What is the Golgi complex in plant cells? ER and Golgi complex
The cell is an integral system
A living cell is a unique, perfect, smallest unit of the body; it is designed to use oxygen and nutrients as efficiently as possible while performing its functions. Vital organelles for the cell are the nucleus, ribosomes, mitochondria, endoplasmic reticulum, and Golgi apparatus. Let's talk about the latter in more detail.
What it is
This membrane organelle is a complex of structures that remove substances synthesized in it from the cell. Most often it is located close to the outer cell membrane.
Golgi apparatus: structure
It consists of membrane-shaped “sacs” called cisterns. The latter have an elongated shape, slightly flattened in the middle and expanded at the edges. The complex also contains round Golgi vesicles - small membrane structures. The cisternae are “folded” into stacks called dictyosomes. The Golgi apparatus contains various types of “sacs”; the entire complex is divided into certain parts according to the degree of distance from the nucleus. There are three of them: the cis section (closer to the nucleus), the middle section, and the trans section - the farthest from the nucleus. They are characterized by a different composition of enzymes, and therefore the work performed. There is one feature in the structure of dictyosomes: they are polar, that is, the section closest to the nucleus only receives vesicles coming from the endoplasmic reticulum. The part of the “stack” facing the cell membrane only forms and releases them.

Golgi apparatus: functions
The main tasks performed are the sorting of proteins, lipids, mucous secretions and their removal. Non-protein substances secreted by the cell and carbohydrate components of the outer membrane also pass through it. At the same time, the Golgi apparatus is not at all an indifferent mediator that simply “transmits” substances; processes of activation and modification (“maturation”) take place in it:
- Sorting of substances, transport of proteins. The distribution of protein substances occurs into three streams: for the membrane of the cell itself, export, and lysosomal enzymes. In addition to proteins, the first stream also includes fats. An interesting fact is that any export substances are transported inside the bubbles. But proteins intended for the cell membrane are embedded in the membrane of the transport vesicle and move in this way.
- The release of all products produced in the cell. The Golgi apparatus “packs” all products, both protein and other nature, into secretory vesicles. All substances are released through the complex interaction of the latter with the cell membrane.
- Synthesis of polysaccharides (glycosaminoglycans and components of the cell wall glycocalyx).
- Sulfation, glycosylation of fats and proteins, partial proteolysis of the latter (necessary to convert them from an inactive form to an active one) - these are all processes of “maturation” of proteins necessary for their future full-fledged work.

Finally
Having examined how the Golgi complex is structured and works, we are convinced that it is the most important and integral part of any cell (especially secretory ones). A cell that does not produce substances for export also cannot do without this organelle, since the “completion” of the cell membrane and other important internal processes of life depend on it.
The Golgi complex was discovered by Camillo Golgi in 1898. This structure is present in the cytoplasm of almost all eukaryotic (constituting higher organisms) cells, especially secretory cells in animals.
Golgi complex. Structure.
The structure is represented by a stack of flattened membrane sacs. They are called tanks. This stack of sacs is connected to the Golgi system). At one end of the stacks of sacs, new cisternae are constantly being formed by the fusion of vesicles that bud from the endoplasmic reticulum (a network of cavities). At the other end of the stack on the inside of the tank, they complete maturation and break up again into bubbles. This is how the tanks in the hill gradually move towards the inner side from the outer side.
In the cisterns of the structure, the maturation of proteins intended for secretion, transmembrane proteins, lysosome proteins, and others occurs. Maturing substances move sequentially through the organelle cisterns. The final folding of proteins and their modifications - phosphorylation and glycosylation - occur in them.
Characterized by the presence of a number of individual dictyosomes (stacks). There are often several stacks connected by tubes or one large stack.
Contains four main sections: trans-Golgi network, cis-Golgi, trans-Golgi and medial-Golgi. An intermediate compartment (separate region) is also attached to the structure. It is represented by a cluster of membrane vesicles in the space between the reitculum and the cis-Golgi.
The entire apparatus is a very polymorphic (diverse) organelle. Even at different stages of development of one cell, the Golgi complex can look different.
The device also differs in its asymmetry. The cisterns located closer to the cell nucleus (cis-Golgi) contain the most immature proteins. These tanks are joined by continuous membrane vesicles - vesicles. Different tanks contain different resident enzymes (catalytic), which suggests that different processes occur in them with maturing proteins.
