Чим є комплекс гольджі у рослинних клітинах. ЕПС та комплекс Гольджі
Клітина - цілісна система
Жива клітина - унікальна досконала дрібна одиниця організму, вона влаштована так, щоб максимально ефективно використовувати кисень і поживні речовини, виконуючи свої функції. Життєво важливими для клітин органелами є ядро, рибосоми, мітохондрії, ендоплазматичний ретикулум, апарат Гольджі. Ось про останнє і поговоримо докладніше.
Що це таке
Ця мембранна органела є комплексом структур, які виводять із клітини синтезовані в ній речовини. Найчастіше вона розташовується поблизу зовнішньої клітинної мембрани.
Апарат Гольджі: будова
Він складається з утворених мембранами "мішочків", званих цистернами. Останні мають витягнуту форму, трохи сплющені посередині і розширені по краях. Також у комплексі є круглі бульбашки Гольджі – дрібні мембранні структури. Цистерни складені стопочками, які називаються диктіосоми. Апарат Гольджі містить різні типи "мішочків", весь комплекс ділять на деякі частини за рівнем віддаленості від ядра. Розрізняють їх три: цис-відділ (ближче до ядра), серединний, і транс-відділ – найдальший від ядра. Вони характеризуються різним складом ферментів, отже, і виконуваної роботою. У будові диктіос є одна особливість: вони полярні, тобто найближчий до ядра відділ тільки приймає бульбашки, що йдуть від ендоплазматичного ретикулума. Частина “стопки”, звернена до мембрани клітини, лише формує та віддає їх.

Апарат Гольджі: функції
Основними завданнями є сортування білків, ліпідів, слизових секретів та їх виведення. Також через нього проходять небілкові речовини, що виділяються клітиною, вуглеводні компоненти зовнішньої мембрани. При цьому апарат Гольджі зовсім не є індиферентним посередником, який просто передає речовини, в ньому йдуть процеси активізації та модифікації (дозрівання):
- Сортування речовин, транспортування білків. Розподіл білкових речовин відбувається на три потоки: для мембрани самої клітини, експортні, лізосомальні ферменти. До першого потоку крім білків включаються і жири. Цікавий факт, що будь-які експортні речовини переносяться усередині бульбашок. А ось призначені для мембрани клітини білки вбудовуються в мембрану транспортної бульбашки та переміщуються таким чином.
- Виділення всіх продуктів, вироблених у клітині. Апарат Гольджі "упаковує" всю продукцію, як білкову, так і іншої природи, в секреторні бульбашки. Усі речовини виділяються назовні шляхом складної взаємодії останніх із клітинною мембраною.
- Синтез полісахаридів (глікозаміногліканів та компонентів глікоколіксу клітинної стінки).
- Сульфатування, глікозилювання жирів і білків, частковий протеоліз останніх (необхідний для переведення їх з неактивної форми в активну), - це всі процеси "дозрівання" білків, необхідні для майбутньої повноцінної роботи.

На закінчення
Розглянувши те, як влаштований та працює комплекс Гольджі, переконуємось, що він є найважливішою та невід'ємною частиною будь-якої клітини (особливо секреторних). Клітина, що не продукує речовин на експорт, також не може обійтися без цієї органели, оскільки від неї залежить укомплектованість клітинної мембрани та інші важливі внутрішні процеси життєдіяльності.
Комплекс Гольджі було виявлено Камілло Гольджі у 1898 році. Ця структура присутня в цитоплазмі майже всіх еукаріотичних (що становлять вищі організми) клітин, особливо, секреторних клітин у тварин.
Гольджі комплекс. Будова.
Структура представлена чаркою сплощених мішечків мембран. Вони називаються цистернами. Ця стопка мішечків пов'язана із системою Гольджі). З одного кінця стопок мішечків постійно відбувається формування нових цистерн злиттям бульбашок, які відбруньковуються від ендоплазматичного ретикулуму (мережі порожнин). З іншого ж кінця стопки з внутрішньої сторони цистерни завершують дозрівання і знову розпадаються на бульбашки. Так відбувається поступове переміщення цистерн у сопці до внутрішньої сторони від зовнішньої сторони.
