต์ซินแน็ปติก
หรือโดยตรง (การจัดหาไอออน
ผ่านตัวรับไอโอโนโทรปิก)
หรือทางอ้อม
การเปิดใช้งานช่องไอออน
ผ่านระบบจีโปรตีน
(ตัวรับเมตาบอโทรปิก).
7. การยับยั้งสารสื่อประสาท
เกิดขึ้นได้ทั้งทาง
การย่อยสลายด้วยเอนไซม์หรือ
โมเลกุลของสารสื่อประสาท
นำมาขึ้นโดยเซลล์
28. 28 ไอโอโนโทรปิกไซแนปส์
28ไอโอโนโทรปิก
ไซแนปส์
29. 29 ไซแนปส์ Metabotropic
30. 30 ตัวรับโพสต์ซินแน็ปติก
ไอโอโนโทรปิก1. รวดเร็ว
2. คอมเพล็กซ์เดียวกับ
ช่องไอออน
3. ทำงานเพื่อ
ช่องเปิด
4. นิโคติน
ตัวรับ cholinergic,
ตัวรับกาบา,
ไกลซีน
เมตาบอโทรปิก
1. ช้า
2. การเปิดใช้งาน
น้ำตกเอนไซม์
3. ต่อมาอาจ
เปิดหรือ
ปิด
(ทางอ้อม) ช่องทาง
4. มัสคารินิก
ตัวรับ cholinergic,
ตัวรับ
ส่วนใหญ่
นิวโรเปปไทด์,
ส่วนใหญ่
ตัวรับ
catecholamines และ
เซโรโทนิน
31. 31
32. 32
คุณสมบัติทางสรีรวิทยาไซแนปส์ทางเคมี:
- การนำทางเดียว
- ความล่าช้าของซินแนปติก
- ลักษณะควอนตัมของการปลดปล่อยผู้ไกล่เกลี่ย
- การลดลงของสารสื่อประสาทด้วยการกระตุ้นเป็นเวลานาน
(ความเมื่อยล้าไซแนปส์)
- ความสามารถในการสร้างไซแนปส์น้อยกว่าเส้นประสาท
- การเปลี่ยนแปลงของจังหวะของการกระตุ้น
- มีความไวสูงต่อการขาด O2 และสารพิษ
33. 33 การจำแนกประเภทของตัวบล็อกประสาทและกล้ามเนื้อ
33 การจำแนกประเภทของตัวบล็อกประสาทและกล้ามเนื้อ1.) ยาชาเฉพาะที่ปิดกั้นการนำของการกระตุ้น
เยื่อ presynaptic (โนโวเคน, ลิโดเคน ฯลฯ )
2.) ตัวบล็อกที่ป้องกันการปล่อยตัวคนกลาง
จากปลาย presynaptic (สารพิษโบทูลินั่ม, Mn,
พรอสตาแกลนดิน)
3.)
บล็อกเกอร์,
ละเมิด
กลับ
การจับกุม
พรีไซแนปติก
พังผืด
สินค้า
ไฮโดรไลซิส
คนกลาง (โคลีน),
จึงป้องกันการสังเคราะห์ซ้ำ
(ฮีโมโคลีน).
4.)
บล็อคเกอร์
ตัวรับ ACh
บน
โพสต์ซินแนปติก
เมมเบรน:
ก.) การแข่งขัน - ทูโบคูรารีน
b.) การกระทำที่ไม่ใช่การแข่งขัน - prestonal, α-bungarotoxin
5.) ตัวบล็อกของการกระทำ anticholinergic - กดดัน
cholinosterase ซึ่งทำให้เกิด depolarization และ
การปิดใช้งานตัวรับ ซึ่งรวมถึงออร์กาโนฟอสฟอรัส
สารประกอบ: ไดคลอร์วอส คาร์โบฟอส
34. 34 ไซแนปส์ไฟฟ้า.
ลักษณะของระบบประสาทส่วนกลาง แต่ยังพบในรอบนอก (หัวใจ, กล้ามเนื้อเรียบ
สิ่งทอ).
เป็นตัวแทนของการติดต่ออย่างใกล้ชิด
เยื่อหุ้มเซลล์สองเซลล์
ความกว้างของรอยแยก synaptic เป็นลำดับความสำคัญ
น้อยกว่าในไซแนปส์ทางเคมี
เยื่อหุ้มเซลล์ทั้งสองร่วมกัน
อินทิกรัลโปรตีนที่ก่อตัวขึ้น
ช่องไอออนระหว่างเซลล์ (Nexuses)
การดำรงอยู่ของพวกเขาลดลงอย่างมาก
ความต้านทานระหว่างเซลล์ซึ่งทำให้
การกระจายที่เป็นไปได้ของทวิภาคี
การสลับขั้วระหว่างเซลล์
35.
35ไซแนปส์ไฟฟ้า
1
3
1 - พรีซินแนปติก
พังผืด
2 - โพสต์ซินแน็ปติก
พังผืด
3 - เชื่อมต่อ
2
3
36. 36 โครงสร้างพื้นฐานของ Nexus (ช่องสัมผัส)
37. 37 โครงสร้างและการทำงานของไซแนปส์ไฟฟ้า
- ความกว้างของซินแนปติกช่องว่าง 5 นาโนเมตร
- เส้นผ่านศูนย์กลางรูพรุน 1 นาโนเมตร
- ปัจจุบันลดลงใน 2-4
ครั้ง
- ความล่าช้าในการถือครอง
0.1 มิลลิวินาที
38.
39ความแตกต่างระหว่างไซแนปส์ไฟฟ้าและ
เคมี:
- ขาด
-
-
ความล่าช้าของซินแนปติก
การถือครองทวิภาคี
เร้าอารมณ์
หมายถึงน่าตื่นเต้น
ประสาท
ไวต่อการเปลี่ยนแปลงน้อยลง
อุณหภูมิ
ความเมื่อยล้าน้อยลงมาก
44. 44 ลำดับชั้นของส่วนประกอบที่หดตัวของโครงสร้างของกล้ามเนื้อโครงร่าง
45 คุณสมบัติทางสรีรวิทยาของกล้ามเนื้อความตื่นเต้นง่าย
การนำไฟฟ้า
ความสามารถ
ที่พัก
การหดตัว
45. 45 คุณสมบัติทางสรีรวิทยาของกล้ามเนื้อ
46คุณสมบัติทางกายภาพของกล้ามเนื้อ
1.Extensibility - เพิ่มขนาด
ภายใต้อิทธิพลของโหลดภายนอก
2. ความยืดหยุ่น - กลับคืนสู่สภาพเดิม
สภาพหลังขนถ่าย.
3. ปั้น - รักษาที่กำหนด
โหลดภายนอก, ความยาว
4.ความหนืด-แรงดึง.
46. 46 คุณสมบัติทางกายภาพของกล้ามเนื้อ
47หน้าที่ของกล้ามเนื้อโครงร่าง
(คิดเป็น 40% ของน้ำหนักตัว)
1. เคลื่อนไหวร่างกายในอวกาศ
2. เคลื่อนไหวร่างกายแต่ละส่วน
สัมพันธ์กับเพื่อน
3. การบำรุงรักษาท่าทาง (ฟังก์ชันคงที่)
4. การเคลื่อนไหวของเลือดและน้ำเหลือง
5. การควบคุมอุณหภูมิ
6. การมีส่วนร่วมในการหายใจ
7. การป้องกันอวัยวะภายใน
8. คลังเก็บน้ำ ไกลโคเจน โปรตีน และเกลือ
9. ตัวรับ (proprio-, baro-, value-,
ตัวรับความร้อน)
47. 47 หน้าที่ของกล้ามเนื้อโครงร่าง (คิดเป็น 40% ของน้ำหนักตัว)
48ประเภทของเส้นใยโครงกระดูก
เฟส
เส้นใยที่รวดเร็ว
ด้วยชนิดไกลโคไลติก
ออกซิเดชัน (สีขาว)
พวกเขามี
บาดแผลที่แข็งแกร่ง,
เส้นใยที่รวดเร็ว
ประเภทออกซิไดซ์
ดำเนินการอย่างรวดเร็ว
บาดแผลที่แข็งแกร่งและ
แต่เหนื่อยเร็ว
เหนื่อยอ่อน
เส้นใยช้า
ประเภทออกซิไดซ์
ทำหน้าที่บำรุงรักษา
ท่าทางของบุคคล หน่วยประสาท
กล้ามเนื้อเหล่านี้มีหนูมากที่สุด เส้นใย
โทนิค
ช้า,
ได้อย่างมีประสิทธิภาพ
ทำงานในสามมิติ
โหมด.
กล้าม
เส้นใย
ไม่
สร้าง PD
และไม่
ปฏิบัติตามกฎหมาย "ทั้งหมดหรือ
ไม่มีอะไร".
แอกซอนของเซลล์ประสาทสั่งการมี
ซินแนปติกมากมาย
ติดต่อ
กับ
พังผืด
เส้นใยกล้ามเนื้อ
48. 48 ประเภทของเส้นใยโครงร่าง
49โหมดของการหดตัวของกล้ามเนื้อ
1. โสด
2. ผลรวม (เต็มและไม่สมบูรณ์)
บาดทะยัก dentate และเรียบ
3. ความถี่ที่เหมาะสมและเชิงลบ
ตัด
4. ติดต่อ
49. 49 โหมดของการหดตัวของกล้ามเนื้อ
50.
51ทฤษฎีการสรุปผลของการหดตัวของกล้ามเนื้อ
1. Helmholtz - หลักการซ้อนทับ:
การเพิ่มแอมพลิจูดของการหดตัวครั้งเดียว
2. Vvedensky - มูลค่ารวม
ขึ้นอยู่กับสถานะการทำงาน
ผ้าเช่น จากเฟสใด (การคำนวณ
หรือความหักเหของแสง) นำไปใช้ต่อไป
ระคายเคือง
3. Babsky - เชื่อมโยงค่าของการรวมกับ
การสะสม ATP และ Ca 2+ ที่เหลือจาก
ตัดก่อนหน้านี้
4. ทฤษฎีสมัยใหม่ - เพิ่มขึ้น
การสร้างสะพานแอคโตไมโอซิน
ไซแนปส์ - โครงสร้างพิเศษที่ให้การถ่ายโอนการกระตุ้นจากเซลล์หนึ่งไปยังอีกเซลล์หนึ่ง แนวคิดของ SINAPSE ได้รับการแนะนำในสรีรวิทยาโดย C. Sherrington (การเชื่อมต่อ, การติดต่อ) ไซแนปส์ให้การสื่อสารการทำงานระหว่างเซลล์แต่ละเซลล์ พวกเขาแบ่งออกเป็นเส้นประสาทประสาท, ประสาทและกล้ามเนื้อและประสาทของเซลล์ประสาทกับเซลล์หลั่ง (neuro-glandular) มีสามส่วนการทำงานในเซลล์ประสาท: โสม เดนไดรต์ และแอกซอน ดังนั้นจึงมีการติดต่อระหว่างเซลล์ประสาทที่เป็นไปได้ทั้งหมด ตัวอย่างเช่น axo-axonal, axo-somatic และ axo-dendritic
การจัดหมวดหมู่.
1) ตามที่ตั้งและเป็นของโครงสร้างที่เกี่ยวข้อง:
- อุปกรณ์ต่อพ่วง(ประสาทและกล้ามเนื้อ, ระบบประสาท, ตัวรับ-เซลล์ประสาท);
- ศูนย์กลาง(axo-somatic, axo-dendritic, axo-axonal, somato-dendritic, somato-somatic);
2) กลไกของการกระทำ - กระตุ้นและยับยั้ง;
3) วิธีการส่งสัญญาณ - เคมี,ไฟฟ้า,ผสม.