Golgi complex. Functions.
The tasks of the structure include chemical modification and transport of substances entering it. Proteins that penetrate into the apparatus from the endoplasmic reticulum are the initial substrate for enzymes. After being concentrated and modified, the enzymes in the vesicles are transported to the designated site. For example, this could be the area where a new kidney is forming. With the participation of cytoplasmic microtubules, the transfer process is most active.
The Golgi complex also performs the task of attaching carbohydrate groups to proteins and the subsequent use of these proteins in the construction of the membrane of lysosomes and cells.
In some algae, cellulose fibers are synthesized in the structure of the apparatus.
The functions of the Golgi complex are quite diverse. Among them are:
- Sorting, removal, accumulation of secretory products.
- Accumulation of lipid molecules and formation of lipoproteins.
- Completion of protein modification (post-translational), namely glycosylation, sulfation, etc.
- Formation of lysosomes.
- Participation in the formation of acrosomes.
- Polysaccharide synthesis for the formation of waxes, glycoproteins, mucus, gum, matrix substances in plants (pectins, hemicellulose and others).
- Formation of contractile vacuoles in protozoa.
- Formation of the cell plate in plant cells after nuclear division.
18. Golgi complex, its structure and functions. Lysosomes. Their structure and functions. Types of lysosomes.
Golgi complex It is a stack of disc-shaped membrane sacs (cisternae), somewhat expanded closer to the edges, and an associated system of Golgi vesicles. A number of individual stacks (dictyosomes) are found in plant cells; animal cells often contain one large or several stacks connected by tubes.
1. Accumulates and removes organic substances synthesized in the endoplasmic reticulum
2. Forms lysosomes
3. Formation of carbohydrate components of the glycocalyx - mainly glycolipids.
Lysosomes are an integral part of the cell composition. They are a type of vesicle. These cellular helpers, being part of the vacuome, are covered with a membrane and filled with hydrolytic enzymes. The importance of the existence of lysosomes inside the cell is ensured by the secretory function, which is necessary in the process of phagocytosis and autophagocytosis.
Perform digestive function- digest food particles and remove dead organelles.
Primary lysosomes- these are small membrane vesicles that have a diameter of about one hundred nm, filled with homogeneous fine contents, which are a set of hydrolytic enzymes. Lysosomes contain about forty enzymes.
Secondary lysosomes are formed by the fusion of primary lysosomes with endocytic or pinocytotic vacuoles. To put it another way, secondary lysosomes are intracellular digestive vacuoles, the enzymes of which are supplied by primary lysosomes, and the material for digestion is supplied by the endocytic (pinocytotic) vacuole.
19. Eps, its varieties, role in the processes of synthesis of substances.
Endoplasmic reticulum in different cells it can be presented in the form of flattened cisterns, tubules or individual vesicles. The wall of these formations consists of a bilipid membrane and some proteins included in it and delimits the internal environment of the endoplasmic reticulum from the hyaloplasm.
There are two types of endoplasmic reticulum:
granular (granular or rough);
non-grainy or smooth.
The outer surface of the membranes of the granular endoplasmic reticulum contains attached ribosomes. There may be both types of endoplasmic reticulum in the cytoplasm, but usually one form predominates, which determines the functional specificity of the cell. It should be remembered that the two named varieties are not independent forms of the endoplasmic reticulum, since one can trace the transition of the granular endoplasmic reticulum to the smooth one and vice versa.
Functions of the granular endoplasmic reticulum:
synthesis of proteins intended for removal from the cell (“for export”);
separation (segregation) of the synthesized product from the hyaloplasm;
condensation and modification of synthesized protein;
transport of synthesized products into the tanks of the lamellar complex or directly from the cell;
synthesis of bilipid membranes.
The smooth endoplasmic reticulum is represented by cisterns, wider channels and individual vesicles, on the outer surface of which there are no ribosomes.
Functions of smooth endoplasmic reticulum:
participation in glycogen synthesis;
lipid synthesis;
detoxification function - neutralization of toxic substances by combining them with other substances.
The lamellar Golgi complex (reticular apparatus) is represented by a cluster of flattened cisterns and small vesicles bounded by a bilipid membrane. The lamellar complex is divided into subunits - dictyosomes. Each dictyosome is a stack of flattened cisternae, along the periphery of which small vesicles are localized. At the same time, in each flattened cistern, the peripheral part is somewhat expanded, and the central part is narrowed.
