Максимальная нагрузка g. Единицы силы
Он, с одной стороны, помогает очистить генофонд от вредных мутаций, со-став-ляю-щих «ге-не-ти-чес-кий груз» , с другой стороны, может привести к вымиранию вида . Зависит это от характера полового отбора. Если он негативно влияет на при-спо-соб-лен-ность вида, то это ведёт к вымиранию . А если, как в случае с людьми, он будет только устранять генетический груз, то роль полового отбора будет ис-клю-чи-тель-но положительной .
Неважно насколько возвышенными и неземными нам кажутся любовные чувства, на самом деле, пред-по-чте-ния и и в отношении друг друга во многом рациональны . И даже такой, казалось бы, странный критерий отбора, как , на самом деле, отлично объясняется . Причём следует заметить, что половой отбор так же по-ло-жи-тель-но сказывается на скорости эволюции . А в случае с людьми половой отбор является единст-вен-ным методом отбора. Связано это с развитием технологий. Люди больше прак-ти-чес-ки не подвержены ес-тест-вен-но-му отбору , . Теперь выживают все, и это хорошо. Но многие из «выживших» оставляют потомство, и это плохо.
Из-за того, что практически все люди, не зависимо от своего здоровья, могут оставлять потомство, генофонд че-ло-ве-чест-ва ухудшается. Это не призыв становится «чайлдфри». Это констатация факта. Мы, как вид, перестали отсеивать негативные мутации посредством ес-тест-вен-но-го отбора. И это плохо! А единственным гуманным решением этой проблемы является развитие , способной исправить без жертв то, что природа исправляет жестокостью. Но пока что, в случае с людьми, отсеивание негативных мутаций происходит только за счёт полового отбора. И поэтому в нашем случае он играет ис-клю-чи-тель-но по-ло-жи-тель-ную роль. Хоть за это и приходится платить горькими юношескими слезами.
Негативная роль полового отбора
В случае с животными, находящимися в постоянной опасности быть жестоко убитыми и съеденными, роль полового отбора может быть не настолько по-ло-жи-тель-ной , . Связано это с тем, что повышение кон-ку-рен-то-спо-соб-нос-ти в половом отборе может негативно отразиться на выживаемости. Особи могут хуже при-спо-саб-ли-вать-ся к естественным условиям. Это отлично видно на экс-пе-ри-мен-тах с рыбами – . Самки предпочитают более ярких или пятнистых самцов, поэтому в условиях отсутствия хищников самцы становятся очень при-вле-ка-тель-ны-ми. Если же в аквариум запустить хищника, то через несколько поколений самцы станут менее яркими. Связано это с ролью ес-тест-вен-но-го и полового отбора.

Самых ярких самцов съедят хищники, поэтому они не оставят потомства или оставят, но меньше, в результате чего их мутации постепенно эли-ми-ни-ру-ют-ся. Самые тусклые самцы не пройдут половой отбор, поэтому их геном так же не будет широко представлен в популяции. Поэтому самым рас-про-стра-нён-ным фенотипом будет нечто промежуточное. Но в данном случае роль полового отбора очевидно негативная. Вместо того чтобы позволить самцам адап-ти-ро-вать-ся, самки вынуждают их оставаться яркими, в результате чего популяция рыб меньше, чем могла бы быть. И, в итоге, такой половой отбор может сократить про-дол-жи-тель-ность жизни всего вида . Поэтому не берите пример с рыб, и любите друг друга не за яркость, а за содержание!
Источники
Ncbi.nlm.nih.gov/pmc/articles/PMC1070813/
Ncbi.nlm.nih.gov/pmc/articles/PMC3151377/
Ncbi.nlm.nih.gov/pmc/articles/PMC2702796/
Ncbi.nlm.nih.gov/pmc/articles/PMC3684741/
Ncbi.nlm.nih.gov/pmc/articles/PMC1691857/
Nature.com/articles/s41586-018-0020-7
Ncbi.nlm.nih.gov/pmc/articles/PMC5319324/
Ncbi.nlm.nih.gov/pmc/articles/PMC5290736/
Ncbi.nlm.nih.gov/pmc/articles/PMC3210352/
Ncbi.nlm.nih.gov/pmc/articles/PMC4394296/
Ncbi.nlm.nih.gov/pmc/articles/PMC3721656/
Ncbi.nlm.nih.gov/pmc/articles/PMC3193187/
Academic.oup.com/beheco/article/14/2/194/192089
Sciencedirect.com/science/article/pii/S000334729690010X
Ncbi.nlm.nih.gov/pmc/articles/PMC3141438/
Ncbi.nlm.nih.gov/pmc/articles/PMC3797706/
Половой отбор – форма естественного отбора, основанная на соперничестве особей одного пола за спаривание с особями противоположного пола. При этом приспособленность генотипа (фенотипа) оценивается не по его выживаемости, а по его участию в размножении . Значение полового отбора заключается в активизации процесса размножения вида. Встречаются две формы.
