تنتمي الطحالب الخضراء المزرقة إلى المملكة. عمل مستقل
من بين الكائنات الحية الموجودة حاليًا، هناك تلك التي يتم مناقشة انتمائها إلى أي كائن باستمرار. يحدث هذا مع مخلوقات تسمى البكتيريا الزرقاء. على الرغم من أنه ليس لديهم حتى اسم دقيق. المرادفات كثيرة جدًا:
- طحلب اخضر مزرق؛
- السيانوبيونات.
- كسارات فيكوكروم.
- سماوي.
- الطحالب الوحل وغيرها.
لذلك اتضح أن البكتيريا الزرقاء صغيرة تمامًا، ولكنها في نفس الوقت كائن معقد ومتناقض يتطلب دراسة متأنية ودراسة بنيتها من أجل تحديد انتمائها التصنيفي الدقيق.
تاريخ الوجود والاكتشاف
إذا حكمنا من خلال بقايا الحفريات، فإن تاريخ وجود الطحالب الخضراء المزرقة يعود إلى الماضي البعيد، منذ عدة (3.5) مليار سنة. أصبحت هذه الاستنتاجات ممكنة من خلال دراسات علماء الحفريات الذين قاموا بتحليل الصخور (أقسامها) في تلك الأوقات البعيدة.
تم العثور على البكتيريا الزرقاء على سطح العينات، والتي لم يختلف تركيبها عن الأشكال الحديثة. يشير هذا إلى درجة عالية من قدرة هذه المخلوقات على التكيف مع الظروف المعيشية المختلفة وقدرتها على التحمل الشديد وبقائها. من الواضح أنه على مدى ملايين السنين حدثت تغيرات كثيرة في درجة الحرارة وتكوين الغاز للكوكب. ومع ذلك، لم يؤثر أي شيء على جدوى السماوي.
في العصر الحديث، البكتيريا الزرقاء هي كائن حي وحيد الخلية تم اكتشافه في وقت واحد مع أشكال أخرى من الخلايا البكتيرية. أي أنطونيو فان ليفينهوك ولويس باستور وباحثين آخرين في القرنين الثامن عشر والتاسع عشر.
وقد خضعوا لدراسة أكثر شمولاً في وقت لاحق، مع تطور المجهر الإلكتروني وأساليب وأساليب البحث الحديثة. تم تحديد الميزات التي تمتلكها البكتيريا الزرقاء. يتضمن هيكل الخلية عددًا من الهياكل الجديدة غير الموجودة في الكائنات الأخرى.

تصنيف
تظل مسألة تحديد انتمائهم التصنيفي مفتوحة. حتى الآن، لا يُعرف سوى شيء واحد: البكتيريا الزرقاء هي بدائيات النوى. وهذا ما تؤكده ميزات مثل:
- غياب النواة والميتوكوندريا والبلاستيدات الخضراء.
- وجود مورين في جدار الخلية.
- جزيئات الريبوسومات S في الخلية.
ومع ذلك، فإن البكتيريا الزرقاء هي بدائيات النوى، ويبلغ عددها حوالي 1500 ألف نوع. تم تصنيفهم جميعًا ودمجهم في 5 مجموعات مورفولوجية كبيرة.
- كروكوكال. مجموعة كبيرة إلى حد ما توحد الأشكال الانفرادية أو الاستعمارية. يتم تجميع التركيزات العالية من الكائنات الحية معًا بواسطة مخاط مشترك يفرزه جدار الخلية لكل فرد. من حيث الشكل، تشمل هذه المجموعة هياكل على شكل قضيب وكروية.
- الجنبية. تشبه إلى حد كبير الأشكال السابقة، ولكن تظهر ميزة في شكل تكوين الخلايا البيضية (المزيد عن هذه الظاهرة لاحقًا). البكتيريا الزرقاء المدرجة هنا تنتمي إلى ثلاث فئات رئيسية: Pleurocaps، Dermocaps، Myxosarcina.
- أوكسيلاتوريا. السمة الرئيسية لهذه المجموعة هي أن جميع الخلايا متحدة في بنية مخاطية مشتركة تسمى المشعرات. يحدث الانقسام دون تجاوز هذا الخيط، في الداخل. تشتمل المذبذبات على خلايا نباتية حصرية تنقسم إلى نصفين لا جنسيًا.
- الأنفية. مثيرة للاهتمام لحبهم للتبريد. إنهم قادرون على العيش في الصحاري الجليدية المفتوحة، وتشكيل طبقات ملونة عليها. ما يسمى بظاهرة "ازدهار الصحاري الجليدية". أشكال هذه الكائنات هي أيضًا خيطية على شكل ثلاثية الألوان، لكن التكاثر جنسي بمساعدة الخلايا المتخصصة - الأكياس غير المتجانسة. يمكن تضمين الممثلين التاليين هنا: Anabens، Nostoks، Calothrix.
- ستيجونيماتودز. مشابهة جدا للمجموعة السابقة. يكمن الاختلاف الرئيسي في طريقة التكاثر - فهي قادرة على الانقسام عدة مرات داخل خلية واحدة. الممثل الأكثر شعبية لهذه الجمعية هو فيشريلا.
وهكذا يتم تصنيف السيانيد حسب المعايير المورفولوجية، حيث تطرح أسئلة كثيرة حول الباقي والنتائج المربكة. لم يتمكن علماء النبات وعلماء الأحياء الدقيقة حتى الآن من التوصل إلى قاسم مشترك في تصنيف البكتيريا الزرقاء.

بيئات
نظرًا لوجود تكيفات خاصة (الكيسات غير المتجانسة، والخلايا البيضية، والثيلاكويدات غير العادية، والفجوات الغازية، والقدرة على تثبيت النيتروجين الجزيئي، وغيرها)، استقرت هذه الكائنات في كل مكان. إنهم قادرون على البقاء على قيد الحياة حتى في أقسى الظروف التي لا يمكن أن يوجد فيها أي كائن حي. على سبيل المثال، الينابيع الساخنة المحبة للحرارة، والظروف اللاهوائية مع جو كبريتيد الهيدروجين، مع درجة حموضة أقل من 4.
البكتيريا الزرقاء هي كائن حي يعيش بهدوء على رمال البحر والنتوءات الصخرية وكتل الجليد والصحاري الساخنة. يمكنك التعرف وتحديد وجود السيانيد من خلال الطلاء الملون المميز الذي تشكله مستعمراتها. يمكن أن يختلف اللون من الأزرق والأسود إلى الوردي والأرجواني.
يطلق عليها اسم اللون الأزرق والأخضر لأنها غالبًا ما تشكل طبقة مخاطية زرقاء اللون على سطح المياه العذبة أو المالحة العادية. وتسمى هذه الظاهرة "زهرة الماء". يمكن رؤيته في أي بحيرة تقريبًا تبدأ في النمو والمستنقعات.

ملامح بنية الخلية
تمتلك البكتيريا الزرقاء البنية المعتادة للكائنات بدائية النواة، ولكن هناك بعض الخصائص المميزة.
المخطط العام لبنية الخلية هو كما يلي:
- جدار الخلية مصنوع من السكريات والمورين.
- هيكل بيليبيد.
- السيتوبلازم مع مادة وراثية موزعة بحرية في شكل جزيء DNA؛
- ثيلاكويدات، التي تؤدي وظيفة التمثيل الضوئي وتحتوي على أصباغ (الكلوروفيل، الزانثوفيل، الكاروتينات).

أنواع الهياكل المتخصصة
بادئ ذي بدء، هذه هي الأكياس غير المتجانسة. هذه الهياكل ليست أجزاء، بل الخلايا نفسها كجزء من المشعرات (خيط استعماري مشترك يوحده المخاط). عند النظر إليها تحت المجهر، فإنها تختلف في تركيبها، حيث أن وظيفتها الرئيسية هي إنتاج إنزيم يسمح بتثبيت النيتروجين الجزيئي من الهواء. لذلك، لا توجد أصباغ عمليا في الأكياس غير المتجانسة، ولكن هناك الكثير من النيتروجين.
ثانيا، هذه هي الهرمونات - المناطق الممزقة من المشعرات. بمثابة مواقع التكاثر.
الخلايا البيضية هي خلايا ابنة فريدة من نوعها، مشتقة بشكل جماعي من خلية أم واحدة. في بعض الأحيان يصل عددهم إلى ألف في فترة القسمة الواحدة. Dermocaps وغيرها من Pleurocapsodiums قادرة على هذه الميزة.
Akinetes هي خلايا خاصة في حالة راحة ومدرجة في trichomes. وتتميز بجدار خلوي أكثر ضخامة غنيًا بالسكريات. دورهم يشبه الأكياس غير المتجانسة.
فجوات غازية - تحتوي عليها جميع البكتيريا الزرقاء. يشير هيكل الخلية في البداية إلى وجودها. دورهم هو المشاركة في عمليات ازدهار المياه. اسم آخر لمثل هذه الهياكل هو الكربوكسيسومات.
وهي موجودة بالتأكيد في الخلايا النباتية والحيوانية والبكتيرية. ومع ذلك، في الطحالب الخضراء المزرقة هذه الادراج مختلفة بعض الشيء. وتشمل هذه:
- الجليكوجين.
- حبيبات متعدد الفوسفات
- السيانوفيسين هو مادة خاصة تتكون من الأسبارتات والأرجينين. يعمل على تراكم النيتروجين، حيث أن هذه الادراج موجودة في الأكياس غير المتجانسة.
هذا ما تمتلكه البكتيريا الزرقاء. الأجزاء الرئيسية والخلايا والعضيات المتخصصة هي التي تسمح للسيانيد بالقيام بعملية التمثيل الضوئي، ولكنها في نفس الوقت تصنف على أنها بكتيريا.

التكاثر
هذه العملية ليست صعبة بشكل خاص، لأنها نفس البكتيريا العادية. يمكن للبكتيريا الزرقاء أن تنقسم نباتيًا، أو أجزاء من ثلاثية الألوان، أو خلية عادية إلى قسمين، أو تقوم بالعملية الجنسية.
في كثير من الأحيان تشارك الخلايا المتخصصة، والكيسات غير المتجانسة، والخلايا الحركية، والخلايا البيضية في هذه العمليات.
طرق النقل
يتم تغطية خلية البكتيريا الزرقاء من الخارج وأحيانًا أيضًا بطبقة من عديد السكاريد الخاص الذي يمكن أن يشكل كبسولة مخاطية حولها. بفضل هذه الميزة يتم تنفيذ حركة السماوي.
لا توجد سوط أو نواتج خاصة. لا يمكن تنفيذ الحركة إلا على سطح صلب بمساعدة المخاط، في انقباضات قصيرة. تتمتع بعض المذبذبات بطريقة غير عادية للغاية في الحركة - فهي تدور حول محورها وتتسبب في نفس الوقت في دوران المشعرات بأكملها. هكذا تحدث الحركة على السطح.

القدرة على تثبيت النيتروجين
تقريبا كل البكتيريا الزرقاء لديها هذه الميزة. وهذا ممكن بسبب وجود إنزيم النيتروجيناز، القادر على تثبيت النيتروجين الجزيئي وتحويله إلى شكل من المركبات القابلة للهضم. يحدث هذا في الهياكل غير المتجانسة. وبالتالي، فإن تلك الأنواع التي لا تمتلكها غير قادرة على الخروج من فراغ.
بشكل عام، هذه العملية تجعل البكتيريا الزرقاء مخلوقات مهمة جدًا للحياة النباتية. من خلال الاستقرار في التربة، يساعد السيانيد ممثلي النباتات على امتصاص النيتروجين المرتبط ويعيشون حياة طبيعية.
الأنواع اللاهوائية
بعض أشكال الطحالب الخضراء المزرقة (على سبيل المثال، Oscillatoria) قادرة على العيش في ظروف لا هوائية تمامًا وجو من كبريتيد الهيدروجين. في هذه الحالة، تتم معالجة المركب داخل الجسم، ونتيجة لذلك، يتم تكوين الكبريت الجزيئي وإطلاقه في البيئة.
أساس تمييز الطحالب إلى أصناف رئيسية (الممالك، الأقسام، الطبقات، إلخ) هو الخصائص التالية: نوع أصباغ التمثيل الضوئي، وبالتالي لون الخلايا؛ وجود الأسواط وبنيتها وعددها وطريقة التصاقها بالخلية؛ التركيب الكيميائي لجدار الخلية والأغشية الإضافية. التركيب الكيميائي للمواد الاحتياطية. عدد الخلايا في الثالوس وطريقة تفاعلها.
بحلول بداية القرن العشرين، تم تمييز فئات الطحالب التالية وفقًا لباشر:
– الطحالب الخضراء المزرقة
– الطحالب الحمراء
– الطحالب الخضراء
– الطحالب الذهبية
- الطحالب ذات اللون الأصفر والأخضر، أو السوطيات المتغايرة - Xanthophyceae، أو Heterocontae؛
– الدياتومات – العصوية، أو الدياتوميات؛
- الطحالب الدينوفيسية
– طحالب الكريبتوفيسيان – Cryptophyceae;
– طحالب يوجلينوفيسي.
تتميز كل فئة بمجموعة محددة من الأصباغ، وهو منتج احتياطي يتم ترسبه في الخلية أثناء عملية التمثيل الضوئي، وإذا كانت هناك أسواط، فهي بنيتها.
يتم تجميع الطحالب الدقيقة بدائية النواة في مملكة فرعية - السيانوبيونتا. وهذا يشمل جميع الطحالب الخضراء المزرقة أو السيانيد. هذه كائنات بسيطة البنية تتكيف للعيش في الماء. تتجلى الروابط التاريخية لهذه الطحالب مع البكتيريا في بنية الخلايا. لكنها تختلف عن البكتيريا في وجود الكلوروفيل "أ" ونادرا جدا - "ب". أثناء عملية التمثيل الضوئي يطلقون الأكسجين.
