Hva gjør den cytoplasmatiske membranen. Strukturen til cellemembranen
Under celleveggen til bakterien er den cytoplasmatiske membranen (CPM). Det skiller innholdet i cellen fra celleveggen og er en essensiell struktur for enhver celle.
Tykkelsen på CPM for bakterier er vanligvis ca. 6-8 nm. Den står for opptil 15 % av cellens tørre masse. Den består av lipider (15-45%), proteiner (45-60%) og en liten mengde karbohydrater (ca. 10%). Lipider er representert av fosfolipider - opptil 30% av den tørre massen av membranen. Blant dem dominerer fosfatidylglyserol og difosfatidylglyserid (kardiolipin), en essensiell komponent i eukaryote mitokondrielle membraner. Mindre mengder inneholder fosfatidylinositol og fosfatidyl-
etanolamin. I tillegg til fosfolipider ble det funnet ulike glykolipider, små mengder karotenoider og kinoner i membranen. I sammensetningen av lipider avledet fra glyserol, er fettsyrer som er atypiske for membraner - mettede eller enumettede med 16-18 karbonatomer, samt syrer som ikke finnes i eukaryote membraner - cyklopropan og forgrenede fettsyrer med 15-17 karbonatomer. identifisert. Settet av fettsyrer, samt membranlipidene som består av dem, er artsspesifikke for prokaryoter.
Membranlipider er små polare molekyler som bærer hydrofile (hoder) og hydrofobe (haler) grupper. I vannmiljø de danner spontant et lukket bimolekylært lag - et dobbeltlag. Dette laget fungerer som en betydelig barriere for ioner og polare forbindelser. Organisert i et bimolekylært lag danner lipider den strukturelle basis av membranen, opprettholder mekanisk stabilitet og gir den hydrofobicitet.
Proteiner utgjør mer enn halvparten av tørrvekten til membranen. Det finnes over 20 forskjellige typer. Basert på forskjeller i styrken til bindingen med lipider og plassering i membranen, deles proteiner inn i integrale og perifere. Integrerte proteiner er nedsenket i det hydrofobe området av membranen, hvor de danner mange bindinger med hydrokarbonkjeder av lipider,
skaper lipoproteinkomplekser. Perifere proteiner er lokalisert på overflaten av det hydrofile laget og fester seg ofte til integrerte proteiner (fig. 3.14).
Fig.3.14. Struktur cytoplasmatisk membran: 1 - lipider; 2 - glykoproteiner; 3 - perifere proteiner; 4 - integrerte proteiner
Membranproteiner kan deles inn i to grupper i henhold til deres funksjoner i membraner: strukturelle og dynamiske.
Funksjoner strukturelle proteiner begrenset til å opprettholde den strukturelle integriteten til membranen. De er plassert på overflaten av det hydrofile lipidlaget, og fungerer som en molekylær bandasje.
Dynamiske proteiner inkluderer proteiner som er direkte involvert i alle prosesser som skjer på membranen. De er delt inn i tre klasser: transport, som deltar i transporten av forbindelser inn og ut av cellen; katalytisk, fungerer som enzymer i reaksjoner som skjer på membranen; reseptorproteiner som spesifikt binder visse forbindelser (toksiner, hormoner) på utsiden av membranen.
Karbohydrater i membranen er ikke i fri tilstand, men er forbundet med proteiner og lipider for å danne glykoproteiner. De er som
Som regel er de bare lokalisert på den ytre overflaten av membranen og fungerer som reseptorer for gjenkjennelse av miljøfaktorer.
Den cytoplasmatiske membranen til bakterier, som alle andre biologiske membraner, er en asymmetrisk flytende krystallstruktur. Asymmetri skyldes forskjeller i den kjemiske strukturen til proteinmolekyler og deres plassering i lipid-dobbeltlaget i membranen. Noen proteiner er plassert på overflaten av dobbeltlaget, andre er nedsenket i dets tykkelse, og andre passerer gjennom fra den indre til den ytre overflaten av dobbeltlaget. Den strengt definerte orienteringen til membranproteiner skyldes i sin tur det faktum at de syntetiseres og inkorporeres i membranen asymmetrisk. De ytre og indre overflatene av membranen er også forskjellige i enzymatisk aktivitet. Avhengig av forholdene (for eksempel temperatur), kan CPM være i forskjellige fasetilstander: flytende eller krystallinsk. Under overgangen av en flytende krystallfase til en annen, endres mobiliteten til membrankomponentene og tettheten til dens pakking, noe som igjen fører til brudd på dens funksjonelle aktivitet.
Strukturell organisering og funksjonen til den cytoplasmatiske membranen. For å forklare arten og mekanismen til en rekke funksjoner til CPM, er den mest egnede væske-mosaikkmodellen for organisering av biologiske membraner, foreslått av R. Singer og A. Nicholson i 1972. I følge denne modellen er membraner to- dimensjonale løsninger av globulære proteiner og lipider orientert på en bestemt måte. Lipider danner et dobbeltlag der de hydrofile "hodene" til molekylene er vendt utover, og de hydrofobe "halene" er nedsenket i tykkelsen på membranen, samtidig som de har tilstrekkelig fleksibilitet. Membranlipider og mange proteiner beveger seg fritt i dobbeltlaget, men kun i lateral retning (lateral diffusjon). I tverrretningen, det vil si fra den ene overflaten av membranen til den motsatte, kan ikke proteiner bevege seg, og lipider beveger seg ekstremt sakte (1 gang på flere timer). Årsaken til fraværet eller lav aktivitet av tverrspredning ser ut til å være den asymmetriske fordelingen av lipider:
noen lipider er mer i den ytre delen av dobbeltlaget, andre - i det indre. Konsekvensen av dette er ulik elektrontetthet (ledningsevne) til dobbeltlaget i tverrretningen.
CPM er i flytende krystall eller flytende tilstand bare under visse, såkalte
biologiske temperaturer. Med en reduksjon i temperatur (under smeltepunktet, Tm), går lipider over i en krystallinsk tilstand, viskositeten øker til membranen størkner. Temperaturverdien som gjør at membranen stivner bestemmes av innholdet av umettede og
forgrenede fettsyrer. Jo flere av dem i membranen, jo lavere er overgangstemperaturen til lipider fra flytende-krystallinsk tilstand til krystallinsk tilstand.
Prokaryoter har evnen til å regulere membranfluiditet ved å endre antall dobbeltbindinger og kjedelengden til fettsyremolekyler. I E. coli, når temperaturen på mediet synker fra 42°C til 27°C, synker forholdet mellom mettede og umettede fettsyrer i membranen fra 1,6 til 1,0, dvs. innholdet av umettede fettsyrer når nivå av mettede. Dette forhindrer en økning i viskositeten og sikrer at cellene forblir fysiologisk aktive ved lave temperaturer.
CMP utfører en rekke vitale funksjoner i prokaryoter. I utgangspunktet bestemmes de av proteinene lokalisert i den, som fungerer som kanaler, reseptorer, energiregeneratorer, enzymer, transportfunksjoner og andre. CPM er den viktigste osmotiske barrieren, som, på grunn av tilstedeværelsen av membrantransportmekanismer, selektivt går inn i stoffer i cellen og fjerner metabolske produkter fra den. Den selektive permeabiliteten til CPM skyldes de substratspesifikke permeasene lokalisert i den, som aktivt transporterer ulike organiske og mineralske stoffer gjennom membranen. CPM inneholder enzymer for biosyntese av membranlipider og makromolekyler som utgjør celleveggen, ytre membran, kapsler. CPM er stedet for lokalisering av redoksenzymer som utfører
elektrontransport, oksidativ og fotosyntetisk fosforylering, elektrokjemisk energigenerering
transmembranpotensial (A//+) og kjemisk (ATP). CPM
n
utfører viktige funksjoner i biosyntesen og translokasjonen av utskilte proteiner av gramnegative bakterier. Biosyntesen av disse proteinene utføres på ribosomer festet til CPM. Gram-negative bakterier har spesielle reseptorproteiner på CPM som "gjenkjenner" signaler fra den store ribosomale underenheten om bindingen av ribosomet og starten av proteinsyntesen. Membranreseptorproteiner samhandler med den store underenheten av ribosomet, og danner et ribosommembrankompleks, hvor syntesen av utskilte proteiner utføres. På denne måten syntetiserer for eksempel E. coli alkalisk fosfatase, Bac. subtilis - a-amylase. CPM sikrer også overføringen av disse proteinene til det periplasmatiske rommet. Rollen til CMP i reguleringen av celledeling, kromosom- og plasmidreplikasjon og påfølgende segregering av disse genetiske elementene mellom nydannede datterceller er stor.