In 1898, the Italian scientist C. Golgi, using the properties of binding heavy metals (osmium and silver) with cellular structures, identified mesh formations in nerve cells, which he called the “internal mesh apparatus” (Fig. 174). Further improvement of the metal staining method (impregnation) made it possible to verify that network structures (Golgi apparatus) are found in all cells of any eukaryotic organisms. Typically, the elements of the Golgi apparatus are located near the nucleus, near the cell center (centriole). Areas of the Golgi apparatus, clearly identified by the impregnation method, had the appearance of complex networks in some cells, where the cells were connected to each other or were presented in the form of separate dark areas lying independently of each other (dictyosomes), having the form of rods, grains, concave disks and etc. (Fig. 175). There is no fundamental difference between the reticular and diffuse forms of the Golgi apparatus, since a change in the forms of this organelle is often observed in the same cells. Elements of the Golgi apparatus are often associated with vacuoles, which is especially characteristic of secreting cells.
It was found that the morphology of AG changes depending on the stages of cellular secretion, which served as the basis for D.N. Nasonov (1924) put forward the hypothesis that AG is an organelle that ensures the separation and accumulation of substances in a wide variety of cells.
For a long time, it was not possible to detect elements of the Golgi apparatus in plant cells using conventional microtechnical methods. However, with the advent of electron microscopy, AG elements were discovered in all plant cells, where they are located along the cell periphery.
Fine structure of the Golgi apparatus
An electron microscope shows that the Golgi apparatus is represented by membrane structures collected together in a small zone (Fig. 176, 177). A separate zone of accumulation of these membranes is dictyosome(Fig. 178). In the dictyosome, flat membrane sacs, or cisterns, are located close to each other (at a distance of 20-25 nm) in the form of a stack, between which thin layers of hyaloplasm are located. Each individual tank has a diameter of about 1 μm and variable thickness; in the center its membranes can be close together (25 nm), and at the periphery they can have expansions, ampoules, the width of which is not constant. The number of such bags in a stack usually does not exceed 5-10. In some single-celled organisms their number can reach 20. In addition to densely located flat cisterns, many vacuoles are observed in the AG zone. Small vacuoles are found mainly in the peripheral areas of the AG zone; sometimes you can see how they are laced from the ampullary extensions at the edges of the flat cisterns. It is customary to distinguish in the dictyosome zone the proximal or developing, cis-section, and the distal or mature, trans-section (Fig. 178). Between them is the middle or intermediate section of the AG.
During cell division, the reticulate forms of AG disintegrate into dictyosomes, which are passively and randomly distributed among daughter cells. As cells grow, the total number of dictyosomes increases.
In secreting cells, the AG is usually polarized: its proximal part faces the cytoplasm and nucleus, and the distal part faces the cell surface. In the proximal area, the stacks of closely spaced cisterns are adjacent to a zone of small smooth vesicles and short membrane cisterns. In samples of preparatively isolated AG zones with negative contrast, it is clear that a network-like or sponge-like system of membrane cavities adjoins the proximal part of the dictyosome. It is believed that this system may represent a zone of transition of ER elements into the zone of the Golgi apparatus (Fig. 179).
In the middle part of the dictyosome, the periphery of each cistern is also accompanied by a mass of small vacuoles about 50 nm in diameter.
In the distal or trans-section of dictyosomes, the last membrane flat cistern is adjacent to a section consisting of tubular elements and a mass of small vacuoles, often having fibrillar pubescence along the surface on the side of the cytoplasm - these are pubescent or bordered vesicles of the same type as the bordered vesicles during pinocytosis. This is the so-called trans-Golgi apparatus network(TGN), where the separation and sorting of secreted products occurs. Even more distal is a group of larger vacuoles - this is the product of the fusion of small vacuoles and the formation of secretory vacuoles.
When studying thick sections of cells using a megavolt electron microscope, it was found that in cells individual dictosomes can be connected to each other by a system of vacuoles and cisterns. So a loose three-dimensional network is formed, which is visible in a light microscope. In the case of the diffuse form of AG, each individual section is represented by a dictyosome. In plant cells, the diffuse type of AG organization predominates; usually, on average, there are about 20 dictyosomes per cell. In animal cells, centrioles are often associated with the membrane zone of the Golgi apparatus; between the bundles of microtubules extending radially from them lie groups of stacks of membranes and vacuoles, which concentrically surround the cell center. This connection likely reflects the involvement of microtubules in vacuole movement.
