Hvilke molekyler utgjør cellemembranen? Biologiske mekanismer som involverer cellemembranen
Blant Hovedfunksjonene til cellemembranen kan skilles ut som barriere, transport, enzym og reseptor. Cellemembranen (også kalt plasmalemma, plasma- eller cytoplasmatisk membran) beskytter innholdet i cellen eller dens organeller fra miljøet, gir selektiv permeabilitet for stoffer, enzymer er lokalisert på den, samt molekyler som kan "fange" forskjellige kjemiske og fysiske signaler.
Denne funksjonaliteten er gitt av den spesielle strukturen til cellemembranen.
I utviklingen av livet på jorden kunne en celle generelt bare dannes etter at det dukket opp en membran som separerte og stabiliserte det indre innholdet, og forhindret det i å gå i oppløsning.
Når det gjelder å opprettholde homeostase (selvregulering av den relative konstanten til det indre miljøet) barrierefunksjonen til cellemembranen er nært knyttet til transporten.
Små molekyler er i stand til å passere gjennom plasmalemmaet uten noen "hjelpere", langs konsentrasjonsgradienten, dvs. fra et område med høy konsentrasjon av et gitt stoff til et område med lav konsentrasjon. Dette er for eksempel tilfellet for gassene som er involvert i respirasjon. Oksygen og karbondioksid diffunderer gjennom cellemembranen i retningen der konsentrasjonen for øyeblikket er lavere.
Siden membranen for det meste er hydrofob (på grunn av det doble lipidlaget), kan polare (hydrofile) molekyler, selv små, ofte ikke trenge gjennom den. Derfor fungerer en rekke membranproteiner som bærere av slike molekyler, binder seg til dem og transporterer dem gjennom plasmalemmaet.
Integrerte (membran-penetrerende) proteiner opererer ofte etter prinsippet om å åpne og lukke kanaler. Når et molekyl nærmer seg et slikt protein, kobles det til det, og kanalen åpner seg. Dette stoffet eller et annet går gjennom proteinkanalen, hvoretter konformasjonen endres, og kanalen lukkes for dette stoffet, men kan åpne for passasje av en annen. Natrium-kalium-pumpen fungerer i henhold til dette prinsippet, og pumper kaliumioner inn i cellen og pumper natriumioner ut av den.
Enzymatisk funksjon av cellemembranen i større grad implementert på membranene til celleorganeller. De fleste av proteinene som syntetiseres i cellen utfører en enzymatisk funksjon. Når de setter seg ned på membranen i en bestemt rekkefølge, organiserer de en transportør når reaksjonsproduktet katalysert av ett enzymprotein går over til det neste. En slik "pipeline" stabiliserer overflateproteinene i plasmalemmaet.
Til tross for universaliteten til strukturen til alle biologiske membraner (de er bygget i henhold til et enkelt prinsipp, de er nesten like i alle organismer og i forskjellige membrancellestrukturer), kan deres kjemiske sammensetning fortsatt variere. Det er mer flytende og mer faste, noen har mer sikre proteiner, andre mindre. I tillegg er forskjellige sider (indre og ytre) av samme membran også forskjellige.
Membranen som omgir cellen (cytoplasmatisk) på utsiden har mange karbohydratkjeder knyttet til lipider eller proteiner (som et resultat dannes glykolipider og glykoproteiner). Mange av disse karbohydratene reseptor funksjon, som er mottakelig for visse hormoner, fanger opp endringer i fysiske og kjemiske indikatorer i miljøet.
Hvis for eksempel et hormon binder seg til sin cellulære reseptor, så endrer karbohydratdelen av reseptormolekylet sin struktur, etterfulgt av en endring i strukturen til den assosierte proteindelen som trenger inn i membranen. På neste trinn startes eller stoppes forskjellige biokjemiske reaksjoner i cellen, dvs. metabolismen endres, og den cellulære responsen på "irritanten" begynner.
I tillegg til de oppførte fire funksjonene til cellemembranen, skilles andre ut: matrise, energi, markering, dannelse av intercellulære kontakter, etc. De kan imidlertid betraktes som "underfunksjoner" av de som allerede er vurdert.
cellemembran
Bilde av en cellemembran. Små blå og hvite kuler tilsvarer de hydrofobe "hodene" til fosfolipidene, og linjene festet til dem tilsvarer de hydrofile "halene". Figuren viser kun integrerte membranproteiner (røde kuler og gule spiraler). Gule ovale prikker inne i membranen - kolesterolmolekyler Gulgrønne kjeder av perler på utsiden av membranen - oligosakkaridkjeder som danner glykokalyxen
Den biologiske membranen inkluderer også ulike proteiner: integral (penetrerer membranen gjennom), semi-integral (nedsenket i den ene enden inn i det ytre eller indre lipidlaget), overflate (plassert på den ytre eller ved siden av de indre sidene av membranen). Noen proteiner er kontaktpunktene til cellemembranen med cytoskjelettet inne i cellen, og celleveggen (hvis noen) utenfor. Noen av de integrerte proteinene fungerer som ionekanaler, ulike transportører og reseptorer.
Funksjoner
- barriere - gir en regulert, selektiv, passiv og aktiv metabolisme med omgivelsene. For eksempel beskytter peroksisommembranen cytoplasmaet mot peroksider som er farlige for cellen. Selektiv permeabilitet betyr at permeabiliteten til en membran for ulike atomer eller molekyler avhenger av deres størrelse, elektrisk ladning og kjemiske egenskaper. Selektiv permeabilitet sikrer separasjon av cellen og cellulære rom fra miljøet og forsyner dem med de nødvendige stoffene.