Alle prokaryoter, sammen med den cytoplasmatiske membranen, inneholder dens derivater - intracellulære membraner som utfører spesialiserte funksjoner. Den cytoplasmatiske membranen er i stand til å danne alle slags invaginasjoner (invaginasjoner). Disse intussusceptionene utgjør intracellulære membraner som varierer i lengde, innpakning og lokalisering i cytoplasma. De kan samles i komplekse baller - lamellformede, bikakeformasjoner eller rørformasjoner. Mindre komplekse membraner ser ut som enkle løkker eller tubuli av forskjellige lengder. Uavhengig av kompleksiteten i organiseringen av intracellulære membraner, er de alle derivater av den cytoplasmatiske membranen. Størrelsen på deres aktive overflate overstiger den cytoplasmatiske membranen. Dette gir grunnlag for å bedømme den høye funksjonelle aktiviteten til disse strukturene i celler.
Et spesielt rikt intracellulært membranapparat ble funnet i nitrogenfikserende og fotosyntetiske bakterier, Brucella og nitrifiserende bakterier. Hos fotosyntetiske bakterier (Rhodospirillum rubrum) ser membranene ut som lukkede vesikler - vesikler. Dannelsen deres begynner med invaginasjonen av den cytoplasmatiske membranen, som deretter danner en tubuli. Innsnevringer vises på røret, og deler det inn i en serie bobler. Disse vesiklene kalles kromatoforer. De inneholder lysabsorberende pigmenter - bakterioklorofyller og karotenoider, elektrontransportenzymer - ubiquinoner og cytokromer, komponenter i fosforyleringssystemet. Hos noen fotosyntetiske prokaryoter, spesielt lilla svovelbakterier og cyanobakterier, er fotosynteseapparatet representert ved stabler av membraner som har en flatformig form og, i analogi med kloroplastgranaen til grønne planter, kalles thylakoider (fig. 3.15).  De konsentrerer fotosyntesepigmenter, enzymer i elektrontransportkjeden og fosforyleringssystemer. Et trekk ved thylakoider av cyanobakterier er mangelen på forbindelse med den cytoplasmatiske membranen. Dette den eneste gruppen prokaryoter med et differensiert membransystem.
De konsentrerer fotosyntesepigmenter, enzymer i elektrontransportkjeden og fosforyleringssystemer. Et trekk ved thylakoider av cyanobakterier er mangelen på forbindelse med den cytoplasmatiske membranen. Dette den eneste gruppen prokaryoter med et differensiert membransystem.
Hos nitrifiserende bakterier har det intracellulære membranapparatet form av plater, eller lameller, bestående av flate vesikler (fig. 3.16).  Av de intracellulære membranene, de fleste kompleks struktur har mesosomer. De er spiralvridde, flate eller sfærisk rørformede legemer. Mesosomer dannes i løpet av celledelingsperioden i dannelsessonen av tverrskilleveggen. De deltar i kromosomreplikasjon og distribusjon av genomer mellom datterceller, i syntesen av celleveggstoffer. Delta
Av de intracellulære membranene, de fleste kompleks struktur har mesosomer. De er spiralvridde, flate eller sfærisk rørformede legemer. Mesosomer dannes i løpet av celledelingsperioden i dannelsessonen av tverrskilleveggen. De deltar i kromosomreplikasjon og distribusjon av genomer mellom datterceller, i syntesen av celleveggstoffer. Delta
mesosomer i celledeling indikerer dens forbindelse med nukleoidens DNA. Velutviklede mesosomer finnes bare i grampositive bakterier.
Informasjonen som er akkumulert til dags dato tyder på at membranstrukturene til bakterier er tilstrekkelig differensierte og gir forløpet til ulike metabolske prosesser i et bur.
- Cytoplasma og cytoplasmatiske inneslutninger
Det er to fraksjoner i cytoplasmaet. En av dem inneholder strukturelle elementer: ribosomer, aerosomer,
karboksysomer, reserveinneslutninger, genetisk apparatur, Den andre fraksjonen inneholder en kompleks blanding av løselig RNA, enzymatiske proteiner, pigmenter, mineraler, produkter og substrater for metabolske reaksjoner. Denne fraksjonen kalles cytosol.
På grunn av tilstedeværelsen av ulike organiske forbindelser cytoplasmaet til bakterieceller er preget av økt viskositet. Det er 800-8000 ganger viskositeten til vann (nærmer seg viskositeten til glyserin). Unge celler i lagfasen eller tidlige stadier logaritmisk fase, har en lavere viskositet av cytoplasma; i aldring - viskositeten øker, ligner en gel i konsistensen. Graden av viskositet til cytoplasmaet karakteriserer ikke bare cellens alder, men også dens fysiologiske aktivitet. En økning i viskositeten til cytoplasmaet i gamle kulturer er en av faktorene som forårsaker en reduksjon i den fysiologiske aktiviteten til cellene. Cytoplasmaet er mediet som forbinder alle intracellulære strukturer til et enkelt system.
Ribosomer. Cytoplasmaet til en bakteriecelle inneholder konstant strukturer med en sfærisk form, 15-20 nm i størrelse, molekylær vekt 3106.
Ribosomer består av 60-65% ribosomalt RNA og 35-40% protein. Sistnevnte er rike på essensielle aminosyrer. Under ultrasentrifugering setter bakterielle ribosomer seg med en hastighet på ca. 70 Svedberg-enheter (S)7, som de kalles 708-ribosomer for. De cytoplasmatiske ribosomer av eukaryoter er større og kalles 80S ribosomer (deres sedimentasjonskonstant er 80S).
Hvert ribosom består av to underenheter: 30S og 50S, som er forskjellige i størrelsen på RNA-molekyler og mengden protein de inneholder. Den store underenheten (50S) inneholder to rRNA-molekyler, 5S og 23S, og 35 molekyler av forskjellige proteiner. Den lille underenheten (30S) inneholder ett 16 rRNA-molekyl og 21 annen type proteiner. Antall ribosomer i en celle er ikke konstant - fra 5 000 til 90 000. Det bestemmes av cellens alder og betingelsene for å dyrke bakterier. Minimumsmengden er inneholdt i begynnelsen av lagfasen, og maksimum - i den eksponentielle fasen av kulturvekst. I Escherichia coli i perioden med aktiv vekst på et komplett næringsmedium, syntetiseres 5-6 ribosomer på 1 sekund. De fleste av dem i cytoplasmaet til bakterier er i fri tilstand, og resten -
S \u003d 1 swedberg enhet \u003d 10 "13 cm (s) feltenheter.
forenes av tråder av messenger RNA til polysomer. Antall ribosomer i polysomer kan nå flere titalls. Dette indikerer en høy proteinsynteseaktivitet i cellen, siden ribosomer er stedet for proteinsyntese. De kalles billedlig "fabrikker" av protein.
Gassvakuoler (aerosomer). Disse strukturene er iboende bare i noen vann- og jordbakterier. De finnes i fototrofiske svovelbakterier, fargeløse filamentøse bakterier og bakterier av slekten Renobacter. Cellen deres inneholder opptil 40-60 (fig. 3.17). Gassvakuoler er omgitt av tynne 
Ris. 3.17. Renobacter vocuolatum-celle med aerosomer (forstørrelse x 70 000)
proteinmembran. Innvendig inneholder de gassbobler, hvis antall ikke er konstant. Sammensetningen og trykket til gass i bobler og aerosomer som helhet bestemmes av mengden gasser som er oppløst i miljøet. Aerosomer er enten i en komprimert tilstand eller fylt med et gassmiljø. Tilstanden deres reguleres av mediets hydrostatiske trykk. En kraftig trykkøkning fører til kompresjon av aerosomene og cellene mister sin oppdrift.
Aerosomer regulerer oppdriften til cellen, og gjør det mulig å flytte den til gunstige forhold med lufting, belysning og næringsinnhold. En spesiell funksjon er deres engangsdrift i gassfylt tilstand. Etter kompresjon under påvirkning av hydrostatisk trykk, blir de ikke etterfylt med gass og
blir gradvis ødelagt. Cellen kan reprodusere dem bare ved å omdannes.