При первой форме отбора самка пассивна, а самцы борются за право продолжать род, причем побеждает более сильный. В этом направлении у самца в ряду поколений увеличивались физическая сила, активность, более мощно развивались копыта, рога, клыки, шпоры (у петухов) и т. д.
При второй форме активна самка, которая выбирает более привлекательного самца для продолжения рода, что привело к конкуренции между самцами за более красивый и яркий наряд (большинство птиц и бабочек). Для каждого самца это может послужить причиной гибели, но для популяции в целом имеет положительное значение, так как повышает шансы этих самцов на продление рода.
Концепция полового отбора, объясняет происхождение многих признаков, которые, на первый взгляд, являются бесполезными или даже вредными и для особи, и для вида. К таким признакам относятся:
- сильная разветвлённость рогов у оленей, ослабляющая их значение как органов защиты или нападения,
- длинный тяжёлый хвост у самцов некоторых птиц во время брачного периода.
Половой отбор является прогрессивной формой отбора , поскольку «борьба» между самцами не ставит вопрос о выживаемости, о борьбе за условия, необходимые для жизни (пища, жизненное пространство и т. д.). Следовательно, половой отбор не требует гибели «побежденных»: «побеждённые», как правило, выживают и могут быть даже более долговечными, чем победители, а в последующий сезон спаривания могут оказаться «победителями».
Половой отбор сыграл значительную роль в эволюции человека. Основы концепции полового отбора разработаны Ч. Дарвином (Происхождение человека и половой отбор, 1871).
Естественный отбор подразделяется на индивидуальный и групповой . Индивидуальный отбор сводится к дифференциальному размножению отдельных особей, обладающих преимуществами в борьбе за существование в пределах популяций. Индивидуальный отбор основан на соревновании особей внутри популяций
Естественный отбор, «перерабатывая» незначительные наследственные различия особей и «складывая» их в определенном направлении, способствует постепенному отклонению потомков от предков. Любые признаки и свойства видов и более крупных таксонов формируются в процессе отбора особей на основе оценки их индивидуальных различий. На этом постоянном фоне индивидуального отбора в природе и осуществляется групповой отбор - преимущественное размножение особей какой-либо группы.
При групповом отборе в эволюции могут закрепляться признаки, благоприятные для группы, но не всегда благоприятные для особей. В групповом отборе группы особей соревнуются друг с другом в создании и поддержании целостности над организменных систем.
Родственные особи обладают большим сходством генотипов, чем неродственные. Поэтому если какой-либо признак у части родственных особей будет содействовать выживанию ближних, то такой признак может закрепиться в эволюции даже при условии его отрицательного влияния на непосредственного носителя. Например, особь, предупреждающая криком о появлении хищника своих родичей, чаще всего оказывается жертвой нападающего. Однако все остальные особи этой группы будут спасены, а поскольку альтруистический признак характерен хотя бы и для некоторых из них тоже, этот признак будет распространяться естественным отбором (отбором сородичей). Предполагается, что именно групповым отбором закреплены в эволюции свойства, связанные с регуляцией численности популяции. Групповой отбор может привести к вытеснению одной из конкурирующих групп и тем самым к уменьшению группового разнообразия либо к возникновению новых различий между формами и тем самым к понижению давления отбора. Наблюдения показали, что виды антилоп африканских саванн поедают разные части травянистых растений (одни едят только мягкие верхушки трав с цветками, другие - только сухие соломинки трав, третьи - колючие листья и т. п.). Такое положение - результат группового межвидового отбора, способствующего увеличению «суммы жизни» на единице площади.
Во всех без исключения случаях групповой отбор основан на внутрипопуляционном естественном отборе . Это и понятно, так как конкуренция видов в процессе эволюции осуществляется через конкуренцию их индивидов. Возникновение эволюционных новшеств происходит только при индивидуальном отборе, а групповой отбор выбирает уже из готовых приспособлений, возникших на внутривидовом уровне. Промежуточное положение занимает групповой внутривидовой отбор - отбор разных семей, популяций, групп популяций.
половой отбор
форма естественного отбора у животных, основанного на соперничестве особей одного пола (преимущественно мужского) за обладание особями другого пола, ведущая к уменьшению потомства у менее приспособленных. В результате полового отбора у многих видов животных возникли вторичные половые признаки (яркая брачная окраска, рога и др.).
Половой отбор
особая форма естественного отбора, определяющая возникновение в процессе эволюции вторичных половых признаков. К таким признакам относятся: яркая брачная окраска оперения уток, тетеревов и многих др. птиц, «танцы» насекомых, токование птиц, «турнирные бои» самцов птиц и млекопитающих, разнообразная звуковая сигнализация самцов, служащая для привлечения самок, пахучие железы для привлечения особей противоположного пола у насекомых, млекопитающих и т.д. Резко выделяющиеся признаки (окраска и т.п.) развиваются преимущественно у самцов; самки (особенно в период размножения), как правило, оказываются более защищенными покровительственной окраской и формой, соответствующим поведением и др. Первичной основой действия П. о. было расхождение в опознавательных признаках самца и самки. Тем самым, вероятно, облегчались встречи разнополых особей одного вида и предупреждались скрещивания с особями др. видов. Впоследствии особи с более резко выраженными половыми признаками легче привлекали особей др. пола, что вело к их преимущественному размножению. П. о. ≈ один из факторов, определяющих выработку этологических (поведенческих) механизмов изоляции. Иногда действие П. о. вступает в противоречие с действием др. направлений естественного отбора: сохраняются генотипы, которые увеличивают успешность размножения, но не увеличивают жизнеспособности вида в целом. Однако это обстоятельство не даёт основания противопоставлять П. о. естественному отбору и считать его самостоятельным фактором эволюции. Понятие П. о. выдвинуто и обосновано Ч. Дарвином (1859, и особенно 1871). См. также Косвенный отбор, Половой диморфизм.
