Strukturen til nevronet kort. Typer motoriske nevroner
Sist oppdatert: 10/10/2013
Populærvitenskapelig artikkel om nerveceller: strukturen, likhetene og forskjellene mellom nevroner med andre celler, prinsippet om overføring av elektriske og kjemiske impulser.
Nevron er en nervecelle som er hovedbyggesteinen for nervesystemet. Nevroner ligner andre celler på mange måter, men det er en viktig forskjell mellom et nevron og andre celler: Nevroner er spesialiserte på å overføre informasjon gjennom hele kroppen.
Disse høyt spesialiserte cellene er i stand til å overføre informasjon både kjemisk og elektrisk. Det finnes også flere forskjellige typer nevroner som utfører forskjellige funksjoner i menneskekroppen.
Sensoriske (sensitive) nevroner formidler informasjon fra sensoriske reseptorceller til hjernen. Motoriske (motoriske) nevroner overfører kommandoer fra hjernen til musklene. Interneuroner (interneuroner) er i stand til å kommunisere informasjon mellom ulike nevroner i kroppen.
Nevroner sammenlignet med andre celler i kroppen vår
Likheter med andre celler:
- Nevroner, som andre celler, har en kjerne som inneholder genetisk informasjon.
- Nevroner og andre celler er omgitt av en kappe som beskytter cellen.
- Cellelegemene til nevroner og andre celler inneholder organeller som støtter cellelivet: mitokondrier, Golgi-apparatet og cytoplasmaet.
Forskjellene som gjør nevronene unike
I motsetning til andre celler slutter nevroner å reprodusere kort tid etter fødselen. Derfor har noen deler av hjernen flere nevroner ved fødselen enn senere, fordi nevroner dør, men ikke beveger seg. Til tross for at nevroner ikke formerer seg, har forskere bevist at nye forbindelser mellom nevroner oppstår gjennom hele livet.
Nevroner har en membran som er designet for å sende informasjon til andre celler. er spesielle enheter som sender og mottar informasjon. Intercellulære forbindelser kalles synapser. Nevroner frigjør kjemiske forbindelser (nevrotransmittere eller nevrotransmittere) til synapser for å kommunisere med andre nevroner.
Strukturen til et nevron
Et nevron har bare tre hoveddeler: et akson, en cellekropp og dendritter. Imidlertid varierer alle nevroner litt i form, størrelse og egenskaper avhengig av nevronens rolle og funksjon. Noen nevroner har bare noen få grener av dendritter, mens andre forgrener seg sterkt for å motta en stor mengde informasjon. Noen nevroner har korte aksoner, mens andre kan være ganske lange. Det lengste aksonet i menneskekroppen strekker seg fra bunnen av ryggraden til stortåen, lengden er omtrent 0,91 meter (3 fot)!
Mer om strukturen til et nevron
handlingspotensial
Hvordan sender og mottar nevroner informasjon? For at nevroner skal kommunisere, må de overføre informasjon både i selve nevronet og fra nevronet til det neste nevronet. Både elektriske signaler og kjemiske sendere brukes til denne prosessen.
Dendritter mottar informasjon fra sensoriske reseptorer eller andre nevroner. Denne informasjonen sendes deretter til cellekroppen og til aksonet. Når denne informasjonen forlater aksonet, beveger den seg ned langs aksonet via et elektrisk signal kalt et aksjonspotensial.
Kommunikasjon mellom synapser
Så snart den elektriske impulsen når aksonet, må informasjonen mates til dendrittene til det tilstøtende nevronet gjennom synaptisk kløft.I noen tilfeller kan det elektriske signalet bygge bro over gapet mellom nevronene nesten umiddelbart og fortsette sin reise.
I andre tilfeller trenger nevrotransmittere å videresende informasjon fra ett nevron til det neste. Nevrotransmittere er kjemiske transmittere som frigjøres fra aksoner for å krysse den synaptiske kløften og nå reseptorene til andre nevroner. I en prosess som kalles «reuptake», fester nevrotransmittere seg til reseptoren og absorberes av nevronet for gjenbruk.
nevrotransmittere
Det er en integrert del av vår daglige funksjon. Det er ennå ikke kjent nøyaktig hvor mange nevrotransmittere som finnes, men forskere har allerede funnet mer enn hundre av disse kjemiske transmitterne.
Hvilken effekt har hver nevrotransmitter på kroppen? Hva skjer når en sykdom eller medisin møter disse kjemiske transmitterne? Her er noen av de viktigste nevrotransmitterne, deres kjente effekter og sykdommer forbundet med dem.
Nervesystemet kontrollerer, koordinerer og regulerer det koordinerte arbeidet til alle organsystemer, og opprettholder konstansen i sammensetningen av dets indre miljø (på grunn av dette fungerer menneskekroppen som en helhet). Med deltakelse av nervesystemet er organismen forbundet med det ytre miljøet.
nervevev
Nervesystemet dannes nervevev som består av nerveceller nevroner og liten satellittceller (gliaceller), som er omtrent 10 ganger flere enn nevroner.
Nevroner gi de grunnleggende funksjonene til nervesystemet: overføring, prosessering og lagring av informasjon. Nerveimpulser er elektriske og forplanter seg langs nevronenes prosesser.
satellittceller utføre ernæringsmessige, støttende og beskyttende funksjoner, fremme vekst og utvikling av nerveceller.
Strukturen til et nevron
Nevronet er den grunnleggende strukturelle og funksjonelle enheten i nervesystemet.
Den strukturelle og funksjonelle enheten i nervesystemet er nervecellen - nevron. Dens hovedegenskaper er eksitabilitet og ledningsevne.
Nevronet består av kropp Og prosesser.
Korte, sterkt forgrenede skudd - dendritter, gjennom dem kommer nerveimpulser til kroppen nervecelle. Det kan være en eller flere dendritter.
Hver nervecelle har en lang prosess - akson som impulser rettes mot fra cellekroppen. Lengden på aksonet kan nå flere titalls centimeter. Ved å kombinere til bunter dannes aksoner nerver.

De lange prosessene i nervecellen (aksonene) er dekket med myelinskjede. Akkumuleringer av slike prosesser, dekket myelin(hvit fettlignende substans), i sentralnervesystemet danner de den hvite substansen i hjernen og ryggmargen.
Korte prosesser (dendritter) og nevronlegemer har ikke myelinskjede, så de er grå i fargen. Deres ansamlinger danner den grå substansen i hjernen.