Secretory function of the Golgi apparatus
Membrane elements of AG are involved in the segregation and accumulation of products synthesized in the ER, and participate in their chemical rearrangements and maturation: this is mainly the rearrangement of the oligosaccharide components of glycoproteins in the composition of water-soluble secretions or in the composition of membranes (Fig. 180).
In the AG tanks, the synthesis of polysaccharides occurs, their interaction with proteins, leading to the formation of mucoproteins. But most importantly, with the help of elements of the Golgi apparatus, the process of removing ready-made secretions outside the cell occurs. In addition, AG is a source of cellular lysosomes.
The participation of AG in the processes of excretion of secretory products has been very well studied using the example of exocrine pancreatic cells. These cells are characterized by the presence of a large number of secretory granules (zymogen granules), which are membrane vesicles filled with protein content. The proteins of zymogen granules include various enzymes: proteases, lipases, carbohydrates, nucleases. During secretion, the contents of these zymogen granules are released from the cells into the lumen of the gland, and then flows into the intestinal cavity. Since the main product excreted by pancreatic cells is protein, the sequence of incorporation of radioactive amino acids into different parts of the cell was studied (Fig. 181). For this purpose, animals were injected with tritium-labeled amino acid (3H-leucine) and the localization of the label was monitored over time using electron microscopic autoradiography. It turned out that after a short period of time (3-5 min) the label was localized only in the basal areas of the cells, in areas rich in granular ER. Since the label was included in the protein chain during protein synthesis, it was clear that protein synthesis did not occur either in the AG zone or in the zymogen granules themselves, but it was synthesized exclusively in the ergastoplasm on ribosomes. Somewhat later (after 20-40 minutes), a label other than ergastoplasma was found in the zone of AG vacuoles. Consequently, after synthesis in ergastoplasm, the protein was transported to the AG zone. Even later (after 60 min), the label was already detected in the zone of zymogen granules. Subsequently, the mark could be seen in the lumen of the acini of this gland. Thus, it became clear that AG is an intermediate link between the actual synthesis of the secreted protein and its removal from the cell. The processes of protein synthesis and excretion were also studied in detail in other cells (mammary gland, intestinal goblet cells, thyroid gland, etc.), and the morphological features of this process were studied. The exported protein synthesized on ribosomes is separated and accumulates inside the ER cisterns, through which it is transported to the AG membrane zone. Here, small vacuoles containing the synthesized protein are split off from the smooth areas of the ER and enter the vacuole zone in the proximal part of the dictyosome. At this point, the vacuoles can merge with each other and with the flat cis cisternae of the dictyosome. In this way, the protein product is transferred already inside the cavities of the AG tanks.
As proteins in the cisternae of the Golgi apparatus are modified, they are transported from cisternae to cisternae into the distal part of the dictyosome by means of small vacuoles until they reach the tubular membrane network in the trans region of the dictyosome. In this area, small bubbles containing an already mature product are separated. The cytoplasmic surface of such vesicles is similar to the surface of bordered vesicles, which are observed during receptor pinocytosis. The separated small vesicles merge with each other, forming secretory vacuoles. After this, the secretory vacuoles begin to move towards the cell surface, come into contact with the plasma membrane, with which their membranes fuse, and thus the contents of these vacuoles appear outside the cell. Morphologically, this process of extrusion (throwing out) resembles pinocytosis, only with the reverse sequence of stages. It's called exocytosis.
This description of events is only a general diagram of the participation of the Golgi apparatus in secretory processes. the matter is complicated by the fact that the same cell can participate in the synthesis of many secreted proteins, can isolate them from each other and direct them to the cell surface or into lysosomes. In the Golgi apparatus, there is not just a “pumping” of products from one cavity to another, but also their gradual “maturation”, modification of proteins, which ends with the “sorting” of products sent either to lysosomes, or to the plasma membrane, or to secretory vacuoles.
Modification of proteins in the Golgi apparatus
Proteins synthesized in the ER enter the cis-zone of the Golgi apparatus after primary glycosylation and reduction of several saccharide residues there. Ultimately, all proteins there have the same oligosaccharide chains, consisting of two molecules of N-acetylglucosamine, six molecules of mannose (Fig. 182). In cis-cisternae, secondary modification of oligosaccharide chains begins and their sorting into two classes. As a result, oligosaccharides on hydrolytic enzymes intended for lysosomes (mannose-rich olgosaccharides) are phosphorylated, and oligosaccharides of other proteins sent to secretory granules or to the plasma membrane undergo complex transformations, losing a number of sugars and adding galactose, N-acetylglucosamine and sialic acids .