قسم Cyanophyta - الطحالب الخضراء المزرقة أو السيانيا
معظم البكتيريا الزرقاء عبارة عن ضوئيات إجبارية، ومع ذلك، فهي قادرة على الوجود على المدى القصير بسبب انهيار الجليكوجين المتراكم في الضوء في دورة فوسفات البنتوز المؤكسدة وفي عملية تحلل السكر.
البكتيريا الزرقاء، وفقًا للنسخة المقبولة عمومًا، كانت "المبدعين" للغلاف الجوي الحديث المحتوي على الأكسجين على الأرض، مما أدى إلى "كارثة الأكسجين" - وهو تغيير عالمي في تكوين الغلاف الجوي للأرض حدث في بداية العصر الحديث. البروتيروزويك (منذ حوالي 2.4 مليار سنة) مما أدى إلى إعادة الهيكلة اللاحقة للمحيط الحيوي والتجلد الهوروني العالمي. في الوقت الحاضر، باعتبارها مكونًا مهمًا من العوالق البحرية، توجد البكتيريا الزرقاء في بداية معظم سلاسل الغذاء وتنتج جزءًا كبيرًا من الأكسجين (لم يتم تحديد المساهمة بدقة: تتراوح التقديرات على الأرجح من 20٪ إلى 40٪). أصبحت البكتيريا الزرقاء Synechocystis أول كائن حي يقوم بالتمثيل الضوئي وتم تسلسل جينومه بالكامل. يتم النظر في إمكانية استخدام البكتيريا الزرقاء في إنشاء دورات دعم الحياة المغلقة، وكذلك كغذاء جماعي أو إضافات غذائية. تصنيف:
– طلب الكروكوكاليس – الكروكوكاليس :
فئة البكتيريا الكروية.
- ترتيب Nostocales - Nostokovae.
- ترتيب المذبذبات - المذبذبات.
– طلب Pleurocapsales - Pleurocapsaceae.
- طلب بروكلوراليس - بروكلوروفيت؛
– رتبة Stigoneomatales – Stigoneomaceae.
الطحالب الدقيقة حقيقية النواة هي نباتات أحادية أو متعددة الخلايا، ذات ألوان مختلفة، وهي نباتات ذاتية التغذية في المقام الأول، وتعيش معظمها في الماء. تحتوي بلاستيدات هذه الطحالب على الكلوروفيل وفي أغلب الأحيان كلوروفيل إضافي "ب" و"ج" والكاروتينات والزانثوفيل ونادرًا الفيكوبيلين. يعمل الماء كمانح للإلكترون لعملية التمثيل الضوئي. في السابق، كانت الطحالب تصنف كجزء من المملكة النباتية، حيث كانت تشكل مجموعة منفصلة. ومع ذلك، مع تطور الأساليب الوراثية الجزيئية لعلم اللاهوت النظامي، أصبح من الواضح أن هذه المجموعة غير متجانسة للغاية من الناحية التطورية. حاليًا، تصنف الطحالب إلى مملكتين من حقيقيات النوى: الكروميستا والبروتيستا.
طحالب تنتمي إلى مملكة كروميستا
عادةً ما يحتوي كروميو التمثيل الضوئي في البلاستيدات الخضراء الخاصة بهم على الكاروتينويد فوكوكسانثين، وهو ليس من سمات النباتات، وأحيانًا أصباغ محددة أخرى، بالإضافة إلى الكلوروفيل ج. ميزة أخرى لخلايا الكروم هي وجود اثنين من السوط حقيقيات النواة، أحدهما عادة ما يكون ريشيًا - له فروع أنبوبية من الخيط الرئيسي. غالبًا ما تكون البلاستيدات الخضراء والنواة محاطة بغشاء مشترك، بينما تحتوي البلاستيدات الخضراء على حبيبات حساسة للضوء (وصمة العار) التي تدرك التغيرات في شدة الضوء وتحدد الانجذاب الضوئي. المواد الاحتياطية للكروم ليست النشا، كما هو الحال في النباتات، ولكن مادة الليوكوسين الشبيهة بالدهون، أو عديد السكاريد عشب البحر أو غيرها من السكريات المحددة.
– مملكة الطحالب الفرعية ( Phycobionta ) :
قسم العصوية – الدياتومات:
وباعتبارها العنصر الأكثر أهمية في العوالق البحرية، تشكل الدياتومات ما يصل إلى ربع إجمالي المواد العضوية على الكوكب.
فقط الكوكايين، الشكل متنوع. في الغالب انفرادي، وأقل استعماريًا في كثير من الأحيان. معظم ممثلي هذا التقسيم أحادي الخلية، ولكن توجد أيضًا أشكال خيطية ومشتركة. تلعب الدياتومات دورًا مهمًا جدًا في العلاقات الغذائية للكائنات المائية، كونها المكون الرئيسي للعوالق النباتية، وكذلك الرواسب السفلية. كونها كائنات حية تقوم بالتمثيل الضوئي، فهي بمثابة المصدر الرئيسي للغذاء لحيوانات المياه العذبة والبحرية. ويعتقد أنها تمثل ما يصل إلى ربع عملية التمثيل الضوئي التي تحدث على كوكبنا.
تحتوي البلاستيدات الخضراء الدياتومية على الكلوروفيل a وc، بالإضافة إلى الفوكوزانثين. التكاثر لا جنسي بشكل رئيسي - عن طريق انقسام الخلايا. يعمل الليوكوسين كمادة احتياطية.
في الدياتومات، يتم تمثيل مرحلة السوط فقط عن طريق الأمشاج الذكرية (في بعض الأنواع). لذلك، تتحرك الأشكال المتحركة بسبب التدفق الموجه للسيتوبلازم في منطقة التماس الصدفة، حيث يحد السيتوبلازم والغشاء البيئة. تحتوي هذه الكائنات الحية على أصداف ثنائية الصدف فريدة من نوعها، تتكون من السيليكا (SiO 2 ∙nH 2 O) وتشكل نصفين يتناسبان مع بعضهما البعض. تتميز الأبواب الصدفية بزخارف رائعة وتبدو جميلة جدًا. من المعروف أن أكثر من 10 آلاف نوع من الدياتومات تتميز بتنوعها المذهل وتخريمها الرائع.
عندما تموت الخلايا، لا يتم تدمير الهياكل العظمية السيليكونية، بل تتراكم على مدى عشرات الملايين من السنين في قاع المسطحات المائية. تسمى هذه الرواسب "التراب الدياتومي" وتستخدم كمادة كاشطة للتلميع والتصفية أيضًا.
قسم Chrysophyta – الطحالب الذهبية:
وهي تشمل بشكل رئيسي الطحالب المجهرية ذات ظلال مختلفة من اللون الأصفر.
معظم الطحالب الذهبية هي كائنات أحادية الخلية، وفي كثير من الأحيان مستعمرة، وحتى في كثير من الأحيان كائنات متعددة الخلايا.
في الأساس، الطحالب الذهبية عبارة عن خليط من الكائنات الحية، أي أنها تحتوي على بلاستيدات، فهي قادرة على امتصاص المركبات العضوية الذائبة و/أو جزيئات الطعام. بالنسبة للبعض، يعتمد نوع التغذية (ذاتية التغذية، أو مختلطة التغذية، أو غيرية التغذية) على الظروف البيئية أو الحالة الخلوية.
يحدث التكاثر الخضري عن طريق الانقسام الطولي للخلية إلى النصف أو عن طريق أجزاء من مستعمرة الثالوس. يتم التكاثر اللاجنسي باستخدام أبواغ حيوانية أحادية أو ثنائية السوط، أو، بشكل أقل شيوعًا، الأبواغ المسطحة والأميبات. من الأفضل وصف التكاثر الجنسي في الممثلين الذين لديهم منازل بسبب تكوين الزيجوت الملاحظ جيدًا. تلتصق الخلايا ببعضها البعض في منطقة فتحة المنزل، وتندمج البروتوبلاست الخاصة بها لتشكل الزيجوت.
هناك أكثر من ألف نوع موصوف من الطحالب الذهبية، يتم تمثيل معظمها بأشكال أحادية الخلية متحركة بسبب السوط، ولكن هناك أيضًا أنواع خيطية ومستعمرة. بعض الممثلين لديهم بنية خلية أميبية ويختلفون عن الأميبات فقط في وجود البلاستيدات الخضراء.
تفتقر العديد من النباتات الكريسوفيتية إلى جدار الخلية ولكنها تحتوي على قشور السيليكا أو عناصر هيكلية. المادة الاحتياطية هي الكريسولامين. تتمثل أصباغ التمثيل الضوئي في الكلوروفيل a وc، بالإضافة إلى الكاروتينات والزانثوفيل، التي تعطي الخلايا لونًا بنيًا ذهبيًا.
تعيش الطحالب الذهبية، كقاعدة عامة، في العوالق، ولكن توجد أيضًا أشكال سفلية متصلة. هم جزء من نيوستون. تم العثور على معظم الطحالب الذهبية بشكل رئيسي في أحواض المياه العذبة ذات المناخات المعتدلة، حيث تصل إلى أكبر تنوع للأنواع في المياه الحمضية لمستنقعات الإسفاجنوم، والتي ترتبط بتكوين الفوسفاتيز الحمضية بدلاً من القلوية. فهي تتطلب بشكل غير عادي محتوى الحديد في الماء، والذي يستخدم لتخليق السيتوكروم. ويعيش عدد أقل من الأنواع في البحار والبحيرات المالحة، ويوجد عدد قليل منها في التربة. تصل الطحالب الذهبية إلى أقصى نمو لها في موسم البرد: فهي تهيمن على العوالق في أوائل الربيع وأواخر الخريف والشتاء. في هذا الوقت، يلعبون دورًا مهمًا كمنتجين للإنتاج الأولي ويعملون كغذاء للعوالق الحيوانية. بعض الطحالب الذهبية (Uroglena، Dinobryon، Mallomonas، Synura، Prymnesium parvum)، التي تنمو بكميات كبيرة، يمكن أن تسبب ازدهار الماء.
وتستخدم أكياس الطحالب الذهبية الموجودة في الرواسب الموجودة في قاع الخزانات كمؤشرات بيئية لدراسة الظروف البيئية في الماضي والحاضر. تعمل الطحالب الذهبية على تحسين نظام الغاز في الخزانات وهي مهمة في تكوين الطمي والسابروبيل.
قسم الكريبتوفيتا - الطحالب الكريبتوفيت:
الكربتوفيت هي مجموعة صغيرة ولكنها مهمة بيئيًا وتطوريًا من الكائنات الحية التي تعيش في المياه البحرية والقارية. جميعها تقريبًا عبارة عن سوطيات متحركة أحادية الخلية، وبعض الممثلين قادرون على تشكيل مرحلة سعفية. جنس واحد فقط، Bjornbergiella (معزول عن تربة جزر هاواي) قادر على تكوين ثالي خيطي بسيط (موقع هذا الجنس غير مفهوم تمامًا، وفي عدد من الأنظمة لا يتم تصنيفه على أنه نبات كريبتوفيت)؛ وجود الأشكال الاستعمارية محل خلاف أيضًا.
من بين الكائنات الكريبتومونية هناك ذاتية التغذية، ومغايرة التغذية (السابروفتات والباجوتروفات) والمختلطة. يحتاج معظمهم إلى فيتامين ب12 والثيامين، والبعض الآخر يحتاج إلى البيوتين. يمكن أن تستخدم الكريبتوموناد مصادر الأمونيوم والنيتروجين العضوي، لكن الكائنات البحرية أقل قدرة على تحويل النترات إلى نتريت مقارنة بالطحالب الأخرى. المواد العضوية تحفز نمو الكريبتومونات.
الطريقة الرئيسية لتكاثر الكريبتوموناد هي الطريقة الخضرية، وذلك بسبب انقسام الخلايا إلى نصفين باستخدام ثلم الانقسام. في هذه الحالة، يحدث غزو البلازما بدءًا من النهاية الخلفية للخلية. في أغلب الأحيان، تظل الخلية المنقسمة متحركة. الحد الأقصى لمعدل النمو للعديد من الكريبتومونات هو تقسيم واحد يوميًا عند درجة حرارة حوالي 20 درجة مئوية. يحفز نقص النيتروجين والضوء الزائد تكوين المراحل الخاملة. تحاط الأكياس المريحة بمصفوفة سميكة خارج الخلية.
تعد النباتات الخفية ممثلًا نموذجيًا للعوالق، وتوجد أحيانًا في طمي البحيرات المالحة وبين المخلفات في المسطحات المائية العذبة. إنهم يحتلون مكانة بارزة في المياه قليلة التغذية والمعتدلة والمرتفعة والمياه العذبة والبحرية.
يفضل ممثلو المياه العذبة الخزانات الاصطناعية والطبيعية ذات المياه الراكدة - خزانات الترسيب والبرك المختلفة (البيولوجية والتقنية وتربية الأسماك) وهي أقل شيوعًا في الخزانات والبحيرات. وهي تشكل تجمعات كبيرة في البحيرات على أعماق تتراوح بين 15-23 مترًا، عند تقاطع طبقات المياه الغنية والفقيرة بالأكسجين، حيث يكون مستوى الضوء أقل بكثير مما هو عليه على السطح. الممثلون عديمو اللون شائعون في المياه الملوثة بالمواد العضوية، فهي وفيرة في مياه الصرف الصحي، لذلك يمكن أن تكون بمثابة مؤشر لتلوث المياه بالمواد العضوية.
من بين النباتات الخفية، هناك sphagnophylls نموذجية تعيش في المستنقعات ذات قيم الرقم الهيدروجيني المنخفضة، في حين يمكن لعدد من الأنواع أن تتطور في نطاق واسع من الرقم الهيدروجيني.
قسم هابتوفيتا - طحالب هابتوفيت:
النباتات الفردوسية هي مجموعة من الكائنات الأولية ذاتية التغذية، أو التناضحية، أو البلعمية التي تعيش في النظم البيئية البحرية. عادة ما تكون النباتات الفردوسية وحيدة الخلية، ولكن توجد أيضًا أشكال استعمارية. وعلى الرغم من صغر حجمها، إلا أن هذه الكائنات تلعب دورًا كبيرًا ومهمًا جدًا في الدورات الجيوكيميائية للكربون والكبريت.