У цистернах структури відбувається дозрівання білків, призначених для секреції, трансмембранних білків лізосом та інших. Дозріваючі речовини переміщаються послідовно цистернами органели. У них відбувається остаточне згортання білків та їх модифікації – фосфорилювання та глікозилювання.
Характеризуються наявністю низки окремих диктіосів (стопок). У часто присутній кілька стопок з'єднаних трубками або одна велика стопка.
Містить чотири головні відділи: транс-Гольджі мережу, цис-Гольджі, транс-Гольджі та медіал-Гольджі. До структури приєднують також проміжний компартмент (відокремлену область). Він представлений скупченням мембранних бульбашок у проміжку між реіткулумом та цис-гольджі.
Весь апарат є органелою поліморфною (різноманітною). Навіть на різних етапах розвитку однієї клітини комплекс Гольджі може виглядати по-різному.
Відрізняється апарат та асиметрією. Розташовані ближче до клітинного ядра цистерни (цис-Гольджі) містять у собі незрілі білки. До цистерн цим приєднуються безперервно мембранні бульбашки - везикули. У різних цистернах містяться різні резидентні ферменти (каталітичні), що говорить про те, що в них з білками, що дозрівають, відбуваються послідовно різні процеси.
Гольджі комплекс. Опції.
До завдань структури відносяться хімічна модифікація та транспорт речовин, що надходять до нього. Білки, що проникають в апарат з ендоплазматичного ретикулуму, є для ферментів вихідним субстратом. Пройшовши концентрування та модифікацію, ферменти у бульбашках транспортуються до призначеної ділянки. Наприклад, це може бути область формування нової нирки. За участю цитоплазматичних мікротрубочок процес перенесення здійснюється найактивніше.
Комплекс Гольджі виконує також завдання приєднання вуглеводних груп до білків і подальшого використання цих білків при побудові мембрани лізосом і клітин.
В окремих водоростей у структурі апарату здійснюється синтез целюлозних волокон.
Функції комплексу Гольджі досить різноманітні. Серед них слід виділити:
- Сортування, виведення, накопичення секреторних продуктів.
- Накопичення ліпідних молекул та формування ліпопротеїдів.
- Завершення модифікації білків (посттрансляційної), а саме глікозування, сульфатування та інше.
- Формування лізосом.
- Участь у освіті акросоми.
- Полісахаридний синтез для формування восків, глікопротеїдів, слизів, камеді, речовин матриксу у рослин (пектинів, геміцелюлози та інших).
- Освіта скорочуваних вакуолей у найпростіших.
- Освіта клітинної платівки у рослинних клітинах після поділу ядра.
18. Комплекс Гольджі, його будова та функції. Лізосоми. Їх будова та функції. Типи лізосом.
Комплекс Гольджіє стопкою дископодібних мембранних мішечків (цистерн), дещо розширених ближче до країв, і пов'язану з ними систему бульбашок Гольджі. У рослинних клітинах виявляється ряд окремих стопок (диктіосоми), у тваринних клітинах часто міститься одна велика або кілька, з'єднаних трубками, стопок.
1. Накопичує та виводить органічні речовини, що синтезуються в ендоплазматичній мережі
2. Утворює лізосоми
3. Формування вуглеводних компонентів глікокаліксу – в основному, гліколіпідів.
Лізосомиє невід'ємною частиною складу клітини. Вони є різновидом везикул. Ці клітинні помічники, будучи частиною вакуома, покриті оболонкою з мембрани та наповнені гідролітичними ферментами. Важливість існування лізосом усередині клітини забезпечена секреторною функцією, яка необхідна у процесі фагоцитозу та аутофагоцитозу.
Виконують травну функцію- перетравлюють харчові частинки та видаляють відмерлі органоїди.
Первинні лізосоми- це дрібні мембранні бульбашки, які мають деаметр близько ста нм, заповнені гомогенним дрібнодисперсним вмістом, що є набір гідролітичних ферментів. У лізосомах є близько сорока ферментів.
Вторинні лізосомиутворюються при злитті первинних лізосом з ендоцитозними або піноцитозними вакуолями. Якщо сказати інакше, то вторинні лізосоми – це внутрішньоклітинні травні вакуолі, ферменти яких поставляються первинними лізосомами, а матеріал для перетравлення – ендоцитозною (піноцитозною) вакуоллю.