Nevroner kobles til hverandre på denne måten: aksonet til ett nevron blir med kroppen, dendrittene eller aksonet til et annet nevron. Kontaktpunktet mellom en nevron og en annen kalles synapse. Det er 1200–1800 synapser på kroppen til ett nevron.
Synapse - rommet mellom naboceller der den kjemiske overføringen av en nerveimpuls fra en nevron til en annen finner sted.

Hver Synapsen består av tre divisjoner:
- membran dannet av en nerveende presynaptisk membran);
- cellekroppsmembraner postsynaptisk membran);
- synaptisk spalte mellom disse membranene

Den presynaptiske delen av synapsen inneholder et biologisk aktivt stoff ( formidler), som sikrer overføring av en nerveimpuls fra en nevron til en annen. Under påvirkning av en nerveimpuls går nevrotransmitteren inn i den synaptiske kløften, virker på den postsynaptiske membranen og forårsaker eksitasjon av neste nevron i cellekroppen. Således, gjennom synapsen, overføres eksitasjon fra en nevron til en annen.
Spredningen av eksitasjon er forbundet med en slik egenskap av nervevevet som ledningsevne.
Typer nevroner
Nevroner varierer i form

Avhengig av funksjonen som utføres, skilles følgende typer nevroner ut:
- nevroner, overføre signaler fra sanseorganene til CNS(ryggmarg og hjerne) følsom. Kroppene til slike nevroner er plassert utenfor sentralnervesystemet, i nerveknutene (ganglia). Et ganglion er en samling av nervecellelegemer utenfor sentralnervesystemet.
- nevroner, overføre impulser fra ryggmargen og hjernen til muskler og indre organer kalt motor. De gir overføring av impulser fra sentralnervesystemet til arbeidsorganene.
- Kommunikasjon mellom sensoriske og motoriske nevroner gjennomført gjennom interkalære nevroner gjennom synaptiske kontakter i ryggmargen og hjernen. Interkalære nevroner ligger i CNS (dvs. kroppene og prosessene til disse nevronene strekker seg ikke utover hjernen).

Samlingen av nevroner i sentralnervesystemet kalles kjerne(hjernekjernen, ryggmargen).
Ryggmargen og hjernen er forbundet med alle organer nerver.
Nerver- mantelstrukturer, bestående av bunter av nervefibre, hovedsakelig dannet av aksoner av nevroner og neurogliaceller.

Nerver gir en forbindelse mellom sentralnervesystemet og organer, blodårer og huden.
Det utføres i henhold til tre hovedgrupper av tegn: morfologisk, funksjonell og biokjemisk.
1. Morfologisk klassifisering av nevroner(i henhold til funksjonene i strukturen). Etter antall skudd nevroner er delt inn i unipolar(med en gren), bipolar ( med to prosesser ) , pseudo-unipolar(falsk unipolar), multipolar(har tre eller flere prosesser). (Figur 8-2). Sistnevnte er mest i nervesystemet.
Ris. 8-2. Typer nerveceller.
1. Unipolar nevron.
2. Pseudo-unipolar nevron.
3. Bipolar nevron.
4. Multipolar nevron.
Nevrofibriller er synlige i cytoplasmaet til nevronene.
(Ifølge Yu. A. Afanasiev og andre).
Pseudo-unipolare nevroner kalles fordi, når aksonet og dendritten beveger seg bort fra kroppen, passer først tett til hverandre, og skaper inntrykk av en prosess, og først deretter divergerer på en T-formet måte (disse inkluderer alle reseptorneuroner i spinale og kraniale ganglier). Unipolare nevroner finnes bare i embryogenese. Bipolare nevroner er bipolare celler i netthinnen, spiral og vestibulære ganglier. Etter form opptil 80 varianter av nevroner er beskrevet: stjerneformet, pyramideformet, pæreformet, fusiform, arachnid, etc.
2. Funksjonell(avhengig av utført funksjon og plass i refleksbuen): reseptor, effektor, interkalær og sekretorisk. Reseptor(sensitive, afferente) nevroner, ved hjelp av dendritter, oppfatter effekten av det ytre eller indre miljøet, genererer en nerveimpuls og overfører den til andre typer nevroner. De finnes bare i spinalgangliene og sensoriske kjerner i kranienervene. Effektor(efferente) nevroner overfører eksitasjon til arbeidsorganene (muskler eller kjertler). De er lokalisert i de fremre hornene i ryggmargen og autonome nerveganglier. Innsetting(assosiative) nevroner er lokalisert mellom reseptor- og effektorneuroner; etter deres antall mest, spesielt i sentralnervesystemet. sekretoriske nevroner(nevrosekretoriske celler) spesialiserte nevroner som fungerer som endokrine celler. De syntetiserer og skiller ut nevrohormoner i blodet og er lokalisert i den hypotalamiske regionen av hjernen. De regulerer aktiviteten til hypofysen, og gjennom den mange perifere endokrine kjertler.
3. Mekler(i henhold til den kjemiske naturen til den utskilte mediatoren):
Kolinerge nevroner (mediator acetylkolin);
Aminergiske (mediatorer - biogene aminer, som noradrenalin, serotonin, histamin);
GABAergic (mediator - gamma-aminosmørsyre);
Aminosyre-ergisk (mediatorer - aminosyrer som glutamin, glycin, aspartat);
Peptidergisk (mediatorer - peptider, som opioide peptider, substans P, kolecystokinin, etc.);
Purinergisk (mediatorer - purinnukleotider, som adenin), etc.
Den indre strukturen til nevroner
Kjerne nevroner er vanligvis store, avrundede, med fint spredt kromatin, 1-3 store nukleoler. Dette gjenspeiler den høye intensiteten til transkripsjonsprosesser i nevronkjernen.
Celleveggen Et nevron er i stand til å generere og lede elektriske impulser. Dette oppnås ved å endre den lokale permeabiliteten til ionekanalene for Na + og K +, endre det elektriske potensialet og raskt flytte det langs cytolemmaet (depolarisasjonsbølge, nerveimpuls).
I cytoplasmaet til nevroner er alle organeller for generelle formål godt utviklet. Mitokondrier er mange og gir høye energibehov til nevronet, assosiert med en betydelig aktivitet av syntetiske prosesser, ledning av nerveimpulser og drift av ionepumper. De er preget av rask slitasje (Figur 8-3). Golgi kompleks veldig godt utviklet. Det er ingen tilfeldighet at denne organellen først ble beskrevet og demonstrert i løpet av cytologi i nevroner. Med lysmikroskopi oppdages det i form av ringer, filamenter, korn som ligger rundt kjernen (diktyosomer). En rekke lysosomer gi konstant intensiv ødeleggelse av bærbare komponenter i nevroncytoplasma (autofagi).