Лит.: Дарвин Ч., Происхождение видов путём естественного отбора, пер. с англ., Соч., т. 3, М. ≈ Л., 1939; его же, Происхождение человека и половой отбор, пер. с англ., там же, т. 5, М., 1953; Шмальгаузен И. И., Проблемы дарвинизма, 2 изд., Л., 1969.
А. В. Яблоков.
Википедия
Половой отбор
Половой отбор - процесс, в основе которого лежит конкуренция за полового партнёра между особями одного пола , что влечёт за собой выборочное спаривание и производство потомства. Этот механизм может быть причиной эволюции некоторых характерных черт и приводить к их усилению. В пределах вида один из полов (практически всегда женский) играет роль ограниченного ресурса для другого (практически всегда мужского).
Половой отбор является частным случаем естественного отбора.
Некоторыми современными этологами высказывается мнение, что чувство любви и самопожертвования является следствием полового отбора и внутриродственной селекции на генетическом уровне.
Отбор на уровне одного гена, до сих пор рассматривавшийся как чересчур упрощенная, но полезная абстракция, был, по-видимому, реальным и даже главным селективным процессом на ранних стадиях органической эволюции, когда преобладающей единицей организации были геноподобные частицы. В современном мире существует процесс, близкий к отбору на уровне гена, дифференциальное размножение вирусных частиц или бактериальных клеток, различающихся по одному вирусному или бактериальному гену.
Некоторые генетически контролируемые отклонения мейоза у Drosophila melanogaster приводят к тому, что одна из хромосом гомологичной пары включается в более эффективно функционирующие сперматозоиды, чем другая. Нарушенное расщепление хромосом в мейозе ведёт к повышению частоты хромосомы одного типа в гаметном фонде по сравнению с частотой её гомолога. Этот процесс известен под названием мейотического драйва. Подобные же изменения частот наблюдаются у генов, локализованных в этих двух гомологичных хромосомах, и у фенотипов, детерминируемых этими генами. В одном случае со отношение полов изменяется в сторону избытка самок, в другом происходит повышение частоты одной рецессивной летали (Sandler, Novitski, 1957; Hiraizumi, Sandler, Crow, 1960). Мейотический драйв по существу представляет собой процесс дифференциальной репродукции гомологичных хромосом, т. е. отбор на уровне хромосом.
У цветковых растений мужские гаметы находятся в самостоятельной структуре мужском гаметофите, состоящем из пыльцевого зерна и пыльцевой трубки. Гетерозиготное растение продуцирует генетически различные классы пыльцы; в некоторых случаях пыльцевые зерна расщепляются по генетическим факторам, влияющим на жизнеспособность пыльцы, на её способность к прорастанию или на скорость роста пыльцевых трубок. Кроме того, пыльца обычно продуцируется и попадает на рыльце в больших количествах, чем это нужно для оплодотворения, так что между пыльцевыми зернами или пыльцевыми трубками возникает конкуренция. В тех случаях, когда пыльца расщепляется по факторам роста, в результате этой конкуренции некоторые классы мужских гамет оказываются более эффективными, чем другие при оплодотворении, и, следовательно, чаще вносят вклад в образование зародышей или эндосперма, т. е. происходит отбор на уровне гамет.
Допустим, что какой-то ген-маркер, детерминирующий определённый внешне заметный морфологический признак, сцеплен с геном, детерминирующим рост пыльцевой трубки. В таком случае среди потомков вместо ожидаемого менделенского расщепления по этому гену будет наблюдаться отклоняющееся соотношение: избыток среди семян или проростков морфологического типа, детерминируемого аллелем-маркером, привнесенным пыльцой, превосходящей в селективном отношении пыльцу другого класса, и снижение доли противоположного морфологического типа. Изменение соотношений при расщеплении, обусловленное низкой селективной ценностью некоторых классов пыльцы, хорошо известно у разных видов цветковых растений.
Хорошо проанализирован случай сцепленных генов Su (определяющего тип эндосперма) и Ga (контролирующего рост пыльцевой трубки) у кукурузы (Zea mays ). Гетерозиготы Su/su обычно дают зерна, разделяющиеся по характеру эндосперма на два класса (крахмалистый и сахаристый эндосперм) в менделевских соотношениях. Допустим, что аллель su (сахаристый эндосперм) сцеплен с аллелем ga (медленный рост пыльцевой трубки) в гетерозиготе (Su Ga/su ga) и такая гетерозигота используется в качестве отцовской особи, так что сцепленный участок su-ga передается через пыльцу. В этом случае в следующем поколении наблюдается заметная нехватка зерен сахаристого типа (Mangelsdorf, Jones, 1926).