- transport - gjennom membranen foregår det en transport av stoffer inn i cellen og ut av cellen. Transport gjennom membranene gir: tilførsel av næringsstoffer, fjerning av sluttprodukter av metabolisme, sekresjon av forskjellige stoffer, dannelse av ioniske gradienter, opprettholdelse av den optimale konsentrasjonen av ioner i cellen, som er nødvendig for funksjon av cellulære enzymer.
Partikler som av en eller annen grunn ikke er i stand til å krysse fosfolipid-dobbeltlaget (for eksempel på grunn av hydrofile egenskaper, siden membranen er hydrofob inni og ikke lar hydrofile stoffer passere gjennom, eller på grunn av deres store størrelse), men nødvendige for cellen , kan penetrere membranen gjennom spesielle bærerproteiner (transportører) og kanalproteiner eller ved endocytose.
Ved passiv transport krysser stoffer lipid-dobbeltlaget uten energiforbruk langs konsentrasjonsgradienten ved diffusjon. En variant av denne mekanismen er forenklet diffusjon, der et spesifikt molekyl hjelper et stoff å passere gjennom membranen. Dette molekylet kan ha en kanal som lar bare én type stoff passere gjennom.
Aktiv transport krever energi, da den skjer mot en konsentrasjonsgradient. Det er spesielle pumpeproteiner på membranen, inkludert ATPase, som aktivt pumper kaliumioner (K+) inn i cellen og pumper natriumioner (Na+) ut av den. - matrise - gir en viss relativ posisjon og orientering av membranproteiner, deres optimale interaksjon.
- mekanisk - sikrer cellens autonomi, dens intracellulære strukturer, samt forbindelse med andre celler (i vev). Cellevegger spiller en viktig rolle i å gi mekanisk funksjon, og hos dyr - intercellulær substans.
- energi - under fotosyntese i kloroplaster og cellulær respirasjon i mitokondrier, opererer energioverføringssystemer i deres membraner, der proteiner også deltar;
- reseptor - noen proteiner som ligger i membranen er reseptorer (molekyler som cellen oppfatter visse signaler med).
For eksempel virker hormoner som sirkulerer i blodet kun på målceller som har reseptorer som tilsvarer disse hormonene. Nevrotransmittere (kjemikalier som leder nerveimpulser) binder seg også til spesifikke reseptorproteiner på målceller. - enzymatiske - membranproteiner er ofte enzymer. For eksempel inneholder plasmamembranene til tarmepitelceller fordøyelsesenzymer.
- implementering av generering og ledning av biopotensialer.
Ved hjelp av membranen opprettholdes en konstant konsentrasjon av ioner i cellen: konsentrasjonen av K + ion inne i cellen er mye høyere enn utenfor, og konsentrasjonen av Na + er mye lavere, noe som er veldig viktig, siden dette opprettholder potensialforskjellen over membranen og genererer en nerveimpuls. - cellemarkering - det er antigener på membranen som fungerer som markører - "merker" som gjør at cellen kan identifiseres. Dette er glykoproteiner (det vil si proteiner med forgrenede oligosakkaridsidekjeder festet til dem) som spiller rollen som "antenner". På grunn av mylderet av sidekjedekonfigurasjoner er det mulig å lage en spesifikk markør for hver celletype. Ved hjelp av markører kan celler gjenkjenne andre celler og handle i samspill med dem, for eksempel når de danner organer og vev. Det lar også immunsystemet gjenkjenne fremmede antigener.
Struktur og sammensetning av biomembraner
Membraner er sammensatt av tre klasser av lipider: fosfolipider, glykolipider og kolesterol. Fosfolipider og glykolipider (lipider med karbohydrater festet til dem) består av to lange hydrofobe hydrokarbon-"haler" som er assosiert med et ladet hydrofilt "hode". Kolesterol stivner membranen ved å okkupere det ledige rommet mellom de hydrofobe lipidhalene og hindre dem i å bøye seg. Derfor er membraner med lavt kolesterolinnhold mer fleksible, mens de med høyt kolesterolinnhold er mer stive og sprø. Kolesterol fungerer også som en "stopper" som forhindrer bevegelse av polare molekyler fra og inn i cellen. En viktig del av membranen består av proteiner som trenger inn i den og er ansvarlige for ulike egenskaper til membraner. Deres sammensetning og orientering i forskjellige membraner er forskjellig.
Cellemembraner er ofte asymmetriske, det vil si at lagene er forskjellige i lipidsammensetning, overgangen til et individuelt molekyl fra ett lag til et annet (den s.k. flip flop) er vanskelig.
Membranorganeller
Disse er lukkede enkelt- eller sammenkoblede deler av cytoplasmaet, atskilt fra hyaloplasmaet med membraner. Enkeltmembranorganeller inkluderer endoplasmatisk retikulum, Golgi-apparater, lysosomer, vakuoler, peroksisomer; til to-membran - kjerne, mitokondrier, plastider. Strukturen til membranene til forskjellige organeller er forskjellig i sammensetningen av lipider og membranproteiner.
Selektiv permeabilitet
Cellemembraner har selektiv permeabilitet: glukose, aminosyrer, fettsyrer, glyserol og ioner diffunderer sakte gjennom dem, og membranene selv regulerer aktivt denne prosessen til en viss grad - noen stoffer passerer gjennom, mens andre ikke gjør det. Det er fire hovedmekanismer for inntreden av stoffer i cellen eller fjerning av dem fra cellen til utsiden: diffusjon, osmose, aktiv transport og ekso- eller endocytose. De to første prosessene er passive i naturen, det vil si at de ikke krever energi; de to siste er aktive prosesser knyttet til energiforbruk.