19. Епс, її різновиди, що у процесах синтезу речовин.
Ендоплазматична мережау різних клітинах може бути представлена у формі сплощених цистерн, канальців або окремих везикул. Стінка цих утворень складається з біліпідної мембрани та включених до неї деяких білків і відмежовує внутрішнє середовище ендоплазматичної мережі від гіалоплазми.
Розрізняють два різновиди ендоплазматичної мережі:
зерниста (гранулярна або шорстка);
незерниста або гладка.
На зовнішній поверхні мембран зернистої ендоплазматичної мережі містяться прикріплені рибосоми. У цитоплазмі можуть бути обидва різновиди ендоплазматичної мережі, але зазвичай переважає одна форма, що і зумовлює функціональну специфічність клітини. Слід пам'ятати, що названі два різновиди не є самостійними формами ендоплазматичної мережі, оскільки можна простежити перехід зернистої ендоплазматичної мережі в гладку і навпаки.
Функції зернистої ендоплазматичної мережі:
синтез білків, призначених для виведення із клітини ("на експорт");
відокремлення (сегрегація) синтезованого продукту від гіалоплазми;
конденсація та модифікація синтезованого білка;
транспорт синтезованих продуктів до цистерн пластинчастого комплексу або безпосередньо з клітини;
синтез біліпідних мембран.
Гладка ендоплазматична мережа представлена цистернами, ширшими каналами та окремими везикулами, на зовнішній поверхні яких відсутні рибосоми.
Функції гладкої ендоплазматичної мережі:
участь у синтезі глікогену;
синтез ліпідів;
дезінтоксикаційна функція – нейтралізація токсичних речовин, за допомогою з'єднання їх з іншими речовинами.
Пластинчастий комплекс Гольджі (сітчастий апарат) представлений скупченням сплощених цистерн та невеликих везикул, обмежених біліпідною мембраною. Пластинчастий комплекс поділяється на субодиниці – диктіосоми. Кожна диктіосома є стопкою сплощених цистерн, по периферії яких локалізуються дрібні бульбашки. При цьому у кожній сплощеній цистерні периферична частина дещо розширена, а центральна звужена.
У 1898 р. італійський учений К. Гольджі, використовуючи властивості зв'язування важких металів (осмія і срібла) з клітинними структурами, виявив у нервових клітинах сітчасті освіти, що він назвав “внутрішнім сітчастим апаратом” (рис. 174). Подальше вдосконалення методу фарбування металами (імпрегнації) дало змогу переконатися, що сітчасті структури (апарат Гольджі) зустрічаються у всіх клітинах будь-яких еукаріотних організмів. Зазвичай елементи апарату Гольджі розташовані біля ядра поблизу клітинного центру (центріолі). Ділянки апарату Гольджі, що чітко виявляються методом імпрегнації, мали в деяких клітинах вид складних мереж, де осередки були пов'язані один з одним або представлялися у вигляді окремих темних ділянок, що лежать незалежно одна від одної (диктіосоми), що мають вид паличок, зерен, дисків увігнутих і і т.д. (Рис. 175). Між сітчастою і дифузною формою апарату Гольджі немає принципової відмінності, оскільки часто в тих самих клітинах спостерігається зміна форм цього органоїду. Елементи апарату Гольджі часто пов'язані з вакуолями, що особливо характерно для клітин, що секретують.
Було виявлено, що морфологія АГ змінюється залежно від стадій клітинної секреції, що стало підставою Д.М. Насонову (1924) висунути гіпотезу про те, що АГ є органоїдом, що забезпечує сепарацію та накопичення речовин у різних клітинах.
Довгий час у рослинних клітинах не вдавалося виявити елементів апарату Гольджі звичайними методами мікротехніки. Однак з появою методу електронної мікроскопії елементи АГ були виявлені у всіх рослинних клітинах, де вони розташовані на периферії клітини.