4) สารเคมีถูกจัดประเภทตามผู้ไกล่เกลี่ยด้วยความช่วยเหลือของการถ่ายโอน - cholinergic, adrenergic, serotonergic, glycinergic. เป็นต้น
โครงสร้างไซแนปส์
ไซแนปส์ประกอบด้วยองค์ประกอบหลักดังต่อไปนี้:
เมมเบรน Presynaptic (ในประสาทและกล้ามเนื้อไซแนปส์ - นี่คือแผ่นท้าย):
เมมเบรนโพสต์ซินแน็ปติก;
แหว่ง synaptic รอยแหว่งซินแนปติกนั้นเต็มไปด้วยเนื้อเยื่อเกี่ยวพันที่มีโอลิโกแซ็กคาไรด์ ซึ่งมีบทบาทเป็นโครงสร้างรองรับสำหรับเซลล์ที่สัมผัสกันทั้งสองเซลล์
ระบบการสังเคราะห์และการปลดปล่อยคนกลาง
ระบบการปิดใช้งานของมัน
ในประสาทและกล้ามเนื้อไซแนปส์ เมมเบรน presynaptic เป็นส่วนหนึ่งของเยื่อหุ้มเส้นประสาทที่สิ้นสุดในบริเวณที่สัมผัสกับเส้นใยกล้ามเนื้อ เมมเบรน postynaptic เป็นส่วนหนึ่งของเมมเบรนของเส้นใยกล้ามเนื้อ
โครงสร้างของประสาทและกล้ามเนื้อไซแนปส์
1 - เส้นใยประสาทไมอิลิน
2 - เส้นประสาทที่สิ้นสุดด้วยถุงไกล่เกลี่ย;
3 - เยื่อหุ้มเซลล์ subsynaptic ของเส้นใยกล้ามเนื้อ
4 - แหว่ง synaptic;
เมมเบรน 5-postsynaptic ของเส้นใยกล้ามเนื้อ
6 - ไมโอไฟบริล;
7 - ซาร์โคพลาสซึม;
8 - ศักยภาพการทำงานของเส้นใยประสาท;
9 - ศักยภาพของแผ่นท้าย (EPSP):
10 - ศักยภาพการทำงานของเส้นใยกล้ามเนื้อ
ส่วนของเมมเบรนโพสซินแนปติกที่อยู่ตรงข้ามกับพรีซินแนปติกเรียกว่า ซับซินแนปติกเมมเบรน คุณสมบัติของเมมเบรน subsynaptic คือการมีอยู่ของตัวรับพิเศษที่ไวต่อผู้ไกล่เกลี่ยและการปรากฏตัวของช่องทางที่ขึ้นอยู่กับเคมี ในโพสซินแนปติกเมมเบรน นอกซับซินแนปติก มีช่องแรงดันไฟฟ้า
กลไกการส่งผ่านการกระตุ้นใน synapses กระตุ้นทางเคมี. ในปี 1936 Dale ได้พิสูจน์ว่าเมื่อเส้นประสาทสั่งการถูกกระตุ้น อะซิติลโคลีนจะถูกปลดปล่อยออกมาในกล้ามเนื้อโครงร่างที่ส่วนท้ายของมัน ในประสาทที่มีการส่งผ่านสารเคมีการกระตุ้นจะถูกส่งด้วยความช่วยเหลือของผู้ไกล่เกลี่ย (ตัวกลาง) ผู้ไกล่เกลี่ยเป็นสารเคมีที่รับประกันการส่งผ่านของการกระตุ้นในประสาท ผู้ไกล่เกลี่ยในไซแนปส์ของประสาทและกล้ามเนื้อคืออะซิติลโคลีนในไซแนปส์ของเส้นประสาทที่กระตุ้นและยับยั้ง - อะซิติลโคลีน, คาเทโคลามีน - อะดรีนาลีน, นอร์อิพิเนฟริน, โดปามีน เซโรโทนิน; กรดอะมิโนที่เป็นกลาง - กลูตามีน, แอสปาร์ติก; กรดอะมิโนที่เป็นกรด - ไกลซีน, กรดแกมมา - อะมิโนบิวทีริก; โพลีเปปไทด์: สาร P, เอนเคฟาลิน, โซมาโตสแตติน; สารอื่นๆ: ATP, ฮีสตามีน, พรอสตาแกลนดิน
ผู้ไกล่เกลี่ยแบ่งออกเป็นหลายกลุ่มขึ้นอยู่กับธรรมชาติของพวกเขา:
โมโนเอมีน (acetylcholine, dopamine, norepinephrine, serotonin.);
กรดอะมิโน (กรดแกมมาอะมิโนบิวทีริก - GABA, กรดกลูตามิก, ไกลซีน ฯลฯ );
Neuropeptides (สาร P, endorphins, neurotensin, ACTH, angiotensin, vasopressin, somatostatin เป็นต้น)
การสะสมของผู้ไกล่เกลี่ยในการก่อตัวของ presynaptic เกิดขึ้นเนื่องจากการขนส่งจากบริเวณ perinuclear ของเซลล์ประสาทด้วยความช่วยเหลือของ axstock ที่รวดเร็ว การสังเคราะห์สื่อกลางที่เกิดขึ้นในขั้วซินแนปติกจากผลิตภัณฑ์ที่แตกแยก การนำสารสื่อประสาทกลับมาใช้ใหม่จาก synaptic cleft
ปลายประสาท presynaptic มีโครงสร้างสำหรับการสังเคราะห์สารสื่อประสาท หลังจากการสังเคราะห์ สารสื่อประสาทจะถูกบรรจุเป็นถุง เมื่อถูกกระตุ้น synaptic vesicles เหล่านี้จะหลอมรวมกับ presynaptic membrane และสารสื่อประสาทจะถูกปล่อยเข้าไปใน synaptic cleft มันแพร่กระจายไปยังเมมเบรนโพสซินแนปติกและจับกับตัวรับที่เฉพาะเจาะจง อันเป็นผลมาจากการก่อตัวของสารสื่อประสาท-ตัวรับคอมเพล็กซ์ เยื่อโพสต์ซินแนปติกจะซึมผ่านไปยังแคตไอออนและดีโพลาไรซ์ได้ ซึ่งส่งผลให้มีศักยภาพในการโพสต์ซินแนปติกแบบกระตุ้นและจากนั้นจึงมีศักยภาพในการดำเนินการ ตัวกลางถูกสังเคราะห์ขึ้นในปลายพรีซินแนปติกจากวัสดุที่จัดหามาโดยการขนส่งด้วยแอกซอน คนกลาง "ปิดใช้งาน" นั่นคือ ถูกแยกออกหรือถูกลบออกจากรอยแหว่งซินแนปติกโดยกลไกการขนส่งแบบย้อนกลับไปยังปลายทางพรีซินแนปติก
คุณค่าของแคลเซียมไอออนในการหลั่งสารสื่อกลาง.
การหลั่งของผู้ไกล่เกลี่ยเป็นไปไม่ได้หากไม่มีแคลเซียมไอออนเข้าร่วมในกระบวนการนี้ เมื่อมีการดีโพลาไรเซชันของเยื่อพรีไซแนปติก แคลเซียมจะเข้าสู่ปลายพรีไซแนปติกผ่านช่องแคลเซียมที่มีแรงดันไฟฟ้าเฉพาะในเมมเบรนนี้ ความเข้มข้นของแคลเซียมในแอกโซพลาสซึมคือ 110 -7 M โดยมีแคลเซียมเข้ามาและเพิ่มความเข้มข้นเป็น 110 - 4 M เกิดการหลั่งสารไกล่เกลี่ย ความเข้มข้นของแคลเซียมใน axoplasm หลังจากสิ้นสุดการกระตุ้นจะลดลงโดยการทำงานของระบบ: การขนส่งที่ใช้งานจากเทอร์มินัล, การดูดซึมโดยไมโตคอนเดรีย, การจับกันโดยระบบบัฟเฟอร์ภายในเซลล์ ที่เหลือ การระบายของถุงน้ำที่ผิดปกติเกิดขึ้น โดยไม่เพียงแต่ปล่อยโมเลกุลเดี่ยวของผู้ไกล่เกลี่ยเท่านั้น แต่ยังมีการปลดปล่อยส่วนต่าง ๆ ควอนตัมของผู้ไกล่เกลี่ยด้วย ควอนตัมของ acetylcholine รวมประมาณ 10,000 โมเลกุล
ไซแนปส์เป็นสถานที่ติดต่อของเซลล์ประสาทกับเซลล์ประสาทหรืออวัยวะบริหารอื่น ไซแนปส์ทั้งหมดแบ่งออกเป็นกลุ่มต่อไปนี้:
1.โดยกลไกการส่งผ่าน:
ก. ไฟฟ้า. การกระตุ้นจะถูกส่งผ่านสนามไฟฟ้า ดังนั้นจึงสามารถส่งได้ทั้งสองทิศทาง มีไม่กี่คนในระบบประสาทส่วนกลาง
ข. เคมี. การกระตุ้นผ่านพวกมันจะถูกส่งผ่านความช่วยเหลือของ FAV ซึ่งเป็นสารสื่อประสาท ส่วนใหญ่อยู่ในคมช.
วี. ผสม
2. โดยการแปล:
ก. เซ็นทรัล ตั้งอยู่ใน ทส.
ข. อุปกรณ์ต่อพ่วง, นอกนั้น. เหล่านี้คือไซแนปส์ประสาทและกล้ามเนื้อและไซแนปส์ของส่วนต่อพ่วงของระบบประสาทอัตโนมัติ
3. ตามสรีรวิทยา:
ก. น่าตื่นเต้น
ข. เบรค
4. ขึ้นอยู่กับสารสื่อประสาทที่ใช้ในการส่ง:
ก. Cholinergic - ตัวกลาง acetylcholine (ACh)
ข. Adrenergic - norepinephrine (NA)
วี. Serotonergic - เซโรโทนิน (ST)
d. Glycinergic - กรดอะมิโนไกลซีน (GLI)
อี GABAergic - กรดแกมมาอะมิโนบิวทีริก (GABA)
อี โดปามีน - โดปามีน (DA)
และ. Peptidergic mediators เป็นนิวโรเปปไทด์ โดยเฉพาะอย่างยิ่ง บทบาทของสารสื่อประสาทนั้นดำเนินการโดยสาร P, เปปไทด์ opioid β-endorphin เป็นต้น
สันนิษฐานว่ามีประสาทซึ่งทำหน้าที่ของผู้ไกล่เกลี่ยโดยฮีสตามีน, เอทีพี, กลูตาเมต, แอสพาเตต
5. ตามตำแหน่งของไซแนปส์:
ก. Axo-dendritic (ระหว่างแอกซอนของหนึ่งกับเดนไดรต์ของเซลล์ประสาทที่สอง)
ข. แอกโซ-แอกซอน
วี. แอกโซโซมาติก
Dendro-โซมาติก
e. Dendro-เดนไดรติก
สามประเภทแรกเป็นเรื่องธรรมดาที่สุด
โครงสร้างของ synapses ทางเคมีทั้งหมดมีความคล้ายคลึงกันโดยพื้นฐาน ตัวอย่างเช่น ไซแนปส์ axo-dendritic ประกอบด้วยองค์ประกอบต่อไปนี้:
1. Presynaptic End หรือ Terminal (ปลายแอกซอน)
2. Synaptic plaque ความหนาของส่วนท้าย
3. เยื่อพรีซินแนปติกที่หุ้มส่วนปลายของพรีซินแนปติก
4. Synaptic vesicles ในแผ่นโลหะที่มีสารสื่อประสาท
5. เยื่อโพสต์ไซแนปติก (Postsynaptic membrane) ครอบคลุมบริเวณเดนไดรต์ที่อยู่ติดกับแผ่นโลหะ
6. Synaptic cleft ที่แยกเยื่อก่อนและหลัง Synaptic กว้าง 10-50 nM
7. ตัวรับเคมี (Chemoreceptors) ซึ่งเป็นโปรตีนที่สร้างขึ้นในเยื่อหุ้มเซลล์หลังไซแนปติกและจำเพาะสำหรับสารสื่อประสาท ตัวอย่างเช่น ใน cholinergic synapses เหล่านี้คือ cholinergic receptors, adrenergic synapses คือ adrenoreceptors เป็นต้น ข้าว.