Når aerosomer er fylt med gass, holdes bakterier på overflaten av vannet, når de komprimeres, synker de ned i tykkelsen eller legger seg til bunnen av reservoaret. Denne særegne bevegelsesmetoden ble utviklet i løpet av evolusjonen hovedsakelig i bakterier som mangler flageller, og følgelig evnen til å bevege seg aktivt.
Phycobilisomes. Disse intracellulære strukturene er karakteristiske for cyanobakterier. De ser ut som granuler med en diameter på 28-55 nm, de er stedet for lokalisering av vannløselige pigmenter - fykobiliproteiner, som bestemmer fargen på cyanobakterier og deltar i fotosyntesen.
Klorosomer, eller klorobiumvesikler, er strukturer der fotosynteseapparatet til grønne bakterier av slekten Chlorobium er lokalisert. De har en langstrakt form, 100-150 nm lang, 50-70 nm bred, omgitt av en enkeltlags proteinmembran. Klorosomer er lokalisert i et tett lag under den cytoplasmatiske membranen, men er fysisk atskilt fra den. Klorosomene til grønne bakterier inneholder fotosyntesepigmenter - bakterioklorofyller, som absorberer lyskvanter og overfører energi til reaksjonssentrene for fotosyntesen.
Karboksysomer. I bur visse typer fototrofe (cyanobakterier, noen lilla bakterier) og kjemolitotrofe (nitrifiserende bakterier) prokaryoter inneholder strukturer som har form som et polyeder, 90-500 nm i størrelse. I samsvar med funksjonen de utfører, kalles de karboksysomer. De inneholder enzymet ribulosedifosfatkarboksylase, som katalyserer reaksjonen av karbondioksidbinding med ribulosedifosfat i Calvin-syklusen. Hos autotrofe bakterier er de stedet for karbondioksidfiksering. Karboksysomer er omgitt av en enkeltlags proteinmembran, som beskytter enzymet mot virkningene av intracellulære proteaser.
Reservenæringsstoffer* I tillegg til de beskrevne strukturelle elementene, inneholder cytoplasmaet til bakterier granuler av ulike former og størrelser i form av inneslutninger. Deres tilstedeværelse i
cellen er ustabil og er assosiert med sammensetningen av næringsmediet og den fysiologiske tilstanden til kulturen. Mange cytoplasmatiske inneslutninger består av forbindelser som tjener som en energikilde og en kilde til næringsstoffer. De dannes vanligvis i kulturer på ferske, næringsrike medier, når celleveksten hemmes av en eller annen grunn, eller etter slutten av en periode med aktiv vekst. Den kjemiske sammensetningen av inneslutningene er forskjellig og ikke den samme for forskjellige typer bakterie. De kan være polysakkarider, lipider, krystaller og granuler av uorganiske stoffer.
Av polysakkaridene bør først og fremst nevnes stivelse, glykogen og et stivelseslignende stoff, granulose. Det vanligste er glykogen. Den finnes i basiller, salmonella, Escherichia coli, sardiner osv. I spore-anaerober av slekten Clostridium inneholder cellene små granulosa-granulat. Disse inneslutningene brukes av cellen som kilder til energi og karbon.
Lipider akkumuleres i cytoplasmaet til bakterier i form av små dråper og korn. I mange bakterier er lipidinneslutninger representert av poly-p-hydroksysmørsyre, som ofte utgjør opptil 50 % av den tørre bakteriebiomassen. Spesielt rike på denne forbindelsen er bakterier av slekten Bacillus og fototrofe bakterier. Poly-p-hydroksysmørsyre syntetiseres i store mengder under vekst av mikroorganismer på medier rike på karbohydrater. I hver polylaktidkjede utgjør p-hydroksysmørsyrerester opptil 60 %, og derfor er denne forbindelsen et ideelt "lager" av energi for bakterier. Noen mikroorganismer akkumulerer voks og nøytralt fett (triglyserider). Så i mykobakterier og actinomycetes utgjør voks noen ganger opptil 40% av tørrmassen, gjærceller av slekten Candida, Rhodotorula er rike på nøytrale fettstoffer, antallet når nesten 60%.
Alle lipidinneslutninger i mikroorganismer tjener som en kilde til energi og karbon.
I cellene til mange bakterier finnes ofte spesielle inneslutninger, kalt volute korn. Av kjemisk natur er volutin et polyfosfat. Volutin navn
kommer fra artsnavnet til svovelbakterien Spirillum volutans, der disse inneslutningene først ble beskrevet. Volyutin har egenskapen til metakromasi, dvs. forårsaker misfarging av enkelte fargestoffer. Hvis bakteriene er farget med metylenblått eller toluidinblått, blir volutinkornene lilla eller rød-fiolett farge. I denne forbindelse kalte forskerne W. Babesh og E. Ernst, som først beskrev disse inneslutningene, dem metakromatiske korn. Volutin-korn er sfæriske, opptil 0,5 µm i størrelse. De dannes under forhold med god ernæring av mikroorganismer, spesielt på medier rike på karbohydrater, og også i nærvær av glyserol i mediet. Volutin finnes i cellene til både patogene og saprofytiske bakterier, for eksempel i spirilla, Azotobacter, årsaken til difteri.
Volutin brukes av cellen hovedsakelig som en kilde til fosfatgrupper og delvis til energi.
I fargeløse og lilla svovelbakterier, når sulfider oksideres inne i cellen, avsettes mineralsvovel i form av dråper. Svovel akkumuleres i medier rike på hydrogensulfid H2S. Når sulfider er oppbrukt fra miljøet, bruker bakterier intracellulært svovel. For fargeløse svovelbakterier tjener den som en energikilde; for fotosyntetiske lilla svovelbakterier fungerer den som elektrondonor.
I cyanobakterier er reservestoffet cyanofycin. Ego er et polypeptid som består av arginin og asparaginsyre. Det tjener som en kilde til nitrogen når det er mangel på det i miljøet. Akkumuleringen av cyanofycingranuler skjer i den stasjonære fasen av kulturvekst og kan være opptil 8 % av cellens tørre masse.
cytoplasmatisk membran eller plasmalemma(lat. membrana - hud, film) - den tynneste filmen ( 7– 10nm), som avgrenser det indre innholdet i cellen fra miljø kun synlig med et elektronmikroskop.
Av kjemisk organisasjon plasmalemma er et lipoproteinkompleks - molekyler lipider Og proteiner.
Den er basert på et lipid-dobbeltlag bestående av fosfolipider, i tillegg er glykolipider og kolesterol tilstede i membranene. Alle av dem har egenskapen amfipatrisitet, d.v.s. de har hydrofile ("vannelskende") og hydrofobe ("vannfryktende") ender. Hydrofile polare "hoder" av lipidmolekyler (fosfatgruppe) vender mot utsiden av membranen, og hydrofobe ikke-polare "haler" (fettsyrerester) vender mot hverandre, noe som skaper et bipolart lipidlag. Lipidmolekyler er mobile og kan bevege seg i sitt monolag eller sjelden - fra ett monolag til et annet. Lipidmonolag er asymmetriske, det vil si at de er forskjellige i lipidsammensetning, noe som gir spesifisitet til membraner selv innenfor samme celle. Lipid-dobbeltlaget kan være i en flytende eller fast krystall.
Proteiner er den andre essensielle komponenten i plasmalemmaet. Mange membranproteiner er i stand til å bevege seg i membranens plan eller rotere rundt deres akse, men kan ikke bevege seg fra den ene siden av lipid-dobbeltlaget til den andre.
Lipider gir de grunnleggende strukturelle egenskapene til membranen, mens proteiner gir dens funksjoner.
Funksjonene til membranproteiner er forskjellige: opprettholde strukturen til membraner, motta og konvertere signaler fra miljøet, transportere visse stoffer, katalysere reaksjoner som oppstår på membraner.
Det finnes flere modeller av strukturen til den cytoplasmatiske membranen.
①. SANDWICH MODELL(ekorn– lipider– proteiner)
I 1935 engelske vitenskapsmenn Danieli Og Dawson uttrykte ideen om et lag-for-lag-arrangement i membranen til proteinmolekyler (mørke lag i et elektronmikroskop), som ligger utenfor, og lipidmolekyler (lette lag) - inne . I lang tid var det en idé om en enkelt trelagsstruktur av alle biologiske membraner.
En detaljert studie av membranen ved hjelp av et elektronmikroskop viste at lyslaget faktisk er representert av to lag fosfolipider - dette lipidlag, og dens vannløselige deler er hydrofile hoder rettet mot proteinlaget, og uløselig (fettsyrerester) - hydrofobe haler vendt mot hverandre.