Тонка будівля апарату Гольджі
В електронному мікроскопі видно, що апарат Гольджі представлений мембранними структурами, зібраними разом у невеликій зоні (рис. 176, 177). Окрема зона скупчення цих мембран є диктіосомою(Рис. 178). У диктіосомі щільно один до одного (на відстані 20-25 нм) розташовані у вигляді стоси плоскі мембранні мішки, або цистерни, між якими розташовуються тонкі прошарки гіалоплазми. Кожна окрема цистерна має діаметр близько 1 мкм та змінну товщину; у центрі її мембрани може бути зближені (25 нм), але в периферії мати розширення, ампули, ширина яких непостійна. Кількість таких мішків у стосі зазвичай не перевищує 5-10. У деяких одноклітинних їх кількість може сягати 20 штук. Крім щільно розташованих пласких цистерн у зоні АГ спостерігається безліч вакуолей. Дрібні вакуолі зустрічаються головним чином периферичних ділянках зони АГ; іноді видно, як вони відшнуровуються від ампулярних розширень на краях плоских цистерн. Прийнято розрізняти в зоні диктіосоми проксимальну або формувану, цис- ділянку, і дистальну або зрілу, транс-дільницю (рис. 178). Між ними розташовується середня або проміжна ділянка артеріальної гіпертензії.
Під час поділу клітин сітчасті форми АГ розпадаються до диктіосу, які пасивно і випадково розподіляються по дочірніх клітинах. У разі зростання клітин загальна кількість диктіосом збільшується.
У секретуючих клітинах зазвичай АГ поляризований: його проксимальна частина звернена до цитоплазми та ядра, а дистальна – до поверхні клітини. У проксимальній ділянці до стоси зближених цистерн примикає зона дрібних гладких бульбашок і коротких мембранних цистерн. У зразках препаративно виділених зон АГ при негативному контрастуванні видно, що до проксимальної частини диктіосоми примикає сіткоподібна або губкоподібна система мембранних порожнин. Вважається, що ця система може являти собою зону переходу елементів ЕР в зону апарату Гольджі (рис. 179).
У середній частині диктіосоми периферія кожної цистерни супроводжується масою дрібних вакуолей близько 50 нм в діаметрі.
У дистальному або транс-ділянці диктіосом до останньої мембранної плоскої цистерні примикає ділянку, що складається з трубчастих елементів і масою дрібних вакуолей, часто мають фібрилярну опушеність по поверхні з боку цитоплазми - це опушені або облямовані бульбашки такого ж типу, як і облямовування. Це – так звана транс-мережа апарату Гольджі(TGN), де відбувається поділ і сортування продуктів, що секретуються. Ще дистальніше розташовується група великих вакуолей - це вже продукт злиття дрібних вакуолей і утворення секреторних вакуолей.
При вивченні товстих зрізів клітин у мегавольтний електронний мікроскоп було знайдено, що в клітинах окремі диктосоми можуть бути пов'язані між собою системою вакуолей та цистерн. Так що утворюється пухка тривимірна мережа, що виявляється у світловому мікроскопі. У разі дифузної форми АГ кожна окрема його ділянка представлена диктіосомою. У клітин рослин переважає дифузний тип організації АГ, зазвичай, у середньому на клітину припадає близько 20 диктіосом. У клітинах тварин часто із зоною мембран апарату Гольджі асоційовані центріолі; між радіально відходять від них пучків мікротрубочок лежать групи стосів мембран і вакуолей, які концентрично оточують клітинний центр. Цей зв'язок, ймовірно, відображає участь мікротрубочок у русі вакуолей.
Секреторна функція апарату Гольджі
Мембранні елементи АГ беруть участь у сегрегації та накопиченні продуктів, синтезованих в ЕР, беруть участь у їх хімічних перебудовах, дозріванні: це, головним чином, перебудова олігосахаридних компонентів глікопротеїнів у складі водорозчинних секретів або у складі мембран (рис. 180).
У цистернах АГ відбувається синтез полісахаридів, їхній взаємозв'язок з білками, що призводить до утворення мукопротеїдів. Але головне, з допомогою елементів апарату Гольджі відбувається процес виведення готових секретів межі клітини. Крім того, АГ є джерелом клітинних лізосом.