R  er. 8-3. Ultrastrukturell organisering av nevronkroppen.
er. 8-3. Ultrastrukturell organisering av nevronkroppen.
D. Dendritter. A. Axon.
1. Nucleus (kjerne er vist med en pil).
2. Mitokondrier.
3. Golgi-kompleks.
4. Kromatofilt stoff (områder med granulært cytoplasmatisk retikulum).
5. Lysosomer.
6. Axon-bakke.
7. Nevrotubuli, nevrotråder.
(Ifølge V. L. Bykov).
For normal funksjon og fornyelse av nevronstrukturer må proteinsynteseapparatet være godt utviklet i disse (fig. 8-3). Granulært cytoplasmatisk retikulum i cytoplasma til nevroner danner klynger som er godt farget med grunnleggende fargestoffer og er synlige under lysmikroskopi i form av klumper kromatofilt stoff(basofilt, eller tigerstoff, Nissl-stoff). Begrepet "Nissl-stoff" er bevart til ære for vitenskapsmannen Franz Nissl, som først beskrev det. Klumper av kromatofilt stoff er lokalisert i perikarya til nevroner og dendritter, men finnes aldri i aksoner, hvor det proteinsynteserende apparatet er dårlig utviklet (fig. 8-3). Ved langvarig irritasjon eller skade på en nevron brytes disse ansamlingene av det granulære cytoplasmatiske retikulum opp i separate elementer, som på det lysoptiske nivået manifesteres ved forsvinningen av Nissl-stoffet ( kromatolyse, tigrolyse).
cytoskjelett nevroner er godt utviklet, danner et tredimensjonalt nettverk, representert av nevrofilamenter (6-10 nm tykke) og nevrotubuli (20-30 nm i diameter). Nevrofilamenter og neurotubuli er forbundet med hverandre ved hjelp av tverrgående broer, når de er festet, kleber de sammen til 0,5–0,3 μm tykke bunter, som er farget med sølvsalter. På lysoptisk nivå beskrives de under navnet nevrofibriller. De danner et nettverk i perikaryonene til nevrocytter, og i prosessene ligger de parallelt (fig. 8-2). Cytoskjelettet opprettholder celleformen, og gir også en transportfunksjon - det er involvert i transport av stoffer fra perikaryon til prosessene (aksonal transport).
Inkluderinger i cytoplasmaet til nevronen er representert av lipiddråper, granulat lipofuscin- "aldringspigment" - gul-brun farge av lipoprotein natur. De er gjenværende kropper (telolisosomer) med produkter av ufordøyde nevronstrukturer. Tilsynelatende kan lipofuscin også akkumuleres i ung alder, med intensiv funksjon og skade på nevroner. I tillegg er det pigmentinneslutninger i cytoplasmaet til nevronene til substantia nigra og den blå flekken i hjernestammen. melanin. Mange nevroner i hjernen inneholder inneslutninger glykogen.
Nevroner er ikke i stand til å dele seg, og med alderen avtar antallet gradvis på grunn av naturlig død. Ved degenerative sykdommer (Alzheimers sykdom, Huntingtons sykdom, parkinsonisme) øker intensiteten av apoptose og antallet nevroner i visse deler av nervesystemet synker kraftig.
Nevroner er delt inn i reseptor, effektor og intercalary.
Kompleksiteten og mangfoldet av funksjonene til nervesystemet bestemmes av samspillet mellom nevroner. Denne interaksjonen er et sett med forskjellige signaler som overføres mellom nevroner eller muskler og kjertler. Signaler sendes ut og forplantes av ioner. Ioner genererer en elektrisk ladning (aksjonspotensial) som beveger seg gjennom kroppen til nevronet.
Av stor betydning for vitenskapen var oppfinnelsen av Golgi-metoden i 1873, som gjorde det mulig å farge enkelte nevroner. Begrepet "nevron" (tysk nevron) for å referere til nerveceller ble introdusert av G. W. Waldeyer i 1891.
Encyklopedisk YouTube
1 / 5
✪ Interneuronale kjemiske synapser
✪ Nevroner
✪ Mystisk hjerne. Andre del. Virkeligheten er prisgitt nevroner.
✪ Hvordan sport stimulerer veksten av nevroner i hjernen?
✪ Strukturen til et nevron
Undertekster
Nå vet vi hvordan en nerveimpuls overføres. La alt begynne med eksitasjon av dendritter, for eksempel denne utveksten av kroppen til en nevron. Eksitasjon betyr å åpne ionekanalene i membranen. Gjennom kanalene kommer ioner inn i cellen eller kommer ut av cellen. Dette kan føre til hemming, men i vårt tilfelle virker ionene elektrotonisk. De endrer det elektriske potensialet på membranen, og denne endringen i regionen til aksonbakken kan være nok til å åpne natriumionekanaler. Natriumioner kommer inn i cellen, ladningen blir positiv. Dette åpner kaliumkanaler, men denne positive ladningen aktiverer neste natriumpumpe. Natriumioner kommer inn i cellen igjen, og dermed sendes signalet videre. Spørsmålet er, hva skjer i krysset mellom nevroner? Vi var enige om at det hele begynte med eksitasjonen av dendrittene. Som regel er kilden til eksitasjon en annen nevron. Dette aksonet vil også overføre eksitasjon til en annen celle. Det kan være en muskelcelle eller en annen nervecelle. Hvordan? Her er aksonterminalen. Og her kan det være en dendritt av et annet nevron. Dette er et annet nevron med sitt eget akson. Dendritten hans er spent. Hvordan skjer dette? Hvordan går impulsen fra aksonet til en nevron til dendritten til en annen? Overføring fra akson til akson, fra dendritt til dendritt, eller fra akson til cellekropp er mulig, men som oftest overføres impulsen fra akson til nevrondendritter. La oss ta en nærmere titt. Vi er interessert i hva som skjer i den delen av bildet, som jeg vil sirkle rundt i en boks. Aksonterminalen og dendritten til neste nevron faller inn i rammen. Så her er aksonterminalen. Det ser omtrent slik ut under forstørrelse. Dette er aksonterminalen. Her er dens indre innhold, og ved siden av den er dendritten til et nabonevron. Dette er hvordan dendritten til et nabonevron ser ut under forstørrelse. Her er hva som er inne i det første nevronet. Aksjonspotensialet beveger seg over membranen. Til slutt, et sted på den aksonterminale membranen, blir det intracellulære potensialet positivt nok til å åpne natriumkanalen. Før handlingspotensialets ankomst er det stengt. Her er kanalen. Det slipper natriumioner inn i cellen. Det er her det hele starter. Kaliumioner forlater cellen, men så lenge den positive ladningen forblir, kan den åpne andre kanaler, ikke bare natrium. Det er kalsiumkanaler i enden av aksonet. Jeg skal male rosa. Her er kalsiumkanalen. Den er vanligvis lukket og lar ikke toverdige kalsiumioner passere gjennom. Dette er en spenningsstyrt kanal. Som natriumkanaler åpnes det når det intracellulære potensialet blir positivt nok til å slippe kalsiumioner inn i cellen. Toverdige kalsiumioner kommer inn i cellen. Og dette øyeblikket er fantastisk. Dette er kationer. Det er en positiv ladning inne i cellen på grunn av natriumioner. Hvordan kommer kalsium dit? Kalsiumkonsentrasjonen skapes ved hjelp