بالإضافة إلى التغذية الضوئية، فإن العديد من النباتات اللمسية قادرة على الامتصاص التناضحي والبلعمي للعناصر الغذائية، لذا فإن التغذية المختلطة هي ظاهرة شائعة بالنسبة لها.
تعيش معظم نباتات البرامينسيوفيت في البحار، وتفضل المناطق المفتوحة، ويوجد عدد قليل منها فقط في المياه العذبة والمالحة. تحقق نباتات البريمنسيوفيت أعظم تنوع بيولوجي لها في المياه التي تحتوي على كميات قليلة من العناصر الغذائية، وفي المياه المحيطية المفتوحة شبه الاستوائية، حيث توجد حتى على أعماق تزيد عن 200 متر.
تلعب بعض نباتات البريمنسيوفيت دورًا سلبيًا في الطبيعة. وهكذا، فإن الأنواع التي تشكل المكورات الحجرية تشارك في ظاهرة الاحتباس الحراري كعناصر أساسية في التوازن العالمي لثاني أكسيد الكربون. يمكن أن تسبب "إزهارًا" من الماء ، والذي يسمى "الأبيض" نظرًا لقدرة العصعصيات على عكس الضوء.
قسم Xanthophyta – الطحالب الصفراء والخضراء:
الطحالب التي تكون البلاستيدات الخضراء صفراء أو خضراء أو صفراء.
من بين النباتات الزانثوفيتية توجد أشكال سوطية أحادية الخلية، بالإضافة إلى أشكال استعمارية وخيطية ومختلطة. يتم تمثيل الأخير من خلال جنس Vaucheria المنتشر ("شعر الماء"). غالبًا ما توجد هذه الطحالب الخيطية المتفرعة بشكل ضعيف في الطين الساحلي الذي تغمره المياه بشكل دوري.
عرفت معظم النباتات الصفراء الخضراء تكاثرًا نباتيًا ولا جنسيًا.
يتم تضمين الطحالب الخضراء الصفراء في مجموعات بيئية مختلفة - العوالق ، وفي كثير من الأحيان البيريفيتون والقاع. الغالبية العظمى منها هي أشكال تعيش حرة.
طحالب تنتمي إلى مملكة الطلائعيات
تعد احتجاجات التمثيل الضوئي، جنبًا إلى جنب مع البروتستانت غير المتجانسة، جزءًا من أنواع مختلطة - دينوفلاجيلاتا (دينوفلاجيلات) وأوغلينيدا (أوجلينا)، وتمثلها أيضًا مجموعات كبيرة من الطحالب الخضراء والحمراء. الدينوفلاجيلات. تُسمى الطحالب التي تنتمي إلى شعبة Dinoflagelata بالطحالب النارية (Pyrrhophyta) لقدرتها على التلألؤ البيولوجي - ظاهرة التلألؤ، أو انبعاث الضوء المرئي.
معظم الطحالب النارية هي عبارة عن أشكال أحادية الخلية تحتوي على سوطين، وغالبًا ما تكون في أشكال معقدة ومتنوعة للغاية، مع صفائح السليلوز الكثيفة التي تشكل جدارًا خلويًا يشبه الخوذة أو الدرع. بعضها كبير جدًا، يصل قطره إلى 2 مم. ونظرًا لوجود عدد كبير من الخلايا الطبقية (الحويصلات الهوائية) تحت الغشاء البلازمي، تنتمي هذه الطحالب إلى مجموعة خاصة من الطلائعيات - الحويصلات.
تحتوي الدينوفلاجيلات الضوئية عادةً على الكلوروفيل a وc، بالإضافة إلى الكاروتينات، وتكون خلاياها ملونة باللون الذهبي أو البني. المادة الاحتياطية هي النشا. غالبًا ما تدخل هذه الطحالب في علاقات تكافلية مع الكائنات البحرية (الإسفنج وقناديل البحر وشقائق النعمان البحرية والمرجان والحبار وغيرها). وفي هذه الحالة تفقد صفائحها السليلوزية وتظهر كخلايا كروية ذهبية تسمى زوزانثيلا. في مثل هذه الأنظمة التكافلية، يزود الحيوان الدينوفلاجيلات بثاني أكسيد الكربون الذي تحتاجه لعملية التمثيل الضوئي ويوفر الحماية، بينما تزود الطحالب الحيوان بالأكسجين والمواد العضوية.
الطريقة الرئيسية للتكاثر اللاجنسي للدينوفلاجيلات هي التقسيم الطولي، وتكوين الأبواغ الحيوانية أقل شيوعًا. بعض الأنواع قادرة على التكاثر الجنسي أثناء الزواج المتساوي، وفي بعض الأحيان متباين الزواج.
من المعروف أن حوالي 2000 نوع من الدينوفلاجيلات الحية تعيش في أغلب الأحيان في البحار، وفي كثير من الأحيان في مسطحات المياه العذبة. ممثلو التمثيل الضوئي من هذا النوع هم مكونات عالية الإنتاجية للعوالق البحرية، ومع ذلك، فهي قادرة على التسبب في تفشي الأمراض على نطاق واسع وموت الأسماك والمحاريات والحيوانات الأخرى. ويفسر ذلك بالتطور السريع غير المعتاد لبعض الطحالب النارية القادرة على إنتاج السموم التي تعد من بين السموم العصبية القوية. ونتيجة لذلك، تلحق أضرار جسيمة بمصايد الأسماك البحرية، وبالإضافة إلى ذلك، يتعرض الأشخاص الذين يأكلون الأسماك أو المحار التي أكلت الطحالب السامة للتسمم.
قسم الكلوروفيتا - الطحالب الخضراء:
قسم الطحالب الأكثر شمولاً في هذا الوقت. ووفقا للتقديرات التقريبية، يشمل ذلك حوالي 500 جنس ومن 13000 إلى 20000 نوع. تتميز جميعها في المقام الأول باللون الأخضر النقي لثاليها، المشابه للون النباتات العليا والناجمة عن غلبة الكلوروفيل على الأصباغ الأخرى. نطاق أحجامها كبير جدًا أيضًا - من عدة ميكرونات إلى عدة أمتار.
الأصباغ السائدة في البلاستيدات الخضراء (كما هو الحال في النباتات) هي الكلوروفيل أ و ب، ولهذا السبب يتم تلوين الثالي باللون الأخضر. تشكل الكاروتينات الموجودة في البلاستيدات الخضراء للعديد من الطحالب الخضراء أحادية الخلية تراكمًا على شكل "عين" (وصمة عار). تحتوي العديد من الأنواع على فجوات مقلصة في خلاياها تشارك في تنظيم التناضح. عادة ما تكون الأشكال وحيدة الخلية متحركة بسبب وجود سوطين متطابقين، ولا يتم تغطية الأسواط بفروع أنبوبية، كما هو الحال في الكروميين.
المادة الاحتياطية الرئيسية للطحالب الخضراء هي النشا، وتتكون جدران الخلايا لمعظم الأنواع من السليلوز. هذه الخصائص، بالإضافة إلى التركيب الكيميائي لأصباغ التمثيل الضوئي وبعض السمات الهيكلية للعناصر الخلوية الفردية، تجعل الطحالب الخضراء مشابهة جدًا للنباتات. بالإضافة إلى ذلك، مثل النباتات، تشهد الطحالب الخضراء تغيرًا عبر الأجيال في دورة حياتها. يتيح لنا هذا التشابه اعتبار الطحالب الخضراء بمثابة الأسلاف المباشرين للنباتات البرية. أظهرت دراسة الرنا الريباسي الصغير أن الممثلين الفرديين لهذه المجموعة، وخاصة الطحالب الكاروفيتية، هم أقرب في درجة العلاقة التطورية بالنباتات من الطحالب الأخرى.
يمكن أن يكون تكاثر الطحالب الخضراء نباتيًا ولا جنسيًا وجنسيًا.
تنتشر الطحالب الخضراء على نطاق واسع في جميع أنحاء العالم. يمكن العثور على معظمها في المسطحات المائية العذبة (ممثلي الكاروفيت والكلوروفيسيا)، ولكن هناك العديد من أشكال المياه المالحة والبحرية (معظم ممثلي فئة ulvophyceae). من بينها أشكال العوالق والبيريفيتون والقاعية. هناك طحالب خضراء تكيفت مع الحياة في التربة والموائل الأرضية. يمكن العثور عليها على لحاء الأشجار والصخور والمباني المختلفة وعلى سطح التربة وفي الهواء. يؤدي التطور الهائل للطحالب الخضراء المجهرية إلى "ازدهار" الماء والتربة والثلج ولحاء الأشجار وما إلى ذلك.
قسم يوجلينوفيتا - يوجلينوفيتس:
في اليوجليناسيا، يختلف شكل الجسم من الشكل المغزلي والبيضاوي إلى الورقة المسطحة والشكل الإبري. يتم تقريب الطرف الأمامي من الجسم إلى حد ما، ويمكن تمديد الطرف الخلفي وينتهي بعملية مدببة. يمكن أن تكون الخلايا ملتوية حلزونيا. طول الخلية من 5 إلى 500 ميكرون أو أكثر.
تمتلك الأوغلينية 1، 2، 3، 4 و 7 أسواط مرئية، باستثناء مجموعة صغيرة من الأشكال عديمة السوط، بالإضافة إلى الكائنات الحية المرتبطة بها. تمتد السوط من غزو على شكل قارورة في الطرف الأمامي للخلية - البلعوم (أمبولات).
يتكون نظام الأوجلينويات الحساس للضوء من بنائين. المكون الأول هو الجسم المجاور للسوط (التورم المجاور للقاعدة)، وهو عبارة عن تورم في قاعدة سوط واحد مرئي ويحتوي على فلافينات زرقاء حساسة للضوء. المكون الثاني للنظام هو العينة (وصمة العار)، الموجودة في السيتوبلازم بالقرب من الخزان المقابل للجسم شبه السوطي.
تتميز طحالب الحنديرة بالتغذية الذاتية وغير المتجانسة (الرطبة). في الحالة الأخيرة، تدخل العناصر الغذائية إلى الخلية في شكل مذاب، ويتم امتصاصها من خلال سطحها بالكامل (النوع التناضحي). تتميز بعض الأنواع أيضًا بنظام التغذية البلعمي. هناك ممثلون معروفون للعوز الغذائي لليوجلينا، يعتمدون على فيتامينات ب12 وب.
إذا تمت زراعة الأوجلينا لفترة طويلة في وسط غذائي مناسب في الظلام، فإنها يمكن أن تفقد البلاستيدات الخضراء وتظهر نوعًا من التغذية غير المتجانسة إلى أجل غير مسمى، ولا يختلف في هذه الحالة عن الأوليات. وبالتالي، يمكن اعتبار الأوجلينا من الأوليات ذات وراثة البلاستيدات الخضراء غير المستقرة.
تعيش طحالب اليوجلينا بشكل رئيسي في المياه العذبة، وتفضل المسطحات المائية ذات التدفق البطيء والمحتوى الغني بالمواد العضوية. ويمكن العثور عليها في المناطق الساحلية من البحيرات والأنهار، وفي المسطحات المائية الصغيرة، بما في ذلك البرك، وفي حقول الأرز، وعلى التربة الرطبة. في التربة، تم العثور على ممثلين عديمي اللون على عمق 8-25 سم، ويمكن أن تتسبب الأوجلينويات الملونة في ازدهار الماء وتشكيل طبقة خضراء أو حمراء على سطحها.
إلى حد كبير، تستجيب طحالب الأوجلينا لدرجة تمعدن الماء: فكلما ارتفعت، كلما كان تركيبها النوعي والكمي أضعف. يمكن للبعض تحمل المياه عالية الملوحة.
من بين النباتات اليوجلينية هناك كائنات ذاتية التغذية ضوئية، ومغايرة التغذية (بلعمية التغذية وسابروفتية) ومختلطة التغذية. ثلث الأجناس فقط قادر على القيام بعملية التمثيل الضوئي، والباقي عبارة عن كائنات مغذية وأوسموتروف. حتى euglenaceae الضوئية قادرة على النمو غير المتجانس. معظم الأشكال غيرية التغذية هي كائنات سابروفت، تمتص العناصر الغذائية الذائبة في الماء.
قسم Dinophyta - الطحالب dinophyte:
معظم الممثلين عبارة عن سوطات متناظرة أو غير متناظرة على المستوى الثنائي مع غلاف داخل الخلايا متطور.
تتكاثر بالطرق الخضرية وغير الجنسية والجنسية.
– مملكة باجريانيكي الفرعية (رودوبيونتا):
قسم رودوفيتا - الطحالب الحمراء:
عادة ما تكون هذه نباتات كبيرة جدًا، ولكن توجد أيضًا نباتات مجهرية. من بين الطحالب الحمراء هناك أشكال أحادية الخلية (نادرة للغاية)، خيطية وكاذبة، ولكن لا توجد أشكال حمة حقيقية. تشير البقايا الأحفورية إلى أن هذه مجموعة نباتات قديمة جدًا. عادة ما تكون هذه نباتات كبيرة جدًا، ولكن توجد أيضًا نباتات مجهرية.
تتمتع الطحالب الحمراء بدورة تطور معقدة غير موجودة في الطحالب الأخرى.
يشمل قسم الطحالب الحمراء (Rhodophyta) الأنواع التي تحتوي خلاياها على فئة خاصة من أصباغ التمثيل الضوئي - phycobilins (phycocyanin و phycoerythrin)، والتي تمنحها اللون الأحمر (وبالتالي تسمى الطحالب الأرجوانية). تخفي هذه الأصباغ الإضافية لون صبغة التمثيل الضوئي الرئيسية، الكلوروفيل أ. المادة الاحتياطية السائدة للفطر القرمزي هي عديد السكاريد الشبيه بالنشا. تحتوي جدران خلايا هذه الطحالب على السليلوز أو السكريات الأخرى المضمنة في مصفوفة مخاطية، والتي بدورها ممثلة بالأجار أو الكاراجينان. هذه المكونات تجعل الطحالب الحمراء مرنة وزلقة عند اللمس. يقوم بعض العث الأرجواني بترسيب كربونات الكالسيوم في خلاياه، مما يمنحها الصلابة. تلعب مثل هذه الأشكال دورًا مهمًا في تكوين الشعاب المرجانية.