Den selektive permeabiliteten til membranen under passiv transport skyldes spesielle kanaler - integrerte proteiner. De trenger gjennom og gjennom membranen, og danner en slags passasje. Grunnstoffene K, Na og Cl har sine egne kanaler. Med hensyn til konsentrasjonsgradienten beveger molekylene til disse elementene seg inn og ut av cellen. Ved irritasjon åpner natriumionekanalene seg, og det er en kraftig tilstrømning av natriumioner inn i cellen. Dette resulterer i en ubalanse i membranpotensialet. Etter det gjenopprettes membranpotensialet. Kaliumkanaler er alltid åpne, gjennom hvilke kaliumioner sakte kommer inn i cellen.
se også
Litteratur
- Antonov V. F., Smirnova E. N., Shevchenko E. V. Lipidmembraner under faseoverganger. - M .: Nauka, 1994.
- Gennis R. Biomembraner. Molekylær struktur og funksjoner: oversettelse fra engelsk. = Biomembraner. Molekylær struktur og funksjon (av Robert B. Gennis). - 1. utgave. - M .: Mir, 1997. - ISBN 5-03-002419-0
- Ivanov V. G., Berestovsky T. N. lipid-dobbeltlag av biologiske membraner. - M .: Nauka, 1982.
- Rubin A.B. Biofysikk, lærebok i 2 bind. - 3. utgave, revidert og utvidet. - M .: Moscow University Press, 2004. -
Membraner er ekstremt viskøse og samtidig plastiske strukturer som omgir alle levende celler. Funksjoner cellemembraner:
1. Plasmamembranen er en barriere som opprettholder en annen sammensetning av det ekstra- og intracellulære miljøet.
2. Membraner danner spesialiserte rom inne i cellen, dvs. mange organeller - mitokondrier, lysosomer, Golgi-kompleks, endoplasmatisk retikulum, kjernemembraner.
3. Enzymer involvert i energiomdannelse i prosesser som oksidativ fosforylering og fotosyntese er lokalisert i membraner.
Struktur og sammensetning av membraner
Grunnlaget for membranen er et lipid-dobbeltlag, i dannelsen som fosfolipider og glykolipider deltar. Lipid-dobbeltlaget er dannet av to rader med lipider, hvis hydrofobe radikaler er skjult inni, og de hydrofile gruppene er vendt utover og er i kontakt med det vandige mediet. Proteinmolekyler ser ut til å være "oppløst" i lipid-dobbeltlaget.
Struktur av membranlipider
Membranlipider er amfifile molekyler, pga molekylet har både en hydrofil region (polare hoder) og en hydrofob region, representert av hydrokarbonradikaler av fettsyrer, som spontant danner et dobbeltlag. Det er tre hovedtyper av lipider i membraner: fosfolipider, glykolipider og kolesterol.
Lipidsammensetningen er forskjellig. Innholdet av et eller annet lipid bestemmes tilsynelatende av mangfoldet av funksjoner som utføres av disse lipidene i membraner.
Fosfolipider. Alle fosfolipider kan deles inn i to grupper - glyserofosfolipider og sfinfosfolipider. Glyserofosfolipider er klassifisert som derivater av fosfatidinsyre. De vanligste glyserofosfolipidene er fosfatidylkoliner og fosfatidyletanolaminer. Sfingofosfolipider er basert på aminoalkoholen sfingosin.
Glykolipider. I glykolipider er den hydrofobe delen representert av alkoholceramid, og den hydrofile delen er representert av en karbohydratrest. Avhengig av lengden og strukturen til karbohydratdelen, skilles cerebrosider og gangliosider. Polare "hoder" av glykolipider er plassert på den ytre overflaten av plasmamembraner.
Kolesterol (CS). CS er tilstede i alle membraner av dyreceller. Molekylet består av en stiv hydrofob kjerne og en fleksibel hydrokarbonkjede. Den eneste hydroksylgruppen i 3-posisjonen er det "polare hodet". For en dyrecelle er det gjennomsnittlige molforholdet mellom kolesterol / fosfolipider 0,3-0,4, men i plasmamembranen er dette forholdet mye høyere (0,8-0,9). Tilstedeværelsen av kolesterol i membraner reduserer mobiliteten til fettsyrer, reduserer lateral diffusjon av lipider, og kan derfor påvirke funksjonene til membranproteiner.
Membranegenskaper:
1. Selektiv permeabilitet. Det lukkede dobbeltlaget gir en av hovedegenskapene til membranen: den er ugjennomtrengelig for de fleste vannløselige molekyler, siden de ikke løses opp i dens hydrofobe kjerne. Gasser som oksygen, CO 2 og nitrogen har evnen til lett å trenge inn i cellen på grunn av molekylenes lille størrelse og svak interaksjon med løsemidler. Også molekyler av lipid natur, for eksempel steroidhormoner, trenger lett gjennom dobbeltlaget.
2. Likviditet. Membranene er preget av fluiditet (fluiditet), lipiders og proteiners evne til å bevege seg. To typer fosfolipidbevegelser er mulige: salto (kalt "flip-flop" i vitenskapelig litteratur) og lateral diffusjon. I det første tilfellet snur fosfolipidmolekyler mot hverandre i det bimolekylære laget (eller salto) mot hverandre og bytter plass i membranen, d.v.s. utsiden blir innsiden og omvendt. Slike hopp er forbundet med energiforbruk. Oftere observeres rotasjoner rundt aksen (rotasjon) og lateral diffusjon - bevegelse innenfor laget parallelt med membranoverflaten. Bevegelseshastigheten til molekyler avhenger av mikroviskositeten til membraner, som igjen bestemmes av det relative innholdet av mettede og umettede fettsyrer i sammensetningen av lipider. Mikroviskositeten er lavere hvis umettede fettsyrer dominerer i sammensetningen av lipider, og høyere hvis innholdet av mettede fettsyrer er høyt.