Участь АГ у процесах виведення секреторних продуктів було дуже добре вивчено на прикладі екзокринних клітин підшлункової залози. Для цих клітин характерна наявність великої кількості секреторних гранул (зимогенових гранул), які є мембранними бульбашками, заповненими білковим вмістом. У складі білків зимогенових гранул входять різноманітні ферменти: протеази, ліпази, карбогідрази, нуклеази. При секреції вміст цих зимогенових гранул викидається з клітин у просвіт залози, а потім перетікає в порожнину кишківника. Так як основним продуктом, що виводиться клітинами підшлункової залози, є білок, то досліджували послідовність включення радіоактивних амінокислот до різних ділянок клітини (рис. 181). Для цього тваринам вводили мічену тритієм амінокислоту (3 Н-лейцин) та за допомогою електронно-мікроскопічної радіоавтографії стежили у часі за локалізацією мітки. Виявилося, що через короткий проміжок часу (3-5 хв) мітка локалізувалася тільки в базальних ділянках клітин, ділянки, багатих гранулярним ЕР. Так як мітка включалася в білковий ланцюг під час синтезу білка, то було ясно, що ні в зоні артеріальної гіпертензії, ні в самих зимогенових гранулах синтез білка не відбувається, а він синтезується виключно в ергастоплазмі на рибосомах. Дещо пізніше (через 20-40 хв) мітка крім ергастоплазми була виявлена в зоні вакуолей АГ. Отже, після синтезу в ергастоплазмі білок транспортувався до зони АГ. Ще пізніше (через 60 хв) мітка виявлялася вже й у зоні зимогенових гранул. Надалі мітку можна було бачити у просвіті ацинусів цієї залози. Таким чином, стало ясно, що АГ є проміжною ланкою між власне синтезом білка, що секретується, і виведенням його з клітини. Також докладно процеси синтезу та виведення білків були вивчені на інших клітинах (молочна залоза, келихоподібні клітини кишечника, щитовидна залоза та ін), та були досліджені морфологічні особливості цього процесу. Синтезований на рибосомах білок, що експортується, відділяється і накопичується всередині цистерн ЕР, по яких він транспортується до зони мембран АГ. Тут від гладких ділянок ЕР відщеплюються дрібні вакуолі, що містять синтезований білок, які надходять у зону вакуолей у проксимальній частині диктіосоми. У цьому місці вакуолі можуть зливатися один з одним і плоскими цис-цистернами диктіосоми. Таким чином, відбувається перенесення білкового продукту вже всередині порожнин цистерн АГ.
У міру модифікації білків у цистернах апарату Гольджі, вони за допомогою дрібних вакуолей переносяться від цистерн до цистерни в дистальну частину диктіосоми, поки не досягають трубчастої мембранної мережі в транс-ділянці диктіосоми. У цій ділянці відбувається відщеплення дрібних бульбашок, які вже містять зрілий продукт. Цитоплазматична поверхня таких бульбашок буває подібна до поверхні облямованих бульбашок, які спостерігаються при рецепторному піноцитозі. Дрібні бульбашки, що відокремилися, зливаються один з одним, утворюючи секреторні вакуолі. Після цього секреторні вакуолі починають рухатися до поверхні клітини, стикаються з плазматичною мембраною, з якою зливаються їх мембрани, і таким чином вміст цих вакуолей виявляється за межами клітини. Морфологічно цей процес екструзії (викидання) нагадує піноцитоз, лише зі зворотною послідовністю стадій. Він має назву екзоцитоз.
Такий опис подій є лише загальною схемою участі апарату Гольджі у секреторних процесах. Справа ускладнюється тим, що одна і та ж клітина може брати участь у синтезі багатьох білків, що виділяються, може їх один від одного ізолювати і направляти до клітинної поверхні або ж до складу лізосом. В апараті Гольджі відбувається не просто "перекачування" продуктів з однієї порожнини в іншу, а й поступово йде їх "дозрівання", модифікація білків, яка закінчується "сортуванням" продуктів, що прямують або до лізосом, або до плазматичної мембрани, або до секреторних вакуолів.
Модифікація білків в апараті Гольджі
У цис-зону апарату Гольджі синтезовані в ЕР білки потрапляють після первинного глікозилювання та редукції там же кількох сахаридних залишків. Зрештою, всі білки там мають однакові олігосахаридні ланцюги, що складаються з двох молекул N-ацетилглюкозаміну, шести молекул маннози (рис. 182). У цис-цистернах починається вторинна модифікація олігосахаридних ланцюгів та їх сортування на два класи. В результаті олігосахариди на гідролітичних ферментах, призначених для лізосом (багаті на манозу олгосахариди), фосфорилюються, а олігосахариди інших білків, що направляються в секреторні гранули, або до плазматичної мембрани, піддаються складним перетворенням і втрачають ряд цукрів. алові кислоти .