لا تحتوي الطحالب الحمراء على سوط، ويعيش معظمها أسلوب حياة مستقر، مرتبطًا بالحجارة أو الطحالب الأخرى.
في بحر بارنتس، تعد الطحالب الحمراء ممثلين نموذجيين للنباتات القاعية الساحلية.
تؤكل بعض أنواع الطحالب الحمراء. يتم أيضًا الحصول على عامل التبلور أجار أجار من الطحالب الحمراء.
طحلب اخضر مزرق(السيانوفيتا)، البنادق، بشكل أكثر دقة، بنادق فيكوكروم(Schizophyceae)، الطحالب الغروية (Myxophyceae) - كم عدد الأسماء المختلفة التي تلقتها هذه المجموعة من النباتات ذاتية التغذية القديمة من الباحثين! ولم تهدأ العواطف حتى يومنا هذا. هناك العديد من العلماء المستعدين لاستبعاد اللون الأخضر المزرق من عدد الطحالب وبعضها من المملكة النباتية تمامًا. وليس "بسخفة" بل بثقة تامة أنهم يفعلون ذلك على أساس علمي جدي. والطحالب الخضراء المزرقة هي "المسؤولة" عن هذا المصير. البنية الفريدة للغاية للخلايا والمستعمرات والخيوط، والبيولوجيا المثيرة للاهتمام، والعمر التطوري الكبير - كل هذه الخصائص، منفصلة ومجمعة، توفر الأساس للعديد من التفسيرات لمنهجيات هذه المجموعة من الكائنات الحية.
لا شك أن الطحالب الخضراء المزرقة هي أقدم مجموعة بين الكائنات ذاتية التغذية وبين الكائنات الحية بشكل عام. تم العثور على بقايا كائنات حية مماثلة لها بين الستروماتوليت (تكوينات جيرية ذات سطح درني وبنية داخلية ذات طبقات متحدة المركز من رواسب عصر ما قبل الكمبري)، والتي كان عمرها حوالي ثلاثة مليارات سنة. وكشف التحليل الكيميائي عن وجود منتجات تحلل الكلوروفيل في هذه المخلفات. الدليل الخطير الثاني على قدم الطحالب الخضراء المزرقة هو بنية خلاياها. يتم دمجها مع البكتيريا في مجموعة واحدة تسمى الكائنات ما قبل النووية(بروكاريوتا). يقوم علماء التصنيف المختلفون بتقييم رتبة هذه المجموعة بشكل مختلف - من فئة إلى مملكة مستقلة من الكائنات الحية، اعتمادًا على الأهمية التي يعلقونها على الخصائص الفردية أو مستوى البنية الخلوية. لا يزال هناك قدر كبير من عدم اليقين في تصنيف الطحالب الخضراء المزرقة، وتنشأ خلافات كبيرة على كل مستوى من مستويات أبحاثهم.
توجد الطحالب الخضراء المزرقة في جميع أنواع الموائل، ويكاد يكون من المستحيل وجودها، في جميع القارات والمسطحات المائية على الأرض.
بنية الخلية.بناءً على شكل خلاياها النباتية، يمكن تقسيم الطحالب الخضراء المزرقة إلى مجموعتين رئيسيتين:
1) الأنواع التي تحتوي على خلايا كروية أكثر أو أقل (كروية، إهليلجية واسعة النطاق، كمثرية وبيضاوية)؛
2) الأنواع التي تحتوي على خلايا ممدودة بقوة (أو مضغوطة) في اتجاه واحد (إهليلجي ممدود، مغزلي، أسطواني - من أسطواني قصير وبرميلي إلى أسطواني ممدود). تعيش الخلايا بشكل منفصل، وأحيانًا تنضم معًا في مستعمرات أو تشكل خيوطًا (يمكن أن تعيش الأخيرة أيضًا بشكل منفصل أو تشكل خصلًا أو مستعمرات جيلاتينية).
الخلايا لها جدران سميكة إلى حد ما. في جوهرها، يحيط بالبروتوبلاست هنا أربع طبقات من الغشاء: غشاء الخلية المكون من طبقتين مغطى في الأعلى بغشاء متموج خارجي، وبين البروتوبلاست والغشاء يوجد أيضًا غشاء خلية داخلي. فقط الطبقة الداخلية من القشرة والغشاء الداخلي تشارك في تكوين القسم العرضي بين الخلايا في الخيوط؛ الغشاء الخارجي والطبقة الخارجية للقشرة لا تصل إلى هناك.
تمت دراسة بنية جدار الخلية والبنى المجهرية الأخرى لخلايا الطحالب الخضراء المزرقة باستخدام المجهر الإلكتروني (الشكل 49).
على الرغم من أن غشاء الخلية يحتوي على السليلوز، إلا أن الدور الرئيسي يلعبه مواد البكتين والسكريات المخاطية. في بعض الأنواع، تكون أغشية الخلايا مخاطية جيدًا وتحتوي حتى على أصباغ؛ وفي حالات أخرى، يتم تشكيل غلاف مخاطي خاص حول الخلايا، وأحيانًا يكون مستقلاً حول كل خلية، ولكن في أغلب الأحيان يندمج في غلاف مشترك يحيط بمجموعة أو صف كامل من الخلايا، يسمى في الأشكال الخيطية بمصطلح خاص - trichomes. في العديد من الطحالب الخضراء المزرقة، تكون الأشكال ثلاثية الألوان محاطة بأغماد حقيقية - أغلفة. تتكون كل من الحافظات الخلوية والحقيقية من ألياف رقيقة متشابكة. يمكن أن تكون متجانسة أو ذات طبقات: يمكن أن تكون طبقات الخيوط ذات القواعد والقمة المنفصلة متوازية أو مائلة، وأحيانًا على شكل قمع. تنمو الحالات الحقيقية عن طريق وضع طبقات جديدة من المخاط فوق بعضها البعض أو إدخال طبقات جديدة بين الطبقات القديمة. بعض نوستوسي(Nostoc، Anabaena) تتشكل أغلفة الخلايا عن طريق إفراز المخاط من خلال المسام الموجودة في الأغشية.
تفتقر البروتوبلاست للطحالب الخضراء المزرقة إلى نواة مشكلة وكانت تعتبر في السابق منتشرة، وتنقسم فقط إلى جزء محيطي ملون - الكروماتوبلازم - وجزء مركزي عديم اللون - السيتوبلازم. ومع ذلك، فقد أثبتت طرق مختلفة من الفحص المجهري والكيمياء الخلوية، وكذلك الطرد المركزي الفائق، أن مثل هذا التقسيم لا يمكن أن يكون مشروطًا إلا. تحتوي خلايا الطحالب الخضراء المزرقة على عناصر هيكلية محددة جيدًا، ويحدد ترتيبها المختلف الاختلافات بين السينتروبلازم والكروماتوبلازم. يميز بعض المؤلفين الآن ثلاثة مكونات في البروتوبلاست للطحالب الخضراء المزرقة:
1) النواة.
2) لوحات التمثيل الضوئي (الصفائح)؛
3) الريبوسومات والحبيبات السيتوبلازمية الأخرى.
ولكن بما أن النيوكليوبلازم يحتل منطقة البلازمة المركزية، وتقع الصفائح والمكونات الأخرى في منطقة الكروماتوبلازما التي تحتوي على أصباغ، فإن التمييز الكلاسيكي القديم (توجد الريبوسومات في كلا جزأين البروتوبلاست) لا يمكن اعتباره خطأ.
يتم توطين الأصباغ المركزة في الجزء المحيطي من البروتوبلاست في تكوينات تشبه الصفائح - صفائح، والتي توجد في الكروماتوبلازم بطرق مختلفة: بشكل فوضوي، ومعبأة في حبيبات أو موجهة شعاعيًا. تسمى هذه الأنظمة من الصفائح الآن في كثير من الأحيان بـ parachromatophores.
في الكروماتوبلازما، بالإضافة إلى الصفائح والريبوسومات، هناك أيضًا البلاستيدات الخارجية (حبيبات السيانوفيسين التي تتكون من البروتينات الدهنية) وأنواع مختلفة من البلورات. اعتمادًا على الحالة الفسيولوجية وعمر الخلايا، يمكن لجميع هذه العناصر الهيكلية أن تتغير بشكل كبير حتى تختفي تمامًا.
يتكون المركز المركزي لخلايا الطحالب الخضراء المزرقة من الهيالوبلازم وقضبان مختلفة ولييفات وحبيبات. والأخيرة عبارة عن عناصر كروماتين ملطخة بالأصباغ النووية. يمكن عمومًا اعتبار عناصر الهيالوبلازم والكروماتين نظيرًا للنواة، نظرًا لأن هذه العناصر تحتوي على الحمض النووي؛ أثناء انقسام الخلايا، تنقسم طوليًا، ويتم توزيع النصفين بالتساوي بين الخلايا الوليدة. ولكن، على عكس النواة النموذجية، في خلايا الطحالب الخضراء المزرقة، لا يمكن أبدًا اكتشاف الغشاء النووي والنواة حول عناصر الكروماتين. هذا تكوين يشبه النواة في الخلية ويسمى النواة النووية. كما أنه يحتوي على ريبوسومات تحتوي على الحمض النووي الريبي (RNA) والفجوات وحبيبات متعدد الفوسفات.
لقد ثبت أن الأشكال الخيطية لها بلازموديسماتا بين الخلايا. في بعض الأحيان تكون أنظمة صفائح الخلايا المجاورة مترابطة أيضًا. لا ينبغي بأي حال من الأحوال اعتبار الحاجز المستعرض في المشعرات قطعًا من المادة الميتة. هذا هو المكون الحي للخلية، والذي يشارك باستمرار في عملياتها الحياتية، مثل محيط الكائنات الحية ذات السوط.
بروتوبلازم الطحالب الخضراء المزرقة أكثر كثافة من مجموعات النباتات الأخرى. وهي غير متحركة ونادرا ما تحتوي على فجوات مملوءة بعصارة الخلية. تظهر الفجوات في الخلايا القديمة فقط، ويؤدي حدوثها دائمًا إلى موت الخلايا. ولكن في خلايا الطحالب الخضراء المزرقة غالبًا ما توجد فجوات غازية (فراغات كاذبة). هذه تجاويف في البروتوبلازم، مملوءة بالنيتروجين وتعطي الخلية لونًا أسود-بني أو أسود تقريبًا في الضوء المنقول للمجهر. توجد في بعض الأنواع بشكل شبه دائم، ولكن هناك أيضًا أنواع لا يتم العثور عليها فيها. غالبًا ما يعتبر وجودها أو غيابها شخصية مهمة من الناحية التصنيفية، ولكن بالطبع ما زلنا لا نعرف كل شيء عن فجوات الغاز. غالبًا ما توجد في خلايا الأنواع التي تقود نمط حياة العوالق (ممثلو أجناس Anabaena و Aphanizomenon و Rivularia و Microcystis وما إلى ذلك ، الشكل 50 ، 58.1).
 ,
, 
ولا شك أن الفجوات الغازية الموجودة في هذه الطحالب تعمل كنوع من التكيف لتقليل جاذبيتها النوعية، أي لتحسين "الطفو" في عمود الماء. ومع ذلك، فإن وجودها ليس ضروريًا على الإطلاق، وحتى في الألواح الخشبية النموذجية مثل Microcystis aeruginosa وM. flosaquae، يمكن للمرء أن يلاحظ (خاصة في الخريف) الاختفاء شبه الكامل للفجوات الغازية. وفي بعض الأنواع تظهر وتختفي فجأة، وغالبًا لأسباب غير معروفة. ش نوستوك البرقوق(Nostoc pruniforme، الجداول 3، 9)، وهي مستعمرات كبيرة تعيش دائمًا في قاع الخزانات؛ وتظهر في الظروف الطبيعية في الربيع، بعد وقت قصير من ذوبان الجليد. عادة، تكتسب المستعمرات ذات اللون البني المخضر لونًا رماديًا، وأحيانًا حليبيًا، وتختفي تمامًا في غضون أيام قليلة. يُظهر الفحص المجهري للطحالب في هذه المرحلة أن جميع الخلايا الأنفية مملوءة بفجوات غازية (الشكل 50) وقد أصبحت ذات لون بني مسود، على غرار خلايا الأنابينات العوالق. اعتمادًا على الظروف، تستمر الفجوات الغازية لمدة تصل إلى عشرة أيام ولكنها تختفي في النهاية؛ يبدأ تكوين غلاف مخاطي حول الخلايا ويبدأ انقسامها المكثف. كل خيط أو حتى قطعة خيط تؤدي إلى ظهور كائن حي جديد (مستعمرة). يمكن ملاحظة صورة مماثلة أثناء إنبات جراثيم الأنواع النبتية أو العوالق من جليوترشيا. في بعض الأحيان تظهر فجوات الغاز فقط في بعض خلايا المشعرات، على سبيل المثال في منطقة ميريستيمالب، حيث يحدث انقسام الخلايا المكثف ويمكن أن تنشأ الهرمونات، والتي تساعد فجوات الغاز بطريقة أو بأخرى على إطلاقها.
 ,
,
تتشكل فجوات الغاز على حدود الكروماتوبلازما والمركزية وتكون غير منتظمة الشكل تمامًا. في بعض الأنواع التي تعيش في الطبقات العليا من الطمي السفلي (في السابروبيل)، وخاصة في الأنواع الذبذباتتوجد فجوات غازية كبيرة في الخلايا على جوانب الأقسام العرضية. وقد ثبت تجريبياً أن ظهور مثل هذه الفجوات ناتج عن انخفاض كمية الأكسجين المذاب في الوسط مع إضافة منتجات تخمير كبريتيد الهيدروجين إلى الوسط. يمكن الافتراض أن مثل هذه الفجوات تنشأ كمرافق تخزين أو مواقع لترسيب الغازات التي يتم إطلاقها أثناء العمليات الأنزيمية التي تحدث في الخلية.