In this case, a special complex of oligosaccharides appears. Such transformations of oligosaccharides are carried out with the help of enzymes - glycosyltransferases, which are part of the membranes of the Golgi apparatus cisterns. Since each zone in dictyosomes has its own set of glycosylation enzymes, glycoproteins are transferred, as if in a relay race, from one membrane compartment (“floor” in a stack of dictyosome tanks) to another and in each are subjected to the specific action of enzymes. Thus, in the cis-site, phosphorylation of mannoses in lysosomal enzymes occurs and a special mannose-6 group is formed, characteristic of all hydrolytic enzymes, which then enter the lysosomes.
In the middle part of dictyosomes, secondary glycosylation of secretory proteins occurs: additional removal of mannose and addition of N-acetylglucosamine. In the trans region, galactose and sialic acids are added to the oligosaccharide chain (Fig. 183).
These data were obtained using completely different methods. Using differential centrifugation, it was possible to obtain separate heavier (cis-) components of the Golgi apparatus and lighter (trans-) components and determine the presence of glycosidases and their products in them. On the other hand, using monoclonal antibodies to various enzymes using electron microscopy, it was possible to localize them directly on cell sections.
In a number of specialized cells in the Golgi apparatus, the synthesis of polysaccharides themselves occurs.
In the Golgi apparatus of plant cells, the synthesis of cell wall matrix polysaccharides (hemicelluloses, pectins) occurs. In addition, dictyosomes of plant cells are involved in the synthesis and secretion of mucus and mucins, which also include polysaccharides. The synthesis of the main framework polysaccharide of plant cell walls, cellulose, occurs, as already mentioned, on the surface of the plasma membrane.
In the Golgi apparatus of animal cells, the synthesis of long unbranched polysaccharide chains of glucosainoglycans occurs. One of them, hyaluronic acid, which is part of the extracellular matrix of connective tissue, contains several thousand repeating disaccharide blocks. Many glycosainoglycans are covalently linked to proteins and form proteoglycans (mucoproteins). Such polysaccharide chains are modified in the Golgi apparatus and bind to proteins, which are secreted by cells in the form of proteoglycans. Sulfation of glycosainoglycans and some proteins also occurs in the Golgi apparatus.
Protein sorting in the Golgi apparatus
So, at least three streams of non-cytosolic proteins synthesized by the cell pass through the Golgi apparatus: a stream of hydrolytic enzymes into the lysosome compartment, a stream of secreted proteins that accumulate in secretory vacuoles and are released from the cell only upon receipt of special signals, a stream of constantly secreted secretory proteins. Therefore, there must be some special mechanism for the spatial separation of these different proteins and their pathways.
In the cis- and middle zones of dictyosomes, all these proteins go together without separation, they are only separately modified depending on their oligosaccharide markers.
The actual separation of proteins, their sorting, occurs in the trans region of the Golgi apparatus. This process has not been fully deciphered, but using the example of the sorting of lysosomal enzymes, one can understand the principle of selection of certain protein molecules (Fig. 184).
It is known that only precursor proteins of lysosomal hydrolases have a specific oligosaccharide, namely a mannose group. In cis cisternae, these groups are phosphorylated and then, together with other proteins, are transferred from cisternae to cisternae, through the middle zone to the trans region. The membranes of the trans-network of the Golgi apparatus contain a transmembrane protein receptor (mannose-6-phosphate receptor or M-6-P receptor), which recognizes phosphorylated mannose groups of the oligosaccharide chain of lysosomal enzymes and binds to them. This binding occurs at neutral pH values within the cisternae of the trans network. On membranes, these M-6-F receptor proteins form clusters, groups that are concentrated in the zones of formation of small vesicles coated with clathrin. In the trans-network of the Golgi apparatus, their separation, budding and further transfer to endosomes occur. Consequently, M-6-F receptors, being transmembrane proteins, bind to lysosomal hydrolases, separate them, sort them from other proteins (for example, secretory, non-lysosomal) and concentrate them in bordered vesicles. Having separated from the trans-network, these vesicles quickly lose their coat, merge with endosomes, transferring their lysosomal enzymes associated with membrane receptors into this vacuole. As already mentioned, acidification of the environment occurs inside endosomes due to the activity of the proton transporter. Starting at pH 6, lysosomal enzymes dissociate from M-6-P receptors, are activated and begin to work in the cavity of the endolysosome. Sections of membranes, together with M-6-F receptors, are returned by recycling membrane vesicles back into the trans-network of the Golgi apparatus.