 ②. VÆSENDE MOSAIKKMODEL
②. VÆSENDE MOSAIKKMODEL
I 1972.Sanger Og Nicholson beskrevet en modell av membranen som har fått bred aksept. I følge denne modellen danner ikke proteinmolekyler et sammenhengende lag, men er nedsenket i det bipolare lipidlaget på forskjellige dyp i form av en mosaikk. Kuler av proteinmolekyler, som isfjell, er nedsenket i "havet"
lipider: noen er lokalisert på overflaten av bilipidlaget - perifere proteiner, andre er halvt nedsenket i det - semi-integrerte proteiner, tredje - integrerte proteiner- penetrere den gjennom og gjennom, og danner hydrofile porer. Perifere proteiner, som er på overflaten av lipidlaget, er assosiert med hodene til lipidmolekyler ved elektrostatiske interaksjoner. Men de danner aldri et sammenhengende lag og er faktisk ikke proteiner i selve membranen, men kobler den heller til supramembranen eller submembransystemet. overflateapparat celler.
Hovedrollen i organiseringen av selve membranen spilles av integrerte og semi-integrerte (transmembrane) proteiner, som har en kulestruktur og er assosiert med lipidfasen ved hydrofile-hydrofobe interaksjoner. Proteinmolekyler, som lipider, er amfipatriske og deres hydrofobe områder samhandler med de hydrofobe halene av bilipidlaget, mens de hydrofile områdene vender mot vannmiljøet og danner hydrogenbindinger med vann.
③. PROTEIN-KRYSTALL MODELL(lipoprotein mattemodell)
Membraner dannes ved sammenveving av lipid- og proteinmolekyler, som er kombinert med hverandre på grunnlag av hydrofile
hydrofobe interaksjoner.
 |
|||

Proteinmolekyler, som pinner, trenger inn i lipidlaget og utfører funksjonen til et rammeverk i membranen. Etter behandling av membranen med fettløselige stoffer bevares proteinrammeverket, noe som beviser forholdet mellom proteinmolekyler i membranen. Tilsynelatende implementeres denne modellen bare i visse spesielle områder av noen membraner, der det kreves en stiv struktur og nære stabile forhold mellom lipider og proteiner (for eksempel i regionen der enzymet Na-K-ATP-aser).
Den mest universelle modellen som oppfyller termodynamiske prinsipper (prinsipper for hydrofile-hydrofobe interaksjoner), morfo-biokjemiske og eksperimentelle
Antal-cytologiske data er en væske-mosaikkmodell. Alle tre membranmodellene er imidlertid ikke gjensidig utelukkende og kan forekomme i forskjellige områder av samme membran, avhengig av funksjonelle funksjoner av dette området.
MEMBRANEGENSKAPER
1. Evne til selvmontering. Etter destruktive påvirkninger er membranen i stand til å gjenopprette sin struktur, fordi. lipidmolekyler basert på deres Fysiske og kjemiske egenskaper settes sammen til et bipolart lag, som proteinmolekyler deretter er innebygd i.
2. Fluiditet. Membranen er ikke en stiv struktur, de fleste av dens proteiner og lipider kan bevege seg i membranens plan, de svinger konstant på grunn av rotasjons- og oscillerende bevegelser. Dette bestemmer den høye hastigheten av kjemiske reaksjoner på membranen.
3. Semipermeabilitet. Membranene til levende celler passerer, i tillegg til vann, bare visse molekyler og ioner av oppløste stoffer. Dette sikrer opprettholdelsen av den ioniske og molekylære sammensetningen av cellen.
4. Membranen har ingen løse ender. Den lukkes alltid i bobler.
5. asymmetri. Sammensetningen av de ytre og indre lag av både proteiner og lipider er forskjellig.
6. Polaritet. Den ytre siden av membranen har en positiv ladning, mens den indre siden har en negativ ladning.
MEMBRANFUNKSJONER
1) Barriere - Plasmalemmaet skiller cytoplasmaet og kjernen fra det ytre miljøet. I tillegg deler membranen det indre innholdet i cellen i seksjoner (avdelinger), hvor det ofte oppstår motsatte biokjemiske reaksjoner.
2) Reseptor(signal) - på grunn av den viktige egenskapen til proteinmolekyler - denaturering, er membranen i stand til å fange opp ulike endringer i miljøet. Så når en cellemembran blir utsatt for ulike miljøfaktorer (fysiske, kjemiske, biologiske), endrer proteinene som utgjør sammensetningen deres romlige konfigurasjon, som fungerer som et slags signal for cellen.
Dette gir kommunikasjon med det ytre miljø, cellegjenkjenning og deres orientering under vevsdannelse osv. Denne funksjonen er assosiert med aktiviteten til ulike reguleringssystemer og dannelsen av en immunrespons.
3) Utveksling- Membranen inneholder ikke bare strukturelle proteiner som danner den, men også enzymatiske proteiner som er biologiske katalysatorer. De er plassert på membranen i form av en "katalytisk transportør" og bestemmer intensiteten og retningen til metabolske reaksjoner.
4) Transportere– molekyler av stoffer hvis diameter ikke overstiger 50 nm kan trenge gjennom passiv og aktiv transport gjennom porene i membranstrukturen. Store stoffer kommer inn i cellen ved endocytose(transport i membranemballasje), som krever energiforbruk. Dens varianter er fag og pinocytose.
Passiv transport - en transportmåte der overføring av stoffer utføres langs en gradient av kjemisk eller elektrokjemisk konsentrasjon uten forbruk av ATP-energi. Det finnes to typer passiv transport: enkel og tilrettelagt diffusjon. Diffusjon- dette er overføring av ioner eller molekyler fra en sone med høyere konsentrasjon til en sone med lavere konsentrasjon, dvs. langs gradienten.
enkel diffusjon- saltioner og vann trenger gjennom transmembrane proteiner eller fettløselige stoffer langs en konsentrasjonsgradient.
Tilrettelagt diffusjon- spesifikke bærerproteiner binder stoffet og overfører det gjennom membranen etter "ping-pong"-prinsippet. På denne måten passerer sukkerarter og aminosyrer gjennom membranen. Hastigheten for slik transport er mye høyere enn for enkel diffusjon. I tillegg til bærerproteiner er noen antibiotika, som gramitidin og vanomycin, involvert i forenklet diffusjon.
Fordi de gir ionetransport, kalles de ionoforer.
Aktiv transport er en transportmåte der energien til ATP forbrukes, den går mot konsentrasjonsgradienten. Det involverer enzymene ATPase. Den ytre cellemembranen inneholder ATPaser, som transporterer ioner mot en konsentrasjonsgradient, et fenomen som kalles ionepumpen. Et eksempel er natrium-kalium-pumpen. Normalt er det flere kaliumioner i cellen, i eksternt miljø- natriumioner. Derfor, i henhold til lovene for enkel diffusjon, har kalium en tendens til å forlate cellen, og natrium kommer inn i cellen. I motsetning til dette, pumper natrium-kalium-pumpen kaliumioner inn i cellen mot en konsentrasjonsgradient, og frakter natriumioner inn i det ytre miljøet. Dette gjør det mulig å opprettholde konstansen til den ioniske sammensetningen i cellen og dens levedyktighet. I dyrebur en tredjedel av ATP brukes til natrium-kalium-pumpen.
En type aktiv transport er membranpakket transport. endocytose. Store molekyler av biopolymerer kan ikke trenge gjennom membranen, de kommer inn i cellen i en membranpakke. Skille mellom fagocytose og pinocytose. Fagocytose- fangst av faste partikler av cellen, pinocytose- væskepartikler. Disse prosessene er delt inn i stadier:
1) gjenkjennelse av et stoff av membranreseptorer; 2) invaginasjon (invagination) av membranen med dannelse av en vesikkel (vesikkel); 3) løsgjøring av vesikelen fra membranen, dens fusjon med det primære lysosomet og gjenoppretting av integriteten til membranen; 4) frigjøring av ufordøyd materiale fra cellen (eksocytose).
Endocytose er en måte å fôre for protozoer. Pattedyr og mennesker har et retikulo-histio-endotelsystem av celler som er i stand til endocytose - disse er leukocytter, makrofager, Kupffer-celler i leveren.
OSMOTISKE EGENSKAPER TIL CELLEN
Osmose- enveis prosess for vannpenetrering gjennom en semipermeabel membran fra et område med lavere løsningskonsentrasjon til et område med mer høy konsentrasjon. Osmose bestemmer osmotisk trykk.