إن تكوين جهاز الصباغ للطحالب الخضراء المزرقة متنوع للغاية، حيث تم العثور على حوالي 30 صبغة مختلفة داخل الخلايا. وهي تنتمي إلى أربع مجموعات - الكلوروفيل، الكاروتينات، الزانثوفيل والبروتينات الصفراوية. من بين الكلوروفيل، تم إثبات وجود الكلوروفيل أ بشكل موثوق حتى الآن؛ من الكاروتينات - α، β و ε- كاروتين؛ من الزانثوفيل - إشينيون، زياكسانثين، كريبتوكسايتين، ميكسوكسانثوفيل، وما إلى ذلك، ومن البروتينات البيليبروتين - سي فيكوسيانين، سي فيكو إريثرين وألوفيكوسيانين. من المعتاد جدًا أن تحتوي الطحالب الخضراء المزرقة على المجموعة الأخيرة من الأصباغ (توجد أيضًا في الطحالب الأرجوانية وبعض الكريبتومونات) وغياب الكلوروفيل ب. يشير الأخير مرة أخرى إلى أن الطحالب الخضراء المزرقة هي مجموعة قديمة انفصلت واتبعت مسارًا مستقلاً للتطور حتى قبل ظهور الكلوروفيل ب أثناء التطور، والذي تعطي مشاركته في التفاعلات الكيميائية الضوئية لعملية التمثيل الضوئي أعلى كفاءة.
يفسر التنوع والتركيب الفريد لأنظمة الصباغ الممتصة ضوئيًا مقاومة الطحالب الخضراء المزرقة لتأثيرات السواد لفترات طويلة واللاهوائية. وهذا ما يفسر جزئيًا وجودها في ظروف معيشية قاسية - في الكهوف، والطبقات الغنية بكبريتيد الهيدروجين من الطمي السفلي، وفي الينابيع المعدنية.
ناتج عملية التمثيل الضوئي في خلايا الطحالب الخضراء هو بروتين سكري ينشأ في الكروماتوبلازما ويترسب هناك. يشبه البروتين السكري الجليكوجين - من محلول اليود في يوديد البوتاسيوم يتحول إلى اللون البني. تم العثور على حبيبات السكاريد بين صفائح التمثيل الضوئي. تتكون حبيبات السيانوفيسين الموجودة في الطبقة الخارجية للكروماتوبلازم من البروتينات الدهنية. حبيبات الفولوتين الموجودة في السينتروبلازم هي مواد تخزين ذات أصل بروتيني. تظهر حبيبات الكبريت في بلازما سكان الخزانات الكبريتية.
يمكن أن يفسر تنوع تركيبة الصباغ أيضًا تنوع ألوان الخلايا والأشكال ثلاثية الألوان للطحالب الخضراء المزرقة. ويتنوع لونها من الأزرق والأخضر النقي إلى البنفسجي أو المحمر، وأحيانًا إلى الأرجواني أو الأحمر البني، ومن الأصفر إلى الأزرق الشاحب أو الأسود تقريبًا. يعتمد لون البروتوبلاست على الموقع المنهجي للأنواع، وكذلك على عمر الخلايا والظروف المعيشية. في كثير من الأحيان يتم حجبه بواسطة لون المخاط المهبلي أو المخاط الاستعماري. توجد أصباغ أيضًا في المخاط وتعطي الخيوط أو المستعمرات لونًا أصفر أو بني أو محمر أو أرجواني أو أزرق. يعتمد لون المخاط بدوره على الظروف البيئية - على الضوء والكيمياء ودرجة الحموضة في البيئة، وعلى كمية الرطوبة في الهواء (للنباتات الهوائية).
هيكل المواضيع.ينمو عدد قليل من الطحالب الخضراء المزرقة كخلايا فردية، وينمو معظمها في مستعمرات أو خيوط متعددة الخلايا. في المقابل، يمكن للخيوط إما أن تشكل مستعمرات متنية كاذبة، حيث تكون مغلقة بشكل وثيق، وتحتفظ الخلايا بالاستقلال الفسيولوجي، أو يكون لها بنية متجانسة، حيث ترتبط الخلايا على التوالي، وتشكل ما يسمى trichomes. في المشعرات، ترتبط البروتوبلاستات للخلايا المجاورة عن طريق البلازموديسماتا. تسمى الأشكال ثلاثية الألوان، المحاطة بغشاء مخاطي، بالخيوط.
يمكن أن تكون الأشكال الخيطية بسيطة أو متفرعة. إن التفرع في الطحالب الخضراء المزرقة ذو شقين - حقيقي وكاذب (الشكل 51). يسمى هذا التفرع عندما ينشأ فرع جانبي نتيجة انقسام خلية واحدة بشكل عمودي على الخيط الرئيسي (رتبة Stigonematales). التفرع الكاذب هو تكوين فرع جانبي عن طريق تمزيق المشعرات وكسرها عبر المهبل إلى الجانب عند أحد الطرفين أو كليهما. في الحالة الأولى يتحدثون عن واحد، في الثانية - حول التفرع الخاطئ المزدوج (أو المقترن). يمكن اعتبار التفرع الكاذب سمة متفرعة على شكل حلقة لعائلة Scytonemataceae والتفرع النادر على شكل حرف V - نتيجة الانقسام المتكرر ونمو خليتين ثلاثيتي الألوان متجاورتين في اتجاهين متعاكسين بالنسبة للمحور الطويل للخيط.
تحتوي العديد من الطحالب الخضراء المزرقة الخيطية على خلايا غريبة تسمى الكيسات غير المتجانسة. لديهم غلاف من طبقتين محدد جيدًا، وتكون المحتويات دائمًا خالية من أصباغ الاستيعاب (عديم اللون أو مزرق أو مصفر)، وفجوات الغاز وحبوب المواد الاحتياطية. تتشكل من خلايا نباتية في أماكن مختلفة من المشعرات، اعتمادًا على الموقع المنهجي للطحالب: على طرفي المشعرات (Rivularia، Calothrix، Gloeotrichia) وكلاهما (Anabaenopsis، Cylindrospermum) - القاعدية والمحطة؛ في المشعرة بين الخلايا الخضرية، أي المقحم (Nostoc، Anabaena، Nodularia) أو على جانب المشعرة - بشكل جانبي (في بعض Stigonematales). تم العثور على الكيسات غير المتجانسة منفردة أو عدة (2-10) على التوالي. اعتمادًا على الموقع، تظهر سدادة واحدة (في الكيسة غير المتجانسة الطرفية والجانبية) أو اثنتين، وأحيانًا حتى ثلاثة (في الكيسة غير المتجانسة) في كل كيسة غير متجانسة، والتي تسد المسام من الداخل بين الكيسة غير المتجانسة والخلايا النباتية المجاورة (الشكل 5، 2). .

تسمى الأكياس غير المتجانسة بالغموض النباتي. تحت المجهر الضوئي، تبدو فارغة، ولكن في بعض الأحيان، ولمفاجأة كبيرة للباحثين، تنبت فجأة، مما يؤدي إلى ظهور أشكال ثلاثية الألوان جديدة. أثناء التفرع الكاذب وأثناء فصل الخيوط، غالبًا ما تنفجر trichomes بالقرب من الأكياس غير المتجانسة، كما لو كان نمو trichomes محدودًا بها. ولهذا السبب، كانت تسمى سابقًا الخلايا الحدودية. يتم ربط الخيوط ذات الأكياس غير المتجانسة القاعدية والطرفية بالركيزة باستخدام الأكياس غير المتجانسة. في بعض الأنواع، ترتبط الكيسات غير المتجانسة بتكوين خلايا الراحة - الجراثيم: فهي تقع بجوار الكيسة غير المتجانسة، واحدة تلو الأخرى (في Culindrospermum، Gloeotrichia، Anabaenopsis raciborskii) أو على كلا الجانبين (في بعض Anabaena). من الممكن أن تكون الأكياس غير المتجانسة عبارة عن مستودعات لبعض المواد الاحتياطية أو الإنزيمات. ومن المثير للاهتمام أن نلاحظ أن جميع أنواع الطحالب الخضراء المزرقة القادرة على تثبيت النيتروجين في الغلاف الجوي تحتوي على أكياس غير متجانسة.
التكاثر.النوع الأكثر شيوعًا للتكاثر في الطحالب الخضراء المزرقة هو انقسام الخلايا إلى قسمين. بالنسبة للأشكال وحيدة الخلية، فهذه هي الطريقة الوحيدة؛ في المستعمرات والخيوط يؤدي إلى نمو الخيوط أو المستعمرة.
تتشكل الأشكال ثلاثية الألوان عندما لا تتحرك الخلايا المنقسمة في نفس الاتجاه بعيدًا عن بعضها البعض. إذا تم انتهاك الترتيب الخطي، تظهر مستعمرة ذات خلايا مرتبة عشوائيًا. عند الانقسام في اتجاهين متعامدين في مستوى واحد، تتشكل مستعمرة صفائحية بترتيب منتظم للخلايا على شكل رباعيات (Merismopedia). تحدث التجمعات الحجمية على شكل حزم عندما تنقسم الخلايا إلى ثلاث مستويات (Eucapsis).
يتميز ممثلو بعض الأجناس (Gloeocapsa، Microcystis) أيضًا بالانقسام السريع مع تكوين العديد من الخلايا الصغيرة - الخلايا النانوية - في الخلية الأم.
تتكاثر الطحالب الخضراء المزرقة أيضًا بطرق أخرى - عن طريق تكوين الجراثيم (خلايا الراحة)، والأبواغ الخارجية والأندوسبورات، والهرمونات، والأبواغ الهرمونية، والجونيديا، والمكورات، والمكورات المسطحة. أحد أكثر أنواع تكاثر الأشكال الخيطية شيوعًا هو تكوين الهرمونات. تعتبر طريقة التكاثر هذه مميزة جدًا لبعض الطحالب الخضراء المزرقة لدرجة أنها كانت بمثابة اسم للفئة بأكملها هرمونيوم(الهرمونية). يُطلق على الهرمونات عادةً اسم أجزاء من الأشكال ثلاثية الألوان التي تنقسم إليها الأخيرة. إن تكوين الهورمونيوم ليس مجرد فصل ميكانيكي لمجموعة مكونة من خليتين أو ثلاث أو أكثر. يتم فصل الهرمونات عن طريق موت بعض الخلايا النخرية، ثم بمساعدة إفراز المخاط، فإنها تنزلق من المهبل (إذا كان هناك واحدة)، وتقوم بحركات تذبذبية، وتتحرك في الماء أو على طول الركيزة. كل تجانس يمكن أن يؤدي إلى فرد جديد. إذا كانت مجموعة من الخلايا، على غرار الهورمونيوم، مغطاة بغشاء سميك، يطلق عليها اسم hormospore (hormocyst)، والتي تؤدي في نفس الوقت وظائف التكاثر وتتحمل الظروف غير المواتية.
في بعض الأنواع، يتم فصل شظايا وحيدة الخلية عن الثالوس، والتي تسمى الغونيديا، أو المكورات، أو المكورات المسطحة. تحتفظ الغونيديا بالغشاء المخاطي. تفتقر المكورات إلى أغشية محددة بوضوح؛ المكورات Planococci عارية أيضًا، ولكن، مثل الهرمونات، لديها القدرة على التحرك بنشاط.
لا تزال أسباب حركة الهورمونات والمكورات المسطحة والأشكال ثلاثية الألوان الكاملة (في Oscillatoriaceae) بعيدة عن التوضيح. إنها تنزلق على طول المحور الطولي، وتتأرجح من جانب إلى آخر، أو تدور حوله. تعتبر القوة الدافعة هي إفراز المخاط، وتقلص الترايخوم على طول المحور الطولي، وتقلص الغشاء الخارجي المتموج، وكذلك الظواهر الحركية الكهربائية.
الأعضاء التناسلية الشائعة جدًا هي الجراثيم، خاصة في الطحالب من رتبة Nostocales. وهي وحيدة الخلية، وعادة ما تكون أكبر من الخلايا النباتية وتنشأ منها، عادة من خلية واحدة. ومع ذلك، في ممثلي بعض الأجناس (Gloeotrichia، Anabaena) يتم تشكيلها نتيجة اندماج العديد من الخلايا النباتية، ويمكن أن يصل طول هذه الجراثيم إلى 0.5 ملم. من الممكن أن تحدث إعادة التركيب أيضًا أثناء عملية الدمج هذه، ولكن حتى الآن لا توجد بيانات دقيقة حول هذا الأمر.
تُغطى الجراثيم بغشاء سميك مكون من طبقتين، الطبقة الداخلية منه تسمى الإندوسبوريوم، والطبقة الخارجية تسمى الإكسوسبوريوم. تكون الأصداف ناعمة أو منقطة بالحليمات، عديمة اللون، صفراء أو بنية. بفضل الأصداف السميكة والتغيرات الفسيولوجية في البروتوبلاست (تراكم المواد الاحتياطية، واختفاء أصباغ الاستيعاب، وأحيانًا زيادة عدد حبيبات السيانوفيسين)، يمكن أن تظل الجراثيم قابلة للحياة لفترة طويلة في ظروف غير مواتية وتحت مجموعة متنوعة من التأثيرات القوية. (في درجات حرارة منخفضة وعالية، أثناء التجفيف والإشعاع القوي) . في ظل ظروف مواتية، تنبت الجراثيم، وتنقسم محتوياتها إلى خلايا - يتم تشكيل sporogormogonium، وتخاط القشرة، أو تنكسر أو تفتح بغطاء، ويخرج الهرمون.