При цьому виникає спеціальний комплекс олігосахаридів. Такі перетворення олігосахаридів здійснюються за допомогою ферментів – глікозилтрансфераз, що входять до складу мембран цистерн апарату Гольджі. Так як кожна зона в диктіосомах має свій набір ферментів глікозилювання, то глікопротеїди ніби по естафеті переносяться з одного мембранного відсіку ("поверху" в стосі цистерн диктіосоми) в інший і в кожному піддаються специфічному впливу ферментів. Так у цис-ділянці відбувається фосфорилювання маноз у лізосомних ферментах і утворюється особлива маннозо-6-угруповання, характерна для всіх гідролітичних ферментів, які потім потраплять у лізосоми.
У середній частині диктіос протікає вторинне глікозилювання секреторних білків: додаткове видалення маннози і приєднання N-ацетилглюкозаміну. У транс-ділянці до олігосахаридного ланцюга приєднуються галактоза та сіалові кислоти (рис. 183).
Ці дані були отримані різними методами. За допомогою диференціального центрифугування вдалося отримати роздільні більш важкі (цис-) компоненти апарату Гольджі та легші (транс-) та визначити в них наявність глікозидаз та їх продуктів. З іншого боку, використовуючи моноклональні антитіла до різних ферментів за допомогою електронної мікроскопії, вдалося їх локалізувати прямо на зрізах клітин.
У ряді спеціалізованих клітин в апараті Гольджі відбувається синтез власне полісахаридів.
В апараті Гольджі рослинних клітин відбувається синтез полісахаридів матриксу клітинної стінки (геміцелюлози, пектини). Крім того, диктіосоми рослинних клітин беруть участь у синтезі та виділенні слизів та муцинів, до складу яких входять також полісахариди. Синтез ж основного каркасного полісахариду рослинних клітинних стінок, целюлози, відбувається, як говорилося, на поверхні плазматичної мембрани.
В апараті Гольджі клітин тварин відбувається синтез довгих нерозгалужених полісахаридних ланцюгів глюкозаїногліканів. Один з них, гіалуронова кислота, що входить до складу позаклітинного матриксу сполучної тканини, містить кілька тисяч дисахаридних блоків, що повторюються. Багато глюкозаїноглікан ковалентно пов'язані з білками і утворюють протеоглікани (мукопротеїни). Такі полісахаридні ланцюги модифікуються в апараті Гольджі та зв'язуються з білками, які у вигляді протеогліканів секретуються клітинами. В апараті Гольджі відбувається також сульфатування глюкозаїногліканів та деяких білків.
Сортування білків в апараті Гольджі
Отже, через апарат Гольджі проходить принаймні три потоки синтезованих клітиною нецитозольних білків: потік гідролітичних ферментів в компартмент лізосом, потік білків, що накопичуються в секреторних вакуолях, і виділяються з клітини тільки після отримання спеціальних сигналів, потік секреторних білків, що постійно виділяються. Отже, має бути якийсь спеціальний механізм просторового поділу цих різних білків та їх шляхів прямування.
У цис- та середніх зонах диктіосом всі ці білки йдуть разом без поділу, вони лише окремо модифікуються залежно від їх олігосахаридних маркерів.
Власне поділ білків, їх сортування відбувається в транс-ділянці апарату Гольджі. Цей процес не до кінця розшифрований, але на прикладі сортування лізосомних ферментів можна зрозуміти принцип відбору певних білкових молекул (рис. 184).