3. Asymmetri av membraner. Overflatene til den samme membranen er forskjellige i sammensetningen av lipider, proteiner og karbohydrater (tverrgående asymmetri). For eksempel dominerer fosfatidylkoliner i det ytre laget, mens fosfatidyletanolaminer og fosfatidylseriner dominerer i det indre laget. Karbohydratkomponentene til glykoproteiner og glykolipider kommer til den ytre overflaten og danner en kontinuerlig pose kalt glykokalyxen. Det er ingen karbohydrater på den indre overflaten. Proteiner - hormonreseptorer er lokalisert på den ytre overflaten av plasmamembranen, og enzymene regulert av dem - adenylatcyklase, fosfolipase C - på innsiden, etc.
Membranproteiner
Membranfosfolipider fungerer som et løsningsmiddel for membranproteiner, og skaper et mikromiljø der sistnevnte kan fungere. Proteiner står for 30 til 70% av massen av membraner. Antall ulike proteiner i membranen varierer fra 6-8 i det sarkoplasmatiske retikulum til mer enn 100 i plasmamembranen. Dette er enzymer, transportproteiner, strukturelle proteiner, antigener, inkludert antigener it, reseptorer for ulike molekyler.
Ved lokalisering i membranen deles proteiner inn i integrert (delvis eller fullstendig nedsenket i membranen) og perifert (plassert på overflaten). Noen integrerte proteiner krysser membranen én gang (glykoforin), mens andre krysser membranen mange ganger. For eksempel krysser den retinale fotoreseptoren og β 2 -adrenerge reseptoren dobbeltlaget 7 ganger.
Perifere proteiner og domener av integrerte proteiner lokalisert på den ytre overflaten av alle membraner er nesten alltid glykosylerte. Oligosakkaridrester beskytter proteinet mot proteolyse og er også involvert i ligandgjenkjenning eller adhesjon.
Cytoplasma- en obligatorisk del av cellen, innelukket mellom plasmamembranen og kjernen; Det er delt inn i hyaloplasma (hovedstoffet i cytoplasma), organeller (permanente komponenter i cytoplasma) og inneslutninger (midlertidige komponenter i cytoplasma). Den kjemiske sammensetningen av cytoplasma: grunnlaget er vann (60-90% av den totale massen av cytoplasma), ulike organiske og uorganiske forbindelser. Cytoplasmaet er alkalisk. Et karakteristisk trekk ved cytoplasmaet til en eukaryot celle er konstant bevegelse ( syklose). Det oppdages først og fremst ved bevegelse av celleorganeller, slik som kloroplaster. Hvis bevegelsen til cytoplasmaet stopper, dør cellen, siden den bare er i konstant bevegelse kan utføre sine funksjoner.
Hyaloplasma ( cytosol) er en fargeløs, slimete, tykk og gjennomsiktig kolloidal løsning. Det er i det alle metabolske prosesser finner sted, det gir sammenkoblingen av kjernen og alle organeller. Avhengig av overvekt av den flytende delen eller store molekyler i hyaloplasma, skilles to former for hyaloplasma: sol- mer flytende hyaloplasma og gel- tettere hyaloplasma. Gjensidige overganger er mulige mellom dem: gelen blir til en sol og omvendt.
Funksjoner av cytoplasma:
- integrering av alle komponenter i cellen i et enkelt system,
- miljø for passasje av mange biokjemiske og fysiologiske prosesser,
- miljø for eksistensen og funksjonen til organeller.
Cellevegger
Cellevegger begrense eukaryote celler. Minst to lag kan skilles i hver cellemembran. Det indre laget er ved siden av cytoplasmaet og er representert ved plasmamembran(synonymer - plasmalemma, cellemembran, cytoplasmatisk membran), som det ytre laget dannes over. I en dyrecelle er den tynn og kalles glykokalyx(dannet av glykoproteiner, glykolipider, lipoproteiner), i en plantecelle - tykk, kalt celleveggen(dannet av cellulose).
Alle biologiske membraner har felles strukturelle trekk og egenskaper. Foreløpig generelt akseptert flytende mosaikkmodell av membranstrukturen. Grunnlaget for membranen er et lipid-dobbeltlag, hovedsakelig dannet av fosfolipider. Fosfolipider er triglyserider hvor en fettsyrerest er erstattet med en fosforsyrerest; delen av molekylet der resten av fosforsyre befinner seg kalles det hydrofile hodet, delene hvor fettsyrerester befinner seg kalles hydrofobe haler. I membranen er fosfolipider ordnet på en strengt ordnet måte: de hydrofobe halene til molekylene vender mot hverandre, og de hydrofile hodene vender utover, mot vannet.
I tillegg til lipider inneholder membranen proteiner (i gjennomsnitt ≈ 60%). De bestemmer de fleste av de spesifikke funksjonene til membranen (transport av visse molekyler, katalyse av reaksjoner, mottak og konvertering av signaler fra miljøet, etc.). Skill: 1) perifere proteiner(plassert på den ytre eller indre overflaten av lipid-dobbeltlaget), 2) semi-integrerte proteiner(nedsenket i lipid-dobbeltlaget til forskjellige dybder), 3) integrerte eller transmembrane proteiner(gjennomtrenge membranen gjennom og gjennom, mens den er i kontakt med både det ytre og indre miljøet i cellen). Integrerte proteiner kalles i noen tilfeller kanaldannende, eller kanal, siden de kan betraktes som hydrofile kanaler som polare molekyler passerer inn i cellen (lipidkomponenten i membranen ville ikke slippe dem gjennom).