تم العثور على Endo- و exospores بشكل رئيسي في الممثلين فئة الحرباء(تشامايسفونوفيسيا). تتشكل الأبواغ الداخلية بأعداد كبيرة (أكثر من مائة) في الخلايا الأم المتضخمة. يحدث تكوينها بنجاح (نتيجة لسلسلة من الانقسامات المتعاقبة للبروتوبلاست للخلية الأم) أو في وقت واحد (من خلال التفكك المتزامن للخلية الأم إلى العديد من الخلايا الصغيرة). تنفصل الأبواغ الخارجية، عند تشكلها، عن البروتوبلاست للخلية الأم وتخرج. في بعض الأحيان لا تنفصل عن الخلية الأم، ولكنها تشكل سلاسل عليها (على سبيل المثال، في بعض أنواع الشامايسفون).
التكاثر الجنسي غائب تمامًا في الطحالب الخضراء المزرقة.
طرق الأكل والبيئة.من المعروف أن معظم الطحالب الخضراء المزرقة قادرة على تصنيع جميع المواد الموجودة في خلاياها باستخدام الطاقة الضوئية. تتشابه عمليات التمثيل الضوئي التي تحدث في خلايا الطحالب الخضراء المزرقة من حيث المبدأ مع العمليات التي تحدث في الكائنات الحية الأخرى التي تحتوي على الكلوروفيل.
يعد نوع التغذية الضوئية هو النوع الرئيسي بالنسبة لهم، ولكنه ليس الوحيد. بالإضافة إلى التمثيل الضوئي الحقيقي، فإن الطحالب الخضراء المزرقة قادرة على الاختزال الضوئي، والتغذية الضوئية، والتغذية الذاتية، والتغذية غير المتجانسة، وحتى التغذية الكاملة. إذا كانت هناك مواد عضوية في البيئة، فإنها تستخدمها كمصادر إضافية للطاقة. نظرًا لقدرتها على التغذية المختلطة (التغذية المختلطة)، فإنها يمكن أن تكون نشطة حتى في الظروف القاسية بالنسبة للحياة ذاتية التغذية. في مثل هذه الموائل لا توجد منافسة تقريبًا، وتحتل الطحالب الخضراء المزرقة موقعًا مهيمنًا.
في ظروف الإضاءة السيئة (في الكهوف، في الآفاق العميقة للخزانات)، يتغير تكوين الصباغ في خلايا الطحالب الخضراء المزرقة. هذه الظاهرة التي تسمى التكيف اللوني هي تغير تكيفي في لون الطحالب تحت تأثير التغيرات في التركيب الطيفي للضوء نتيجة زيادة عدد الصبغات التي لها لون مكمل للون الأشعة الساقطة. تحدث التغيرات في لون الخلية (الكلور) أيضا في حالة عدم وجود مكونات معينة في البيئة، في وجود مواد سامة، وكذلك أثناء الانتقال إلى نوع التغذية غير المتجانسة.
ومن بين الطحالب الخضراء المزرقة هناك أيضًا مجموعة من الأنواع، والتي يكون أمثالها نادرًا بشكل عام بين الكائنات الحية الأخرى. هذه الطحالب قادرة على تثبيت النيتروجين في الغلاف الجوي، ويتم دمج هذه الخاصية مع عملية التمثيل الضوئي. حوالي مائة من هذه الأنواع معروفة الآن. كما سبقت الإشارة، فإن هذه القدرة مميزة فقط للطحالب التي تحتوي على أكياس غير متجانسة، وليس جميعها.
تقتصر معظم الطحالب المثبتة للنيتروجين باللون الأزرق والأخضر على الموائل الأرضية. من المحتمل أن استقلالهم الغذائي النسبي كمثبتات للنيتروجين في الغلاف الجوي هو الذي يسمح لهم باستعمار الصخور غير المأهولة دون أدنى أثر للتربة، كما لوحظ في جزيرة كراكاتوا عام 1883: بعد ثلاث سنوات من الانفجار البركاني، تم العثور على تراكمات مخاطية. على الرماد والطف، تتكون من ممثلي أجناس Anabaena، Gloeocapsa، Nostoc، Calothrix، Phormidium، إلخ. المستوطنون الأوائل لجزيرة سورتسي، التي نشأت نتيجة ثوران بركان تحت الماء عام 1963 بالقرب من الجنوب ساحل أيسلندا، كانوا أيضًا مثبتين للنيتروجين. من بينها بعض أنواع العوالق المنتشرة على نطاق واسع والتي تسبب ازدهار الماء (Anabaena circinalis، A. cylindrica، A. flos-aquae، A. lemmermannii، A. scheremievii، A. spiroides، Anabaenopsis Circinalis، Gloeotrichia echinulata).
تعتبر درجة الحرارة القصوى لوجود خلية حية ومستوعبة +65 درجة مئوية، ولكن هذا ليس الحد الأقصى للطحالب الخضراء المزرقة (انظر مقال عن طحالب الينابيع الساخنة). تتحمل الطحالب الخضراء المزرقة المحبة للحرارة درجات الحرارة المرتفعة هذه بسبب الحالة الغروية الغريبة للبروتوبلازم، والتي تتخثر ببطء شديد عند درجات الحرارة المرتفعة. أكثر الحيوانات المحبة للحرارة شيوعًا هي الكوزموبوليتانية Mastigocladus laminosus وPhormidium laminosum. يمكن للطحالب الخضراء المزرقة تحمل درجات الحرارة المنخفضة. تم تخزين بعض الأنواع لمدة أسبوع عند درجة حرارة الهواء السائل (-190 درجة مئوية) دون تلف. لا توجد درجة حرارة كهذه في الطبيعة، ولكن في القارة القطبية الجنوبية، عند درجة حرارة -83 درجة مئوية، تم العثور على الطحالب الخضراء المزرقة (نوستوك) بكميات كبيرة.
في القارة القطبية الجنوبية وفي المرتفعات، بالإضافة إلى درجات الحرارة المنخفضة، يؤثر الإشعاع الشمسي المرتفع أيضًا على الطحالب. للحد من الآثار الضارة للإشعاع قصير الموجة، اكتسبت الطحالب الخضراء المزرقة عددًا من التعديلات أثناء التطور. وأهمها إفراز المخاط حول الخلايا. يعد مخاط المستعمرات والأغشية المخاطية للمهبل الخيطي غلافًا وقائيًا جيدًا يحمي الخلايا من الجفاف ويعمل في نفس الوقت كمرشح يزيل التأثيرات الضارة للإشعاع. اعتمادا على شدة الضوء، يتم ترسيب صبغة أكثر أو أقل في المخاط، ويصبح ملونا في جميع أنحاء سمكه أو طبقاته.
إن قدرة المخاط على امتصاص الماء بسرعة والاحتفاظ به لفترة طويلة تسمح للطحالب الخضراء المزرقة بالنمو بشكل طبيعي في المناطق الصحراوية. يمتص المخاط الحد الأقصى من رطوبة الليل أو الصباح، وتنتفخ المستعمرات، ويبدأ الاستيعاب في الخلايا. بحلول منتصف النهار، تجف المستعمرات الجيلاتينية أو مجموعات الخلايا وتتحول إلى قشور سوداء اللون. ويظلون على هذه الحالة حتى الليلة التالية، عندما يبدأ امتصاص الرطوبة مرة أخرى.
لحياة نشطة، الماء البخاري يكفي لهم.
الطحالب الخضراء المزرقة شائعة جدًا في التربة والمجتمعات الأرضية، وتوجد أيضًا في الموائل الرطبة، وكذلك على لحاء الأشجار، وعلى الحجارة، وما إلى ذلك. غالبًا ما لا يتم تزويد جميع هذه الموائل بالرطوبة باستمرار وتكون مضاءة بشكل غير متساو (لمزيد من التفاصيل، راجع المقالات حول الطحالب الأرضية والتربة).
توجد أيضًا الطحالب الخضراء المزرقة في المجتمعات المحبة للتبريد - على الجليد والثلج. عملية التمثيل الضوئي ممكنة بالطبع فقط عندما تكون الخلايا محاطة بطبقة من الماء السائل، وهو ما يحدث هنا في ضوء الشمس الساطع على الثلج والجليد.
الإشعاع الشمسي على الأنهار الجليدية وحقول الثلج مكثف للغاية، وجزء كبير منه عبارة عن إشعاع قصير الموجة، مما يسبب تكيفات وقائية في الطحالب. تتضمن مجموعة cryobionts عددًا من أنواع الطحالب الخضراء المزرقة، ولكن بشكل عام، يفضل ممثلو هذا القسم الموائل ذات درجات الحرارة المرتفعة (لمزيد من التفاصيل، راجع المقال عن طحالب الثلج والجليد).

تسود الطحالب الخضراء المزرقة في عوالق المسطحات المائية الغنية بالمغذيات، حيث يؤدي تطورها الضخم غالبًا إلى ازدهار الماء. يتم تسهيل نمط حياة العوالق لهذه الطحالب عن طريق فجوات الغاز في الخلايا، على الرغم من أنه ليس كل مسببات الأمراض الزهرية تحتوي عليها (الجدول 4). تعتبر الإفرازات أثناء الحياة ومنتجات التحلل بعد الوفاة لبعض هذه الطحالب الخضراء المزرقة سامة. يبدأ التطور الشامل لمعظم الطحالب الخضراء المزرقة العوالق عند درجات حرارة عالية، أي في النصف الثاني من الربيع والصيف وأوائل الخريف. لقد ثبت أن درجة الحرارة المثلى لمعظم الطحالب الخضراء المزرقة في المياه العذبة تبلغ حوالي +30 درجة مئوية. هناك استثناءات. تتسبب بعض أنواع المذبذبات في "تفتح" الماء تحت الجليد، أي عند درجة حرارة حوالي 0 درجة مئوية. تتطور الأنواع عديمة اللون والمحبة لكبريتيد الهيدروجين بكميات كبيرة في الطبقات العميقة من البحيرات. من الواضح أن بعض مسببات الأمراض المزهرة تتوسع خارج نطاقها بسبب النشاط البشري. وهكذا، لم يتم العثور على أنواع من جنس Anabaenopsis خارج المناطق الاستوائية وشبه الاستوائية لفترة طويلة، ولكن تم العثور عليها بعد ذلك في المناطق الجنوبية من المنطقة المعتدلة، ومنذ عدة سنوات تطورت في خليج هلسنكي. سمحت درجات الحرارة المناسبة وزيادة التخثث (التلوث العضوي) لهذا الكائن بالتطور بأعداد كبيرة شمال خط العرض 60.
"تفتح" المياه بشكل عام، وخاصة بسبب الطحالب الخضراء المزرقة، يعتبر كارثة طبيعية، حيث تصبح المياه غير صالحة لأي شيء تقريبًا. في الوقت نفسه، يزداد التلوث الثانوي والطمي للخزان بشكل ملحوظ، حيث تصل الكتلة الحيوية للطحالب في الخزان "المزهر" إلى قيم كبيرة (متوسط الكتلة الحيوية - ما يصل إلى 200 جم / م 3، الحد الأقصى - ما يصل إلى 450-500 جم / م 3)، ومن بين الطحالب الخضراء المزرقة هناك عدد قليل جدًا من الأنواع التي يمكن أن تستهلكها الكائنات الحية الأخرى كغذاء.
العلاقات بين الطحالب الخضراء المزرقة والكائنات الحية الأخرى متعددة الأوجه. الأنواع من أجناس Gloeocapsa وNostoc وScytonema وStigonema وRivularia وCalothrix هي phycobionts في الأشنات. تعيش بعض الطحالب الخضراء المزرقة في كائنات حية أخرى كممثلات. تعيش أنواع Anabaena و Nostoc في غرف الهواء في طحالب Anthoceros و Blasia. تعيش Anabaena azollae في أوراق سرخس الماء Azolla americana، وفي المساحات بين الخلايا في Cycas وZamia-Nostoc punctiforme (لمزيد من التفاصيل، راجع المقال حول تعايش الطحالب مع الكائنات الحية الأخرى).
وبالتالي، توجد الطحالب الخضراء المزرقة في جميع القارات وفي جميع أنواع الموائل - في الماء وعلى الأرض، في المياه العذبة والمالحة، في أي مكان وفي كل مكان.
ويرى العديد من المؤلفين أن جميع الطحالب الخضراء المزرقة موجودة في كل مكان وعالمية، ولكن هذا أبعد ما يكون عن الواقع. لقد تمت بالفعل مناقشة التوزيع الجغرافي لجنس Anabaenopsis أعلاه. لقد أثبتت الدراسات التفصيلية أنه حتى الأنواع الشائعة مثل Nostoc pruniforme ليست عالمية. تقتصر بعض الأجناس (على سبيل المثال، Nostochopsis، Camptylonemopsis، Raphidiopsis) تمامًا على مناطق ذات مناخات حارة أو دافئة، Nostoc flagelliforme - إلى المناطق القاحلة، والعديد من الأنواع من جنس Chamaesiphon - إلى الأنهار والجداول ذات المياه الباردة والواضحة في البلدان الجبلية.
يعتبر قسم الطحالب الخضراء المزرقة أقدم مجموعة من النباتات ذاتية التغذية على وجه الأرض. إن البنية البدائية للخلية، وغياب التكاثر الجنسي والمراحل السوطية، كلها أدلة جدية على قدمها. من الناحية الخلوية، تشبه الطحالب الخضراء المزرقة البكتيريا، كما توجد بعض أصباغها (البروتينات الصفراوية) أيضًا في الطحالب الحمراء. ومع ذلك، مع الأخذ في الاعتبار مجموعة كاملة من الخصائص المميزة للقسم، يمكن افتراض أن الطحالب الخضراء المزرقة هي فرع مستقل من التطور. منذ أكثر من ثلاثة مليارات سنة، ابتعدوا عن الجذع الرئيسي لتطور النبات وشكلوا فرعًا مسدودًا.
عند الحديث عن الأهمية الاقتصادية للخضراء الزرقاء، يجب وضع المقام الأول على دورها كعوامل مسببة لـ "ازدهار" الماء. وهذا للأسف دور سلبي. تكمن قيمتها الإيجابية في المقام الأول في قدرتها على امتصاص النيتروجين الحر. وفي البلدان الشرقية، تُستخدم الطحالب الخضراء المزرقة كغذاء، وفي السنوات الأخيرة وجد بعضها طريقها إلى أحواض الاستزراع الشامل للإنتاج الصناعي للمواد العضوية.