Most likely, that part of the proteins that accumulates in secretory vacuoles and is removed from the cell after receiving a signal (for example, nervous or hormonal) undergoes the same selection and sorting procedure on the receptors of the trans-cisterns of the Golgi apparatus. These secretory proteins first enter small vacuoles, also coated with clathrin, which then merge with each other. In secretory vacuoles, accumulated proteins often aggregate in the form of dense secretory granules. This results in an increase in protein concentration in these vacuoles by approximately 200 times compared to its concentration in the Golgi apparatus. Then these proteins, as they accumulate in secretory vacuoles, are released from the cell by exocytosis, when the cell receives the corresponding signal.
The third stream of vacuoles, associated with constant, constitutive secretion, also emanates from the Golgi apparatus. Thus, fibroblasts secrete a large amount of glycoproteins and mucins that are part of the main substance of connective tissue. Many cells constantly secrete proteins that facilitate their binding to substrates; there is a constant flow of membrane vesicles to the cell surface, carrying elements of the glycocalyx and membrane glycoproteins. This flow of components released by the cell is not subject to sorting in the receptor trans-system of the Golgi apparatus. The primary vacuoles of this flow also split off from the membranes and are related in their structure to bordered vacuoles containing clathrin (Fig. 185).
Concluding the consideration of the structure and operation of such a complex membrane organelle as the Golgi apparatus, it is necessary to emphasize that despite the apparent morphological homogeneity of its components, the vacuole and the cisterna, in fact, it is not just a collection of vesicles, but a slender, dynamic, complexly organized, polarized system.
In the AG, not only the transport of vesicles from the ER to the plasma membrane occurs. There is retrograde transport of vesicles. Thus, vacuoles split off from secondary lysosomes and return, together with receptor proteins, to the trans-AG zone. In addition, there is a flow of vacuoles from the trans zone to the cis zone of the AG, as well as from the cis zone to the endoplasmic reticulum. In these cases, the vacuoles are coated with proteins of the COP I complex. It is believed that various secondary glycosylation enzymes and receptor proteins in membranes are returned in this way.
These features of the behavior of transport vesicles gave rise to the hypothesis that there are two types of transport of AG components (Fig. 186).
According to one of them, the oldest, there are stable membrane components in the AG, to which substances are relayed from the ER using transport vacuoles. According to an alternative model, AG is a dynamic derivative of the ER: membrane vacuoles split off from the ER merge with each other into a new cis-tank, which then moves through the entire AG zone and finally breaks up into transport vesicles. According to this model, retrograde COP I vesicles return resident Ag proteins to younger cisternae. Thus, it is assumed that the transition zone of the ER represents a “maternity hospital” for the Golgi apparatus.
A- Granular cytoplasmic reticulum.
B- Microbubbles.
B-Microfilaments.
G-Tank.
D- Vacuoles.
Answer: B, D, D.
16. Indicate what functions the Golgi complex performs:
A- Protein synthesis.
B- Formation of complex chemical compounds (glycoproteins, lipoproteins).
B- Formation of primary lysosomes.
D- Participation in the removal of secretory product from the cell.
D- Formation of hyaloplasm.
Answer: B, C, D.
What structural elements of the cell are most actively involved in exocytosis?
A- Cytolemma.
B- Cytoskeleton.
B- Mitochondria.
G-Ribosomes.
Answer: A, B.
18 . What determines the specificity of the protein being synthesized?
A- Messenger RNA.
B- Ribosomal RNA.
D- Membranes of the cytoplasmic reticulum.
Answer: A, B
19 . Which structural elements are actively involved in execution?
Phagocytic function?
A- Karyolemma.
B- Endoplasmic reticulum.
B- Cytolemma.
G-Lysosomes.
D- Microfilaments.
Answer: B, D, D.
20 .What structural components of the cell determine the basophilia of the cytoplasm?
A- Ribosomes.
B- Agranular endoplasmic reticulum.
B- Lysosomes.