A - hydrofilt hode av fosfolipidet; C, hydrofobe haler av fosfolipidet; 1 - hydrofobe områder av proteiner E og F; 2, hydrofile områder av protein F; 3 - en forgrenet oligosakkaridkjede festet til et lipid i et glykolipidmolekyl (glykolipider er mindre vanlige enn glykoproteiner); 4 - forgrenet oligosakkaridkjede festet til et protein i et glykoproteinmolekyl; 5 - hydrofil kanal (fungerer som en pore som ioner og noen polare molekyler kan passere gjennom).
Membranen kan inneholde karbohydrater (opptil 10%). Karbohydratkomponenten i membraner er representert av oligosakkarid- eller polysakkaridkjeder assosiert med proteinmolekyler (glykoproteiner) eller lipider (glykolipider). I utgangspunktet er karbohydrater plassert på den ytre overflaten av membranen. Karbohydrater gir reseptorfunksjoner til membranen. I dyreceller danner glykoproteiner et epimembrankompleks, glykokalyxen, flere titalls nanometer tykk. Mange cellereseptorer er lokalisert i den, med dens hjelp oppstår celleadhesjon.
Molekyler av proteiner, karbohydrater og lipider er mobile, i stand til å bevege seg i membranens plan. Tykkelsen på plasmamembranen er omtrent 7,5 nm.
Membranfunksjoner
Membranene utfører følgende funksjoner:
- separasjon av cellulært innhold fra det ytre miljøet,
- regulering av metabolisme mellom cellen og miljøet,
- oppdeling av cellen i rom ("rom"),
- plassering av "enzymatiske transportører",
- gir kommunikasjon mellom celler i vevet til flercellede organismer (adhesjon),
- signalgjenkjenning.
Det viktigste membran egenskap- selektiv permeabilitet, dvs. membraner er svært permeable for noen stoffer eller molekyler og dårlig permeable (eller helt ugjennomtrengelige) for andre. Denne egenskapen ligger til grunn for den regulerende funksjonen til membraner, som sikrer utveksling av stoffer mellom cellen og det ytre miljøet. Prosessen der stoffer passerer gjennom cellemembranen kalles transport av stoffer. Skill: 1) passiv transport- prosessen med å overføre stoffer, uten energi; 2) aktiv transport- prosessen med å sende stoffer, gå med kostnadene for energi.
På passiv transport stoffer beveger seg fra et område med høyere konsentrasjon til et område med lavere, dvs. langs konsentrasjonsgradienten. I enhver løsning er det molekyler av løsningsmidlet og det løste stoffet. Prosessen med bevegelse av oppløste molekyler kalles diffusjon, bevegelsen av løsemiddelmolekyler kalles osmose. Hvis molekylet er ladet, påvirkes transporten av den elektriske gradienten. Derfor snakker man ofte om en elektrokjemisk gradient, som kombinerer begge gradientene sammen. Transporthastigheten avhenger av gradientens størrelse.
Følgende typer passiv transport kan skilles: 1) enkel diffusjon- transport av stoffer direkte gjennom lipid-dobbeltlaget (oksygen, karbondioksid); 2) diffusjon gjennom membrankanaler- transport gjennom kanaldannende proteiner (Na+, K+, Ca 2+, Cl -); 3) forenklet diffusjon- transport av stoffer ved hjelp av spesielle transportproteiner, som hver er ansvarlig for bevegelsen av visse molekyler eller grupper av beslektede molekyler (glukose, aminosyrer, nukleotider); 4) osmose- transport av vannmolekyler (i alle biologiske systemer er vann løsningsmidlet).
Nødvendighet aktiv transport oppstår når det er nødvendig å sikre overføring av molekyler gjennom membranen mot den elektrokjemiske gradienten. Denne transporten utføres av spesielle bærerproteiner, hvis aktivitet krever energiforbruk. Energikilden er ATP-molekyler. Aktiv transport inkluderer: 1) Na + /K + -pumpe (natrium-kalium pumpe), 2) endocytose, 3) eksocytose.

Arbeid Na + /K + -pumpe. For normal funksjon må cellen opprettholde et visst forhold mellom K + og Na + ioner i cytoplasmaet og i det ytre miljø. Konsentrasjonen av K + inne i cellen bør være betydelig høyere enn utenfor den, og Na + - omvendt. Det skal bemerkes at Na + og K + fritt kan diffundere gjennom membranporene. Na+/K+-pumpen motvirker utjevningen av disse ionekonsentrasjonene og pumper aktivt Na+ ut av cellen og K+ inn i cellen. Na + /K + -pumpen er et transmembranprotein som er i stand til konformasjonsendringer, slik at det kan feste både K + og Na + . Driftssyklusen til Na + /K + -pumpen kan deles inn i følgende faser: 1) festing av Na + fra innsiden av membranen, 2) fosforylering av pumpeproteinet, 3) frigjøring av Na + i det ekstracellulære plass, 4) festing av K+ fra utsiden av membranen, 5) defosforylering av pumpeproteinet, 6) frigjøring av K+ i det intracellulære rommet. Natrium-kalium-pumpen bruker nesten en tredjedel av all energien som er nødvendig for cellens levetid. I løpet av én driftssyklus pumper pumpen ut 3Na+ fra cellen og pumper inn 2K+.
Endocytose- prosessen med absorpsjon av store partikler og makromolekyler i cellen. Det finnes to typer endocytose: 1) fagocytose- fangst og absorpsjon av store partikler (celler, celledeler, makromolekyler) og 2) pinocytose- fangst og absorpsjon av flytende materiale (løsning, kolloidal løsning, suspensjon). Fenomenet fagocytose ble oppdaget av I.I. Mechnikov i 1882. Under endocytose danner plasmamembranen en invaginasjon, kantene smelter sammen, og strukturene avgrenset fra cytoplasmaet av en enkelt membran snøres inn i cytoplasmaet. Mange protozoer og noen leukocytter er i stand til fagocytose. Pinocytose observeres i epitelcellene i tarmen, i endotelet til blodkapillærene.