สารสื่อประสาทอย่างง่ายถูกสังเคราะห์ที่ส่วนปลายของพรีซินแนปติก ส่วนสารสื่อประสาทเปปไทด์ถูกสังเคราะห์ในโสมของเซลล์ประสาท จากนั้นจึงเคลื่อนย้ายไปตามแอกซอนไปยังส่วนปลาย
J กลไกการส่งสารกระตุ้นในไซแนปส์เคมี
ผู้ไกล่เกลี่ยที่มีอยู่ในถุงซินแนปติกจะเกิดขึ้นในร่างกายของเซลล์ประสาท (และเข้าสู่จุดสิ้นสุดของซินแนปติกโดยผ่านแอกซอนทั้งหมด) หรือในแผ่นโลหะซินแนปติก สำหรับการสังเคราะห์สื่อกลางนั้นจำเป็นต้องใช้เอนไซม์ที่เกิดขึ้นในร่างกายของเซลล์บนไรโบโซม ในแผ่นโลหะ synaptic โมเลกุลของผู้ไกล่เกลี่ยจะสะสมและ "บรรจุ" ไว้ในถุงซึ่งจะถูกเก็บไว้จนกว่าจะปล่อยออกมา พบ (A. Fett และ B. Katz, 1952) ว่าถุงหนึ่งประกอบด้วยโมเลกุล acetylcholine ตั้งแต่ 3 ถึง 10,000 โมเลกุล . ปริมาณนี้เรียกว่าควอนตัมของผู้ไกล่เกลี่ย เมื่อเส้นประสาทระคายเคืองในส่วน presynaptic ของ synapse ตุ่ม 250 ถึง 500 จะถูกทำลาย การมาถึงของกระแสประสาท (PD) ในแผ่น synaptic ทำให้เกิดการสลับขั้วของ presynaptic membrane และเพิ่มความสามารถในการซึมผ่านของไอออน Ca2+ ไอออน Ca2+ ที่เข้าสู่ synaptic plaque ทำให้เกิดการหลอมรวมของ synaptic vesicles กับ presynaptic membrane และปล่อยสารของพวกมัน (exocytosis) เข้าไปใน synaptic cleft หลังจากปล่อยตัวไกล่เกลี่ยแล้ว วัสดุตุ่มจะถูกนำมาใช้เพื่อสร้างตุ่มใหม่ โมเลกุลของทรานสมิตเตอร์จะแพร่ผ่านรอยแยกไซแนปติกและจับกับรีเซพเตอร์ที่อยู่บนเยื่อหุ้มโพสซินแนปติก ซึ่งสามารถจดจำโครงสร้างโมเลกุลของผู้ไกล่เกลี่ยได้ การแพร่กระจายของสื่อกลางผ่าน synaptic cleft ใช้เวลาประมาณ 0.5 มิลลิวินาที เมื่อโมเลกุลของตัวรับจับกับสื่อกลาง โครงร่างของมันจะเปลี่ยนไป ซึ่งนำไปสู่การเปิดช่องไอออนและการเข้ามาของไอออนในเซลล์หลังซินแนปติก ทำให้เกิดการสลับขั้วหรือไฮเปอร์โพลาไรเซชัน เมมเบรน ขึ้นอยู่กับลักษณะของตัวกลางที่ปล่อยออกมาและโครงสร้างของตัวรับโมเลกุล โมเลกุลตัวกลาง หลังจากทำหน้าที่กับตัวรับแล้ว จะถูกกำจัดออกจากรอยแหว่งไซแนปติกทันทีโดยการดูดซึมกลับโดยเยื่อพรีซินาติก หรือโดยการแพร่ หรือโดยเอนไซม์ ไฮโดรไลซิส อะซิติลโคลีนถูกไฮโดรไลซ์โดยเอนไซม์อะซิติลโคลีนเอสเตอเรสที่อยู่บนเยื่อโพสซินแนปติก จากนั้นผลิตภัณฑ์จากความแตกแยกจะถูกดูดซึมกลับเข้าไปในคราบจุลินทรีย์และเปลี่ยนเป็นอะเซทิลโคลีนอีกครั้ง นอร์-อะดรีนาลีนถูกไฮโดรไลซ์โดยเอนไซม์โมโนเอมีนออกซิเดส ศักยภาพของโพสต์ซินแนปติกที่กระตุ้นและยับยั้ง ในประสาทกระตุ้น excitatory ช่องโซเดียมและโพแทสเซียมที่เฉพาะเจาะจงจะเปิดขึ้นภายใต้การกระทำของ acetylcholine และไอออน Na + เข้าสู่เซลล์และไอออน K + ปล่อยให้เป็นไปตามการไล่ระดับความเข้มข้น เป็นผลให้เกิดการสลับขั้วของเมมเบรนโพสต์ซินแน็ปติก เรียกว่าศักยภาพโพสซินแน็ปทิกกระตุ้น (EPSP) แอมพลิจูดมีขนาดเล็ก แต่ระยะเวลานานกว่าศักยภาพในการดำเนินการ ในไซแนปส์แบบยับยั้ง การปลดปล่อยตัวกลางจะเพิ่มการซึมผ่านของเมมเบรนโพสซินแนปติกโดยการเปิดช่องเฉพาะสำหรับ K+ และ SG ไอออน ไอออนเหล่านี้เคลื่อนที่ไปตามการไล่ระดับความเข้มข้นทำให้เกิดไฮเปอร์โพลาไรเซชันของเมมเบรน เรียกว่า ศักยภาพในการยับยั้งโพสต์ซินแนปติก (IPSP)
ประสาทไฟฟ้า
ไซแนปส์ไฟฟ้ามีโครงสร้างพิเศษ ความกว้างของรอยแยกซินแนปติกคือ 2–3 นาโนเมตร และความต้านทานรวมต่อกระแสไฟฟ้าจากด้านข้างของเยื่อหุ้มและของเหลวที่อุดรอยแยกนั้นน้อยมาก ไอออนที่มีกระแสไฟฟ้าไม่สามารถผ่านเยื่อหุ้มไขมันได้ ดังนั้น ไอออนเหล่านี้จะถูกส่งผ่านแชนเนลโปรตีน การเชื่อมต่อระหว่างเซลล์ดังกล่าวเรียกว่า nexuses หรือ "gap junctions" (รูปที่ 42) ในแต่ละเซลล์ของเยื่อหุ้มเซลล์ที่อยู่ติดกันจะมีการกระจายเป็นระยะๆ<<коннексоны>> เจาะทะลุความหนาทั้งหมดของเมมเบรน ตั้งอยู่ในลักษณะที่จุดสัมผัสของเซลล์อยู่ตรงข้ามกันและช่องว่างอยู่ในบรรทัดเดียวกัน ช่องที่เกิดขึ้นในลักษณะนี้มีเส้นผ่านศูนย์กลางขนาดใหญ่ ซึ่งหมายถึงการนำไฟฟ้าสูงสำหรับไอออน แม้แต่โมเลกุลที่ค่อนข้างใหญ่ก็สามารถผ่านเข้าไปได้ รอยต่อของช่องว่างเป็นเรื่องปกติในระบบประสาทส่วนกลางและมีแนวโน้มที่จะเชื่อมต่อกลุ่มของเซลล์ที่ทำงานพร้อมกัน
แรงกระตุ้นจะผ่านไซแนปส์โดยไม่ชักช้า สามารถดำเนินการได้ทั้งสองทิศทาง และการส่งผ่านจะไม่ได้รับผลกระทบจากยาหรือสารเคมีอื่นๆ
วันที่ 22 ประสาทและกล้ามเนื้อประสาท
ชุมทางประสาทและกล้ามเนื้อเป็นไซแนปส์ชนิดพิเศษระหว่างส่วนปลายของเซลล์ประสาทสั่งการ (โมโตนิวรอน) และเอ็นโดไมเซียมของเส้นใยกล้ามเนื้อ เส้นใยกล้ามเนื้อแต่ละเส้นมีพื้นที่พิเศษ - แผ่นท้ายของมอเตอร์ซึ่งแอกซอนของเซลล์ประสาทสั่งการแตกกิ่งก้านซึ่งสร้างกิ่งก้านที่ไม่มีไมอีลินที่วิ่งในร่องตื้น ๆ ตามพื้นผิวของเยื่อหุ้มกล้ามเนื้อ เยื่อหุ้มเซลล์ของกล้ามเนื้อ - ซาร์โคเลมมา - ก่อให้เกิดรอยพับลึกจำนวนมากที่เรียกว่า โพสซินแนปติกพับ ไซโตพลาสซึมของปลายเซลล์ประสาทสั่งการนั้นคล้ายกับเนื้อหาของแผ่นซินแนปติก กลไกการถ่ายโอนการกระตุ้นจะเหมือนกัน อันเป็นผลมาจากการกระตุ้นของเซลล์ประสาทสั่งการ ทำให้เกิดการสลับขั้วของพื้นผิวของซาร์โคเลมมา ซึ่งเรียกว่า end plate potential (EPP) ขนาดของศักยภาพนี้เพียงพอที่จะสร้างศักยภาพในการดำเนินการที่แพร่กระจายไปตาม sarcolemma ลึกเข้าไปในเส้นใยและทำให้กล้ามเนื้อหดตัว
เซลล์ประสาทที่ 23 เป็นหน่วยโครงสร้างและหน้าที่หลักของระบบประสาท เซลล์ประสาทเป็นเซลล์ที่มีความเชี่ยวชาญสูงซึ่งได้รับการดัดแปลงสำหรับการรับ การเข้ารหัส การประมวลผล การบูรณาการ การจัดเก็บ และการส่งข้อมูล เซลล์ประสาทประกอบด้วยร่างกายและกระบวนการสองประเภท: เดนไดรต์ที่แตกแขนงสั้นและกระบวนการที่ยาว - แอกซอน (รูปที่ 42) ตัวเซลล์มีขนาดเส้นผ่านศูนย์กลาง 5 ถึง 150 ไมครอน มันเป็นศูนย์กลางการสังเคราะห์ทางชีวภาพของเซลล์ประสาทซึ่งกระบวนการเมแทบอลิซึมที่ซับซ้อนเกิดขึ้น ร่างกายประกอบด้วยนิวเคลียสและไซโตพลาสซึมซึ่งมีออร์แกเนลล์จำนวนมากที่เกี่ยวข้องกับการสังเคราะห์โปรตีนในเซลล์ (โปรตีน) แอกซอน กระบวนการใยยาวของแอกซอนออกจากตัวเซลล์ซึ่งทำหน้าที่ส่งข้อมูล แอกซอนถูกหุ้มด้วยเยื่อไมอีลินชนิดพิเศษที่สร้างสภาวะที่เหมาะสมที่สุดสำหรับการส่งสัญญาณ ปลายแอกซอนแตกแขนงออกมาก แขนงปลายของมันติดต่อกับเซลล์อื่นๆ จำนวนมาก (เส้นประสาท กล้ามเนื้อ ฯลฯ) กลุ่มแอกซอนก่อตัวเป็นเส้นใยประสาท