av en ionepumpe. Jeg har allerede snakket om natrium-kalium-pumpen, det er en lignende pumpe for kalsiumioner. Dette er proteinmolekyler innebygd i membranen. Membranen er fosfolipid. Den består av to lag fosfolipider. Som dette. Det er mer som en ekte cellemembran. Her er også membranen to-lags. Dette er åpenbart, men jeg skal avklare i tilfelle. Også her er det kalsiumpumper som fungerer på samme måte som natrium-kalium-pumper. Pumpen mottar et ATP-molekyl og et kalsiumion, spalter fosfatgruppen fra ATP og endrer konformasjonen, og skyver kalsium ut. Pumpen er konstruert på en slik måte at den pumper kalsium ut av cellen. Det forbruker energien til ATP og gir en høy konsentrasjon av kalsiumioner utenfor cellen. I hvile er konsentrasjonen av kalsium utenfor mye høyere. Når et aksjonspotensial mottas, åpnes kalsiumkanaler, og kalsiumioner fra utsiden kommer inn i aksonterminalen. Der binder kalsiumioner seg til proteiner. Og la oss nå se hva som faktisk skjer på dette stedet. Jeg har allerede nevnt ordet "synapse". Kontaktpunktet mellom aksonet og dendritten er synapsen. Og det er en synapse. Det kan betraktes som et sted hvor nevroner kobles til hverandre. Dette nevronet kalles presynaptisk. Jeg skriver det ned. Du må kjenne vilkårene. presynaptisk. Og dette er postsynaptisk. Postsynaptisk. Og rommet mellom disse aksonene og dendritten kalles synaptisk spalte. synaptisk spalte. Det er et veldig, veldig smalt gap. Nå snakker vi om kjemiske synapser. Vanligvis, når folk snakker om synapser, mener de kjemiske. Det finnes også elektriske, men vi skal ikke snakke om dem ennå. Tenk på en konvensjonell kjemisk synapse. I en kjemisk synapse er denne avstanden bare 20 nanometer. Cellen har i gjennomsnitt en bredde på 10 til 100 mikron. En mikron er 10 til minus sjette potens av meter. Det er 20 ganger 10 til minus niende potens. Dette er et veldig smalt gap, hvis vi sammenligner størrelsen med størrelsen på cellen. Det er vesikler inne i aksonterminalen til det presynaptiske nevronet. Disse vesiklene er koblet til cellemembranen fra innsiden. Her er boblene. De har sin egen lipid-dobbeltlagsmembran. Bobler er beholdere. Det er mange av dem i denne delen av cellen. De inneholder molekyler som kalles nevrotransmittere. Jeg skal vise dem i grønt. Nevrotransmittere inne i vesiklene. Jeg tror dette ordet er kjent for deg. Mange medisiner for depresjon og andre psykiske problemer virker spesifikt på nevrotransmittere. Nevrotransmittere Nevrotransmittere i vesiklene. Når spenningsstyrte kalsiumkanaler åpner seg, kommer kalsiumioner inn i cellen og binder seg til proteiner som holder vesiklene. Vesiklene holdes på den presynaptiske membranen, det vil si denne delen av membranen. De holdes tilbake av proteiner fra SNARE-gruppen. Proteiner fra denne familien er ansvarlige for membranfusjon. Det er hva disse proteinene er. Kalsiumioner binder seg til disse proteinene og endrer konformasjonen deres slik at de trekker vesiklene så nærme cellemembranen at vesikkelmembranene smelter sammen med den. La oss se på denne prosessen mer detaljert. Etter at kalsium binder seg til SNARE-familiens proteiner på cellemembranen, trekker de vesiklene nærmere den presynaptiske membranen. Her er boblen. Slik går den presynaptiske membranen. Mellom seg er de forbundet med proteiner fra SNARE-familien, som tiltrakk boblen til membranen og er lokalisert her. Resultatet ble membranfusjon. Dette fører til at nevrotransmittere fra vesiklene kommer inn i den synaptiske kløften. Dette er hvordan nevrotransmittere frigjøres til synaptisk spalte. Denne prosessen kalles eksocytose. Nevrotransmittere forlater cytoplasmaet til det presynaptiske nevronet. Du har sikkert hørt navnene deres: serotonin, dopamin, adrenalin, som både er et hormon og en nevrotransmitter. Noradrenalin er både et hormon og en nevrotransmitter. Alle av dem er sannsynligvis kjent for deg. De går inn i den synaptiske kløften og binder seg til overflatestrukturene til membranen til det postsynaptiske nevronet. postsynaptisk nevron. La oss si at de binder seg her, her og her til spesifikke proteiner på overflaten av membranen, som et resultat av at ionekanaler aktiveres. Eksitasjon skjer i denne dendritten. La oss si at bindingen av nevrotransmittere til membranen fører til åpning av natriumkanaler. Membrannatriumkanaler åpner seg. De er senderavhengige. På grunn av åpningen av natriumkanaler kommer natriumioner inn i cellen, og alt gjentas igjen. Et overskudd av positive ioner vises i cellen, dette elektrotoniske potensialet sprer seg til området av aksonbakken, deretter til neste nevron, og stimulerer det. Slik skjer det. Det er mulig ellers. Anta at i stedet for å åpne natriumkanaler, vil kaliumionkanaler åpne seg. I dette tilfellet vil kaliumioner gå ut langs konsentrasjonsgradienten. Kaliumioner forlater cytoplasmaet. Jeg vil vise dem som trekanter. På grunn av tap av positivt ladede ioner avtar det intracellulære positive potensialet, som et resultat av at genereringen av et aksjonspotensial i cellen er vanskelig. Jeg håper dette er forståelig. Vi startet med spenning. Et aksjonspotensial genereres, kalsium kommer inn, innholdet i vesiklene kommer inn i synaptisk spalte, natriumkanaler åpnes og nevronet stimuleres. Og hvis du åpner kaliumkanaler, vil nevronet bremse ned. Synapser er veldig, veldig, veldig mange. Det er billioner av dem. Cerebral cortex alene antas å inneholde mellom 100 og 500 billioner synapser. Og det er bare bjeffingen! Hvert nevron er i stand til å danne mange synapser. På dette bildet kan synapser være her, her og her. Hundre og tusenvis av synapser på hver nervecelle. Med en nevron, en annen, tredje, fjerde. Et stort antall forbindelser ... enormt. Nå ser du hvor komplekst alt som har med menneskesinnet å gjøre er ordnet. Håper du finner det nyttig. Undertekster fra Amara.org-fellesskapet
Strukturen til nevroner
cellekropp
Kroppen til en nervecelle består av protoplasma (cytoplasma og kjerne), avgrenset på utsiden av en membran av lipid-dobbeltlag. Lipider er sammensatt av hydrofile hoder og hydrofobe haler. Lipider er ordnet i hydrofobe haler til hverandre, og danner et hydrofobt lag. Dette laget lar bare fettløselige stoffer (f.eks. oksygen og karbondioksid) passere gjennom. Det er proteiner på membranen: i form av kuler på overflaten, på hvilke utvekster av polysakkarider (glycocalix) kan observeres, på grunn av hvilke cellen oppfatter ytre irritasjon, og integrerte proteiner som trenger gjennom membranen, der det er ioner kanaler.