Dialyse– enveisdiffusjon av oppløste stoffer.
En løsning der det osmotiske trykket er det samme som i celler kalles isotonisk. Når en celle er nedsenket i en isotonisk løsning, endres ikke volumet. En isotonisk løsning kalles fysiologisk– Dette er en 0,9 % natriumkloridløsning, som er mye brukt i medisin ved alvorlig dehydrering og tap av blodplasma.
En løsning hvis osmotiske trykk er høyere enn i celler kalles hypertonisk.
Celler i en hypertonisk løsning mister vann og skrumper. Hypertone løsninger er mye brukt i medisin. En gasbind som er dynket i en hypertonisk løsning absorberer puss godt.
En løsning hvor konsentrasjonen av salter er lavere enn i cellen kalles hypotonisk. Når en celle er nedsenket i en slik løsning, strømmer vann inn i den. Cellen svulmer, dens turgor øker, og den kan kollapse. Hemolyse- ødeleggelse av blodceller i en hypotonisk løsning.
Osmotisk trykk i menneskekroppen som helhet reguleres av systemet med utskillelsesorganer.
Forrige123456789Neste
SE MER:
cellemembran også kalt plasma (eller cytoplasmatisk) membran og plasmalemma. Denne strukturen skiller ikke bare det indre innholdet i cellen fra det ytre miljøet, men går også inn i sammensetningen av de fleste celleorganeller og kjerner, som igjen skiller dem fra hyaloplasma (cytosol) - den viskøse-flytende delen av cytoplasma. La oss bli enige om å ringe cytoplasmatisk membran en som skiller innholdet i cellen fra det ytre miljøet. De resterende begrepene refererer til alle membraner.
Strukturen til cellemembranen
Grunnlaget for strukturen til cellens (biologiske) membran er et dobbelt lag av lipider (fett). Dannelsen av et slikt lag er assosiert med egenskapene til molekylene deres. Lipider løses ikke opp i vann, men kondenserer i det på sin egen måte. Den ene delen av et enkelt lipidmolekyl er et polart hode (det tiltrekkes av vann, dvs. hydrofilt), og den andre er et par lange ikke-polare haler (denne delen av molekylet frastøtes av vann, dvs. hydrofob) . Denne strukturen til molekylene gjør at de «gjemmer» halene sine for vannet og vender polarhodene mot vannet.
Som et resultat dannes et lipid-dobbeltlag, der de ikke-polare halene er inne (mot hverandre), og de polare hodene vender ut (til det ytre miljø og cytoplasma). Overflaten til en slik membran er hydrofil, men innvendig er den hydrofob.
I cellemembraner dominerer fosfolipider blant lipider (de er komplekse lipider). Hodene deres inneholder en rest av fosforsyre. I tillegg til fosfolipider er det glykolipider (lipider + karbohydrater) og kolesterol (tilhører steroler). Sistnevnte gir membranen stivhet, og ligger i tykkelsen mellom halene til de gjenværende lipidene (kolesterol er fullstendig hydrofobt).
På grunn av elektrostatisk interaksjon festes visse proteinmolekyler til de ladede hodene til lipider, som blir overflatemembranproteiner. Andre proteiner samhandler med ikke-polare haler, synker delvis ned i dobbeltlaget eller trenger gjennom det.
Således består cellemembranen av et dobbeltlag av lipider, overflate (perifere), nedsenkede (semi-integrerte) og penetrerende (integrerte) proteiner. I tillegg noen proteiner og lipider utenfor membraner er knyttet til karbohydratkjeder.

Dette flytende mosaikkmodell av membranstrukturen ble fremsatt på 70-tallet av XX-tallet. Før dette ble det antatt en sandwichmodell av strukturen, ifølge hvilken lipid-dobbeltlaget er plassert inne, og på innsiden og utsiden er membranen dekket med kontinuerlige lag av overflateproteiner. Akkumuleringen av eksperimentelle data motbeviste imidlertid denne hypotesen.
Tykkelsen på membraner i forskjellige celler er omtrent 8 nm. Membraner (til og med forskjellige sider av en) skiller seg fra hverandre i prosentdel forskjellige typer lipider, proteiner, enzymatisk aktivitet osv. Noen membraner er mer flytende og mer permeable, andre er tettere.
pauser cellemembran flettes lett sammen på grunn av de fysisk-kjemiske egenskapene til lipid-dobbeltlaget. I membranens plan beveger lipider og proteiner (med mindre de er fiksert av cytoskjelettet).
Funksjoner av cellemembranen
De fleste av proteinene nedsenket i cellemembranen utfører en enzymatisk funksjon (de er enzymer). Ofte (spesielt i membranene til celleorganeller) er enzymer ordnet i en bestemt rekkefølge slik at reaksjonsproduktene katalysert av ett enzym går over til det andre, deretter det tredje osv. Det dannes en transportør som stabiliserer overflateproteiner, fordi de ikke tillate enzymer å svømme langs lipid-dobbeltlaget.
Cellemembranen utfører en avgrensende (barriere) funksjon fra omgivelsene og samtidig en transportfunksjon. Det kan sies at dette er dens viktigste formål. Den cytoplasmatiske membranen, som har styrke og selektiv permeabilitet, opprettholder konstanten til den indre sammensetningen av cellen (dens homeostase og integritet).
I dette tilfellet skjer transport av stoffer forskjellige måter. Transport langs en konsentrasjonsgradient innebærer bevegelse av stoffer fra et område med høyere konsentrasjon til et område med lavere (diffusjon). Så for eksempel diffunderer gasser (CO 2, O 2).
Det er også transport mot konsentrasjonsgradienten, men med energiforbruk.
Transport er passiv og lett (når den blir hjulpet av en form for overføring).
Til). Passiv diffusjon over cellemembranen er mulig for fettløselige stoffer.
Det er spesielle proteiner som gjør membraner permeable for sukker og andre vannløselige stoffer. Disse bærerne binder seg til transporterte molekyler og drar dem over membranen.
3. Funksjoner og struktur av cytoplasmatisk membran
Slik transporteres glukose inn i de røde blodcellene.
Spennende proteiner, når de kombineres, kan danne en pore for bevegelse av visse stoffer gjennom membranen. Slike bærere beveger seg ikke, men danner en kanal i membranen og fungerer på samme måte som enzymer, og binder et spesifikt stoff. Overføringen utføres på grunn av en endring i konformasjonen av proteinet, på grunn av hvilke kanaler som dannes i membranen. Et eksempel er natrium-kalium-pumpen.
Transportfunksjonen til den eukaryote cellemembranen realiseres også gjennom endocytose (og eksocytose). Gjennom disse mekanismene kommer store molekyler av biopolymerer, til og med hele celler, inn i cellen (og ut av den). Endo- og eksocytose er ikke karakteristisk for alle eukaryote celler (prokaryoter har det ikke i det hele tatt). Så endocytose observeres i protozoer og lavere virvelløse dyr; hos pattedyr absorberer leukocytter og makrofager skadelige stoffer og bakterier, dvs. endocytose utfører beskyttende funksjon for kroppen.
Endocytose er delt inn i fagocytose(cytoplasma omslutter store partikler) og pinocytose(fangst av væskedråper med stoffer oppløst i den). Mekanismen til disse prosessene er omtrent den samme. Absorberte stoffer på celleoverflaten er omgitt av en membran. En vesikkel (fagocytisk eller pinocytisk) dannes, som deretter beveger seg inn i cellen.
Eksocytose er fjerning av stoffer fra cellen ved den cytoplasmatiske membranen (hormoner, polysakkarider, proteiner, fett, etc.). Disse stoffene er innelukket i membranvesikler som passer til cellemembranen. Begge membranene smelter sammen og innholdet er utenfor cellen.
Den cytoplasmatiske membranen utfører en reseptorfunksjon. For å gjøre dette, på dens ytre side er det strukturer som kan gjenkjenne en kjemisk eller fysisk stimulus. Noen av proteinene som trenger inn i plasmalemmaet er koblet fra utsiden til polysakkaridkjeder (danner glykoproteiner). Dette er særegne molekylære reseptorer som fanger opp hormoner. Når et bestemt hormon binder seg til reseptoren, endrer det strukturen. Dette utløser igjen den cellulære responsmekanismen. Samtidig kan kanaler åpne seg, og visse stoffer kan begynne å komme inn i cellen eller fjernes fra den.