G- Peroxisomes.
D-Golgi complex.
E- Granular endoplasmic reticulum.
Answer: A, E.
21 . Which of the following organelles have a membrane structure?
A - Cellular center.
B- Mitochondria.
B- Golgi complex.
G-Ribosomes.
D- Cytoskeleton.
Answer: B, C.
22 .What do mitochondria and peroxisomes have in common?
A- They belong to organelles with a membrane structure.
B- They have a double membrane.
D- These are organelles of general importance.
Answer: A, B, D.
What functions do lysosomes perform in a cell?
A- Protein biosynthesis
B- Participation in phagocytosis
B- Oxidative phosphorylation
D- Intracellular digestion
Answer: B.G.
What is the structural organization of lysosomes?
A- Surrounded by a membrane.
B- Filled with hydrolytic enzymes.
D- Formed in the Golgi complex.
Answer: A, B, D.
25. Glycocalyx:
A- Located in the smooth endoplasmic reticulum.
B- Located on the outer surface of the cytolemma.
B- Formed by carbohydrates.
D- Participates in cell adhesion and cell recognition.
D- Located on the inner surface of the cytolemma.
Answer: B, C, D.
26. Marker enzymes of lysosomes:
A-Acid phosphatase.
B-ATPase.
B- Hydrolases.
G- Catalase and oxidases.
Answer: A, B.
What is the importance of the nucleus in the life of a cell?
A- Storage of hereditary information.
B- Energy storage center.
B- Intracellular metabolism control center.
D- Place of formation of lysosomes.
D- Reproduction and transmission of genetic information to daughter cells.
Answer: A, B, D.
28. What does not apply to the structural components of the nucleus:
A- Karyolemma.
B- Nucleoli.
B- Karyoplasm.
G-Ribosomes.
D- Chromatin, chromosomes.
E- Peroxisomes.
Answer: G, E.
What is transported from the nucleus through nuclear pores into the cytoplasm?
A- DNA fragments.
B- Ribosomal subunits.
B- Messenger RNAs.
D- Fragments of the endoplasmic reticulum.
Answer: B, C.
What is the nuclear-cytoplasmic ratio and how does it change with increasing functional activity of the cell?
A- Position of the nucleus in the cytoplasm.
B- Core shape.
B- The ratio of the size of the nucleus to the size of the cytoplasm.
D- Reduced with increased functional activity of the cell.
Answer: B, G.
What is true for nucleoli?
A- Clearly visible during mitosis.
B- They consist of granular and fibrillar components.
B- Nucleolar granules are subunits of ribosomes.
G- Nucleolar threads - ribonucleoproteins
Answer: B, C, D.
Which of the following signs refer to necrosis?
A- This is genetically programmed cell death
B- At the beginning of apoptosis, the synthesis of RNA and protein increases.
B- membranes are destroyed
G-enzymes of lysosomes enter the cytoplasm
D- Fragmentation of the cytoplasm with the formation of apoptotic bodies
Answer: B, G.
Everything is true except
1.Function of the Golgi complex (all are correct except):
A - sorting of proteins into transport vesicles
B- glycosylation of proteins
B- recycling of membranes of secretory granules after exocytosis
G - packaging of secretory product
D - synthesis of steroid hormones
2. Microtubules provide (all are true except):
A - organization of the internal space of the cell
B- maintaining cell shape
B- polarization of the cell during division
G- form the contractile apparatus
D- organization of the cytoskeleton
E-transport of organelles
3. Specialized structures built on the basis of the cytoskeleton include (all are true except):
A- cilia, flagella
B - basal striation
B- microvilli
4. Localization of cilia (all are true except):
A- epithelium of the mucous membrane of the airways
B- epithelium of the proximal nephron
B- epithelium of the mucous membrane of the female reproductive tract
G - epithelium of the mucous membrane of the vas deferens
5. Localization of microvilli (all are true except):
A- epithelium of the mucous membrane of the small intestine
B- epithelium of the tracheal mucosa
B- epithelium of the proximal nephron
6. Basal striation (all are true except):
A- ensures the transport of substances against a concentration gradient
B - area of the cell where highly energy-intensive processes take place
B - area of the cell where simple diffusion of ions occurs
D - where reabsorption of elements of primary urine occurs in the proximal tubule of the nephron
D- participates in the concentration of salivary secretion
7. Brush border (all are correct except):
A- located on the apical surface of cells
B- increases the suction surface area