Nevronet består av en kropp med en diameter på 3 til 130 mikron. Kroppen inneholder en kjerne (med et stort antall atomporer) og organeller (inkludert en høyt utviklet grov ER med aktive ribosomer, Golgi-apparatet), samt prosesser. Det er to typer prosesser: dendritter og aksoner. Nevronet har et utviklet cytoskjelett som trenger inn i prosessene. Cytoskjelettet opprettholder cellens form, trådene fungerer som "skinner" for transport av organeller og stoffer pakket i membranvesikler (for eksempel nevrotransmittere). Cytoskjelettet til et nevron består av fibriller med forskjellige diametre: Mikrotubuli (D = 20-30 nm) - består av proteinet tubulin og strekker seg fra nevronet langs aksonet, opp til nerveendene. Nevrofilamenter (D = 10 nm) - sammen med mikrotubuli gir intracellulær transport av stoffer. Mikrofilamenter (D = 5 nm) - består av aktin- og myosinproteiner, de er spesielt uttalt i voksende nerveprosesser og i neuroglia. ( neuroglia, eller ganske enkelt glia (fra andre greske νεῦρον - fiber, nerve + γλία - lim), - et sett med hjelpeceller i nervevevet. Det utgjør omtrent 40 % av volumet av CNS. Antall gliaceller er i gjennomsnitt 10-50 ganger større enn antallet nevroner.)
I kroppen til nevronet avsløres et utviklet syntetisk apparat, den granulære ER av nevronet farges basofilt og er kjent som "tigroiden". Tigroiden trenger inn i de innledende delene av dendrittene, men er lokalisert i en merkbar avstand fra begynnelsen av aksonet, som fungerer som et histologisk tegn på aksonet. Nevroner er forskjellige i form, antall prosesser og funksjoner. Avhengig av funksjonen skilles sensitiv, effektor (motorisk, sekretorisk) og interkalær. Sensoriske nevroner oppfatter stimuli, konverterer dem til nerveimpulser og overfører dem til hjernen. Effektor (av lat. effectus - handling) - de utvikler og sender kommandoer til arbeidsorganene. Intercalary - utføre en forbindelse mellom sensoriske og motoriske nevroner, delta i informasjonsbehandling og kommandogenerering.
Det skilles mellom anterograd (bort fra kroppen) og retrograd (mot kroppen) aksontransport.
Dendritter og akson
Aksjonspotensialskaping og ledningsmekanisme
I 1937 bestemte John Zachary Jr. at det gigantiske aksonet for blekksprut kunne brukes til å studere de elektriske egenskapene til aksoner. Blekksprutaksoner ble valgt fordi de er mye større enn menneskelige. Hvis du setter inn en elektrode inne i aksonet, kan du måle membranpotensialet.
Aksonmembranen inneholder spenningsstyrte ionekanaler. De lar aksonet generere og lede elektriske signaler gjennom kroppen kalt aksjonspotensialer. Disse signalene genereres og forplantes av elektrisk ladede natrium (Na+), kalium (K+), klor (Cl-), kalsium (Ca2+) ioner.
Trykk, strekk, kjemiske faktorer eller en endring i membranpotensial kan aktivere en nevron. Dette skjer på grunn av åpningen av ionekanaler som lar ioner krysse cellemembranen og følgelig endre membranpotensialet.
Tynne aksoner bruker mindre energi og metabolske stoffer for å utføre et aksjonspotensial, men tykke aksoner gjør at det kan ledes raskere.
For å utføre aksjonspotensialer raskere og mindre energikrevende, kan nevroner bruke spesielle gliaceller til å belegge aksoner kalt oligodendrocytter i CNS eller Schwann-celler i det perifere nervesystemet. Disse cellene dekker ikke aksonene fullstendig, og etterlater hull på aksonene åpne for ekstracellulært materiale. I disse hullene er det en økt tetthet av ionekanaler. De kalles intercepts Ranvier. Gjennom dem går aksjonspotensialet gjennom det elektriske feltet mellom hullene.
Klassifisering
Strukturell klassifisering
Basert på antall og arrangement av dendritter og aksoner, er nevroner delt inn i ikke-aksonale, unipolare nevroner, pseudo-unipolare nevroner, bipolare nevroner og multipolare (mange dendritiske stammer, vanligvis efferente) nevroner.
Aksonløse nevroner- små celler, gruppert nær ryggmargen i de intervertebrale gangliene, som ikke har anatomiske tegn på separasjon av prosesser i dendritter og aksoner. Alle prosesser i en celle er veldig like. Det funksjonelle formålet med aksonløse nevroner er dårlig forstått.
Unipolare nevroner- nevroner med én prosess, er til stede, for eksempel, i den sensoriske kjernen til trigeminusnerven i midthjernen. Mange morfologer mener at unipolare nevroner ikke finnes i menneskekroppen og høyere virveldyr.
Multipolare nevroner- Nevroner med ett akson og flere dendritter. Denne typen nerveceller dominerer i sentralnervesystemet.