Reseptorfunksjonen til cellemembraner er godt studert basert på virkningen av hormonet insulin. Når insulin binder seg til sin glykoproteinreseptor, aktiveres den katalytiske intracellulære delen av dette proteinet (enzymet adenylatcyklase). Enzymet syntetiserer syklisk AMP fra ATP. Allerede aktiverer eller hemmer det ulike enzymer i cellulær metabolisme.
Reseptorfunksjonen til den cytoplasmatiske membranen inkluderer også gjenkjennelse av naboceller av samme type. Slike celler er festet til hverandre ved hjelp av ulike intercellulære kontakter.
I vev, ved hjelp av intercellulære kontakter, kan celler utveksle informasjon med hverandre ved hjelp av spesialsyntetiserte lavmolekylære stoffer. Et eksempel på en slik interaksjon er kontakthemming, når celler slutter å vokse etter å ha mottatt informasjon om at den ledige plassen er opptatt.
Intercellulære kontakter er enkle (membraner av forskjellige celler er ved siden av hverandre), låsing (invaginasjon av membranen til en celle inn i en annen), desmosomer (når membranene er forbundet med bunter av tverrgående fibre som trenger inn i cytoplasmaet). I tillegg er det en variant av intercellulære kontakter på grunn av mediatorer (mellomledd) - synapser. I dem overføres signalet ikke bare kjemisk, men også elektrisk. Synapser overfører signaler mellom nerveceller, så vel som fra nerve til muskel.
celleteori
I 1665 fant R. Hooke, som undersøkte et kutt av en trekork under et mikroskop, tomme celler, som han kalte "celler". Han så bare skjellene til planteceller, og lang tid skallet ble ansett som den viktigste strukturelle komponenten i cellen. I 1825 beskrev J. Purkinė protoplasmaet til celler, og i 1831 beskrev R. Brown kjernen. I 1837 kom M. Schleiden til at planteorganismer består av celler, og hver celle inneholder en kjerne.
1.1. Ved å bruke dataene akkumulert på den tiden, T.
Den cytoplasmatiske membranen, dens funksjoner og struktur
Schwann formulerte i 1839 hovedbestemmelsene i celleteorien:
1) buret er grunnleggende strukturell enhet planter og dyr;
2) prosessen med celledannelse bestemmer vekst, utvikling og differensiering av organismer.
I 1858 R. Virchow - grunnleggeren patologisk anatomi- supplert den cellulære teorien med den viktige bestemmelsen at en celle bare kan komme fra en celle (Omnis cellula e cellula) som følge av dens deling. Han fant at grunnlaget for alle sykdommer er endringer i cellenes struktur og funksjon.
1.2. Moderne celleteori inkluderer følgende bestemmelser:
1) celle - den viktigste strukturelle, funksjonelle og genetiske enheten til levende organismer, den minste enheten av levende ting;
2) cellene til alle encellede og flercellede organismer er like i struktur, kjemisk sammensetning og de viktigste manifestasjonene av livsprosesser;
3) hver ny celle dannes som et resultat av delingen av den opprinnelige (moder)cellen;
4) cellene til flercellede organismer er spesialiserte: de utfører ulike funksjoner og danner vev
5) cellen er et åpent system som strømmer av materie, energi og informasjon passerer og transformeres gjennom
Strukturen og funksjonene til den cytoplasmatiske membranen
Cellen er åpen selvregulerende system, der det er en konstant flyt av materie, energi og informasjon. Disse strømmene mottas spesielt apparat celler som inkluderer:
1) supramembranøs komponent - glykokalyx;
2) elementær biologisk membran eller deres kompleks;
3) submembranstøtte-kontraktilt kompleks av hyaloplasma;
4) anabole og katabolske systemer.
Hovedkomponenten i dette apparatet er den elementære membranen.
Cellen inneholder ulike typer membraner, men prinsippet for deres struktur er det samme.
I 1972 foreslo S. Singer og G. Nicholson en væske-mosaikkmodell av den elementære membranstrukturen. I følge denne modellen er den også basert på bilipidlaget, men proteinene er plassert annerledes i forhold til dette laget. Noen proteinmolekyler ligger på overflaten av lipidlagene (perifere proteiner), noen penetrerer ett lipidlag (semi-integrerte proteiner), og noen penetrerer begge lipidlagene (integrerte proteiner). Lipidlaget er i væskefasen ("lipidhavet"). På den ytre overflaten av membranene er det et reseptorapparat - glykokalyxen, dannet av forgrenede molekyler av glykoproteiner, "gjenkjennende" visse stoffer og strukturer.
2.3. Membranegenskaper: 1) plastisitet, 2) semipermeabilitet, 3) selvlukkende evne.
2.4. Membranfunksjoner: 1) strukturelle - en membran som strukturell komponent er en del av de fleste organeller (membranprinsippet for strukturen til organeller); 2) barriere og regulatorisk - opprettholder konstansen til den kjemiske sammensetningen og regulerer alle metabolske prosesser (metabolske reaksjoner forekommer på membraner); 3) beskyttende; 4) reseptor.
Celle - den viktigste strukturelle, funksjonelle, genetiske enheten i organiseringen av levende ting, et elementært levende system. En celle kan eksistere som en separat organisme (bakterier, protozoer), eller som en del av vevet til flercellede organismer. Begrepet "celle" ble foreslått av den engelske oppdageren Robert Hooke i 1665.
Hovedbestemmelsene i celleteorien ble formulert av botanikeren Schleiden (1838) og zoolog-fysiologen Schwann (1839). I 1858 supplerte Virchow bestemmelsene med en uttalelse om celledeling.
De viktigste bestemmelsene i moderne celleteori:
Alle levende organismer består av celler. En celle er en enhet av struktur, funksjon, reproduksjon og individuell utvikling av levende organismer. Det er ikke liv utenfor cellen.
Cellene til alle organismer er like i struktur og kjemisk sammensetning.
Celler kan kun dannes fra celler ved deling.
Den cellulære strukturen til alle levende organismer er bevis på opprinnelsesenheten.
Moderne definisjon av en celle:
en celle er et åpent biologisk system, begrenset av en semipermeabel membran, bestående av en kjerne og cytoplasma, i stand til selvregulering og selvreproduksjon.
To typer organismer har en cellulær struktur - prokaryoter (bakterier og blågrønnalger) og eukaryoter (fig. 2.1). Eukaryote celler består av et overflateapparat (cytoplasmatisk membran), cytoplasma og kjerne.
cytoplasmatiske membraner
Cytoplasmatiske membraner utfører en rekke viktige funksjoner: barriere (avgrensende), regulatorisk (regulerer metabolske strømmer), transport (sikrer selektiv permeabilitet av stoffer gjennom passiv og aktiv transport), strukturell, utveksling. Biologiske membraner bygges først og fremst av lipider, proteiner og
karbohydrater (fig. 2.2). Flere modeller av strukturen til cytoplasmatiske membraner er foreslått («sandwich»-modellen - Danieli og Dawson-modellen, Lenard-modellen, etc.). Tilsynelatende, avhengig av funksjonen, er det flere typer membraner. For tiden er væske-mosaikk-modellen foreslått av Singer-Nicholson (1972) lagt til grunn. I følge denne modellen inkluderer membranene et bimolekylært lag av lipider, som inkluderer proteinmolekyler.
Lipider er vannuløselige stoffer. De har et polart (ladet) hode og lange uladede (ikke-polare) karbohydratkjeder. Lipidmolekyler vender mot hverandre med upolare ender, og deres polare poler (hoder) forblir utenfor og danner hydrofile overflater (fig. 2.3).
Membranproteiner kan deles inn i tre grupper: perifere (den svakeste assosiert med membranen), nedsenket (semi-integrert) og penetrerende (integrert), og danner porene i membranen. Funksjonelt er membranproteiner delt inn i enzymatiske, transport-, strukturelle og regulatoriske.
På den ytre overflaten av plasmamembranen er protein- og lipidmolekyler assosiert med karbohydratkjeder, og danner en glykokalyx. Karbohydratkjeder fungerer som reseptorer, cellen får evnen til å spesifikt reagere på ytre påvirkninger. Dermed forårsaker interaksjonen av et hormon med sin "egen" reseptor fra utsiden en endring i strukturen til det integrerte proteinet, noe som fører til utløsning av en cellulær respons. Spesielt kan en slik respons manifestere seg i dannelsen av "kanaler" gjennom hvilke løsninger av visse stoffer begynner å komme inn i cellen eller fjernes fra den.