Eksocytose- den omvendte prosessen med endocytose: fjerning av forskjellige stoffer fra cellen. Under eksocytose smelter vesikkelmembranen sammen med den ytre cytoplasmatiske membranen, innholdet i vesikkelen fjernes utenfor cellen, og dens membran er inkludert i den ytre cytoplasmatiske membranen. På denne måten skilles hormoner ut fra cellene i de endokrine kjertlene, og i protozoer blir det ufordøyd mat igjen.
Gå til forelesninger nummer 5"Celleteori. Typer mobilorganisasjon»
Gå til forelesninger nummer 7"Eukaryot celle: struktur og funksjoner til organeller"
Kort beskrivelse:
Sazonov V.F. 1_1 Strukturen til cellemembranen [Elektronisk ressurs] // Kinesiolog, 2009-2018: [nettsted]. Dato for oppdatering: 06.02.2018...__.201_). _Strukturen og funksjonen til cellemembranen er beskrevet (synonymer: plasmalemma, plasmolemma, biomembran, cellemembran, ytre cellemembran, cellemembran, cytoplasmatisk membran). Denne innledende informasjonen er nødvendig både for cytologi og for å forstå prosessene med nervøs aktivitet: nervøs eksitasjon, hemming, arbeidet med synapser og sensoriske reseptorer.
cellemembran (plasma EN lemma eller plasma O lemma)
Begrepsdefinisjon
Cellemembranen (synonymer: plasmalemma, plasmolemma, cytoplasmatisk membran, biomembran) er en trippel lipoprotein (dvs. "fett-protein") membran som skiller cellen fra miljøet og utfører en kontrollert utveksling og kommunikasjon mellom cellen og dens omgivelser.
Hovedsaken i denne definisjonen er ikke at membranen skiller cellen fra miljøet, men bare at den kobler til celle med miljøet. Membranen er aktiv strukturen til cellen, fungerer den hele tiden.
En biologisk membran er en ultratynn bimolekylær film av fosfolipider dekket med proteiner og polysakkarider. Denne cellulære strukturen ligger til grunn for barriere-, mekaniske- og matriseegenskapene til en levende organisme (Antonov VF, 1996).
Figurativ representasjon av membranen
For meg fremstår cellemembranen som et gittergjerde med mange dører i, som omgir et bestemt territorium. Alle små levende skapninger kan fritt bevege seg frem og tilbake gjennom dette gjerdet. Men større besøkende kan bare komme inn gjennom dørene, og selv da ikke alle. Ulike besøkende har kun nøkler til sine egne dører, og de kan ikke gå gjennom andres dører. Så gjennom dette gjerdet er det stadig strømmer av besøkende frem og tilbake, fordi hovedfunksjonen til membrangjerdet er todelt: å skille territoriet fra det omkringliggende rommet og samtidig forbinde det med det omkringliggende rommet. For dette er det mange hull og dører i gjerdet - !
Membranegenskaper
1. Permeabilitet.
2. Semi-permeabilitet (delvis permeabilitet).
3. Selektiv (synonym: selektiv) permeabilitet.
4. Aktiv permeabilitet (synonym: aktiv transport).
5. Kontrollert permeabilitet.
Som du kan se, er hovedegenskapen til membranen dens permeabilitet med hensyn til forskjellige stoffer.
6. Fagocytose og pinocytose.
7. Eksocytose.
8. Tilstedeværelsen av elektriske og kjemiske potensialer, mer presist, potensialforskjellen mellom de indre og ytre sidene av membranen. I overført betydning kan man si det "membranen gjør cellen til et "elektrisk batteri" ved å kontrollere ionestrømmer". Detaljer: .
9. Endringer i elektrisk og kjemisk potensial.
10. Irritabilitet. Spesielle molekylære reseptorer lokalisert på membranen kan forbindes med signal(kontroll)stoffer, som et resultat av at tilstanden til membranen og hele cellen kan endres. Molekylære reseptorer utløser biokjemiske reaksjoner som respons på kombinasjonen av ligander (kontrollstoffer) med dem. Det er viktig å merke seg at signalstoffet virker på reseptoren fra utsiden, mens endringene fortsetter inne i cellen. Det viser seg at membranen overførte informasjon fra miljøet til det indre miljøet i cellen.
11. Katalytisk enzymatisk aktivitet. Enzymer kan være innebygd i membranen eller assosiert med overflaten (både innenfor og utenfor cellen), og der utfører de sin enzymatiske aktivitet.
12. Endre formen på overflaten og området. Dette gjør at membranen kan danne utvekster utover eller omvendt invaginasjoner inn i cellen.
13. Evnen til å danne kontakter med andre cellemembraner.
14. Vedheft - evnen til å holde seg til faste overflater.
Kort liste over membranegenskaper
- Permeabilitet.
- Endocytose, eksocytose, transcytose.
- Potensialer.
- Irritabilitet.
- enzymatisk aktivitet.
- Kontakter.
- Adhesjon.
Membranfunksjoner
1. Ufullstendig isolasjon av internt innhold fra det ytre miljøet.
2. Hovedsaken i arbeidet med cellemembranen er Utveksling diverse stoffer mellom cellen og det ekstracellulære miljøet. Dette skyldes slike egenskaper ved membranen som permeabilitet. I tillegg regulerer membranen denne utvekslingen ved å regulere dens permeabilitet.