Pseudo-unipolare nevroner- er unike i sitt slag. En prosess går fra kroppen, som umiddelbart deler seg i en T-form. Hele denne enkeltkanalen er dekket med en myelinskjede og representerer strukturelt et akson, men langs en av grenene går eksitasjonen ikke fra, men til nevronkroppen. Strukturelt sett er dendritter forgreninger på slutten av denne (perifere) prosessen. Triggersonen er begynnelsen på denne forgreningen (det vil si at den er plassert utenfor cellekroppen). Slike nevroner finnes i spinalgangliene.
Funksjonell klassifisering
Afferente nevroner(sensitiv, sensorisk, reseptor eller centripetal). Nevroner av denne typen inkluderer primære celler i sanseorganene og pseudo-unipolare celler, der dendritter har frie ender.
Efferente nevroner(effektor, motor, motor eller sentrifugal). Nevroner av denne typen inkluderer endelige nevroner - ultimatum og nest siste - ikke ultimatum.
Assosiative nevroner(intercalary eller interneurons) - en gruppe nevroner kommuniserer mellom efferente og afferente, de er delt inn i intrusion, commissural og projection.
sekretoriske nevroner- nevroner som skiller ut høyaktive stoffer (nevrohormoner). De har et velutviklet Golgi-kompleks, aksonet ender i axovasale synapser.
Morfologisk klassifisering
Den morfologiske strukturen til nevroner er mangfoldig. Ved klassifisering av nevroner brukes flere prinsipper:
- ta hensyn til størrelsen og formen på kroppen til nevronet;
- antallet og arten av forgreningsprosesser;
- aksonlengde og tilstedeværelsen av spesialiserte slirer.
I henhold til cellens form kan nevroner være sfæriske, granulære, stjerneformede, pyramideformede, pæreformede, spindelformede, uregelmessige osv. Størrelsen på nevronkroppen varierer fra 5 mikron i små granulære celler til 120-150 mikron i gigantiske pyramidale nevroner.
I henhold til antall prosesser skilles følgende morfologiske typer nevroner ut:
- unipolare (med én prosess) nevrocytter, tilstede for eksempel i den sensoriske kjernen til trigeminusnerven i midthjernen;
- pseudo-unipolare celler gruppert nær ryggmargen i de intervertebrale gangliene;
- bipolare nevroner (har ett akson og en dendritt) lokalisert i spesialiserte sanseorganer - netthinnen, olfaktorisk epitel og pære, auditive og vestibulære ganglier;
- multipolare nevroner (har ett akson og flere dendritter), dominerende i CNS.
Utvikling og vekst av en nevron
Spørsmålet om nevronal deling er for tiden diskutabelt. Ifølge en versjon utvikler nevronet seg fra en liten forløpercelle, som slutter å dele seg selv før den frigjør prosessene sine. Aksonet begynner å vokse først, og dendrittene dannes senere. En fortykkelse vises på slutten av utviklingsprosessen til nervecellen, som baner vei gjennom det omkringliggende vevet. Denne fortykkelsen kalles vekstkjeglen til nervecellen. Den består av en flatet del av nervecellens prosess med mange tynne ryggrader. Mikrospinulene er 0,1 til 0,2 µm tykke og kan være opptil 50 µm lange; det brede og flate området til vekstkjeglen er omtrent 5 µm bredt og langt, selv om formen kan variere. Mellomrommene mellom vekstkjeglens mikrorygger er dekket med en foldet membran. Mikrorygger er i konstant bevegelse - noen trekkes inn i vekstkjeglen, andre forlenges, avviker i forskjellige retninger, berører underlaget og kan feste seg til det.
Vekstkjeglen er fylt med små, noen ganger sammenkoblede, uregelmessig formede membranøse vesikler. Under de foldede områdene av membranen og i ryggradene er en tett masse av sammenfiltrede aktinfilamenter. Vekstkjeglen inneholder også mitokondrier, mikrotubuli og neurofilamenter som ligner på de som finnes i kroppen til en nevron.
Mikrotubuli og neurofilamenter forlenges hovedsakelig ved tilsetning av nylig syntetiserte underenheter i bunnen av nevronprosessen. De beveger seg med en hastighet på omtrent en millimeter per dag, som tilsvarer hastigheten til langsom aksontransport i et modent nevron. Siden den gjennomsnittlige fremdriftshastigheten til vekstkjeglen er tilnærmet den samme, er det mulig at verken montering eller ødeleggelse av mikrotubuli og nevrofilamenter skjer i den andre enden under veksten av nevronprosessen. Nytt membranmateriale legges til på slutten. Vekstkjeglen er et område med rask eksocytose og endocytose, noe som fremgår av de mange vesiklene som finnes her. Små membranvesikler transporteres langs nevronprosessen fra cellekroppen til vekstkjeglen med en strøm av rask aksontransport. Membranmateriale syntetisert i kroppen til nevronet overføres til vekstkjeglen i form av vesikler og inngår her i plasmamembranen ved eksocytose, og forlenger dermed prosessen til nervecellen.
Veksten av aksoner og dendritter innledes vanligvis av en fase med nevronal migrasjon, når umodne nevroner slår seg ned og finner et permanent sted for seg selv.
Egenskaper og funksjoner til nevroner
Egenskaper:
- Tilstedeværelsen av en transmembranpotensialforskjell(opptil 90 mV), er den ytre overflaten elektropositiv i forhold til den indre overflaten.
- Svært høy følsomhet til visse kjemikalier og elektrisk strøm.
- Evnen til å nevrosekretere, det vil si til syntese og frigjøring av spesielle stoffer (nevrotransmittere) til miljøet eller den synaptiske kløften.
- Høyt strømforbruk, et høyt nivå av energiprosesser, som nødvendiggjør en konstant tilførsel av hovedkildene til energi - glukose og oksygen, nødvendig for oksidasjon.
Funksjoner:
- mottaksfunksjon(synapser er kontaktpunkter, vi mottar informasjon i form av en impuls fra reseptorer og nevroner).
- Integrativ funksjon(informasjonsbehandling, som et resultat dannes et signal ved utgangen av nevronet, som bærer informasjonen om alle de summerte signalene).
- Konduktørfunksjon(fra nevronet langs aksonet er det informasjon i form av en elektrisk strøm til synapsen).
- Overføringsfunksjon(en nerveimpuls, etter å ha nådd slutten av aksonet, som allerede er en del av strukturen til synapsen, forårsaker frigjøring av en mediator - en direkte sender av eksitasjon til et annet nevron eller utøvende organ).