En av de viktige funksjonene til membranen er å gi kontakter mellom celler i organer og vev.
Under plasmamembranen, på siden av cytoplasmaet, er det et kortikalt lag og intracellulære fibrillære strukturer som sikrer den mekaniske stabiliteten til membranen.
I planteceller er en tett struktur plassert utenfor membranen - celleveggen, bestående av polysakkarider (cellulose).
En av de viktigste egenskapene til cytoplasmaet er assosiert med evnen til å føre ulike stoffer inn eller ut av cellen. Dette er nødvendig for å opprettholde konstansen i sammensetningen. Små molekyler og ioner passerer gjennom membraner ved passiv og aktiv transport.
Passiv transport skjer uten energiforbruk ved diffusjon, osmose og tilrettelagt diffusjon (Fig. 2.4). Diffusjon -

transport av molekyler og ioner gjennom membranen fra en region med høy til en region med lav konsentrasjon, dvs. langs konsentrasjonsgradienten. Hvis stoffer er godt løselige i fett, trenger de inn i cellen ved enkel diffusjon (oksygen, karbondioksid). Diffusjonen av vann over semipermeable membraner kalles osmose. Vann er også i stand til å passere gjennom membranporene dannet av proteiner og bære molekyler av ioner og stoffer oppløst i det. Tilrettelagt diffusjon er transport av stoffer som er uløselige i fett og ikke passerer gjennom porene gjennom ionekanaler ved hjelp av bærerproteiner.
Aktiv transport av stoffer over membranen skjer med forbruk av ATP-energi og med deltakelse av bærerproteiner. Det utføres mot en konsentrasjonsgradient (dette er hvordan aminosyrer, sukker, kalium, natrium, kalsiumioner, etc.) transporteres. Et eksempel på en aktiv tramport er driften av en kalium-natriumpumpe. Konsentrasjonen av K inne i cellen er 10-20 ganger høyere enn utenfor, og Na er omvendt. For å opprettholde denne konsentrasjonen overføres tre Na-ioner fra cellen for hver to K-ioner inn i cellen. Denne prosessen involverer et protein i membranen, som fungerer som et enzym som bryter ned ATP med frigjøring av energien som er nødvendig for driften av pumpen (fig. 2.5. A).
Overføringen av makromolekyler og store partikler inn i cellen utføres ved endocytose, og fjerning fra cellen - ved eksocytose.
Under endocytose (fig. 2.5. B) danner membranen invaginasjoner eller utvekster, som deretter, snøring av, blir til intracellulære vesikler som inneholder produktet fanget av cellen. Denne prosessen foregår med forbruk av ATP-energi. Det finnes to typer endocytose - fagocytose (absorpsjon av store partikler av cellen) og pinocytose (absorpsjon av flytende stoffer).
Membranen tar del i fjerning av stoffer fra cellen i prosessen med eksocytose. På denne måten fjernes hormoner, proteiner, fettdråper osv. fra cellen.
Den cytoplasmatiske cellemembranen består av tre lag:
Ekstern - protein;
Mellom - bimolekylært lag av lipider;
Intern - protein.
Membrantykkelsen er 7,5-10 nm. Det bimolekylære laget av lipider er matrisen til membranen. Lipidmolekylene i begge lagene samhandler med proteinmolekylene som er nedsenket i dem. Fra 60 til 75% av membranlipidene er fosfolipider, 15-30% kolesterol. Proteiner er hovedsakelig representert av glykoproteiner. Skille integrerte proteiner spenner over hele membranen, og perifert plassert på utsiden eller indre overflate.
integrerte proteiner danner ionekanaler som gir utveksling av visse ioner mellom den ekstra- og intracellulære væsken. De er også enzymer som utfører antigradienttransport av ioner over membranen.
Perifere proteiner er kjemoreseptorer på den ytre overflaten av membranen, som kan samhandle med ulike fysiologisk aktive stoffer.
Membranfunksjoner:
1. Sikrer integriteten til cellen som en strukturell enhet av vevet.
Utfører utveksling av ioner mellom cytoplasma og ekstracellulær væske.
Gir aktiv transport av ioner og andre stoffer inn og ut av cellen.
Produserer oppfatningen og behandlingen av informasjon som kommer til cellen i form av kjemiske og elektriske signaler.
Mekanismer for celleeksitabilitet. Historie om studiet av bioelektriske fenomener.
I utgangspunktet er informasjonen som overføres i kroppen i form av elektriske signaler (for eksempel nerveimpulser). Tilstedeværelsen av animalsk elektrisitet ble først etablert av naturforskeren (fysiologen) L. Galvani i 1786. For å studere atmosfærisk elektrisitet hang han nevromuskulære preparater av froskebein på en kobberkrok. Når disse potene rørte ved jernrekkverket på balkongen, trakk musklene seg sammen. Dette indikerte virkningen av en slags elektrisitet på nerven til det nevromuskulære preparatet. Galvani mente at dette skyldtes tilstedeværelsen av elektrisitet i selve levende vev. Imidlertid fant A. Volta at kilden til elektrisitet er kontaktstedet for to forskjellige metaller - kobber og jern. I fysiologi Galvanis første klassiske opplevelse det anses å berøre nerven til det nevromuskulære preparatet med bimetallisk pinsett laget av kobber og jern. For å bevise sin sak produserte Galvani andre opplevelse. Han kastet enden av nerven som innerverer det nevromuskulære preparatet over kuttet av muskelen hans. Resultatet ble en sammentrekning. Denne opplevelsen overbeviste imidlertid ikke Galvanis samtidige. Derfor gjorde en annen italiener Matteuchi følgende eksperiment. Han la nerven til det ene nevromuskulære froskepreparatet på muskelen til det andre, som trakk seg sammen under påvirkning av en irriterende strøm. Som et resultat begynte det første stoffet også å avta. Dette indikerte overføring av elektrisitet (aksjonspotensial) fra en muskel til en annen. Tilstedeværelsen av en potensiell forskjell mellom de skadede og uskadede delene av muskelen ble først nøyaktig fastslått på 1800-tallet ved hjelp av et strenggalvanometer (amperemeter) Matteuchi. Dessuten hadde snittet negativ ladning, og muskeloverflaten er positiv.
Cytoplasma- en obligatorisk del av cellen, innelukket mellom plasmamembranen og kjernen; Det er delt inn i hyaloplasma (hovedstoffet i cytoplasma), organeller (permanente komponenter i cytoplasma) og inneslutninger (midlertidige komponenter i cytoplasma). Den kjemiske sammensetningen av cytoplasma: grunnlaget er vann (60-90% av den totale massen av cytoplasma), ulike organiske og uorganiske forbindelser. Cytoplasmaet er alkalisk. Trekk eukaryot celle cytoplasma - konstant bevegelse ( syklose). Det oppdages først og fremst ved bevegelse av celleorganeller, slik som kloroplaster. Hvis bevegelsen av cytoplasma stopper, dør cellen, fordi den bare er inne i konstant bevegelse, kan den utføre sine funksjoner.
Hyaloplasma ( cytosol) er en fargeløs, slimete, tykk og gjennomsiktig kolloidal løsning. Det er i det alle metabolske prosesser finner sted, det gir sammenkoblingen av kjernen og alle organeller. Avhengig av overvekt av den flytende delen eller store molekyler i hyaloplasma, skilles to former for hyaloplasma: sol- mer flytende hyaloplasma og gel- tettere hyaloplasma. Gjensidige overganger er mulige mellom dem: gelen blir til en sol og omvendt.
Funksjoner av cytoplasma:
- integrering av alle komponenter i cellen i et enkelt system,
- miljø for passasje av mange biokjemiske og fysiologiske prosesser,
- miljø for eksistensen og funksjonen til organeller.
Cellevegger
Cellevegger begrense eukaryote celler. Minst to lag kan skilles i hver cellemembran. Det indre laget er ved siden av cytoplasmaet og er representert ved plasmamembran(synonymer - plasmalemma, cellemembran, cytoplasmatisk membran), som det ytre laget dannes over. I en dyrecelle er den tynn og kalles glykokalyx(dannet av glykoproteiner, glykolipider, lipoproteiner), i Plante-celle- tykk, kalt celleveggen(dannet av cellulose).