Половой отбор
Половой отбор основан на селективной неравноценности особей одного пола у раздельнополых организмов (обычно у животных). Это особая форма индивидуального отбора, в которой участвуют представители только одного пола (обычно самцы) данной популяции.
Половой отбор начинается с «разделения труда» между гаметами и между их носителями, возникшего на ранних этапах эволюции полового размножения. Специализация яиц и в конечном счете особей женского пола была направлена на обеспечение питания и защиты зародыша, а сперматозоиды и в конечном счете особи мужского пола на нахождение яиц и их оплодотворение. Это разделение функций привело в самом начале к численному преобладанию сперматозоидов над яйцами, к большей подвижности сперматозоидов или самцов по сравнению с яйцеклетками или самками и к развитию у самцов более сильного полового влечения, чем у самок. Все это заложило основы для возникновения селективных различий между самцами.
Половой диморфизм у животных нередко выражается в различиях по двум группам признаков. Первичные половые признаки это признаки, непосредственно относящиеся к размножению. Здесь мы не будем их рассматривать. Вторичные половые признаки, имеющиеся у самцов, помогают им отыскивать брачных партнеров. К этим вторичным половым признакам и обращена теория полового отбора.
Вторичные половые признаки можно разделить на два обширных класса: 1) признаки, относящиеся к размерам, силе и разного рода придаткам (например, более крупные размеры самцов у морских львов или рога у самцов оленей); 2) украшения и демонстрационное поведение (например, яркое оперение селезней, яркие пятна на шее у самцов колибри, особые типы песен, брачное поведение). В «Происхождении человека и половом отборе» Дарвин (1871) рассмотрел известные в его время примеры вторичных половых признаков у самцов животных, принадлежащих к широкому кругу групп, и предложил для их объяснения теорию полового отбора.
Дарвин представлял себе половой отбор как процесс, дополняющий более общий и более широко распространенный процесс естественного отбора. Последний, по теории Дарвина, создал адаптивные признаки данного вида в целом, в том числе изначальные приспособления, общие для обоих полов, а также первичные половые различия, непосредственно связанные с размножением. Естественный отбор в первоначальном: дарвиновском смысле не объяснял вторичные половые признаки. Такие признаки, как рога оленя или яркое оперение селезней, не представляют собой приспособлений, благоприятных для вида в целом; не являются они также необходимыми и для размножения. Однако они, по-видимому, повышают вероятность успешности спариваний для тех самцов, которые ими обладают. Теория полового отбора была введена для того, чтобы объяснить развитие таких специальных мужских признаков.
Дарвин описывает процесс, предложенный им в качестве объяснения, следующим образом (Darwin, 1871) : «…половой отбор… зависит от преимуществ, относящихся исключительно к воспроизведению, которыми обладают одни особи перед другими особями того же пола и вида». Далее он объясняет (я здесь несколько перефразировал утверждение Дарвина, сохранив его терминологию): если оба пола ведут совершенно одинаковый образ жизни, но у самца органы чувств или органы передвижения значительно более развиты, чем у самки, то возможно, что самцам нередко удается приобрести эти признаки не потому, что они лучше приспособлены к выживанию в борьбе за существование, а потому, что они обладают известными преимуществами перед другими самцами при размножении. «В подобных случаях должен был вступать в действие половой отбор».
Изучение полового отбора начиная с 1871 г. шло извилистым путем. Во времена Дарвина эта тема была предметом разногласий. Уоллес (Wallace, 1889) допускал роль полового отбора в развитии признаков, необходимых самцам в драках, но не в развитии демонстрационных признаков. Затем эта проблема была предана забвению. Когда в начале современного периода эволюционных исследований она вновь выступила на сцену в работах Фишара (Fisher, 1930) и Гексли (Huxley, 1938), ситуация была совершенно иной: те же самые явления рассматривались с других альтернативных, точек зрения и теория полового отбора должка была дебютировать вновь. Этот второй дебют оказался удачным: в настоящее время половой отбор является предметом активных исследований и ему посвящена обширная литература .
Для того чтобы оценить современное состояние проблемы естественного отбора, следует прежде всего ясно представить себе огромное разнообразие вторичных половых признаков, несущих самые разные функции в жизни животных. Возникновение всех этих признаков, по-видимому, нельзя отнести за счет отбора какого-то одного типа. Различия в общих размерах между самцами и самками могут быть обусловлены отбором, направленным на экологическую дивергенцию (см, часть ), а также половым отбором. Мощные рога и другое «оружие» самцов может служить для захвата территории или достижения доминирующего положения в сообществе, а также для завоевания самок. Разного рода украшения, демонстрации, песни и запах самцов могут служить стимуляторами при ухаживании и видоспецифичными опознавательными сигналами. В последнем случае они могли возникнуть в результате отбора, направленного на создание репродуктивной изоляции (см. часть ), а также полового отбора.