เดนไดรต์เป็นกระบวนการที่แตกกิ่งก้านสาขามากซึ่งขยายออกจากตัวเซลล์เป็นจำนวนมาก เซลล์ประสาทหนึ่งเซลล์สามารถแยกเดนไดรต์ได้มากถึง 1,000 เดนไดรต์ ร่างกายและเดนไดรต์ถูกปกคลุมด้วยเมมเบรนเดียวและสร้างพื้นผิวที่รับ (รับ) ของเซลล์ มันมีการติดต่อส่วนใหญ่จากเซลล์ประสาทอื่น - ไซแนปส์ ผนังเซลล์ - เมมเบรน - เป็นฉนวนไฟฟ้าที่ดี ทั้งสองด้านของเมมเบรนมีความต่างศักย์ไฟฟ้า - ศักย์ของเมมเบรนซึ่งเป็นระดับที่เปลี่ยนแปลงเมื่อเปิดใช้งานการสัมผัสซินแนปติก ไซแนปส์มีโครงสร้างที่ซับซ้อน (ดูรูปที่ 42) มันเกิดจากเยื่อหุ้มสองอัน: พรีไซแนปติกและโพสต์ซินแนปติก พรีซินแนปติกเมมเบรนอยู่ที่ส่วนท้ายของแอกซอนที่ทำหน้าที่ส่งสัญญาณ โพสต์ซินแนปติก - บนร่างกายหรือเดนไดรต์ที่ส่งสัญญาณ ในไซแนปส์ เมื่อมีสัญญาณมาถึง สารเคมีสองชนิดจะถูกปล่อยออกจากถุงซินแนปติก - สารกระตุ้น (อะซีติลโคลีน, อะดรีนาลีน, นอเรพิเนฟริน) และสารยับยั้ง (เซโรโทนิน, กรดแกมมา-อะมิโนบิวทีริก) สารเหล่านี้ - ผู้ไกล่เกลี่ยที่ทำหน้าที่ในเมมเบรนโพสต์ซินแน็ปติกเปลี่ยนคุณสมบัติในบริเวณที่สัมผัส ด้วยการปลดปล่อยตัวกลางกระตุ้น excitatory ศักยภาพโพสต์ซินแน็ปทิก excitatory (EPSP) เกิดขึ้นในพื้นที่ติดต่อด้วยการกระทำของตัวกลางยับยั้ง ศักยภาพโพสต์ซินแน็ปติกยับยั้ง (IPSP) ตามลำดับ เกิดขึ้น ผลรวมของพวกเขานำไปสู่การเปลี่ยนแปลงในศักยภาพภายในเซลล์ต่อการสลับขั้วหรือไฮเปอร์โพลาไรเซชัน เมื่อสลับขั้ว เซลล์จะสร้างแรงกระตุ้นที่ส่งผ่านแอกซอนไปยังเซลล์อื่นหรืออวัยวะที่ทำงานอยู่ ระหว่างไฮเปอร์โพลาไรเซชัน เซลล์ประสาทจะเข้าสู่สถานะยับยั้งและไม่สร้างกิจกรรมอิมพัลส์ (รูปที่ 43) ความหลากหลายและหลากหลายของไซแนปส์ทำให้เกิดการเชื่อมต่อระหว่างเซลล์ประสาทที่กว้างขวางและการมีส่วนร่วมของเซลล์ประสาทเดียวกันในการเชื่อมโยงการทำงานที่แตกต่างกัน
การจัดหมวดหมู่
การจำแนกโครงสร้าง
ตามจำนวนและการจัดเรียงตัวของเดนไดรต์และแอกซอน เซลล์ประสาทจะแบ่งออกเป็นเซลล์ประสาทที่ไม่ใช่แอกซอน เซลล์ประสาทยูนิโพลาร์ เซลล์ประสาทเทียมยูนิโพลาร์ เซลล์ประสาทสองขั้ว และเซลล์ประสาทหลายขั้ว
เซลล์ประสาทไร้แอกซอน- เซลล์ขนาดเล็กจัดกลุ่มใกล้กับไขสันหลังในปมประสาท intervertebral ซึ่งไม่มีอาการทางกายวิภาคของการแยกกระบวนการออกเป็นเดนไดรต์และแอกซอน กระบวนการทั้งหมดในเซลล์จะคล้ายกันมาก จุดประสงค์การทำงานของเซลล์ประสาทไร้แอกซอนนั้นเข้าใจได้ไม่ดีนัก
เซลล์ประสาทยูนิโพลาร์- มีเซลล์ประสาทที่มีกระบวนการเดียว เช่น ในนิวเคลียสรับความรู้สึกของเส้นประสาทไตรเจมินัลในสมองส่วนกลาง
เซลล์ประสาทสองขั้ว- เซลล์ประสาทที่มีแอกซอนหนึ่งอันและเดนไดรต์หนึ่งอันซึ่งอยู่ในอวัยวะรับความรู้สึกพิเศษ - เรตินา, เยื่อบุผิวรับกลิ่นและกระเปาะ, ปมประสาทหูและขนถ่าย
เซลล์ประสาทหลายขั้ว- เซลล์ประสาทที่มีแอกซอนหนึ่งอันและเดนไดรต์หลายอัน เซลล์ประสาทประเภทนี้มีอิทธิพลเหนือระบบประสาทส่วนกลาง
เซลล์ประสาทหลอกยูนิโพลาร์- มีเอกลักษณ์ในแบบของมัน กระบวนการหนึ่งออกจากร่างกายซึ่งจะแบ่งออกเป็นรูปตัว T ทันที ทางเดินเดี่ยวทั้งหมดนี้ปกคลุมด้วยปลอกไมอีลินและโครงสร้างเป็นตัวแทนของแอกซอน แม้ว่าตามกิ่งก้านสาขาหนึ่ง การกระตุ้นไม่ได้มาจากร่างกายของเซลล์ประสาท โครงสร้าง dendrites เป็นสาขาในตอนท้ายของกระบวนการนี้ (อุปกรณ์ต่อพ่วง) โซนทริกเกอร์เป็นจุดเริ่มต้นของการแตกแขนงนี้ (นั่นคืออยู่นอกเซลล์) เซลล์ประสาทดังกล่าวพบในปมประสาทไขสันหลัง
แม้จะมีลักษณะทั่วไปของคุณสมบัติหลักขององค์กร แต่เคมีซินเซสนั้นแตกต่างกันในผู้ไกล่เกลี่ยที่ใช้ ลักษณะของการกระทำและตำแหน่ง ด้วยเหตุนี้จึงมีหลายวิธีในการจำแนกประเภทของสารเคมี
โดย ประเภทคนกลางไซแนปส์แบ่งออกเป็น cholinergic (ผู้ไกล่เกลี่ย - ACh), กลูตามาเทอจิค (ผู้ไกล่เกลี่ย - กลูตาเมต), adrenergic (ผู้ไกล่เกลี่ย - norepinephrine), โดปามีน (ผู้ไกล่เกลี่ย - โดปามีน) เป็นต้น
โดย ผลประสาทถูกแบ่งออกเป็นกระตุ้นและยับยั้ง
โดย ตำแหน่งในระบบประสาทประสาทแบ่งออกเป็นส่วนกลาง (อยู่ในระบบประสาทส่วนกลาง) และอุปกรณ์ต่อพ่วง (อยู่ในระบบประสาทส่วนปลาย)
ประสาทส่วนปลายคือการสัมผัสของแอกซอนกับกล้ามเนื้อทุกประเภทรวมถึงเซลล์ต่อม ประสาทส่วนปลายมีขนาดใหญ่กว่าศูนย์กลางและมีขนาดถึง 50-100 ไมครอน (รูปที่ 3.26) ดังนั้นในแต่ละเส้นใยกล้ามเนื้อโครงร่างที่โตเต็มที่จะมีไซแนปส์ประสาทและกล้ามเนื้อเพียงอันเดียวที่เกิดจากปลายประสาทของแอกซอนของเซลล์ประสาทสั่งการ
ข้าว. 3.26.
การส่งผ่านแบบซินแนปติกในจานท้ายเกิดขึ้นจากการมีส่วนร่วมของผู้ไกล่เกลี่ย ACh และนำไปสู่การสร้าง PCR แอมพลิจูดสูง (30–40 mV) PPP ดังกล่าวสูงกว่าเกณฑ์การสร้าง AP 2-3 เท่า ดังนั้น presynaptic AP แต่ละอันทำให้เกิดการสร้าง PEP ที่มีแอมพลิจูดสูง ใน 100% ของกรณีจะนำไปสู่การสร้าง AP ของกล้ามเนื้อและการหดตัวของเส้นใยกล้ามเนื้อตามมา
ไซแนปส์กับอวัยวะภายใน (เซลล์กล้ามเนื้อเรียบ คาร์ดิโอไมโอไซต์ หรือเซลล์ต่อม) สร้างแอกซอนของเซลล์ประสาทซิมพาเทติกและพาราซิมพาเทติกหลังปมประสาท ตามกฎแล้วในแอกซอนดังกล่าว การจัดกลุ่มของถุงน้ำและการปล่อยตัวไกล่เกลี่ยจะไม่เกิดขึ้นจากตาเดี่ยวสุดท้าย เช่นเดียวกับในประสาทและกล้ามเนื้อประสาท แต่ตามเส้นทางของแอกซอนจากเส้นเลือดขอดจำนวนมาก มีส่วนขยายดังกล่าวมากถึง 250-300 ต่อความยาวแอกซอน 1 มม. ระยะห่างระหว่างเยื่ออิรีซินแนปติกและโพสซินแนปติกในไซแนปส์ดังกล่าวมีขนาดใหญ่ ตั้งแต่ 80 ถึง 250 นาโนเมตร และสารสื่อประสาทที่ปล่อยออกมาจะส่งการออกฤทธิ์ไปยังเมแทบอโทรปิกไอโอสต์ซินเนติกรีเซพเตอร์
รูปที่ 11a 3.27 แสดงตัวอย่างของไซแนปส์ที่เกิดจากเส้นใยพาราซิมพาเทติก iostganglionic ในเนื้อเยื่อของกล้ามเนื้อเรียบของกระเพาะอาหาร จะเห็นได้ว่าตลอดเส้นทางของ postganglionic parasympathetic axon มี varices จำนวนมากที่มี synaptic vesicles ที่มี ACh mediator ช่อง Ca 2+ อยู่ที่นี่โดยเป็นส่วนหนึ่งของเยื่อหุ้มพรีไซแนปติก ดังนั้นภายใต้อิทธิพลของ AP ที่แพร่กระจายไปตามแอกซอนและเกิดจากการดีโพลาไรเซชัน การเข้าสู่แคลเซียมไอออนในเส้นเลือดขอดจึงเกิดขึ้น การเปิดตัวควอนตัมคนกลาง

ข้าว. 3.27.
เมื่อ ACh โต้ตอบกับ metabotropic mChRs ของเยื่อโพสต์ซินแนปติก หลังจากเกิด synaptic delay ที่ยาวนาน (1.5-2 ms เทียบกับ 0.3-0.5 ms ใน fast synapses) EPSP ที่กินเวลา 20-50 ms จะเกิดขึ้น สำหรับการเกิดขึ้นของ AP ในเซลล์กล้ามเนื้อเรียบนั้น จำเป็นต้องมีแอมพลิจูดเกณฑ์ของ EPSP ที่ 8-25 mV ตามกฎแล้ว สัญญาณ presynaptic เดียว (single AP) ไม่เพียงพอที่จะทำให้แคลเซียมไอออนเข้าสู่เส้นเลือดขอดและกระตุ้นให้เกิดตุ่มนูน (vesicle exocytosis) ดังนั้นการปลดปล่อยคนกลางออกจากเส้นเลือดขอดของแอกซอนหลังปมประสาทจึงดำเนินการภายใต้การกระทำของ APs presynaptic จำนวนหนึ่ง (วอลเลย์) ที่ต่อเนื่องกันเท่านั้น การกระตุ้นการส่งผ่านในการสัมผัสดังกล่าวทำให้เกิดการเปลี่ยนแปลงของเส้นใยกล้ามเนื้อในผนังของอวัยวะภายในหรือทำให้เกิดการหลั่งในเซลล์ต่อม
ไซแนปส์ส่วนกลางมีความหลากหลายทางโครงสร้างที่ใหญ่มาก จำนวนมากที่สุดคือ axodendritic และ axosomatic synapses - การสัมผัสระหว่างปลายประสาทของแอกซอนของเซลล์หนึ่งกับ dendrite หรือร่างกายของเซลล์อื่น (รูปที่ 3.28)

ข้าว. 3.28.