Alle biologiske membraner har felles strukturelle trekk og egenskaper. Foreløpig generelt akseptert flytende mosaikkmodell av membranstrukturen. Grunnlaget for membranen er et lipid-dobbeltlag, hovedsakelig dannet av fosfolipider. Fosfolipider er triglyserider hvor en fettsyrerest er erstattet med en fosforsyrerest; delen av molekylet der resten av fosforsyre befinner seg kalles det hydrofile hodet, delene hvor fettsyrerester befinner seg kalles hydrofobe haler. I membranen er fosfolipider ordnet på en strengt ordnet måte: de hydrofobe halene til molekylene vender mot hverandre, og de hydrofile hodene vender utover, mot vannet.
I tillegg til lipider inneholder membranen proteiner (i gjennomsnitt ≈ 60%). De bestemmer de fleste av de spesifikke funksjonene til membranen (transport av visse molekyler, katalyse av reaksjoner, mottak og konvertering av signaler fra miljøet, etc.). Skill: 1) perifere proteiner(plassert på den ytre eller indre overflaten av lipid-dobbeltlaget), 2) semi-integrerte proteiner(nedsenket i lipid-dobbeltlaget til forskjellige dybder), 3) integrerte eller transmembrane proteiner(gjennomtrenge membranen gjennom og gjennom, mens den er i kontakt med både den ytre og Internt miljø celler). Integrerte proteiner kalles i noen tilfeller kanaldannende, eller kanal, siden de kan betraktes som hydrofile kanaler som polare molekyler passerer inn i cellen (lipidkomponenten i membranen ville ikke slippe dem gjennom).

A - hydrofilt hode av fosfolipidet; C, hydrofobe haler av fosfolipidet; 1 - hydrofobe områder av proteiner E og F; 2, hydrofile områder av protein F; 3 - en forgrenet oligosakkaridkjede festet til et lipid i et glykolipidmolekyl (glykolipider er mindre vanlige enn glykoproteiner); 4 - forgrenet oligosakkaridkjede festet til et protein i et glykoproteinmolekyl; 5 - hydrofil kanal (fungerer som en pore som ioner og noen polare molekyler kan passere gjennom).
Membranen kan inneholde karbohydrater (opptil 10%). Karbohydratkomponenten i membraner er representert av oligosakkarid- eller polysakkaridkjeder assosiert med proteinmolekyler (glykoproteiner) eller lipider (glykolipider). I utgangspunktet er karbohydrater plassert på den ytre overflaten av membranen. Karbohydrater gir reseptorfunksjoner til membranen. I dyreceller danner glykoproteiner et epimembrankompleks, glykokalyxen, flere titalls nanometer tykk. Mange cellereseptorer er lokalisert i den, med dens hjelp oppstår celleadhesjon.
Molekyler av proteiner, karbohydrater og lipider er mobile, i stand til å bevege seg i membranens plan. Tykkelsen på plasmamembranen er omtrent 7,5 nm.
Membranfunksjoner
Membranene utfører følgende funksjoner:
- separasjon av cellulært innhold fra det ytre miljøet,
- regulering av metabolisme mellom cellen og miljøet,
- oppdeling av cellen i rom ("rom"),
- plassering av "enzymatiske transportører",
- gir kommunikasjon mellom celler i vevet til flercellede organismer (adhesjon),
- signalgjenkjenning.
Det viktigste membran egenskap- selektiv permeabilitet, dvs. membraner er svært permeable for noen stoffer eller molekyler og dårlig permeable (eller helt ugjennomtrengelige) for andre. Denne egenskapen ligger til grunn for den regulerende funksjonen til membraner, som sikrer utveksling av stoffer mellom cellen og det ytre miljøet. Prosessen der stoffer passerer gjennom cellemembranen kalles transport av stoffer. Skill: 1) passiv transport- prosessen med å overføre stoffer, uten energi; 2) aktiv transport- prosessen med å sende stoffer, gå med kostnadene for energi.
På passiv transport stoffer beveger seg fra et område med høyere konsentrasjon til et område med lavere, dvs. langs konsentrasjonsgradienten. I enhver løsning er det molekyler av løsningsmidlet og det løste stoffet. Prosessen med bevegelse av oppløste molekyler kalles diffusjon, bevegelsen av løsemiddelmolekyler kalles osmose. Hvis molekylet er ladet, påvirkes transporten av den elektriske gradienten. Derfor snakker man ofte om en elektrokjemisk gradient, som kombinerer begge gradientene sammen. Transporthastigheten avhenger av gradientens størrelse.
Følgende typer passiv transport kan skilles: 1) enkel diffusjon- transport av stoffer direkte gjennom lipid-dobbeltlaget (oksygen, karbondioksid); 2) diffusjon gjennom membrankanaler- transport gjennom kanaldannende proteiner (Na+, K+, Ca 2+, Cl -); 3) forenklet diffusjon- transport av stoffer ved hjelp av spesielle transportproteiner, som hver er ansvarlig for bevegelsen av visse molekyler eller grupper av beslektede molekyler (glukose, aminosyrer, nukleotider); 4) osmose— transport av vannmolekyler (i alt biologiske systemer løsningsmidlet er vann).
Nødvendighet aktiv transport oppstår når det er nødvendig å sikre overføring av molekyler gjennom membranen mot den elektrokjemiske gradienten. Denne transporten utføres av spesielle bærerproteiner, hvis aktivitet krever energiforbruk. Kilden til energi er ATP-molekyler. Aktiv transport inkluderer: 1) Na + /K + -pumpe (natrium-kalium pumpe), 2) endocytose, 3) eksocytose.

Arbeid Na + /K + -pumpe. For normal funksjon må cellen opprettholde et visst forhold mellom K + og Na + ioner i cytoplasmaet og i det ytre miljø. Konsentrasjonen av K + inne i cellen bør være betydelig høyere enn utenfor den, og Na + - omvendt. Det skal bemerkes at Na + og K + fritt kan diffundere gjennom membranporene. Na+/K+-pumpen motvirker utjevningen av disse ionekonsentrasjonene og pumper aktivt Na+ ut av cellen og K+ inn i cellen. Na + /K + -pumpen er et transmembranprotein som er i stand til konformasjonsendringer, slik at det kan feste både K + og Na + . Driftssyklusen til Na + /K + -pumpen kan deles inn i følgende faser: 1) festing av Na + fra innsiden av membranen, 2) fosforylering av pumpeproteinet, 3) frigjøring av Na + i det ekstracellulære plass, 4) festing av K+ fra utsiden av membranen, 5) defosforylering av pumpeproteinet, 6) frigjøring av K+ i det intracellulære rommet. Natrium-kalium-pumpen bruker nesten en tredjedel av all energien som er nødvendig for cellens levetid. I løpet av én driftssyklus pumper pumpen ut 3Na+ fra cellen og pumper inn 2K+.
Endocytose- prosessen med absorpsjon av store partikler og makromolekyler i cellen. Det finnes to typer endocytose: 1) fagocytose- fangst og absorpsjon av store partikler (celler, celledeler, makromolekyler) og 2) pinocytose- fangst og absorpsjon av flytende materiale (løsning, kolloidal løsning, suspensjon). Fenomenet fagocytose ble oppdaget av I.I. Mechnikov i 1882. Under endocytose danner plasmamembranen en invaginasjon, kantene smelter sammen, og strukturene avgrenset fra cytoplasmaet av en enkelt membran snøres inn i cytoplasmaet. Mange protozoer og noen leukocytter er i stand til fagocytose. Pinocytose observeres i epitelcellene i tarmen, i endotelet til blodkapillærene.
Eksocytose- en prosess omvendt til endocytose: utskillelse ulike stoffer fra cellen. Under eksocytose smelter vesikkelmembranen sammen med den ytre cytoplasmatiske membranen, innholdet i vesikkelen fjernes utenfor cellen, og dens membran er inkludert i den ytre cytoplasmatiske membranen. På denne måten skilles hormoner ut fra cellene i de endokrine kjertlene, og i protozoer blir det ufordøyd mat igjen.
Gå til forelesninger nummer 5"Celleteori. Typer mobilorganisasjon»
Gå til forelesninger nummer 7"Eukaryot celle: struktur og funksjoner til organeller"
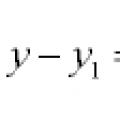 Ligninger av linjer og kurver i planet
Ligninger av linjer og kurver i planet Generalisert homogen likning Generaliserte homogene likninger av andre orden
Generalisert homogen likning Generaliserte homogene likninger av andre orden Takket være kommaet På grunn av det faktum at i deres
Takket være kommaet På grunn av det faktum at i deres