Возникновение всего обширного спектра вторичных половых признаков результат совместного действия различных селективных процессов, поэтому трудно выделить здесь эффекты одного только полового отбора.
В последние годы делаются попытки распространить концепцию полового отбора на двудомные растения. Эти попытки выражаются в истолковании известных фактов биологии размножения растений в терминах сильно расширенной концепции полового отбора. В частности, указывают на очень большое число пыльцевых зерен по сравнению с числом яйцеклеток и возникающую в результате конкуренцию между этими зернами (Stephenson, Bertin, 1983; Willson, 1983). Однако такое положение вещей результат исходного разделения труда между самцами и самками; это непременное предварительное условие для процесса полового отбора, как уже упоминалось, а не его результат. Явление несовместимости у растений, некоторые типы пыльцы которых успешно прорастают лишь в определённом семязачатке, рассматривается как выбор со стороны особи женского пола (Stephenson, Bertin, 1983; Willson, Burley, 1983). В этом случае концепция такого выбора расширяется, охватывая иммунологические реакции организмов, лишенных нервной системы или умственных способностей. Межсамцовый отбор теоретически возможен у двудомных растений. Однако эта проблема в настоящее время далека от решения из-за отсутствия критического подхода, а также необходимых данных.
Признаки самцов, связанные с драками между ними
Роль полового отбора ясно выражена в развитии признаков, связанных с драками между самцами и с явлением доминирования. Для полового отбора этого типа необходимо наличие трёх условий: 1) конкуренция между самцами за самок; 2) генотипические различия между самцами, определяющие их конкурентоспособность в борьбе за самок; 3) репродуктивное преимущество самцов, добившихся успеха, над другими самцами.
Первое условие имеется у млекопитающих и птиц с полигинной (полигамной) системой размножения. У полигинных видов самые сильные самцы собирают вокруг себя гаремы из самок и охраняют самих самок или территорию, на которой они находятся» отгоняя более слабых самцов, так что на долю последних остается мало самок или не остается вовсе.
Полигиния часто встречается у млекопитающих и иногда у птиц. Среди млекопитающих она наблюдается у оленей, крупного рогатого скота, овец, у большинства антилоп, у слонов, тюленей, морских львов, моржей и павианов; среди птиц у кур, фазанов и павлинов. У самцов этих животных хорошо развиты вторичные половые признаки в отличие от самцов родственных неполигинных групп. Так, у полигинных кур, фазанов и павлинов самцы гораздо крупнее, драчливее, и оперение у них наряднее, чем у самок, а у моногинных серой куропатки, грауса и тундряной куропатки различия между особями разного пола относительно невелики. У моржей и морских львов самцы очень крупные; самцов многих копытных украшают рога; у павианов самцы крупные и агрессивные. В отличие от этого у моногинных волков и некоторых моногинных видов обезьян, а также у ряда кошек, у которых детенышей выращивает в основном мать, и у колониальных, но неполигинных грызунов, самцы и самки почти не отличаются друг от друга по размерам и силе (Darwin, 1871, гл. 8).
Поскольку признаки, связанные с драками между самцами, наблюдаются преимущественно в полигинных группах, можно считать, что они возникли в результате полового отбора. Корреляция между диморфизмом по общим размерам и системой спаривания выражена менее ясно, не говоря уже о многочисленных исключениях и осложнениях (Ralls, 1977).
У млекопитающих и птиц наблюдается также и другая корреляция между системой спаривания и вкладом родителей в заботу о потомстве (Trivers, 1972; Zeveloff, Boyce, 1980).
У птиц самцы и самки принимают более или менее одинаковое участие в различных аспектах заботы о потомстве, т. е. в постройке гнезда и выкармливании птенцов. Стратегия размножения, характерная для большинства птиц, делает необходимым сотрудничество обеих родительских особей. Это ведет к моногамии, которая широко распространена у птиц. При моногамии, когда у каждого самца есть брачный партнер, конкуренции между самцами за самок не возникает. Соответственно не развиваются и признаки, необходимые самцам в драках и для завоевания доминирующего положения.
Полигинные птицы, для которых характерно доминирование самцов, представляют собой исключение, подтверждающее правило.
У млекопитающих стратегия размножения основана, напротив, на преимущественной роли матери в заботе о потомстве, что обусловлено вынашиванием зародыша во чреве и выкармливанием детеныша собственным молоком. Вклад самцов в заботу о потомстве часто пренебрежимо мал. Свободные от этой заботы самцы получают возможность конкурировать за самок и устраивать гаремы. Это создаёт почву для полового отбора по признакам, связанным с доминированием самцов, которые часто встречаются у млекопитающих. Моногамные млекопитающие с их равноправием между полами исключение, подтверждающее правило.
Репродуктивное поведение у снежного барана и у благородного оленя
Половой отбор будет эффективным лишь в том случае, если победившие самцы оставят больше выживающих потомков, чем самцы, не достигшие успеха (условие 3, указанное в предыдущем разделе). Сведения об относительной успешности размножения отдельных самцов в природных условиях получить трудно и в литературе их немного. Однако в настоящее время таких данных становится больше, и мы приведем здесь два примера.