อย่างไรก็ตามมีตัวเลือกอื่น ๆ ทั้งหมด: dendro-dendritic, somatodendritic, axo-axonal และ synapses ประเภทอื่น ๆ โครงสร้างพิเศษของขั้วประสาทใน CYS แสดงให้เห็นถึงลักษณะเฉพาะของไซแนปส์ทางเคมี: การปรากฏตัวของถุงไซแนปติก, โซนแอคทีฟในพรีไซแนปทิกบัด และตัวรับโพสต์ซินแนปติกบนเยื่อหุ้มเซลล์เป้าหมาย ความแตกต่างคือขนาดที่เล็กของเซนทรัลไซแนปส์ ดังนั้นในระบบประสาทส่วนกลางในซินเซสเคมีในตา presynaptic จำนวนโซนที่ใช้งานไม่เกิน 10 และส่วนใหญ่จะลดลงเหลือ 1-2 นี่เป็นเพราะขนาดของ presynaptic buds ขนาดเล็ก (1–2 μm)
นอกจากไซแนปส์อย่างง่ายแล้ว ซึ่งประกอบด้วยปลายก่อนและหลังซินแนปติกหนึ่งจุด ไซแนปส์ที่ซับซ้อนยังมีอยู่ในระบบประสาทส่วนกลางด้วย พวกเขาแบ่งออกเป็นหลายกลุ่ม ในกลุ่มของไซแนปส์ที่ซับซ้อนกลุ่มหนึ่ง การสิ้นสุดของแอกซอนแบบพรีซินแนปติกจะก่อตัวเป็นกิ่งก้านสาขาหลายกิ่ง - เยื่อหุ้มเซลล์เจริญที่ลงท้ายด้วยตาขนาดเล็ก ด้วยความช่วยเหลือของพวกเขา แอกซอนจะติดต่อกับเดนไดรต์ของเซลล์ประสาทหลายเซลล์พร้อมกัน ในอีกกลุ่มหนึ่งของไซแนปส์ที่ซับซ้อน ปลายพรีซินแนปติกของแอกซอนต่างๆ จุดสิ้นสุดเหล่านี้ครอบคลุมโซนโพสต์ซินแน็ปติกอย่างใกล้ชิด - ส่วนหัวของกระดูกสันหลัง Synaptic glomeruli ซึ่งเป็นกลุ่มที่มีขนาดกะทัดรัดของกระบวนการของเซลล์ประสาทต่าง ๆ ซึ่งก่อตัวเป็นซินแนปส์ร่วมกันจำนวนมาก มีโครงสร้างที่ซับซ้อนมากยิ่งขึ้น โดยปกติแล้ว glomeruli ดังกล่าวจะถูกล้อมรอบด้วยเซลล์เกลีย (ดูรูปที่ 3.28)
ไซแนปส์ถือได้ว่าเป็นหน่วยการทำงานของเนื้อเยื่อประสาทซึ่งช่วยให้มั่นใจได้ถึงการส่งข้อมูลในระบบประสาท อย่างไรก็ตาม ปฏิสัมพันธ์ของไซแนปส์ที่ทำงานติดกันเป็นเงื่อนไขที่สำคัญเท่าเทียมกันสำหรับการประมวลผลข้อมูลในระบบประสาทส่วนกลาง การมีซินแนปส์ที่ซับซ้อน (โดยเฉพาะซินแนปติก โกลเมอรูลี) ที่ทำให้กระบวนการนี้มีประสิทธิภาพเป็นพิเศษ ดังนั้นจึงเป็นที่ชัดเจนว่าเหตุใดซินแนปส์ที่ซับซ้อนจำนวนมากที่สุดจึงอยู่ในบริเวณของสมองที่มีการประมวลผลสัญญาณที่ซับซ้อนที่สุด - ในเปลือกสมองของสมองส่วนหน้า, เปลือกสมองน้อยและฐานดอก
จำนวนของไซแนปส์บนเยื่อหุ้มเซลล์ประสาทส่วนกลางหนึ่งช่วงโดยเฉลี่ยตั้งแต่ 2-5,000 ถึง 15,000 หรือมากกว่านั้น ตำแหน่งของผู้ติดต่อนั้นแปรปรวนมาก ไซแนปส์มีอยู่ในร่างกายของเซลล์ประสาท เดนไดรต์ของมัน และในแอกซอนในระดับที่น้อยกว่า สิ่งที่สำคัญที่สุดสำหรับกิจกรรมของเซลล์ประสาทคือการสัมผัสกับโสม ฐานของเดนไดรต์ เช่นเดียวกับจุดที่แตกแขนงแรกของเดนไดรต์ ฟังก์ชันพรีซินแนปติกส่วนใหญ่มักดำเนินการโดยการแบ่งส่วนปลายของแอกซอน (ตาพรีซินแนนติก) หรือส่วนขยายขอดตามแอกซอน บ่อยครั้งที่กิ่ง dendritic บาง ๆ สามารถทำหน้าที่เป็นโครงสร้างที่ไม่ใช่ resynaptic ได้
ดังที่เราได้กล่าวไปแล้ว ศักยภาพของโพสต์ซินแนปติกในซิงก์เคมีสามารถเป็นได้ทั้งแบบดีโพลาไรซ์และแบบกระตุ้น (VISI) หรือแบบไฮเปอร์โพลาไรซ์และแบบยับยั้ง (TPSP)
- การแนะนำ
- การจำแนกประเภทของไซแนปส์
- สมมติฐาน porocytosis
- บทสรุป
- บรรณานุกรม
การแนะนำ
จนถึงปัจจุบันมีการสร้างเทคโนโลยีจำนวนมากสำหรับการฝังอวัยวะเทียมต่าง ๆ ซึ่งร่างกายไม่ได้ปฏิเสธเป็นเวลานาน ปัญหาหนึ่งที่ขัดขวางการพัฒนาอุตสาหกรรมนี้คือการบูรณาการระบบประสาทและอุปกรณ์ไซเบอร์เนติกส์ พูดง่ายๆ ก็คือ ในการสร้างการเชื่อมต่อระหว่างเส้นประสาทกับตัวประมวลผลของอวัยวะเทียม
ทางออกของปัญหานี้ - สง่างามและไม่ต้องการการนำอิเล็กโทรดเข้าสู่เนื้อเยื่อประสาทอย่างคร่าวๆ - คือการสร้างการเชื่อมต่อแบบซินแนปติก ไซแนปส์ - ผลิตภัณฑ์จากธรรมชาติ - เป็นรูปแบบในอุดมคติของการรวมการทำงานของปลายประสาทและอวัยวะเอฟเฟกต์ต่าง ๆ (กล้ามเนื้อ, เนื้อเยื่อหลั่ง)
ในการทำเช่นนี้จำเป็นต้องศึกษาโครงสร้างและสรีรวิทยาของไซแนปส์ต่างๆ
ไซแนปส์ กระแสประสาท สรีรวิทยา
ข้อกำหนดทั่วไปและประวัติการค้นพบ
ไซแนปส์เป็นจุดสัมผัสระหว่างเซลล์ประสาท 2 เซลล์หรือระหว่างเซลล์ประสาทกับเซลล์รับเอฟเฟกต์ มันทำหน้าที่ส่งกระแสประสาทระหว่างเซลล์สองเซลล์ และระหว่างการส่งซินแนปติก แอมพลิจูดและความถี่ของสัญญาณจะถูกควบคุม การส่งแรงกระตุ้นจะดำเนินการทางเคมีด้วยความช่วยเหลือของผู้ไกล่เกลี่ยหรือทางไฟฟ้าผ่านทางไอออนจากเซลล์หนึ่งไปยังอีกเซลล์หนึ่ง ตามกฎแล้ว ไซแนปส์ถูกเข้าใจว่าเป็นไซแนปส์ทางเคมีซึ่งส่งสัญญาณโดยใช้สารสื่อประสาท ไซแนปส์โดยทั่วไปคือรูปแบบที่เกิดจากปลายแอกซอนของเซลล์ประสาทหนึ่งและเดนไดรต์ของอีกเซลล์หนึ่ง (แอกโซ-เดนไดรติกซินแนปส์) แต่มีประเภทอื่น: axosomatic, axo-axonal และ dendro-dendritic ไซแนปส์ระหว่างแอกซอนของเซลล์ประสาทสั่งการและใยกล้ามเนื้อโครงร่างเรียกว่า motor end plate หรือทางแยกประสาทและกล้ามเนื้อ ไซแนปส์ในระบบประสาทมีอยู่สองประเภท: ไซแนปส์แบบกระตุ้นและแบบยับยั้ง ในซิงก์กระตุ้นเซลล์หนึ่งทำให้เกิดการกระตุ้นอีกเซลล์หนึ่ง ในกรณีนี้ผู้ไกล่เกลี่ย excitatory ทำให้เกิดการสลับขั้ว - การไหลของไอออน Na + พุ่งเข้าสู่เซลล์ ในการยับยั้งไซแนปส์ เซลล์หนึ่งจะยับยั้งการกระตุ้นของอีกเซลล์หนึ่ง นี่เป็นเพราะตัวกลางที่เป็นตัวยับยั้งทำให้เกิดการไหลของไอออนที่มีประจุลบเข้าไปในเซลล์ ดังนั้นจึงไม่เกิดการสลับขั้ว
แรงกระตุ้นของเส้นประสาทเข้าสู่ไซแนปส์ผ่านทางส่วนปลายของพรีซินแนปติก ซึ่งถูกจำกัดโดยเยื่อหุ้มพรีไซแนปติก (ส่วนพรีไซแนปติก) และรับรู้โดยเยื่อหุ้มโพสซินแนปติก (ส่วนโพสต์ซินแนปติก) รอยแยก synaptic ตั้งอยู่ระหว่างเยื่อหุ้ม ส่วนปลายของพรีไซแนปติกประกอบด้วยไมโทคอนเดรียและถุงพรีไซแนปติกจำนวนมากที่มีสารสื่อประสาท แรงกระตุ้นของเส้นประสาทที่เข้าสู่ปลาย presynaptic ทำให้สื่อกลางถูกปล่อยเข้าไปในรอยแหว่งของ synaptic โมเลกุลของตัวกลางทำปฏิกิริยากับโปรตีนตัวรับเฉพาะของเยื่อหุ้มเซลล์ เปลี่ยนการซึมผ่านของไอออนบางชนิด ซึ่งนำไปสู่การเกิดขึ้นของศักยภาพในการดำเนินการ นอกเหนือจากการประสานทางเคมีแล้ว ยังมีการเชื่อมประสานด้วยไฟฟ้า ซึ่งการส่งผ่านของแรงกระตุ้นเกิดขึ้นโดยตรงในลักษณะของไฟฟ้าชีวภาพระหว่างเซลล์ที่สัมผัสกัน
ขึ้นอยู่กับลักษณะของสัญญาณที่ผ่านไซแนปส์ ไซแนปส์จะแบ่งออกเป็นไซแนปส์ไฟฟ้าและซินแนปส์เคมี สารเคมีซินแนปส์เป็นซินเซสที่ส่งสารด้วยความช่วยเหลือของสารออกฤทธิ์ทางชีวภาพและสารที่ส่งสารคือสารสื่อประสาท
· ในปี พ.ศ. 2440 เชอร์ริงตันได้กำหนดแนวคิดของไซแนปส์
· สำหรับการศึกษาระบบประสาท รวมทั้งการส่งซินแนปติก ในปี พ.ศ. 2449 Golgi และ Ramon y Cajal ได้รับรางวัลโนเบล
· ในปี พ.ศ. 2464 นักวิทยาศาสตร์ชาวออสเตรีย โอ. โลวี (O. Loewi) ได้กำหนดลักษณะทางเคมีของการส่งผ่านการกระตุ้นผ่านไซแนปส์และบทบาทของอะซิติลโคลีนในนั้น ได้รับรางวัลโนเบลในปี พ.ศ. 2479 ร่วมกับ จี. เดล (N. Dale)
· ในปี 1933 นักวิทยาศาสตร์โซเวียต A.V. Kibyakov สร้างบทบาทของอะดรีนาลีนในการส่งสัญญาณแบบซินแนปติก
· 1970 - B. Katz (V. Katz, UK), U. von Euler (U. v. Euler, สวีเดน) และ J. Axelrod (J. Axelrod, USA) ได้รับรางวัลโนเบลจากการค้นพบบทบาทของ norepinephrine ใน synaptic การแพร่เชื้อ.