B - consists of cilia
G- consists of microvilli
D- increases the transport surface in the proximal tubules of the nephron
8. General purpose organoids (all are correct except):
A- mitochondria
B-Golgi complex
G-cilia
D-lysosomes
E-peroxisomes
F-centrioles
Z-elements of the cytoskeleton
9.Function of peroxisomes (all are true except):
A- oxidation of an organic substrate with the formation of hydrogen peroxide
B- synthesis of enzyme - catalase
B- utilization of hydrogen peroxide
10. Ribosomes (all are correct except):
A - with light microscopy, their presence is judged by the pronounced basophilia of the cytoplasm
B- consist of small and large subunits
B- are formed in granular ER
G- consist of rRNA and proteins
D- non-membrane structure
11.Which organelles are well developed in steroid-producing cells (all are true except):
A - granular endoplasmic reticulum
B- agranular endoplasmic reticulum
B-mitochondria with tubular cristae
12. Trophic inclusions (all are true except):
A-carbohydrates
B- mucous membranes
B-proteins
G-lipid
13.Nuclear envelope (all are true except):
A - consists of a single membrane
B - consists of two membranes
B - ribosomes are located on the outside
D - the nuclear lamina is connected to it from the inside
D - permeated with pores
14. Structural components of the kernel (all are true except):
A - nucleoplasm
B-nucleolemma
B-microtubules
G-chromatin
D- nucleoli
15. Structure of a nuclear pore (all are true except):
A - membrane component
B-chromosomal component
B- fibrillar component
G-granular component
16. Nucleolus (all are true except):
A- surrounded by a membrane
B- not surrounded by a membrane
B- its organization involves five pairs of chromosomes
G- contains granular and fibrillar components
17. Nucleolus (all are true except):
A - the amount depends on the metabolic activity of the cell
B- participates in the formation of ribosomal subunits
B- chromosomes 13,14, 15, 21 and 22 are involved in the organization
D - 7, 8, 10, 11 and 23 chromosomes are involved in the organization
D - consists of three components
18. Cellular center (all are correct except):
A- is localized near the nucleus
B- is the center of spindle organization
B- consists of two centrioles
G-centrioles are formed by 9 doublets of microtubules
D-centrioles are duplicated in the S period of interphase
19. Mitochondria (all are correct except):
A - presence of cristae
B- ability to share
20. Functions of actin filaments (all are correct except):
A - cell movement
B- change in cell shape
B- participation in exo- and endocytosis
D - provide movement of cilia
D- are part of microvilli
21. Everything is true for the nucleolus except:
A- Formed in the area of nucleolar organizers (secondary chromosome constrictions)
B- Nucleolar granules extend into the cytoplasm
B- Nucleolar proteins are synthesized in the cytoplasm
D- Nucleolar RNA is formed in the cytoplasm
For compliance
1. Compare the periods of interphase with the processes occurring in them:
1. Presynthetic A - DNA doubling, increased RNA synthesis
2. Synthetic B-synthesis of rRNA, mRNA, tubulins
3. Postsynthetic B-cell growth, preparing them for DNA synthesis
Answer: 1-B; 2-A; 3-B.
2 .Compare the phases of mitosis with the processes occurring in them:
1. Prophase A - formation of the equatorial plate from chromosomes
2. Metaphase B - formation of nucleolemma, despiralization of chromosomes,
nucleolus formation, cytotomy
3. Anaphase B-spiralization of chromosomes, disappearance of the nucleolus,
fragmentation of the nucleolemma
4. Telophase G - divergence of chromatids to opposite poles
Answer: 1-B; 2-A; 3-G; 4-B.
3. A change in the structure of the kernel is called (match):
1.karyolysis A - reduction in size and compaction of chromatin
2.karyorrhexis B - fragmentation
3.karyopyknosis B - dissolution of its components
Answer: 1-B, 2-B, 3-A.
4. Characteristics of the components of the drug:
1.chromophobic A - stained with Sudan dye
2.chromophilic B - not stained with dye
3. Sudanophilic B - stained with dye
 Partial derivatives of a function of three variables
Partial derivatives of a function of three variables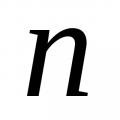 Empirical distribution function Using this sample, construct an empirical distribution function
Empirical distribution function Using this sample, construct an empirical distribution function No money, but you hold on
No money, but you hold on