Клаттон-Брок и др. (Clutton-Brock et al., 1982), изучавшие в течение длительного времени благородного оленя (Cervus elaphus ) в Шотландии, получили данные о репродуктивном успехе отдельных особей. Для этого вида характерен резко выраженный половой диморфизм. Самцы весят почти вдвое (в среднем в 1.7 раза) больше самок. Во время осеннего гона между самцами происходят драки, исход которых определяется общими размерами, силой и величиной рогов у самцов. Победители собирают себе гаремы, причём число самок в гареме варьирует от 1 до 22, в зависимости от бойцовских качеств самца, а побежденные остаются одинокими.
Самцы, добившиеся успеха, покрывают за каждый сезон размножения несколько или много самок, тогда как неудачники иногда не спариваются вовсе. В выборке, состоявшей из 13 самцов, число выживших потомков на одного самца (за всю его жизнь) колебалось от 0 до 25. Это число коррелирует с величиной гарема, которая, как мы видели, зависит от бойцовских качеств самцов. Совершенно очевидно, что различия между самцами в отношении репродуктивного успеха очень велики (Clutton-Brock et al., 1982).
Признаки, определяющие привлекательность самцов
Гораздо менее ясна роль полового отбора в развитии демонстрационных признаков самцов, таких, как разного рода украшения, пение и феромоны, в неполигинных группах животных. Названным признакам можно дать различные объяснения, не обязательно связанные с половым отбором. Кроме того, существуют проблемы, касающиеся адекватности самого селективного механизма. Половой отбор по признакам, связанным с украшениями и демонстрационным поведением самцов, подразумевает выбор, осуществляемый самками сугубо индивидуально, а это фактор, не поддающийся количественной оценке.
Отдают ли самки предпочтение самым привлекательным или самым сильным самцам, и постоянны ли они в своих предпочтениях? А у неполигинных животных, у которых число самок и самцов одинаково и они образуют брачные пары, оставят ли самки, спаривающиеся с самцами, которых они сразу предпочли всем остальным, больше потомков, чем те, которым пришлось выбирать вторично? Половой отбор по признакам, определяющим привлекательность самцов, был бы эффективен, если бы все или большинство самок предпочитали самцов какого-то одного типа, но выполняется ли это условие в природе? Что происходит, если самки различаются в отношении предпочитаемого типа самца? Если же для всех или для большинства самок данного вида существует одинаковый стандарт привлекательности самцов, то каким образом этот общий для всего вида стандарт возник впервые?
В некоторых недавних исследованиях обнаружена связь между предпочтениями самок и репродуктивным успехом самцов избираемого типа. У бабочек Colias (С. eurytheme и С. philodice ) самки делают выбор между самцами на основании кинетики полета. Предпочитаемые самцы достигают наибольшего успеха в спаривании и, по-видимому, обладают также наилучшей способностью к полету (Watt et al., 1986). У лягушки Physalaemus pustalosus в Панаме самки предпочитают определённый тип звуковых сигналов. Издающие такие крики самцы имеют больше шансов на спаривание. Эти успешно спаривающиеся самцы, кроме того, старше и крупнее, так что в этом случае, так же как и у бабочек Colias , по-видимому, играет роль приспособленность (Ryan, 1980; 1983; 1985). Самки бычка Cottus bairdi предпочитают спариваться с крупными самцами, причём успешно спаривающиеся самки откладывают более жизнеспособные яйца (Downhower et al., 1983). В этих и в сходных случаях выбор самок основан на признаке, связанном с общей приспособленностью самцов.
Подобные случаи не позволяют объяснить обширный класс демонстрационных признаков самцов, которые, казалось бы, не дают своим обладателям никаких преимуществ и даже могут оказывать пагубное воздействие на их жизнеспособность. Принято считать, что длинные хвосты у самцов некоторых райских птиц снижают их приспособленность. Брачные призывы самцов панамских лягушек повышают вероятность уничтожения их хищниками летучими мышами (Trachops cirrhosus ) (Tuttle, Ryan, 1981).
Есть основания сомневаться в том, что механизм полового отбора может создавать и действительно создаёт хорошо заметные и не адаптивные демонстрационные признаки самцов. Возникновение этого класса признаков можно, однако, объяснить другим сочетанием селективных сил, действие которых слагается из ряда последовательных стадий. Вначале таким демонстрационным признакам благоприятствует обычный индивидуальный отбор, способствуя ухаживанию и спариванию. Конкретные демонстрационные признаки и ритуалы ухаживания в пределах данного вида, вероятно, могли иметь расовые особенности, как, например, локальные диалекты у некоторых видов птиц. Расовые различия должны были бы вызывать некоторую начальную репродуктивную изоляцию. Дальнейшая дивергенция на видовом уровне обычно должна усиливать эту экологическую изоляцию и лежащие в её основе признаки, как побочный результат такой дивергенции. И наконец, демонстрационные признаки и ритуалы ухаживания дивергентных видов должны дифференцироваться ещё сильнее в результате отбора, направленного на этологическую изоляцию как таковую, с тем чтобы подавить межвидовую гибридизацию (см. часть ).