Nevroner er svært komplekse strukturer. Cellestørrelser er ekstremt forskjellige (fra 4-6 mikron til 130 mikron). Formen på en nevron er også veldig variabel, men alle nerveceller har prosesser (en eller flere) som strekker seg fra kroppen. Mennesker har over en billion (10) nerveceller.
På strengt definerte stadier av ontogenese programmeres den massedød av nevroner sentral- og perifert nervesystem. I løpet av 1 leveår dør omtrent 10 millioner nevroner, og i løpet av livet mister hjernen omtrent 0,1 % av alle nevroner. Døden bestemmes av en rekke faktorer:
de mest aktivt deltakende i intercellulære interaksjoner av nevronet overlever (de vokser raskere, har flere prosesser, flere kontakter med målceller).
det er gener som er ansvarlige for utgangen mellom liv og død.
avbrudd i blodtilførselen.
Etter antall skudd nevroner er delt inn i:
unipolar - ensidig,
bipolar - tosidig,
multipolar - multibehandlet.
Blant unipolare nevroner skilles ekte unipolare ut,
liggende i netthinnen i øyet, og falske unipolare lokalisert i spinalknutene. Falske unipolare celler i utviklingsprosessen var bipolare celler, men så ble en del av cellen trukket inn i en lang prosess, som ofte gjør flere svinger rundt kroppen og deretter forgrener seg i en T-form.
Prosessene til nerveceller er forskjellige i struktur, hver nervecelle har et akson eller neuritt, som kommer fra cellekroppen i form av en tråd som har samme tykkelse langs hele lengden. Aksoner reiser ofte lange avstander. I løpet av nevrittet avgår tynne grener - kollateraler. Aksonet, som overfører prosessen og impulsen i den, går fra cellen til periferien. Aksonet ender med en effektor eller motor som ender i muskel- eller kjertelvev. Lengden på aksonet kan være mer enn 100 cm Det er ikke noe endoplasmatisk retikulum og frie ribosomer i aksonet, så alle proteiner skilles ut i kroppen og transporteres deretter langs aksonet.
Andre prosesser starter fra cellekroppen med bred base og kraftig forgrening. De kalles dendrittiske prosesser eller dendritter og er mottakelige prosesser der impulsen forplanter seg mot cellekroppen. Dendritter ender i sensitive nerveender eller reseptorer som spesifikt oppfatter irritasjoner.
Ekte unipolare nevroner har bare ett akson, og oppfatningen av impulser utføres av hele overflaten av cellen. Det eneste eksemplet på unipotente celler hos mennesker er retinale amokrine celler.
Bipolare nevroner ligger i netthinnen i øyet og har et akson og en forgreningsprosess - en dendritt.
Multipolare nevroner er utbredt og ligger i ryggmargen og hjernen, autonome ganglioner, etc. Disse cellene har ett akson og mange forgrenende dendritter.
Avhengig av plasseringen er nevroner delt inn i sentrale, liggende i hjernen og ryggmargen, og perifere - disse er nevroner av de autonome gangliene, organnerveplexuser og spinalnoder.
Nerveceller samhandler tett med blodårene. Det er 3 interaksjonsalternativer:
Nerveceller i kroppen ligger i form av kjeder, d.v.s. en celle kontakter en annen og overfører sin impuls til den. Slike cellekjeder kalles refleksbuer. Avhengig av posisjonen til nevroner i refleksbuen, har de en annen funksjon. Av funksjon kan nevroner være følsomme, motoriske, assosiative og interkalære. Mellom seg selv eller med målorganet samhandler nerveceller ved hjelp av kjemikalier - nevrotransmittere.
Aktiviteten til et nevron kan induseres av en impuls fra et annet nevron eller være spontant. I dette tilfellet spiller nevronet rollen som en pacemaker (pacemaker). Slike nevroner finnes i en rekke sentre, inkludert det respiratoriske.
Den første sensoriske nevronen i refleksbuen er sansecellen. Irritasjon oppfattes av reseptoren - en følsom slutt, impulsen når cellekroppen langs dendritten, og overføres deretter langs aksonet til en annen nevron. Kommandoen om å handle på arbeidsorganet overføres av et motor- eller effektorneuron. Et effektornevron kan motta en impuls direkte fra en sensitiv celle, da vil refleksbuen bestå av to nevroner.
I mer komplekse refleksbuer er det en mellomledd - en interkalær nevron. Han oppfatter en impuls fra en sensitiv celle og overfører den til en motorisk celle.
Noen ganger kombineres flere celler med samme funksjon (sensorisk eller motorisk) av ett nevron, som konsentrerer impulser fra flere celler i seg selv - dette er assosiative nevroner. Disse nevronene overfører impulsen videre til intercalary- eller effektorneuronene.
I kroppen til en nevron inneholder de fleste nerveceller én kjerne. Flerkjernede nerveceller er karakteristiske for noen perifere ganglier i det autonome nervesystemet. På histologiske preparater ser kjernen til en nervecelle ut som en lys boble med en tydelig skillelig kjerne og noen få klumper av kromatin. Elektronmikroskopi avslører de samme submikroskopiske komponentene som i kjernene til andre celler. Den kjernefysiske konvolutten har mange porer. Kromatin er spredt. En slik struktur av kjernen er karakteristisk for metabolsk aktive kjernefysiske apparater.
Kjernemembranen i prosessen med embryogenese danner dype folder som strekker seg inn i karyoplasmaen. Ved fødselen blir foldingen mye mindre. Hos en nyfødt er det allerede en overvekt av volumet av cytoplasma over kjernen, siden i løpet av embryogeneseperioden er disse forholdene reversert.
Cytoplasmaet til en nervecelle kalles neuroplasma. Den inneholder organeller og inneslutninger.
Golgi-apparatet ble først oppdaget i nerveceller. Det ser ut som en kompleks kurv som omgir kjernen fra alle kanter. Dette er en slags diffus type av Golgi-apparatet. Under elektronmikroskopi består den av store vakuoler, små vesikler og pakker med doble membraner som danner et anastomoserende nettverk rundt nervecellens kjerneapparat. Imidlertid er Golgi-apparatet som oftest plassert mellom kjernen og stedet der aksonet kommer fra - aksonbakken. Golgi-apparatet er stedet for generering av handlingspotensial.