Відомо, що тільки білки-попередники лізосомних гідролаз мають специфічну олігосахаридну, а саме маннозну групу. У цис-цистернах ці угруповання фосфорилируются і далі разом з іншими білками переносяться від цистерни до цистерни, через середню зону транс-ділянку. Мембрани транс-мережі апарату Гольджі містять трансмембранний білок - рецептор (манноза-6-фосфатний рецептор або М-6-Ф-рецептор), який дізнається про фосфорильовані манозні угруповання олігосахаридного ланцюга лізосомних ферментів і зв'язується з ними. Це зв'язування відбувається при нейтральних значеннях рН усередині цистерн транс-мережі. На мембранах ці М-6-Ф-рецепторні білки утворюють кластери, групи, які концентруються у зонах утворення дрібних бульбашок, покритих клатрином. У транс-мережі апарату Гольджі відбувається їх відділення, відбрунькування та подальше перенесення до ендосом. Отже М-6-Ф-рецептори, будучи трансмембранними білками, зв'язуючись з лізосомними гідролазами, відокремлюють їх, відсортують від інших білків (наприклад, секреторних, нелізосомних) і концентрують їх в облямованих бульбашках. Відірвавшись від транс-мережі ці бульбашки швидко втрачають шубу, зливаються з ендосомами, переносячи свої лізосомні ферменти, пов'язані з мембранними рецепторами, у цю вакуолю. Як мовилося раніше, всередині ендосом через активності протонного переносника відбувається закислення середовища. Починаючи з рН 6, лізосомні ферменти дисоціюють від М-6-Ф-рецепторів, активуються і починають працювати в порожнині ендолізосоми. Ділянки мембран разом з М-6-Ф-рецепторами повертаються шляхом рециклізації мембранних бульбашок назад в транс-мережу апарату Гольджі.
Найімовірніше, що та частина білків, яка накопичується в секреторних вакуолях і виводиться з клітини після надходження сигналу (наприклад, нервового або гормонального) проходить таку ж процедуру відбору, сортування на рецепторах транс-цистерн апарату Гольджі. Ці секреторні білки потрапляють спочатку у дрібні вакуолі теж одягнені клатрином, які потім зливаються один з одним. У секреторних вакуолях часто відбувається агрегація накопичених білків як щільних секреторних гранул. Це призводить до підвищення концентрації білка в цих вакуолях приблизно в 200 разів порівняно з його концентрацією в апараті Гольджі. Потім ці білки в міру накопичення секреторних вакуолях викидаються з клітини шляхом екзоцитозу, поле отримання клітиною відповідного сигналу.
Від апарату Гольджі виходить і третій потік вакуолей, пов'язаний із постійною конститутивною секрецією. Так фібробласти виділяють велику кількість глікопротеїдів та муцинів, що входять в основну речовину сполучної тканини. Багато клітин постійно виділяють білки, що сприяють зв'язування їх з субстратами, постійно йде потік мембранних бульбашок до поверхні клітини, що несуть елементи глікоколіксу і мембранних глікопротеїдів. Цей потік компонентів, що виділяються клітиною, не підлягає сортуванню в рецепторній транс-системі апарату Гольджі. Первинні вакуолі цього потоку також відщеплюються від мембран і відносяться за своєю структурою до облямованих вакуолей, що містять клатрин (рис. 185).
Закінчуючи розгляд будови і роботи такої складної мембранної органели, як апарат Гольджі, необхідно підкреслити, що незважаючи на морфологічну однорідність його компонентів, вакуолі і цистерни, насправді, це не просто скупчення бульбашок, а струнка, динамічна складно організована, поляризована система.
В АГ відбувається не лише транспорт везикул від ЕР до плазматичної мембрани. Існує ретроградне перенесення везикул. Так від вторинних лізосом відщеплюються вакуолі та повертаються разом із рецепторними білками в транс-АГ зону. Крім того, існує потік вакуолей від транс-зони до цис-зони АГ, а також від цис-зони до ендоплазматичного ретикулуму. У цих випадках вакуолі одягнені білками COP I-комплексу. Вважається, що таким шляхом повертаються різні ферменти вторинного глікозилювання та рецепторні білки у складі мембран.
Ці особливості поведінки транспортних везикул дали основу гіпотезі існування двох типів транспорту компонентів АГ (рис. 186).
По одному з них, найстарішому, в артеріальній гіпертензії існують стабільні мембранні компоненти, до яких від ЕР естафетно переносяться речовини за допомогою транспортних вакуолей. По альтернативній моделі АГ є динамічним похідним ЕР: мембранні вакуолі, що відщепилися від ЕР, зливаються один з одним у нову цис-цистерну, яка потім просувається через всю зону АГ і в кінці розпадається на транспортні везикули. За цією моделлю ретроградні COP I везикули повертають постійні білки АГ у молодші цистерни. Таким чином передбачається, що перехідна зона ЕР є "пологовим домом" для апарату Гольджі.