การจำแนกประเภทของไซแนปส์
ตามกลไกการส่งกระแสประสาท:
สารเคมี - นี่คือสถานที่สัมผัสใกล้ชิดระหว่างเซลล์ประสาทสองเซลล์สำหรับการส่งกระแสประสาทซึ่งเซลล์ต้นทางปล่อยสารพิเศษซึ่งเป็นสารสื่อประสาทเข้าไปในช่องว่างระหว่างเซลล์ซึ่งการปรากฏตัวของรอยแยก synaptic นั้นกระตุ้นหรือยับยั้ง เซลล์รับ
ไฟฟ้า (ephaps) - สถานที่แนบสนิทของเซลล์คู่หนึ่งซึ่งเยื่อหุ้มเซลล์เชื่อมต่อกันโดยใช้การก่อตัวของโปรตีนพิเศษ - connexons (แต่ละ connexon ประกอบด้วยหน่วยย่อยโปรตีนหกหน่วย) ระยะห่างระหว่างเยื่อหุ้มเซลล์ในไซแนปส์ไฟฟ้าคือ 3.5 นาโนเมตร (ระหว่างเซลล์ปกติคือ 20 นาโนเมตร) เนื่องจากความต้านทานของของเหลวนอกเซลล์มีขนาดเล็ก (ในกรณีนี้) แรงกระตุ้นจะผ่านไซแนปส์โดยไม่ชักช้า ประสาทไฟฟ้ามักจะถูกกระตุ้น
ไซแนปส์แบบผสม - ศักยภาพของการกระทำของพรีซินแนปติกสร้างกระแสที่สลับขั้วของเยื่อหุ้มโพสซินแนปติกของไซแนปส์ทางเคมีทั่วไป โดยที่เมมเบรนก่อนและหลังซินแนปติกไม่พอดีกัน ดังนั้น ในไซแนปส์เหล่านี้ การส่งผ่านสารเคมีจึงทำหน้าที่เป็นกลไกเสริมแรงที่จำเป็น
สารเคมีที่พบบ่อยที่สุด สำหรับระบบประสาทของสัตว์เลี้ยงลูกด้วยนม ไซแนปส์ไฟฟ้ามีลักษณะเฉพาะน้อยกว่าสารเคมี
โดย ที่ตั้ง และ เครื่องประดับ โครงสร้าง :
อุปกรณ์ต่อพ่วง
ประสาทและกล้ามเนื้อ
ระบบประสาท (axo-vasal)
ตัวรับ-เซลล์ประสาท
ศูนย์กลาง
axo-dendritic - กับ dendrites รวมถึง
axo-spiky - มีเงี่ยง dendritic ผลพลอยได้จาก dendrites;
axo-somatic - กับร่างกายของเซลล์ประสาท
แอกโซแอกซอน - ระหว่างแอกซอน
dendro-dendritic - ระหว่าง dendrites;
โดย สารสื่อประสาท :
· aminergic ที่มี biogenic amines (เช่น serotonin, dopamine);
รวมถึง adrenergic ที่มี adrenaline หรือ norepinephrine
cholinergic ที่มี acetylcholine;
Purinergic ที่มีพิวรีน;
peptidergic ที่มีเปปไทด์
ในเวลาเดียวกัน คนกลางเพียงคนเดียวไม่ได้ถูกสร้างในไซแนปส์เสมอไป โดยปกติแล้วตัวไกล่เกลี่ยหลักจะถูกดีดออกพร้อมกับอีกตัวซึ่งทำหน้าที่เป็นโมดูเลเตอร์
ตามสัญญาณของการกระทำ
น่าตื่นเต้น
เบรค.
หากอดีตนำไปสู่การเกิดขึ้นของการกระตุ้นในเซลล์โพสต์ซินแน็ปติก (อันเป็นผลมาจากการได้รับแรงกระตุ้น เมมเบรนจะสลับขั้วในเซลล์ซึ่งอาจทำให้เกิดการกระทำภายใต้เงื่อนไขบางประการ) จากนั้นในทางกลับกัน หยุดหรือป้องกันการเกิดขึ้น ป้องกันการแพร่กระจายของแรงกระตุ้นต่อไป โดยปกติแล้วการยับยั้งคือ glycinergic (ตัวกลาง - ไกลซีน) และ GABAergic synapses (ตัวกลาง - กรดแกมมา - อะมิโนบิวทีริก)
ไซแนปส์ยับยั้งมีสองประเภท:
1) ไซแนปส์ที่ส่วนท้ายของพรีซินแนปติกซึ่งตัวไกล่เกลี่ยถูกปล่อยออกมา โพลาไรซ์โพลาไรเซชันของเมมเบรนโพสต์ซินแนปติกและก่อให้เกิดการยับยั้งศักยภาพของโพสต์ซินแนปติก
2) axo-axonal synapse ให้การยับยั้ง presynaptic Cholinergic synapse (s. cholinergica) - ไซแนปส์ที่ acetylcholine เป็นตัวกลาง
ในบางไซแนปส์มีการบดอัดแบบโพสต์ซินแนปติก - โซนที่มีอิเล็กตรอนหนาแน่นซึ่งประกอบด้วยโปรตีน ตามการมีหรือไม่มี synapses แบบอสมมาตรและสมมาตรนั้นแตกต่างกัน เป็นที่ทราบกันว่าซินแนปส์ของกลูตามาเทอจิคทั้งหมดนั้นไม่สมมาตร ในขณะที่ซินแนปส์ของ GABAergic นั้นมีความสมมาตร
ในกรณีที่ส่วนขยายของซินแนปติกหลายอันสัมผัสกับเยื่อหุ้มโพสซินแนปติก จะเกิดซินแนปส์หลายอัน
อุปกรณ์ที่มีหนามซึ่งส่วนที่ยื่นออกมาสั้น ๆ เดี่ยวหรือหลายอันของเยื่อโพสต์ซินแนปติกของเดนไดรต์สัมผัสกับการขยายตัวของไซแนปติก เป็นรูปแบบพิเศษของซินแนปส์ เครื่องมือ Spiny เพิ่มจำนวนการติดต่อของ synaptic บนเซลล์ประสาทอย่างมีนัยสำคัญ และเป็นผลให้จำนวนข้อมูลที่ประมวลผล ไซแนปส์ที่ "ไม่มีกระดูกสันหลัง" เรียกว่า "นั่ง" ตัวอย่างเช่น GABAergic synapses ทั้งหมดเป็นแบบเซสไซล์
โครงสร้างของไซแนปส์เคมี
ไซแนปส์ส่วนใหญ่ในระบบประสาทของอาณาจักรสัตว์คือไซแนปส์ทางเคมี ลักษณะเหล่านี้มีลักษณะเฉพาะโดยมีลักษณะทั่วไปหลายประการ แม้ว่าอย่างไรก็ตาม ขนาดและรูปร่างของส่วนประกอบก่อนและหลังซินแน็ปติกจะแตกต่างกันอย่างมาก ไซแนปส์ในเปลือกสมองของสัตว์เลี้ยงลูกด้วยนมมีแอกซอนส่วนปลายประมาณ 100 นาโนเมตรและพรีซินแนปติกบัดซึ่งมีเส้นผ่านศูนย์กลางเฉลี่ยประมาณ 1 ไมโครเมตร
ไซแนปส์ทางเคมีประกอบด้วยสองส่วน: พรีซินแนปติกซึ่งเกิดจากส่วนต่อท้ายของแอกซอนของเซลล์ที่มีรูปร่างคล้ายสโมสรและโพสต์ซินแนปติกซึ่งแสดงโดยพื้นที่สัมผัสของพลาสมาเมมเบรนของเซลล์รับ ระหว่างทั้งสองส่วนมีช่องว่าง synaptic - ช่องว่างกว้าง 10-50 นาโนเมตรระหว่างเยื่อหุ้มเซลล์แบบโพสต์ซินแน็ปติกและพรีซินแนปติกซึ่งขอบเสริมด้วยการสัมผัสระหว่างเซลล์
ส่วนของ axolemma ของส่วนต่อขยายรูปกระบองที่อยู่ติดกับ synaptic cleft เรียกว่า presynaptic membrane ส่วนของไซโตเลมมาของเซลล์รับความรู้สึกซึ่งจำกัดรอยแยกไซแนปติกในด้านตรงข้าม เรียกว่า เมมเบรนโพสซินแนปติก (postynaptic membrane) ในทางเคมี ไซแนปส์เป็นส่วนบรรเทาและมีตัวรับจำนวนมาก
ในการขยายตัวของ synaptic มีถุงเล็ก ๆ ที่เรียกว่า presynaptic vesicles หรือ synaptic vesicles ซึ่งมีตัวกลาง (ตัวกลางในการส่งผ่านของการกระตุ้น) หรือเอนไซม์ที่ทำลายตัวกลางนี้ บนโพสต์ซินแน็ปทิกและบ่อยครั้งบนเยื่อหุ้มพรีซินแนปติก มีตัวรับสำหรับตัวกลางหนึ่งตัวหรืออีกตัวหนึ่ง
ขนาดเท่ากันของ presynaptic vesicles ใน synapses ที่ศึกษาทั้งหมด (40-50 นาโนเมตร) ได้รับการพิจารณาเป็นครั้งแรกว่าเป็นหลักฐานว่า vesicle แต่ละอันเป็นกลุ่มขั้นต่ำซึ่งจำเป็นต้องมีการปลดปล่อยเพื่อสร้างสัญญาณ synaptic ถุงน้ำตั้งอยู่ตรงข้ามเยื่อ presynaptic ซึ่งเกิดจากวัตถุประสงค์ในการทำงานเพื่อปลดปล่อยสื่อกลางเข้าไปในรอยแหว่ง synaptic นอกจากนี้ ใกล้ presynaptic vesicle ยังมีไมโตคอนเดรียจำนวนมาก (ผลิต adenosine triphosphate) และโครงสร้างเส้นใยโปรตีนตามลำดับ
รอยแหว่งซินแนปติกคือช่องว่างกว้าง 20 ถึง 30 นาโนเมตรระหว่างพรีซินแนปติกเวสิเคิลและเมมเบรนโพสซินแนปส์ ซึ่งมีโครงสร้างการจับก่อนและหลังไซแนปส์ที่สร้างจากโปรตีโอไกลแคน ความกว้างของ synaptic gap ในแต่ละกรณีนั้นเกิดจากการที่สื่อกลางที่ดึงออกมาจาก presynapse จะต้องผ่านไปยัง postsynapse ในเวลาที่น้อยกว่าความถี่ของสัญญาณประสาทที่มีลักษณะเฉพาะของเซลล์ประสาทที่สร้าง synapse อย่างมีนัยสำคัญ (เวลาที่มัน ใช้เวลาไกล่เกลี่ยเพื่อส่งผ่านจาก pre-ไปยัง postsynaptic membrane ตามลำดับของไมโครวินาที)
เมมเบรนโพสซินแนปติกเป็นของเซลล์ที่รับกระแสประสาท กลไกของการแปลสัญญาณทางเคมีของผู้ไกล่เกลี่ยเป็นศักย์ไฟฟ้าในเซลล์นี้คือตัวรับ - โปรตีนโมเลกุลใหญ่ที่ฝังอยู่ในเยื่อหุ้มโพสต์ซินแนปติก ด้วยความช่วยเหลือของเทคนิค ultramicroscopic พิเศษทำให้ได้รับข้อมูลจำนวนมากพอสมควรเกี่ยวกับโครงสร้างโดยละเอียดของ synapses ในช่วงไม่กี่ปีที่ผ่านมา