Эта гипотеза согласуется с той большой ролью, которую играют демонстрационные признаки самца и выбор со стороны самки в этологической изоляции. Выбор самкой брачного партнера в сущности касается межвидовых отношений в не меньшей мере, чем внутривидовых. Самки обычно принимают самцов своего вида, но отвергают самцов с демонстрационными признакаками чужого вида. Короче говоря, наличие у самцов неадаптивных демонстрационных признаков можно объяснять как результат видообразования.
Связь между этологической изоляцией, видообразованием и половым отбором была отмечена недавно у рыб семейства Cichlidae из африканских озер и у группы Drosophila sitvestris с Гавайских островов (Dominey, 1984; Carson, 1986; Mayr, 1988). Эти авторы считают половой отбор движущей силой в развитии этологической изоляции и видообразовании. Такое объяснение соответствует имеющимся фактам. Равным образом, однако, можно считать, что главным определяющим фактором является видообразование, как это описано выше. Мне, вообще, не кажется необходимым привлекать половой отбор в строгом смысле для объяснения именно этих и других сходных случаев.
Междемовый отбор
Перейдем теперь к вопросу об отборе на популяционном уровне, вызывающему большие разногласия. Междемовый отбор, который часто называют также групповым отбором, представляет собой дифференциальное размножение разных локальных популяций. Мы начнем с рассмотрения междемового отбора у организмов, не образующих сообществ.
Одна из особенностей органического мира, которую трудно объяснить на основе индивидуального отбора, но можно рассматривать как результат междемового отбора, это половое размножение. Хотя и были созданы модели, в которых половому размножению благоприятствует индивидуальный отбор (Williams, 1975), однако они представляются нереалистичными. Половое размножение это тот процесс, который создаёт рекомбинационную изменчивость в скрещивающихся популяциях. От полового размножения выигрывают не родительские генотипы, распадающиеся в процессе рекомбинации, а популяция будущих поколений, у которых возрастает запас изменчивости. Это подразумевает участие в качестве одного из факторов селективного процесса на популяционном уровне .
Социально-групповой отбор
Общественные перепончатокрылые представляют собой один из примеров крайне высокой интегрированности сообществ в мире животных, и у них очень ярко выражены эффекты социально-группового отбора. Отбор этого типа, вероятно, действует также в группах с не столь высокоинтегрированной структурой сообщества, например у птиц и млекопитающих. В части VIII мы попытаемся показать, что некоторые особенности вида Homo sapiens , возможно, возникли в результате социально-группового отбора.
Альтруизм
Альтруистическое поведение животных слагается из разнообразных специфических особенностей поведения. В общем и целом его можно определить как поведение, приносящее пользу другим особям. Его диапазон очень широк от поведения, благоприятствующего другим особям в умеренной степени, до подлинного самопожертвования, вплоть до самоубийства. Альтруистическое поведение может принести пользу потомкам, другим родичам или социальным группам.
Существование различных типов альтруистического поведения позволяет считать, что в их создании могут участвовать разные типы отбора. Рассмотрим три случая.
Первый случай альтруистическое поведение родительских особей по отношению к своим потомкам. К этому типу альтруистического поведения можно отнести общее явление заботы о потомстве у птиц и млекопитающих или особые виды поведения, характерные для чибисов, крикливых зуйков и других землегнездников, которые притворяются ранеными, отвлекая внимание хищника, или отводят его от гнезда. Забота о потомстве явно результат индивидуального отбора, поскольку индивидуальный отбор благоприятствует сохранению генов тех родительских особей, которые оставляют наибольшее число выживающих потомков.
Рассмотрим теперь связанное с самопожертвованием оборонительное поведение рабочих особей у общественных пчел, таких, как Apis mellifera . Когда рабочая пчела использует жало, это равносильно для нее самоубийству, но полезно для колонии, так как предотвращает вторжение врага. Колонии, обороняющиеся лучше других, выживают и размножаются более успешно, чем колонии с менее эффективной защитой. Самопожертвование рабочих пчел, наряду с другими характеристиками касты рабочих, можно адекватно объяснить как результат социально-группового отбора, поскольку оно выгодно колонии пчел в целом.
Третий случай группы примитивных людей, находящихся на стадии собирательства и охоты, примером которых могут служить бушмены юго-западной Африки (Lee, Devore, 1976), Эти сообщества представляют собой организованные группы, в которые входят члены семьи, другие родственники, свойственники, а иногда случайные гости из других групп. В них глубоко укоренен обычай разделения пищи. Если убито крупное животное, его мясо раздается всем членам группы независимо от того, родственники это или случайные посетители. В таких группах развиваются также другие типы кооперативного поведения.
Допустим теперь в порядке обсуждения, что распределение корма и другие аналогичные типы социального поведения имеют какую-то генетическую основу; это позволит нам попытаться изучить типы отбора, которые, возможно, участвуют в развитии такого поведения.