لا يزال تصنيف الطحالب الخضراء المزرقة بعيدًا عن الكمال. إن البساطة المقارنة للتشكل، والعدد الصغير نسبيًا من الشخصيات ذات القيمة من وجهة نظر علم اللاهوت النظامي والتنوع الواسع لبعضها، فضلاً عن التفسيرات المختلفة لنفس الشخصيات، أدت إلى حقيقة أن جميع الشخصيات الموجودة تقريبًا الأنظمة ذاتية بدرجة أو بأخرى وبعيدة عن أن تكون طبيعية. لا يوجد تمييز جيد ومبرر بين الأنواع ككل، ويتم فهم نطاق الأنواع بشكل مختلف في الأنظمة المختلفة. تم تحديد العدد الإجمالي للأنواع في القسم ليكون 1500-2000. حسب النظام الذي اعتمدناه ينقسم قسم الطحالب الخضراء المزرقة إلى 3 أصناف وعدة رتب وعائلات عديدة.
الموسوعة البيولوجية
علم النظم ومهامه هناك فرع خاص من علم الأحياء يسمى علم النظم، يتعامل مع تصنيف الكائنات الحية وتوضيح علاقاتها التطورية. يسمي بعض علماء الأحياء علم اللاهوت النظامي علم التنوع (التنوع... ... الموسوعة البيولوجية
يعد التكافل، أو التعايش بين كائنين حيين، أحد أكثر الظواهر إثارة للاهتمام وغامضة إلى حد كبير في علم الأحياء، على الرغم من أن دراسة هذه القضية لها تاريخ يمتد إلى قرن تقريبًا. تم اكتشاف ظاهرة التكافل لأول مرة على يد السويسري... الموسوعة البيولوجية
1إفيموفا إم في، إيفيموف أ.أ.
يعرض المقال ويحلل بيانات من بعض المؤلفين حول تصنيف الطحالب الخضراء المزرقة (البكتيريا الزرقاء). تم عرض نتائج تحديد أنواع البكتيريا الزرقاء في بعض الينابيع الساخنة في كامتشاتكا.
هناك ما يصل إلى 1500 نوع من الطحالب الخضراء المزرقة. في مصادر أدبية مختلفة من قبل مؤلفين مختلفين، تم ذكرها تحت أسماء مختلفة: cyanea، cyanobionts، cyanophytes، cyanobacteria، cyanella، الطحالب الخضراء المزرقة، الطحالب الخضراء المزرقة، cyanophyceae. يؤدي تطور البحث إلى قيام بعض المؤلفين بتغيير وجهات نظرهم حول طبيعة هذه الكائنات وبالتالي تغيير الاسم. على سبيل المثال، مرة أخرى في عام 2001، V.N. صنفتها نيكيتينا على أنها طحالب وأطلقت عليها اسم النباتات الزرقاء، وفي عام 2003 حددتها بالفعل على أنها نباتات بدائية النوى. في الأساس، يتم اختيار الاسم وفقًا للتصنيف الذي يفضله هذا المؤلف أو ذاك.
ما الذي يجعل الكائنات الحية من مجموعة واحدة لديها العديد من الأسماء، وأسماء مثل البكتيريا الزرقاء والطحالب الخضراء المزرقة تتناقض مع بعضها البعض؟ بسبب عدم وجود نواة فهي تشبه البكتيريا، وبسبب وجود الكلوروفيل أوالقدرة على تصنيع الأكسجين الجزيئي - مع النباتات. وفقًا لـ إي.جي. كوكا، "البنية الفريدة للغاية للخلايا والمستعمرات والخيوط، والبيولوجيا المثيرة للاهتمام، والعمر التطوري الكبير - كل هذه الميزات ... توفر الأساس للعديد من التفسيرات لعلم النظاميات لهذه المجموعة من الكائنات الحية." يعطي Kukk أسماء مثل الطحالب الخضراء المزرقة ( النباتات الزرقاء) ، بنادق فيكوكروم ( الفصام)، الطحالب الغروية ( ميكسوفيسيا) .
النظاميات هي واحدة من الأساليب الرئيسية لدراسة العالم. هدفها هو البحث عن الوحدة في التنوع المرئي للظواهر الطبيعية. لقد احتلت مشكلة التصنيف في علم الأحياء دائمًا وتحتل مكانة خاصة ترتبط بالتنوع الهائل والتعقيد والتقلب المستمر للأشكال البيولوجية للكائنات الحية. البكتيريا الزرقاء هي أوضح مثال على تعدد الأنظمة.
تعود المحاولات الأولى لبناء نظام اللون الأزرق والأخضر إلى القرن التاسع عشر. (أغارد - 1824، كوتزينغ - 1843، 1849، توريت - 1875). استمر كيرشنر في تطوير النظام (1900). منذ عام 1914، بدأت مراجعة كبيرة للنظام، وتم نشر عدد من الأنظمة الجديدة النباتات الزرقاء(إلينكين - 1916، 1923، 1936؛ بورتسي - 1914، 1916، 1917؛ هيتلر - 1925، 1932). تم التعرف على نظام AA باعتباره الأكثر نجاحًا. إلينكينا، نشرت عام 1936. تم الحفاظ على هذا التصنيف حتى يومنا هذا، حيث أثبت أنه مناسب لعلماء الأحياء المائية وعلماء الحفريات الدقيقة.
كان مخطط مفتاح اتحاد الجمهوريات الاشتراكية السوفياتية لطحالب المياه العذبة يعتمد على نظام Elenkin، الذي تم إجراء تغييرات طفيفة عليه. وفقًا لمخطط المحدد، تم تصنيف اللون الأزرق والأخضر على أنه النباتات الزرقاء، مقسمة إلى ثلاث فئات ( الكروكوكسيا, شاميزيفوني, الهرمونية). تنقسم الفصول إلى أوامر، وأوامر إلى عائلات. يحدد هذا المخطط موقع اللون الأزرق والأخضر في نظام النبات.
وبحسب تصنيف باركر (1982) للطحالب فإن الطحالب الخضراء المزرقة تنتمي إلى المملكة بروكاريوتا، قسم سيانوفيكوتا، فصل الطحالب الزرقاء .
كان القانون الدولي لتسميات النباتات يعتبر غير مقبول بالنسبة لبدائيات النوى، وعلى أساسه تم تطوير القانون الدولي الحالي لتسميات البكتيريا. ومع ذلك، تعتبر البكتيريا الزرقاء كائنات "ثنائية العضو" ويمكن وصفها وفقًا لقواعد كل من ICN والقانون النباتي. في عام 1978، اقترحت اللجنة الفرعية المعنية بالبكتيريا الضوئية التابعة للجنة الدولية لعلم البكتيريا النظامية إخضاع التسمية للتصنيف. النباتات الزرقاءقواعد المدونة الدولية لتسمية البكتيريا، وقبل عام 1985، نشرت قوائم بالأسماء المعتمدة حديثًا لهذه الكائنات. ن.ف. أجرت كوندراتيفا تحليلا نقديا لهذا الاقتراح في مقالتها. ويرى المؤلف أن اقتراح علماء البكتيريا “خاطئ وقد تكون له عواقب ضارة على تطور العلم”. يعرض المقال تصنيف بدائيات النوى الذي اعتمده المؤلف. وفقًا لهذا التصنيف، ينتمي اللون الأخضر المزرق إلى المملكة الفائقة بروكاريوتا، مملكة فوتوبروكاريوتا، المملكة الفرعية بروكاريوفيكوبيونتا، قسم النباتات الزرقاء.
S. A. Balandin وآخرون، الذين يصفون مملكة النباتات، يعينون قسم البكتيريا ( البكتيريا) للنباتات السفلية، وقسم الطحالب الخضراء المزرقة ( النباتات الزرقاء- وليس غير ذلك) - للطحالب. وفي الوقت نفسه، لا يزال من غير الواضح ما هي المجموعة التصنيفية للطحالب - ربما مملكة فرعية. وفي الوقت نفسه، عند وصف قسم البكتيريا، يشير المؤلفون إلى ما يلي: "عند تصنيف البكتيريا، يتم التمييز بين عدة فئات: البكتيريا الحقيقية (eubacteria)، والبكتيريا المخاطية،... والبكتيريا الزرقاء (الطحالب الخضراء المزرقة)". ربما يكون الانتماء التصنيفي للبكتيريا الزرقاء سؤالًا مفتوحًا للمؤلفين.
توفر الأدبيات العديد من التصنيفات بناءً على التقسيم إلى مجموعات وفقًا لـ المظهريعلامات. يقوم خبراء التصنيف المختلفون بتقييم رتبة البكتيريا الزرقاء (أو الخضراء المزرقة؟) بشكل مختلف - من فئة إلى مملكة مستقلة من الكائنات الحية. وهكذا، وفقًا لنظام هيكل للممالك الثلاث (1894)، تنتمي جميع البكتيريا إلى المملكة الطلائعيات. يضع نظام الممالك الخمس لويتاكر (1969) البكتيريا الزرقاء في المملكة مونيرا. وفقًا لنظام الكائنات الحية لتاختادجيان (1973)، فإنهم ينتمون إلى المملكة الفائقة بروكاريوتا، مملكة البكتيريا. ومع ذلك، في عام 1977 أ. ينسبهم تختادجيان إلى مملكة دروبيانكا ( ميتشوتا)، المملكة الفرعية Cyanea، أو الطحالب الخضراء المزرقة ( سيانوبيونتا)، قسم النباتات الزرقاء. في الوقت نفسه، يشير المؤلف إلى أن الكثيرين يعينون المملكة بدلا من ذلك ميتشوتا"إنهم يستخدمون اسمًا مؤسفًا مونيرا، الذي اقترحه إي هيكل من أجل "نوع" خالٍ من الأسلحة النووية بروتامويباوالتي تبين أنها مجرد قطعة خالية من الأسلحة النووية من الأميبا العادية." وفقا لقواعد اللجنة الدولية للسلامة النووية الوطنية، يتم تضمين الطحالب الخضراء المزرقة في المملكة الفائقة بدائيات النوى، مملكة ميتشوتا، المملكة الفرعية أوكسيفوتوباكتريوبيونتاكقسم البكتيريا الزرقاء. نظام تصنيف الممالك الخمس وفقًا لمارجيليس وشوارتز يضع البكتيريا الزرقاء في المملكة بدائيات النوى. يحدد تصنيف الممالك الست التي وضعها كافاليير سميث هذه الشعبة البكتيريا الزرقاءإلى الإمبراطورية بروكاريوتا، مملكة بكتيريا، المملكة الفرعية نيجيبكتريا .
في التصنيف الحديث للكائنات الحية الدقيقة، يتم قبول التسلسل الهرمي التالي للأصناف: المجال، الشعبة، الطبقة، الرتبة، العائلة، الجنس، النوع. تم اقتراح تصنيف المجال على أنه أعلى بالنسبة للمملكة من أجل التأكيد على أهمية تقسيم العالم الحي إلى ثلاثة أجزاء - العتيقة, بكتيرياو حقيقيات النوى. وفقا لهذا التسلسل الهرمي، يتم تصنيف البكتيريا الزرقاء في المجال بكتيريا، فصيلة B10 البكتيريا الزرقاءوالتي بدورها تنقسم إلى خمسة أقسام فرعية.
يعرّفها مخطط متصفح التصنيف التابع للمركز الوطني لمعلومات التكنولوجيا الحيوية (NCBI) (2004) على أنها شعبة ويخصصها لمملكة مونيرا.
في السبعينيات تم تطوير القرن الماضي بواسطة K. Woese النشوء والتطورالتصنيف، الذي يعتمد على مقارنة جميع الكائنات الحية باستخدام جين واحد صغير من الرنا الريباسي (rRNA). ووفقا لهذا التصنيف تشكل البكتيريا الزرقاء فرعا منفصلا عن شجرة 16S-rRNA وتنتمي إلى المملكة يوباكتيريا. لاحقًا (1990) عرّف وويز هذه المملكة بأنها بكتيريا، تقسيم جميع الكائنات الحية إلى ثلاث ممالك - بكتيريا, العتيقةو حقيقيات النوى.
من أجل الوضوح، يتم تلخيص المخططات التصنيفية للبكتيريا الزرقاء التي تم تناولها في المقالة في الجدول 1.
الجدول 1. المخططات التصنيفية للبكتيريا الزرقاء
|
المملكة الفرعية |
||||||
|
هيكل، 1894 |
الطلائعيات |
|||||
|
هولرباخ |
بروكاريوتا |
النباتات |
سماوي- |
|||
|
ويتاكر، |
سماوي- بكتيريا |
|||||
|
تختاجيان، 1974 |
بروكاريوتا |
سيانوبيونتا |
النباتات الزرقاء |
|||
|
كوندراتيفا، 1975 |
بروكاريوتا |
صور بروكاريوتا |
بروكاريو-phycobionta |
النباتات الزرقاء |
||
|
يوباكتيريا |
سماوي- بكتيريا |
|||||
|
المدونة الدولية لتسمية البكتيريا، 1978 |
بروكاريوتا |
أوكسي فوتو- بكتيريا- بيونتا |
سماوي- بكتيريا |
|||
|
باركر، 1982 |
بروكاريوتا |
سيانوفيكوتا |
سماوي- phyceae |
|||
|
مارجيليس وشوارتز، 1982 |
بدائيات النوى |
بدائيات النوى |
البكتيريا الزرقاء |
|||
|
محدد البكتيريا بيرجي، 1984-1989 |
بدائيات النوى |
جريسيليكوتس |
أوكسي فوتو- بكتيريا |
|||
|
بكتيريا |
سماوي- بكتيريا |
|||||
|
محدد بكتيريا بيرجي، 1997 |
بدائيات النوى |
جريسيليكوتس |
أوكسي فوتو- بكتيريا |
|||
|
كافالير سميث, |
العلاقات العامةياcaryota |
بكتيريا |
نيجيبكتريا |
البكتيريا الزرقاء |
||
|
متصفح التصنيف NCBI، 2004 |
سماوي- بكتيريا |
|||||
|
بالاندين |
النباتات |
الأعشاب البحرية؟ |
النباتات الزرقاء |
|||
|
النباتات |
النباتات؟ |
البكتيريا |
سماوي- بكتيريا |
تصنيف البكتيريا الزرقاء هو في مرحلة التطوير، وفي جوهره، ينبغي اعتبار جميع الأجناس والأنواع المدرجة مؤقتة وتخضع لتعديلات كبيرة.