ดังนั้นโครงสร้างที่ได้รับคำสั่งของการกดทับเหมือนปล่องภูเขาไฟขนาดเส้นผ่านศูนย์กลาง 10 นาโนเมตรที่กดเข้าด้านในจึงถูกค้นพบบนเยื่อหุ้มพรีซินแนปติก ในตอนแรกพวกมันถูกเรียกว่าซินแนปโทพอร์ แต่ตอนนี้โครงสร้างเหล่านี้ถูกเรียกว่าเวสิเคิลไฟล์แนบไซต์ (VSPs) เต้ารับถูกจัดเรียงเป็นกลุ่มตามคำสั่งของหกช่องแยกจากกันรอบๆ สิ่งที่เรียกว่าส่วนที่ยื่นออกมาอัดแน่น ดังนั้น ส่วนที่ยื่นออกมาหนาแน่นจึงก่อตัวเป็นโครงสร้างสามเหลี่ยมปกติที่ด้านในของเยื่อหุ้มเซลล์ก่อนไซแนปติก และ SSV เป็นรูปหกเหลี่ยม และเป็นบริเวณที่ถุงเปิดออกและขับสารสื่อประสาทเข้าไปในรอยแยกไซแนปติก
โครงสร้างของไซแนปส์ไฟฟ้า
ซึ่งแตกต่างจากไซแนปส์ทางเคมี รอยแยกไซแนปติกในไซแนปส์ไฟฟ้านั้นแคบมาก (ประมาณ 3.5 นาโนเมตร) ผ่านรอยแยกไซแนปติกของไซแนปส์ประเภทนี้ ช่องโปรตีนที่สั่งเชิงพื้นที่พร้อมรูพรุนที่ชอบน้ำ แต่ละช่องมีเส้นผ่านศูนย์กลางประมาณ 5 นาโนเมตร ซึ่งเจาะเยื่อหุ้มเซลล์ก่อนและหลังไซแนปส์ และเรียกว่าคอนเนกซอน ใน protostomes (ไส้เดือนฝอย, หอย, สัตว์ขาปล้อง), connexons เกิดจากโปรตีน pannexins หรือ innexins); ใน deuterostomes (ascidians สัตว์มีกระดูกสันหลัง) connexons ถูกสร้างขึ้นจากโปรตีนประเภทต่างๆ - connexins ซึ่งเข้ารหัสโดยยีนกลุ่มต่างๆ ยังไม่พบ pannexins และ connexins ใน echinoderms; พวกเขาอาจมีโปรตีนอีกตระกูลหนึ่งที่สร้างจุดเชื่อมต่อช่องว่างและไซแนปส์ไฟฟ้า
สัตว์มีกระดูกสันหลังมีทั้งแพนเนกซินและคอนเนกซิน แต่จนถึงตอนนี้ ยังไม่มีการระบุไซแนปส์ไฟฟ้าในสัตว์มีกระดูกสันหลัง ซึ่งช่องทางระหว่างเซลล์จะเกิดขึ้นจากแพนเนกซิน
ไอออนและโมเลกุลขนาดเล็ก รวมทั้งสีย้อมเรืองแสงที่นำเข้าเซลล์เทียม จะผ่านคอนเนกซิน (หรือแพนเนกซิน) ที่เชื่อมต่อเซลล์ประสาททั้งก่อนและหลังซินแน็ปติก การผ่านของสีย้อมเหล่านี้ผ่านไซแนปส์ไฟฟ้าสามารถบันทึกได้แม้จะใช้กล้องจุลทรรศน์แบบใช้แสง
ไซแนปส์ไฟฟ้ายอมให้นำไฟฟ้าได้ทั้งสองทิศทาง (ตรงข้ามกับสารเคมี); อย่างไรก็ตาม เมื่อเร็ว ๆ นี้ ได้มีการค้นพบการแก้ไขไซแนปส์ไฟฟ้า ซึ่งก็คือการส่งสัญญาณประสาทในทิศทางเดียวในสัตว์จำพวกครัสเตเชียบางชนิด
กลไกการส่งกระแสประสาท
การมาถึงของแรงกระตุ้นทางไฟฟ้าไปยังเยื่อพรีไซแนปส์จะกระตุ้นกระบวนการส่งสัญญาณไซแนปติก ซึ่งขั้นตอนแรกคือการที่ไอออน Ca 2+ เข้าสู่พรีไซแนปส์ผ่านเยื่อหุ้มเซลล์ผ่านช่องแคลเซียมเฉพาะที่อยู่ใกล้กับรอยแหว่งไซแนปติก ไอออน Ca 2+ โดยกลไกที่ไม่รู้จักโดยสิ้นเชิง กระตุ้นถุงน้ำที่อัดแน่นที่บริเวณจุดยึด และปล่อยสารสื่อประสาทเข้าไปในรอยแยกไซแนปติก ไอออน Ca 2+ ที่เข้าสู่เซลล์ประสาท หลังจากเปิดใช้งานถุงด้วยสื่อกลาง จะถูกปิดใช้งานในเวลาหลายไมโครวินาทีเนื่องจากการทับถมในไมโตคอนเดรียและถุงพรีไซแนปส์
โมเลกุลสื่อกลางที่ปล่อยออกมาจากพรีไซแนปส์จับกับตัวรับบนเยื่อหุ้มเซลล์หลังไซแนปส์ ซึ่งเป็นผลมาจากการที่ช่องไอออนเปิดในโมเลกุลขนาดใหญ่ของตัวรับ (ในกรณีของตัวรับช่องสัญญาณ ซึ่งเป็นชนิดที่พบบ่อยที่สุด เมื่อตัวรับชนิดอื่นทำงาน กลไกการส่งสัญญาณจะต่างกัน) ไอออนที่เริ่มเข้าสู่เซลล์โพสต์ซินแนปติกผ่านช่องเปิดจะเปลี่ยนประจุของเยื่อหุ้มเซลล์ ซึ่งเป็นโพลาไรเซชันบางส่วน (ในกรณีของไซแนปส์แบบยับยั้ง) หรือดีโพลาไรเซชัน (ในกรณีของไซแนปส์แบบกระตุ้น) ของเมมเบรนนี้ และในฐานะ ผลลัพธ์นำไปสู่การยับยั้งหรือกระตุ้นการสร้างโดยศักยภาพในการดำเนินการของเซลล์โพสต์ซินแนปติก
สมมติฐานควอนตัม-vesicular
เป็นที่นิยมจนกระทั่งเมื่อเร็ว ๆ นี้ในฐานะคำอธิบายสำหรับกลไกการปลดปล่อยสารสื่อประสาทจากพรีไซแนปส์ สมมติฐานของควอนตัม-เวสซิคูลาร์ เอ็กโซไซโทซิส (QVE) บอกเป็นนัยว่า "แพ็คเกจ" หรือควอนตัมของผู้ไกล่เกลี่ยมีอยู่ในถุงเดียวและถูกปล่อยออกมาระหว่างเอ็กโซไซโทซิส ( ในกรณีนี้ เยื่อหุ้มเซลล์จะหลอมรวมเข้ากับเยื่อหุ้มพรีไซแนปติกของเซลล์ ) ทฤษฎีนี้เป็นสมมติฐานที่แพร่หลายมาเป็นเวลานาน แม้ว่าข้อเท็จจริงที่ว่าไม่มีความสัมพันธ์กันระหว่างระดับของการปล่อยสารสื่อประสาท (หรือศักยภาพของโพสต์ซินแนปติก) และจำนวนของถุงน้ำในพรีไซแนปส์ นอกจากนี้ สมมติฐาน CBE ยังมีข้อบกพร่องที่สำคัญอื่นๆ
พื้นฐานทางสรีรวิทยาของการปล่อยสารไกล่เกลี่ยอย่างแม่นยำควรมีปริมาณเท่ากันในแต่ละถุง สมมติฐาน TBE ในรูปแบบคลาสสิกไม่เหมาะสำหรับการอธิบายผลกระทบของควอนตัมที่มีขนาดต่างกัน (หรือปริมาณที่แตกต่างกันของผู้ไกล่เกลี่ย) ที่สามารถปล่อยออกมาได้ในระหว่างการกระตุ้นเอกโซไซโทซิสเพียงครั้งเดียว ในกรณีนี้ เราต้องคำนึงถึงว่าสามารถสังเกตตุ่มขนาดต่างๆ กันในตา presynaptic เดียวกันได้ นอกจากนี้ยังไม่พบความสัมพันธ์ระหว่างขนาดของถุงน้ำและปริมาณของผู้ไกล่เกลี่ยในนั้น (นั่นคือความเข้มข้นในถุงอาจแตกต่างกันด้วย) ยิ่งไปกว่านั้น ในไซแนปส์ของประสาทและกล้ามเนื้อที่ถูกทำลาย เซลล์ Schwann สร้างศักยภาพของโพสต์ซินแนปติกขนาดเล็กจำนวนมากขึ้นกว่าที่สังเกตได้ในไซแนปส์ก่อนการปฏิเสธ แม้ว่าจะไม่มีถุงพรีไซแนปติกที่แปลเป็นภาษาท้องถิ่นในพื้นที่ของปุ่มพรีซินแนปติกในเซลล์เหล่านี้
สมมติฐาน porocytosis
มีหลักฐานการทดลองที่สำคัญว่าสารสื่อประสาทถูกหลั่งเข้าไปในรอยแหว่งไซแนปติกเนื่องจากการเปิดใช้งานแบบซิงโครนัสของกลุ่มหกเหลี่ยมของ MPV (ดูด้านบน) และถุงที่ติดอยู่กับพวกมัน ซึ่งกลายเป็นพื้นฐานสำหรับการกำหนดสมมติฐาน porocytosis สมมติฐานนี้อยู่บนพื้นฐานของการสังเกตว่าถุงเล็ก ๆ ที่ติดอยู่กับ SSV เมื่อได้รับศักยภาพในการดำเนินการ สัญญาพร้อมกันและในเวลาเดียวกันจะหลั่งสารสื่อกลางในปริมาณที่เท่ากันเข้าไปในรอยแหว่งซินแนปติกในแต่ละครั้ง โดยปล่อยเนื้อหาเพียงบางส่วนจาก แต่ละถุงหก คำว่า "porocytosis" มาจากคำภาษากรีก poro (หมายถึงรูขุมขน) และ cytosis (อธิบายถึงการขนส่งสารเคมีผ่านพลาสมาเมมเบรนของเซลล์)
ข้อมูลการทดลองส่วนใหญ่เกี่ยวกับการทำงานของจุดเชื่อมต่อระหว่างเซลล์แบบโมโนซินแนปติกได้มาจากการศึกษาของจุดเชื่อมต่อประสาทและกล้ามเนื้อที่แยกได้ ในประสาทและกล้ามเนื้อประสาทของ SSV โครงสร้างหกเหลี่ยมตามลำดับจะเกิดขึ้น โครงสร้างหกเหลี่ยมเหล่านี้แต่ละโครงสร้างสามารถกำหนดเป็น "ซินแนปโตเมอร์" นั่นคือ โครงสร้างที่เป็นหน่วยพื้นฐานในกระบวนการหลั่งสื่อกลาง ซินแนปโทเมอร์ประกอบด้วย นอกเหนือไปจากรูพรุนจริง โครงสร้างเส้นใยโปรตีนที่มีถุงเรียงตามลำดับเชิงเส้น การมีอยู่ของโครงสร้างที่คล้ายคลึงกันยังได้รับการพิสูจน์สำหรับไซแนปส์ในระบบประสาทส่วนกลาง (CNS)