Индивидуальный отбор, благоприятствующий развитию заботы о потомстве, вероятно, очень интенсивен. Трудно представить себе, однако, чтобы члены сообщества делились пищей только со своими потомками, обделяя других его членов и близких родичей, поскольку поведенческий фенотип и «социальное давление» со стороны других членов группы обычно обладают пластичностью. Приматам, ведущим общественный образ жизни, не свойственны жесткие и строго направленные типы поведения. Поведение, связанное с распределением пищи, должно естественным образом выходить за пределы своих первоначальных целей, т. е. снабжения пищей потомков, и распространяться на всю семью и родственную группу. Следует также ожидать, что социально-групповой отбор должен способствовать развитию такого поведения. Группа в целом зависит от объединения её членов в действиях, связанных с добыванием пищи, которые в сущности обеспечивают выживание, и она должна выигрывать от распределения пищи на широкой основе. Тенденция делиться пищей, усиливаемая социально-групповым отбором, должна распространяться на всех членов группы, как кровных родичей, так и «свойственников» в равной мере. Такое поведение, вероятно, пересекается с типами поведения, создаваемыми в результате индивидуального отбора среди родичей промежуточного ранта. Короче говоря, распределение пищи можно было бы адекватно объяснить как результат совместного действия индивидуального и социально-группового отбора, направленного на создание пластичных культурных традиций.
Кин-отбор
Термин кин-отбор ввел Мэйнард Смит (Maynard Smith, 1964Приведена наиболее подходящая ссылка (прим. сканера) ")">*), определив его как отбор, направленный на сохранение признаков, благоприятствующих выживанию близких родичей данной особи. Такими родичами могут быть либо потомки данной особи, либо её сибсы и другие родичи. В качестве примеров признаков, возникших в результате кин-отбора, он привел заботу о потомстве, имитацию ранения, а также касты стерильных рабочих у общественных насекомых.
Мэйнард Смит и более позние исследователи считают кин-отбор процессом, близким к социально-групповому отбору, и, возможно, промежуточным между индивидуальным и групповым отбором.
Кин-отбор приобрел популярность как концепция и термин и очень широко используется в литературе. Тем не менее я считаю, что он мало что объясняет, но зато способен внести большую путаницу в мышление (см. Grant, 1978). Отчасти кин-отбор более поздний синоним индивидуального отбора, направленного на развитие заботы о потомстве, а отчасти более поздний синоним группового отбора .
Явления, приписываемые кин-отбору, вполне объяснимы этими другими типами отбора, т. е. индивидуальным отбором по заботе о потомстве, или социально-групповым отбором, или совместным действием этих двух процессов.
Некоторые авторы предпочитают термин кин-отбор прежнему термину групповой отбор, потому что он подчеркивает роль родства. Однако в концепции группового отбора глубоко заложено родство. Все социальные группы животных это родственные группы. В отдельном термине кин-отбор нет нужды.
Можем ли мы найти в природе какие-нибудь явления, которые определяются кин-отборам в узком смысле, исключающем такие принятые ранее типы отбора, как индивидуальный отбор и социально-групповой отбор? Некоторые явления, относимые обычно за счет кин-отбора, такие, как альтруизм и касты рабочих у общественных насекомых, мы уже обсудили. Другой набор признаков предупреждающая окраска у несъедобных насекомых в течение некоторого времени считался результатом кин-отбора.
Хищник убивает насекомое, имеющее предупреждающую окраску, обнаруживает, что оно несъедобно, и научается распознавать цветовой сигнал. Это идет на пользу сибсам погибшего насекомого, имеющим такую же окраску. Объяснение предупреждающей окраски на основе кин-отбора исходит из допущения, что хищник убивает несъедобных насекомых, когда пробует их съесть. Викланд и Ярви (Wiklund, Jarvi, 1982) проверили это допущение в экспериментах на большом числе насекомых с предупреждающей окраской, которых они предлагали птицам четырёх видов. Оказалось, что птицы обычно не убивают насекомое. Они «пробуют» его, отвергают и часто (в 84% проб) насекомое остается живым. Поэтому несъедобность и предупреждающая окраска вполне объяснимы на основании одного лишь индивидуального отбора (Wiklund, Jarvi, 1982).
Отбор на видовом уровне
По меньшей мере три типа отбора действуют на видовом уровне: 1) отбор, направленный на развитие экологической дивергенции; 2) замещение видов; 3) отбор, приводящий к репродуктивной изоляции. Эти типы отбора рассматриваются в части , а здесь они перечислены просто для того, чтобы завершить наш обзор.
Обсуждалась также возможность селективных процессов на уровне биотических сообществ. Высказывалось мнение, что более стабильные сообщества, возможно, превосходят менее стабильные по длительности существования в экологических масштабах времени (MacArthur, 1955; Hutchinson, 1959; Dunbar, 1960* 17
 Корпускулярные и волновые свойства частиц
Корпускулярные и волновые свойства частиц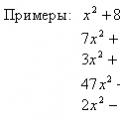 Уравнения онлайн X 5 0 решение
Уравнения онлайн X 5 0 решение Предельные монокарбоновые кислоты
Предельные монокарбоновые кислоты