المبدأ الرئيسي للتصنيف لا يزال المظهري. ومع ذلك، فإن هذا التصنيف مناسب لأنه يسمح بتحديد العينات بطريقة بسيطة إلى حد ما.
الأكثر شعبية هو المخطط التصنيفي لمحدد بيرجي للبكتيريا، والذي يقسم البكتيريا أيضًا إلى مجموعات وفقًا للخصائص المظهرية.
وفقا لنشر دليل بيرجي لتصنيف البكتيريا، تم تجميع جميع الكائنات الحية قبل النووية في المملكة بدائيات النوى، والتي تم تقسيمها إلى أربعة أقسام. تصنف البكتيريا الزرقاء في القسم 1 - جريسيليكوتس، والتي تشمل جميع البكتيريا التي لها جدار خلوي من النوع سالب الجرام، الفئة 3 - البكتيريا المؤكسدة، مرتب البكتيريا الزرقاء .
تُعرّف الطبعة التاسعة من دليل بيرجي للبكتيريا الأقسام بأنها فئات، وتنقسم كل منها إلى مجموعات ليس لها وضع تصنيفي. ومن الغريب أن بعض المؤلفين يفسرون تصنيف الطبعة نفسها من كتاب بيرجي محدد البكتيريا بشكل مختلف. على سبيل المثال، ج.أ. زافارزين - يتوافق بوضوح مع التقسيم إلى مجموعات الواردة في المنشور نفسه: يتم تضمين البكتيريا الزرقاء في المجموعة 11 - البكتيريا الضوئية الأكسجينية. م.ف. جوسيف و إل. يميز Mineev جميع مجموعات البكتيريا حتى المجموعة التاسعة بما في ذلك وفقًا للمحدد، ثم تتبع الاختلافات الجذرية. وهكذا، في المجموعة 11، قام المؤلفون بتضمين التعايش الداخلي للأوالي والفطريات واللافقاريات، وتم تعيين البكتيريا المؤكسدة في المجموعة 19.
وفقا لأحدث طبعة من دليل بيرجي، يتم تضمين البكتيريا الزرقاء في المجال بكتيريا .
يعتمد المخطط التصنيفي لـ "دليل بيرجي للبكتيريا" على عدة تصنيفات: ريبكا، درويت، هيتلر، التصنيف الذي تم إنشاؤه نتيجة لإعادة التقييم النقدي لنظام هيتلر، تصنيف أناغنوستيديس وكوماريك.
يعتمد نظام درويت بشكل أساسي على شكل الكائنات الحية من عينات الأعشاب، مما يجعله غير مقبول للممارسة. يعتمد نظام هيتلر المعقد بشكل حصري تقريبًا على الخصائص المورفولوجية للكائنات الحية من العينات الطبيعية. من خلال إعادة التقييم النقدي لأجناس هيتلر، تم إنشاء نظام آخر يعتمد على الخصائص المورفولوجية وطرق التكاثر. نتيجة لإعادة التقييم النقدي لأجناس هيتلر، تم إنشاء نظام يعتمد بشكل أساسي على الخصائص المورفولوجية وطريقة تكاثر البكتيريا الزرقاء. من خلال إجراء تعديل معقد لنظام هيتلر، مع الأخذ في الاعتبار البيانات المتعلقة بالتشكل والبنية التحتية وطرق التكاثر والتباين، تم إنشاء نظام موسع حديث لـ Anagnostidis وKomarek. أبسط نظام Rippk، الوارد في دليل بيرجي للبكتيريا، يعتمد بشكل حصري تقريبًا على دراسة تلك البكتيريا الزرقاء الموجودة في الثقافات فقط. يستخدم هذا النظام الخصائص المورفولوجية، وطريقة التكاثر، والبنية التحتية للخلية، والخصائص الفسيولوجية، والتركيب الكيميائي، وأحيانا البيانات الوراثية. هذا النظام، مثل نظام Anagnostis وKomárek، هو نظام انتقالي، لأنه يقترب جزئيًا من التصنيف الوراثي، أي. يعكس السلالة والارتباط الوراثي.
وفقا للمخطط التصنيفي لدليل بيرجي للبكتيريا، تنقسم البكتيريا الزرقاء إلى خمس مجموعات فرعية. تشتمل المجموعتان الفرعيتان الأولى والثانية على أشكال أحادية الخلية أو مستعمرات غير خيطية من الخلايا متحدة بطبقات خارجية من جدار الخلية أو مصفوفة تشبه الهلام. تختلف بكتيريا كل مجموعة فرعية في طريقة تكاثرها. تشمل المجموعات الفرعية الثالث والرابع والخامس الكائنات الخيطية. تختلف بكتيريا كل مجموعة فرعية عن بعضها البعض في طريقة انقسام الخلايا، ونتيجة لذلك، في شكل الأشكال ثلاثية الألوان (متفرعة أو غير متفرعة، غير متفرعة أو متعددة التسلسل). تشتمل كل مجموعة فرعية على عدة أجناس من البكتيريا الزرقاء، بالإضافة إلى ما يسمى بـ "المجموعات الثقافية" أو "الأجيال الفائقة"، إلى جانب الأجناس، والتي من المتوقع أيضًا أن يتم تقسيمها إلى عدد من الأجناس الإضافية.
على سبيل المثال، "المجموعة الثقافية" سيانوثيسي(المجموعة الفرعية الأولى) تضم سبع سلالات مدروسة معزولة عن بيئات مختلفة. بشكل عام، تتضمن المجموعة الفرعية الأولى تسعة أجناس ( شاميسفون, سيانوثيسي, جلووباكتر, ميكروسيستيس, جليوكابسا, جلوثيسي, ميكسوباكترون, المكورات العنقودية, Synechocystis). تتضمن المجموعة الفرعية الثانية ستة أجناس ( داء الكروكوسيديا, ديرموكاربا, ديرموكاربيلا, ميكوسارسينا, بليوروكابسا, زينوكوكوس). تتضمن المجموعة الفرعية الثالثة تسعة أجناس ( أرثروسبيرا, كريناليوم, لينجبيا, ميكروكوليوس, المذبذبات, الزائفة, سبيرولينا, ستاريا, الترايكوديميوم). المجموعة الفرعية الرابعة تضم سبعة أجناس ( أنابينا, أفانيزومينون, سليندروسبيرموم, عقيدية, نوستوك, سيتونيما, كالوثريكس). تتضمن المجموعة الفرعية الخامسة أحد عشر جنسًا من البكتيريا الزرقاء الخيطية المحتملة، والتي تتميز بدرجة عالية من التعقيد المورفولوجي والتمايز (خيوط متعددة المقاطعات). هذا نوع من كلوروجلويوبسيس، فيشريلا، جيتلريا، ستيجونيما، سيانوبوتريس، لورييلا، نوستوكوبسيس، ماستيجوكلادوبسيس، ماستيجوكوليوس، ويستيلا، هابالوسيفون.
يقوم بعض المؤلفين، بناءً على تحليل جين الرنا الريباسي 16S، بتصنيف البكتيريا الزرقاء والبروكلوروفيت (الرتبة بروكلوراليس)، وهي مجموعة من بدائيات النوى تم اكتشافها مؤخرًا نسبيًا، وتقوم، مثل البكتيريا الزرقاء، بعملية التمثيل الضوئي الأكسجيني. تشبه البروكلوروفيت في كثير من النواحي البكتيريا الزرقاء، ولكن على عكسها، فهي تحتوي على الكلوروفيل أتحتوي على الكلوروفيل بلا تحتوي على أصباغ فيكوبيلين.
لا يزال هناك قدر كبير من عدم اليقين في تصنيف البكتيريا الزرقاء، وتنشأ خلافات كبيرة على كل مستوى من مستويات أبحاثهم. ولكن، كما يعتقد كوك، فإن الطحالب الخضراء المزرقة هي "المسؤولة" عن هذا المصير.
تم دعم العمل بمنحة بحثية أساسية من فرع الشرق الأقصى للأكاديمية الروسية للعلوم للفترة 2006-2008. "الكائنات الحية الدقيقة في الشرق الأقصى الروسي: التصنيف، والبيئة، وإمكانات التكنولوجيا الحيوية."
فهرس:
- بالاندين إس. إيه.، أبراموفا إل. آي.، بيريزينا إن. إيه. علم النبات العام مع أساسيات علم النبات الجيولوجي. م.: المحكمة الجنائية الدولية "Akademkniga"، 2006. ص 68.
- القاموس الموسوعي البيولوجي / إد. جيلياروف إم إس. م: الموسوعة السوفييتية، 1986. ص 63، 578.
- جيراسيمينكو إل إم، أوشاتينسكايا جي تي. // علم الحفريات البكتيرية. م: بين ران، 2002. ص 36.
- Gollerbakh M.M.، Kosinskaya E.K.، Polyansky V.I. // مفتاح طحالب المياه العذبة في اتحاد الجمهوريات الاشتراكية السوفياتية. م: سوف. العلوم، 1953. المجلد. 2. 665 ص.
- غوسيف إم في، مينيفا إل.إيه. علم الاحياء المجهري. م: الأكاديمية، 2003. 464 ص.
- إلينكين أ.أ. الطحالب الخضراء المزرقة في اتحاد الجمهوريات الاشتراكية السوفياتية. م. ل.: أكاديمية العلوم في اتحاد الجمهوريات الاشتراكية السوفياتية، 1936. 679 ص.
- إمتسيف في.تي.، ميشوستين إي.إن. علم الاحياء المجهري. م: الحبارى، 2005. ص 446.
- زافارزين جي إيه، كولوتيلوفا إن.إن. مقدمة في علم الأحياء الدقيقة للتاريخ الطبيعي. م: دار الكتب "الجامعة"، 2001. 256 ص.
- زافارزين ج. // علم الحفريات البكتيرية. م.: بين ران، 2002. ص 6.
- زاخاروف ب. النظاميات التصنيفية التحويلية. م: منشورات T-vo العلمية KMK، 2005. 164 ص.
- كوندراتييفا إن.في. // بوت. جورن، 1981. T.66. رقم 2. ص 215.
- كوك على سبيل المثال. // الحياة النباتية. م: التربية، 1977. ت.3. ص 78.
- لوبيكينا إي.جي. // مواد بين الجامعات. علمي أسيوط. "عالم النباتات في كامتشاتكا" (6 فبراير 2004). بتروبافلوفسك كامتشاتسكي: KSPU، 2004. ص 122.
- نيتروسوف إيه آي، كوتوفا آي بي. علم الاحياء المجهري. م: الأكاديمية، 2006. 352 ص.
- نيكيتينا ف.ن. // حصيرة. الثاني علمي. أسيوط. "الحفاظ على التنوع البيولوجي في كامتشاتكا والبحار المجاورة." بتروبافلوفسك كامتشاتسكي، 2001. ص 73.
- نيكيتينا ف.ن. // حصيرة. المؤتمر الحادي عشر للروسية. مجتهد. المجتمع (18-22 أغسطس 2003، نوفوسيبيرسك-بارناول). بارنول: دار نشر أزبوكا، 2003. ت3. ص129.
- معرف البكتيريا بيرجي / إد. هولت جيه، كريج إن، سنيث بي، ستالي جيه، ويليامز إس إم: العالم، 1997. المجلد 1. 431 ص.
- جنوب ر.، ويتيك أ. أساسيات علم الطحالب. م: مير، 1990. 597 ص.
- تختادجيان أ.ل. // الحياة النباتية. م: التربية، 1977. ت.1. ص 49.
- تايلور د.، جرين ن.، ستاوت دبليو. علم الأحياء. م: مير، 2004. ت 1. 454 ص.
- Stackebrandt E.، Tindall B.، Ludwig V.، Goodfellow M. // علم الأحياء الدقيقة الحديث. بدائيات النوى. م: مير، 2005. ط2، ص148.
- ياكوفليف جي.بي.، تشيلومبيتكو ف.أ. علم النبات. م: المدرسة العليا، 1990. ص 338.
- دليل بيرجي لعلم الجراثيم المنهجي: Ed.D.R. Boone, R.W. Costenholz: Springer-Verlag NY Berling, Meidelberg, 1984. V. 1.
- دليل بيرجي لعلم الجراثيم المنهجي: الطبعة الثانية Ed.D.R. Boone, R.W. Costenholz: Springer-Verlag NY Berling, Meidelberg, 2001. V. 1.
- كافاليير سميث، T. السلالة البروتستانتية والتصنيف عالي المستوى للبروتوزوا. يورو. جي بروتيستول، 2003. المجلد 39. ص 338.
- ويتاكر، آر.إتش. // العلوم، 1969. المجلد 163. ص 150.
الرابط الببليوغرافي
إفيموفا إم في، إيفيموف أ.أ. الطحالب الخضراء المزرقة أم البكتيريا الزرقاء؟ قضايا النظم // المشاكل الحديثة للعلوم والتعليم. – 2007. – رقم 6-1.;عنوان URL: http://science-education.ru/ru/article/view?id=710 (تاريخ الوصول: 01/02/2020). نلفت انتباهكم إلى المجلات التي تصدرها دار النشر "أكاديمية العلوم الطبيعية"
 الدوق الأكبر سفياتوبولك الثاني إيزياسلافيتش (1093–1113)
الدوق الأكبر سفياتوبولك الثاني إيزياسلافيتش (1093–1113)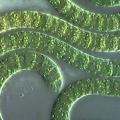 تنتمي الطحالب الخضراء المزرقة إلى المملكة
تنتمي الطحالب الخضراء المزرقة إلى المملكة حول دراسة اللغات الأجنبية في المؤسسات التعليمية
حول دراسة اللغات الأجنبية في المؤسسات التعليمية