ดังที่กล่าวไว้ข้างต้น กลไกของ porocytic สร้างสารสื่อประสาทควอนตัม แต่ไม่มีเยื่อหุ้มของถุงน้ำแต่ละอันที่ผสานเข้ากับเยื่อหุ้มพรีไซแนปติกอย่างสมบูรณ์ ค่าสัมประสิทธิ์การแปรผันเล็กน้อย (น้อยกว่า 3%) สำหรับค่าของศักยภาพโพสต์ซินแนปติกเป็นตัวบ่งชี้ว่าในไซแนปส์เดียวมีซินแนปโตเมอร์ไม่เกิน 200 ตัว ซึ่งแต่ละตัวจะหลั่งควอนตัมตัวส่งสัญญาณหนึ่งตัวเพื่อตอบสนองต่อศักยภาพการกระทำหนึ่งตัว ตำแหน่งการปลดปล่อย 200 ตำแหน่ง (นั่นคือ synaptomers ที่ปล่อยสารสื่อประสาท) ที่พบในเส้นใยกล้ามเนื้อขนาดเล็กทำให้สามารถคำนวณขีดจำกัดควอนตัมสูงสุดที่หนึ่งตำแหน่งการปลดปล่อยต่อไมโครเมตรของความยาวทางแยก synaptic การสังเกตนี้ตัดความเป็นไปได้ของควอนตัมสารสื่อประสาทที่เกิดขึ้นใน ปริมาณหนึ่งถุง
การเปรียบเทียบสมมติฐานของ porocytosis และ quantum-vesicular
การเปรียบเทียบสมมติฐาน TBE ที่ยอมรับเมื่อเร็วๆ นี้กับสมมติฐานของ porocytosis สามารถดำเนินการได้โดยการเปรียบเทียบค่าสัมประสิทธิ์การแปรผันทางทฤษฎีกับการทดลองที่คำนวณสำหรับแอมพลิจูดของศักย์ไฟฟ้าหลังซินแน็ปติกที่สร้างขึ้นเพื่อตอบสนองต่อการปล่อยสารสื่อประสาทแต่ละตัวจากพรีไซแนปส์ สมมติว่ากระบวนการ exocytosis เกิดขึ้นในไซแนปส์ขนาดเล็กที่มีถุงประมาณ 5,000 ถุง (50 ถุงต่อความยาวไซแนปส์แต่ละไมครอน) ควรสร้างศักยภาพหลังการทำซินแนปส์โดยถุงสุ่ม 50 ถุงที่สุ่มเลือก ซึ่งให้ค่าสัมประสิทธิ์การแปรผันทางทฤษฎีที่ 14% ค่านี้มีค่ามากกว่าค่าสัมประสิทธิ์การแปรผันของศักยภาพโพสต์ซินแนปติกที่ได้รับจากการทดลองประมาณ 5 เท่า ดังนั้นจึงอาจโต้แย้งได้ว่ากระบวนการเอ็กโซไซโทซิสในไซแนปส์นั้นไม่ได้สุ่ม (ไม่ตรงกับการกระจายแบบปัวซอง) ซึ่งเป็นไปไม่ได้หาก อธิบายในแง่ของสมมติฐาน TBE แต่สอดคล้องกับสมมติฐาน porocytosis ความจริงก็คือสมมติฐานของ porocytosis สันนิษฐานว่าถุงทั้งหมดที่เกี่ยวข้องกับ presynaptic membrane ขับตัวกลางออกมาพร้อมกัน ในเวลาเดียวกัน จำนวนสื่อกลางที่คงที่ถูกขับเข้าไปในรอยแยกซินแนปติกเพื่อตอบสนองต่อศักยภาพของการกระทำแต่ละอย่าง (ความยั่งยืนเห็นได้จากค่าสัมประสิทธิ์การแปรผันของการตอบสนองหลังซินแนปติกที่ต่ำ) อาจอธิบายได้ด้วยการเปิดตัวสื่อกลางปริมาณเล็กน้อยโดย ถุงจำนวนมาก - ในขณะเดียวกันยิ่งถุงมีส่วนร่วมในกระบวนการมากขึ้น ค่าสัมประสิทธิ์สหสัมพันธ์ก็จะยิ่งน้อยลง แม้ว่าสิ่งนี้จะดูค่อนข้างขัดแย้งจากมุมมองของสถิติทางคณิตศาสตร์
สิ่งที่เรียกว่า "หลักการเดล" (หนึ่งเซลล์ประสาท - หนึ่งสื่อกลาง) นั้นถือว่าผิดพลาด หรือตามที่เชื่อกันในบางครั้ง มันถูกขัดเกลา: ไม่ใช่หนึ่งเดียว แต่สามารถปล่อยตัวกลางหลายตัวออกจากปลายด้านหนึ่งของเซลล์ และชุดของมันจะคงที่สำหรับเซลล์หนึ่งๆ
บทสรุป
ดังนั้นจึงมีการพิจารณาคำถามเกี่ยวกับโครงสร้างและหลักการทำงานของไซแนปส์เคมี และแม้ว่าจะยังมีคำถามที่ต้องการความชัดเจน อย่างไรก็ตาม ความรู้เกี่ยวกับการเชื่อมต่อซินแนปติกระหว่างเนื้อเยื่อประสาทถือเป็นก้าวสำคัญในด้านประสาทวิทยาศาสตร์ เขาคือผู้ที่ยอมทำสิ่งที่แทบจะเป็นไปไม่ได้ - การดำเนินการเพื่อฟื้นฟูกิจกรรมทางประสาท การรวมเครื่องจักรและเนื้อเยื่อที่มีชีวิตเข้าด้วยกันอย่างดีที่สุด และในอนาคต - การอยู่ร่วมกันอย่างแท้จริงของสิ่งมีชีวิตและธรรมชาติประดิษฐ์ที่มนุษย์สร้างขึ้น
บรรณานุกรม
1. ซาเวลิเยฟ ก.ใน.แหล่งที่มาของความแปรปรวนในคุณสมบัติไดนามิกของระบบประสาทในระดับซินแนปติก // เทียม ปัญญา. - NAS ของยูเครน, โดเนตสค์, 2549 - ฉบับที่ 4 - P.323-338
2. ซาเวลิเยฟ ก.ใน.วิธีการจัดระเบียบตนเองของซินแนปติกและปัญหาของซินแนปส์ส่วนปลายของเซลล์ประสาท // นิตยสาร ปัญหา วิวัฒนาการ เปิด ระบบ. - คาซัคสถาน อัลมาตี 2549 - V.8 - ฉบับที่ 2. - S.96-104.
3. เอ็กเซิลส์ ง.ถึง.สรีรวิทยาของไซแนปส์ - ม.: มีร์, 2509. - 397 น.
เอกสารที่คล้ายกัน
สรีรวิทยาของเซลล์ประสาทและโครงสร้าง ห่างไกล ประชิดตัว และการติดต่อสัมพันธ์กัน สาระสำคัญของกลไกการส่งแรงกระตุ้นไฟฟ้าจากเซลล์ประสาทหนึ่งไปยังอีกเซลล์หนึ่งผ่านไซแนปส์ทางเคมี ปัจจัยหลักที่ทำหน้าที่ไกล่เกลี่ย
ภาคนิพนธ์ เพิ่ม 02/10/2558
ความหงุดหงิดเป็นคุณสมบัติหลักของเซลล์ที่มีชีวิต สรีรวิทยาของเซลล์ที่ถูกกระตุ้น โครงสร้างและคุณสมบัติพื้นฐานของเยื่อหุ้มเซลล์และช่องไอออน สรีรวิทยาของเนื้อเยื่อประสาทและไซแนปส์ การจำแนกประเภทของยา antiadrenergic กลไกการออกฤทธิ์
ภาคนิพนธ์ เพิ่ม 03/02/2014
กลไกการส่งผ่านการกระตุ้นในไซแนปส์ทางเคมี คุณสมบัติของโครงสร้าง ประเภทและคุณสมบัติของคนกลาง ไซแนปส์ไฟฟ้าและการยับยั้ง คุณลักษณะของการส่งสัญญาณ วิธีการควบคุมทางเภสัชวิทยาของการกระตุ้น synaptic
งานนำเสนอ เพิ่ม 12/09/2014
เซลล์ประสาทเป็นหน่วยโครงสร้างและการทำงานของระบบประสาท ลักษณะโครงสร้างของเซลล์นี้ การทำงานและความเชี่ยวชาญเฉพาะทาง การก่อตัวของปลอกไมอีลิน เส้นใยที่ไม่มีเยื่อเมือก หลักการและการพิสูจน์การนำกระแสประสาท
งานนำเสนอ เพิ่ม 09/30/2013
ทฤษฎีการเกิดและการดำรงคงอยู่ของศักยภาพเยื่อพัก โครงสร้างของประสาทเคมี การถ่ายโอนข้อมูล ผู้ไกล่เกลี่ยทางเคมีการสร้างศักยภาพในการดำเนินการ การจำแนกประเภทของยาตามการแปลของการกระทำ วิธีการรับสาร
ภาคนิพนธ์ เพิ่ม 03/02/2014
ประเภทของกล้ามเนื้อมนุษย์ คุณสมบัติทางกายภาพและทางสรีรวิทยาของกล้ามเนื้อโครงร่าง ความกว้างของการหดตัวของบาดทะยัก ระดับความดันโลหิตและเลือดไปเลี้ยงอวัยวะต่างๆ ระบบประสาทอัตโนมัติและผู้ไกล่เกลี่ย การกระตุ้นเซลล์กล้ามเนื้อเรียบของร่างกาย
บทคัดย่อ เพิ่ม 03/10/2013
สรีรวิทยาปกติ สรีรวิทยาทางพยาธิวิทยา ตารางลำดับเหตุการณ์ การจำแนกตามกลุ่มและกลุ่มย่อย โครงสร้างทางเคมี กลไกการออกฤทธิ์ แหล่งที่มาของแหล่งกำเนิด เป็นต้น กลไกการออกฤทธิ์ทางชีวภาพของยาในกลุ่มนี้
ภาคนิพนธ์ เพิ่ม 07/03/2551
หลักการทำงานของระบบประสาทส่วนกลางเบื้องต้น. การควบคุมมีสองประเภทหลัก: ทางร่างกายและทางประสาท สรีรวิทยาของเซลล์ประสาท ประเภทของการเชื่อมต่อของเซลล์ประสาท โครงสร้างของไซแนปส์เป็นสถานที่ติดต่อระหว่างเซลล์ประสาทและเซลล์เอฟเฟ็กเตอร์ที่รับสัญญาณ
งานนำเสนอเพิ่ม 04/22/2015
ความสัมพันธ์ระหว่างระบบประสาทและต่อมไร้ท่อ. การเชื่อมต่อของร่างกายระหว่างเซลล์ กลุ่มผู้ไกล่เกลี่ยและควบคุมสารเคมี การจำแนกประเภทของฮอร์โมน กลไกการควบคุมต่อมไร้ท่อของเซลล์ สรีรวิทยาของระบบ hypothalamic-ต่อมใต้สมอง
งานนำเสนอ เพิ่ม 01/26/2014
คำอธิบายวิธีการฝังวัสดุรองรับเทียมสำหรับฟันปลอมแบบถอดได้และแบบถอดไม่ได้ ศึกษาข้อกำหนดพื้นฐานสำหรับการฝัง ประเภทของการเคลือบและวิธีการแปรรูปและสร้างพื้นผิวที่หยาบของรากฟันเทียม สถาปัตยกรรมกระดูก
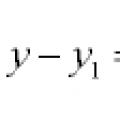 สมการเส้นและเส้นโค้งในระนาบ
สมการเส้นและเส้นโค้งในระนาบ สมการเอกพันธ์ทั่วไป สมการเอกพันธ์ทั่วไปของอันดับสอง
สมการเอกพันธ์ทั่วไป สมการเอกพันธ์ทั่วไปของอันดับสอง ขอบคุณเครื่องหมายจุลภาคเนื่องจากข้อเท็จจริงที่ว่าในพวกเขา
ขอบคุณเครื่องหมายจุลภาคเนื่องจากข้อเท็จจริงที่ว่าในพวกเขา