Mitokondrier ser ut som veldig korte pinner. De finnes i cellekroppen og i alle prosesser. I de terminale grenene av nerveprosessene, dvs. deres akkumulering observeres i nerveendene. Ultrastrukturen til mitokondrier er typisk, men deres indre membran danner ikke et stort antall cristae. De er svært følsomme for hypoksi. Mitokondrier ble først beskrevet i muskelceller av Kelliker for over 100 år siden. I noen nevroner er det anastomoser mellom mitokondrielle cristae. Antall cristae og deres totale overflate er direkte relatert til intensiteten av deres respirasjon. Uvanlig er akkumulering av mitokondrier i nerveender. I prosessene er de orientert med sin lengdeakse langs prosessene.
Cellesenteret i nerveceller består av 2 sentrioler omgitt av en lys sfære, og kommer mye bedre til uttrykk i unge nevroner. I modne nevroner finnes cellesenteret med vanskeligheter, og i den voksne organismen gjennomgår sentrosomet degenerative endringer.
Ved farging av nerveceller med toluoidblått, finnes det klumper av forskjellige størrelser i cytoplasmaet - basofilt stoff, eller Nissls stoff. Dette er et veldig ustabilt stoff: med generell tretthet som følge av langvarig arbeid eller nervøs spenning, forsvinner klumper av Nissl-stoff. Histokjemisk ble det funnet RNA og glykogen i klumpene. Elektronmikroskopiske studier har vist at Nissl-klumper er et endoplasmatisk retikulum. Det er mange ribosomer på membranene i det endoplasmatiske retikulum. Det er også mange frie ribosomer i nevroplasmaet, og danner rosettlignende klynger. Det utviklede granulære endoplasmatiske retikulumet gir syntese av en stor mengde protein. Proteinsyntese observeres bare i nevronets kropp og i dendrittene. Nerveceller er preget av et høyt nivå av syntetiske prosesser, primært protein og RNA.
I retning av aksonet og langs aksonet er det D.C. semi-flytende innhold av nevronet, beveger seg til periferien av neuritten med en hastighet på 1-10 mm per dag. I tillegg til den langsomme bevegelsen av nevroplasma, ble det også funnet rask strøm(fra 100 til 2000 mm per dag), den har en universell karakter. Rask strøm avhenger av prosessene med oksidativ fosforylering, tilstedeværelsen av kalsium, og forstyrres av ødeleggelsen av mikrotubuli og nevrofilamenter. Kolinesterase, aminosyrer, mitokondrier, nukleotider transporteres ved rask transport. Rask transport er nært knyttet til tilførsel av oksygen. 10 minutter etter døden stopper bevegelsen i den perifere nerven til pattedyr. For patologi er eksistensen av aksoplasmatisk bevegelse viktig i den forstand at ulike smittestoffer kan spre seg langs aksonet, både fra periferien av kroppen til sentralnervesystemet, og inne i det. Kontinuerlig aksoplasmatisk transport er en aktiv prosess som krever energi. Noen stoffer har evnen til å bevege seg langs aksonet i motsatt retning ( retrograd transport): acetylkolinesterase, poliomyelittvirus, herpesvirus, tetanustoksin, som produseres av bakterier fanget i et hudsår, når sentralnervesystemet langs aksonet og forårsaker kramper.
Hos en nyfødt er nevroplasmaet fattig på klumper av basofilt stoff. Med alderen observeres en økning i antall og størrelse på klumper.
Spesifikke strukturer av nerveceller er også nevrofibriller og mikrotubuli. nevrofibriller finnes i nevroner under fiksering og i cellekroppen har de et tilfeldig arrangement i form av filt, og i prosessene ligger de parallelt med hverandre. I levende celler ble de funnet ved bruk av fasekontrollfilming.
Elektronmikroskopi avslører homogene filamenter av neuroprotofibriller, bestående av neurofilamenter, i cytoplasmaet til kroppen og prosesser. Nevrofilamenter er fibrillære strukturer med en diameter på 40 til 100 A. De består av spiralvridde filamenter, representert av proteinmolekyler som veier 80 000. Nevrofibriller oppstår fra buntaggregering av nevroprotofibriller som eksisterer in vivo. På et tidspunkt ble funksjonen til å lede impulser tilskrevet nevrofibriller, men det viste seg at etter å ha kuttet nervefiberen, opprettholdes ledning selv når nevrofibriller allerede er i degenerering. Det er klart at hovedrollen i prosessen med impulsledning tilhører den interfibrillære nevroplasmaen. Dermed er den funksjonelle betydningen av nevrofibriller ikke klar.
mikrotubuli er sylindriske. Kjernen deres har lav elektrontetthet. Veggene er dannet av 13 langsgående orienterte fibrillære underenheter. Hver fibrill i sin tur består av monomerer som aggregerer og danner en langstrakt fibrill. De fleste mikrotubuli er plassert på langs i prosessene. Mikrotubuli transporterer stoffer (proteiner, nevrotransmittere), organeller (mitokondrier, vesikler), enzymer for syntese av mediatorer.
Lysosomer i nerveceller er de små, det er få av dem, og deres strukturer skiller seg ikke fra andre celler. De inneholder svært aktiv sur fosfatase. Lysosomer ligger hovedsakelig i kroppen til nerveceller. Med degenerative prosesser øker antallet lysosomer i nevroner.
I nevroplasmaet til nerveceller finnes inneslutninger av pigment og glykogen. To typer pigmenter finnes i nerveceller - lipofuscin, som har en blekgul eller grønngul farge, og melanin, et mørkebrunt eller brunt pigment (for eksempel et svart stoff - substantianigra i bena i hjernen).
Melanin funnet i celler veldig tidlig - ved slutten av det første leveåret. Lipofuscin
akkumuleres senere, men ved fylte 30 år kan det oppdages i nesten alle celler. Pigmenter som lipofuscin spiller en viktig rolle i metabolske prosesser. Pigmenter relatert til kromoproteiner er katalysatorer i redoksprosesser. De er det gamle redokssystemet til nevroplasmaet.
Glykogen akkumuleres i et nevron under en periode med relativ hvile i distribusjonsområdene til Nissl-stoffet. Glykogen finnes i kroppene og proksimale segmenter av dendrittene. Aksoner mangler polysakkarider. Nerveceller inneholder også enzymer: oksidase, fosfatase og kolinesterase. Neuromodulin er et spesifikt aksoplasmatisk protein.
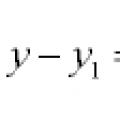 Ligninger av linjer og kurver i planet
Ligninger av linjer og kurver i planet Generalisert homogen likning Generaliserte homogene likninger av andre orden
Generalisert homogen likning Generaliserte homogene likninger av andre orden Takket være kommaet På grunn av det faktum at i deres
Takket være kommaet På grunn av det faktum at i deres