3. En annen viktig funksjon av membranen er skape en forskjell i kjemiske og elektriske potensialer mellom dens indre og ytre side. På grunn av dette, inne i cellen har et negativt elektrisk potensial -.
4. Gjennom membranen utføres også informasjonsutveksling mellom cellen og dens omgivelser. Spesielle molekylære reseptorer lokalisert på membranen kan binde seg til kontrollstoffer (hormoner, mediatorer, modulatorer) og utløse biokjemiske reaksjoner i cellen, noe som fører til ulike endringer i cellen eller i dens strukturer.
Video:Strukturen til cellemembranen
Videoforelesning:Detaljer om strukturen til membranen og transport
Membranstruktur
Cellemembranen har en universell tre-lags struktur. Dets medianfettlag er kontinuerlig, og de øvre og nedre proteinlagene dekker det i form av en mosaikk av individuelle proteinområder. Fettlaget er grunnlaget som sikrer isolering av cellen fra miljøet, isolerer den fra miljøet. I seg selv passerer den vannløselige stoffer svært dårlig, men passerer lett fettløselige. Derfor må permeabiliteten til membranen for vannløselige stoffer (for eksempel ioner) forsynes med spesielle proteinstrukturer - og.
Nedenfor er mikrofotografier av ekte cellemembraner av kontaktceller, oppnådd ved hjelp av et elektronmikroskop, samt en skjematisk tegning som viser den trelagsmembranen og mosaikken til proteinlagene. For å forstørre et bilde, klikk på det.


Separat bilde av det indre lipidlaget (fett) av cellemembranen, gjennomsyret av integrerte innebygde proteiner. De øvre og nedre proteinlagene fjernes for ikke å forstyrre vurderingen av lipid-dobbeltlaget

Figur over: En ufullstendig skjematisk fremstilling av cellemembranen (celleveggen) fra Wikipedia.
Merk at de ytre og indre proteinlagene er fjernet fra membranen her slik at vi bedre kan se det sentrale fettdobbelte lipidlaget. I en ekte cellemembran flyter store protein-"øyer" over og under langs fettfilmen (små kuler i figuren), og membranen viser seg å være tykkere, trelags: protein-fett-protein . Så det er faktisk som en sandwich av to protein-"brødskiver" med et tykt lag "smør" i midten, dvs. har en tre-lags struktur, ikke en to-lags.
I denne figuren tilsvarer små blå og hvite kuler de hydrofile (fuktbare) "hodene" til lipidene, og "strengene" som er festet til dem, tilsvarer de hydrofobe (ikke-fuktbare) "halene". Av proteinene er kun integrerte ende-til-ende membranproteiner (røde kuler og gule helixer) vist. Gule ovale prikker inne i membranen er kolesterolmolekyler Gulgrønne kjeder av perler på utsiden av membranen er oligosakkaridkjeder som danner glykokalyxen. Glycocalyx er som et karbohydrat ("sukker") "lo" på membranen, dannet av lange karbohydrat-proteinmolekyler som stikker ut fra den.
Living er en liten "protein-fettpose" fylt med halvflytende gelélignende innhold, som penetreres av filmer og rør.
Veggene i denne posen er dannet av en dobbel fett- (lipid) film, dekket innvendig og utvendig med proteiner - cellemembranen. Derfor sies membranen å ha tre-lags struktur : proteiner-fett-proteiner. Inne i cellen er det også mange lignende fettmembraner som deler dens indre plass i rom. Cellulære organeller er omgitt av de samme membranene: kjerne, mitokondrier, kloroplaster. Så membranen er en universell molekylær struktur som er iboende i alle celler og alle levende organismer.

Til venstre - ikke lenger en ekte, men en kunstig modell av et stykke av en biologisk membran: dette er et øyeblikksbilde av et fettfosfolipid-dobbeltlag (dvs. et dobbeltlag) i ferd med sin molekylære dynamikkmodellering. Beregningscellen til modellen er vist - 96 PQ-molekyler ( f osfatidil X olin) og 2304 vannmolekyler, totalt 20544 atomer.
Til høyre er en visuell modell av et enkelt molekyl av samme lipid, hvorfra membranlipid-dobbeltlaget er satt sammen. Den har et hydrofilt (vannelskende) hode på toppen, og to hydrofobe (vannfryktende) haler nederst. Dette lipidet har et enkelt navn: 1-steroyl-2-docosahexaenoyl-Sn-glycero-3-fosfatidylkolin (18:0/22:6(n-3)cis PC), men du trenger ikke å huske det med mindre du planlegger å få læreren din til å besvime med dybden av kunnskapen din.
Du kan gi en mer presis vitenskapelig definisjon av en celle:
er et ordnet, strukturert heterogent system av biopolymerer begrenset av en aktiv membran, som deltar i et enkelt sett av metabolske, energi- og informasjonsprosesser, og også opprettholder og reproduserer hele systemet som helhet.
Inne i cellen penetreres også membraner, og mellom membranene er det ikke vann, men en viskøs gel/sol med variabel tetthet. Derfor flyter ikke de interagerende molekylene i cellen fritt, som i et reagensrør med en vandig løsning, men sitter (immobilisert) mest på polymerstrukturene i cytoskjelettet eller intracellulære membraner. Og derfor foregår kjemiske reaksjoner inne i cellen nesten som i et fast legeme, og ikke i en væske. Den ytre membranen som omgir cellen er også dekket av enzymer og molekylære reseptorer, noe som gjør den til en veldig aktiv del av cellen.
Cellemembranen (plasmalemma, plasmolemma) er et aktivt skall som skiller cellen fra miljøet og forbinder den med miljøet. © Sazonov V.F., 2016.
Fra denne definisjonen av en membran følger det at den ikke bare begrenser cellen, men jobber aktivt knytte den til omgivelsene.
Fettet som utgjør membranene er spesielt, så molekylene kalles vanligvis ikke bare fett, men lipider, fosfolipider, sfingolipider. Membranfilmen er dobbel, dvs. den består av to filmer klistret sammen. Derfor skriver lærebøker at bunnen av cellemembranen består av to lipidlag (eller " dobbeltlag", dvs. dobbeltlag). For hvert enkelt lipidlag kan den ene siden fuktes av vann, og den andre ikke. Så disse filmene fester seg sammen med hverandre nøyaktig på sidene som ikke fukter.
bakteriemembran
Skallet til en prokaryot celle av gramnegative bakterier består av flere lag, vist i figuren nedenfor.
Lag av skallet av gram-negative bakterier:
1. Den indre tre-lags cytoplasmatiske membranen, som er i kontakt med cytoplasma.
2. Cellevegg, som består av murein.
3. Den ytre trelags cytoplasmatiske membranen, som har samme system av lipider med proteinkomplekser som den indre membranen.
Kommunikasjon av gram-negative bakterieceller med omverdenen gjennom en så kompleks tre-trinns struktur gir dem ikke en fordel i å overleve under tøffe forhold sammenlignet med gram-positive bakterier som har et mindre kraftig skall. De tåler like dårlig høye temperaturer, høy surhet og trykkfall.




Videoforelesning:Plasmamembran. E.V. Cheval, Ph.D.
Videoforelesning:Membranen som cellegrense. A. Iljaskin
Viktigheten av membranionekanaler
Det er lett å forstå at bare fettløselige stoffer kan komme inn i cellen gjennom membranens fettfilm. Dette er fett, alkoholer, gasser. For eksempel, i erytrocytter passerer oksygen og karbondioksid lett inn og ut direkte gjennom membranen. Men vann og vannløselige stoffer (for eksempel ioner) kan rett og slett ikke passere gjennom membranen inn i noen celle. Dette betyr at de trenger spesielle hull. Men hvis du bare lager et hull i fettfilmen, vil den umiddelbart stramme seg tilbake. Hva å gjøre? En løsning ble funnet i naturen: det er nødvendig å lage spesielle proteintransportstrukturer og strekke dem gjennom membranen. Slik oppnås kanalene for passasje av fettuløselige stoffer - ionekanalene til cellemembranen.
Så, for å gi sin membran ytterligere egenskaper for permeabilitet for polare molekyler (ioner og vann), syntetiserer cellen spesielle proteiner i cytoplasmaet, som deretter integreres i membranen. De er av to typer: transportørproteiner (for eksempel transport ATPaser) og kanaldannende proteiner (kanaldannere). Disse proteinene er innebygd i det doble fettlaget i membranen og danner transportstrukturer i form av transportører eller i form av ionekanaler. Ulike vannløselige stoffer kan nå passere gjennom disse transportstrukturene, som ellers ikke kan passere gjennom den fettholdige membranfilmen.
Generelt kalles proteiner innebygd i membranen også integrert, nettopp fordi de så å si er inkludert i membranens sammensetning og trenger gjennom og gjennom den. Andre proteiner, ikke integrerte, danner, som det var, øyer som "flyter" på overflaten av membranen: enten langs dens ytre overflate eller langs dens indre. Tross alt vet alle at fett er et godt glidemiddel og det er lett å skli på det!
konklusjoner
1. Generelt er membranen tre-lags:
1) det ytre laget av protein "øyer",
2) fet tolags "sjø" (lipiddobbeltlag), dvs. dobbel lipidfilm
3) det indre laget av protein "øyer".
Men det er også et løst ytre lag - glykokalyxen, som er dannet av glykoproteiner som stikker ut av membranen. De er molekylære reseptorer som signalkontroller binder seg til.
2. Spesielle proteinstrukturer er bygget inn i membranen, som sikrer dens permeabilitet for ioner eller andre stoffer. Vi må ikke glemme at noen steder er fetthavet gjennomsyret av integrerte proteiner. Og det er integrerte proteiner som danner spesielle transportstrukturer cellemembran (se avsnitt 1_2 Membrantransportmekanismer). Gjennom dem kommer stoffer inn i cellen, og fjernes også fra cellen til utsiden.
3. Enzymproteiner kan være lokalisert på alle sider av membranen (ytre og indre), samt inne i membranen, noe som påvirker både tilstanden til selve membranen og levetiden til hele cellen.
Så cellemembranen er en aktiv variabel struktur som aktivt jobber i hele cellens interesser og forbinder den med omverdenen, og er ikke bare et "beskyttende skall". Dette er det viktigste å vite om cellemembranen.
I medisin brukes membranproteiner ofte som "mål" for legemidler. Reseptorer, ionekanaler, enzymer, transportsystemer fungerer som slike mål. Nylig, i tillegg til membranen, har gener som er skjult i cellekjernen også blitt mål for medikamenter.
Video:Introduksjon til cellemembranbiofysikk: struktur av membran 1 (Vladimirov Yu.A.)
Video:Historie, struktur og funksjoner til cellemembranen: Struktur av membraner 2 (Vladimirov Yu.A.)
© 2010-2018 Sazonov V.F., © 2010-2016 kineziolog.bodhy.
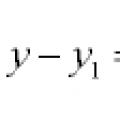 Ligninger av linjer og kurver i planet
Ligninger av linjer og kurver i planet Generalisert homogen likning Generaliserte homogene likninger av andre orden
Generalisert homogen likning Generaliserte homogene likninger av andre orden Takket være kommaet På grunn av det faktum at i deres
Takket være kommaet På grunn av det faktum at i deres