А-гранулярної цитоплазматичної мережі.
Б- Мікропузирків.
В-мікрофіламентів.
Г-Цистерн.
Д-вакуолей.
Відповідь: Б, Г, Д.
16. Вказати, які функції виконує комплекс Гольджі:
А-синтез білка.
Б- Утворення комплексних хімічних сполук (глікопротеїдів, ліпопротеїдів).
В- Освіта первинних лізосом.
Г- Участь у виведенні із клітини секреторного продукту.
Д-Освіта гіалоплазми.
Відповідь: Б,В,Г.
Які структурні елементи клітини найактивніше беруть участь в екзоцитозі?
А-Цитолемма.
Б-Цитоскелет.
В-Мітохондрії.
Г-Рібосоми.
Відповідь: А,Б.
18 . Що визначає специфічність синтезованого білка?
А-Інформаційна РНК.
Б-рибосомна РНК.
Г-мембрани цитоплазматичної мережі.
Відповідь: А,В
19 . Які структурні елементи беруть активну участь у виконанні
Фагоцитарні функції?
А-Каріолемма.
Б-Ендоплазматична мережа.
В-Цитолемма.
Г-Лізосоми.
Д-Мікрофіламенти.
Відповідь: В,Г,Д.
20 .Які структурні компоненти клітини зумовлюють базофілію цитоплазми?
А-Рібосоми.
Б-Агранулярна ендоплазматична мережа.
В-Лізосоми.
Г-пероксисоми.
Д-комплекс Гольджі.
Е-гранулярна ендоплазматична мережа.
Відповідь: А,Е.
21 . Які з перерахованих органел мають мембранну будову?
А-Клітинний центр.
Б-Мітохондрії.
В- Комплекс Гольджі.
Г-Рібосоми.
Д-цитоскелет.
Відповідь: Б,В.
22 .Що спільного між мітохондріями та пероксисомами?
А- Належать до органоїдів мембранної будови.
Б-мають подвійну мембрану.
Д- Це органели загального значення.
Відповідь: А,В,Д.
Які функції у клітині виконують лізосоми?
А-Біосинтез білка
Б- Участь у фагоцитозі
В- Окисне фосфорилювання
Г-Внутрішньоклітинне травлення
Відповідь: Б.Г.
Яка структурна організація лізосом?
А-Оточені мембраною.
В- Заповнені гідролітичними ферментами.
Г- утворюються в комплексі Гольджі.
Відповідь: А,В,Г.
25. Глікокалікс:
А- Знаходиться в гладкій ендоплазматичній мережі.
Б-знаходиться на зовнішній поверхні цитолеми.
В- утворений вуглеводами.
Г-Бере участь у клітинній адгезії та клітинному впізнанні.
Д- знаходиться на внутрішній поверхні цитолеми.
Відповідь: Б,В,Г.
26. Маркерні ферменти лізосом:
А-Кисла фосфатаза.
Б-АТФ-аза.
В-Гідролази.
Г- Каталаза та оксидази.
Відповідь: А,В.
Яке значення ядра у життєдіяльності клітини?
А-Зберігання спадкової інформації.
Б-Центр накопичення енергії.
В- Центр управління внутрішньоклітинним метаболізмом.
Г- Місце утворення лізосом.
Д-Відтворення та передача генетичної інформації дочірнім клітинам.
Відповідь: А,В,Д.
28. Що не стосується структурних компонентів ядра:
А-Каріолемма.
Б-Ядрішки.
В-Каріоплазма.
Г-Рібосоми.
Д-хроматин, хромосоми.
Е-пероксисоми.
Відповідь: Г,Є.
Що транспортується з ядра через ядерні пори до цитоплазми?
А-Фрагменти ДНК.
Б-субодиниці рибосом.
В-Інформаційні РНК.
Г-Фрагменти ендоплазматичної мережі.
Відповідь: Б,В.
Що таке ядерно-цитоплазматичне відношення і як воно змінюється у разі підвищення функціональної активності клітини?
А-Положення ядра в цитоплазмі.
Б-форма ядра.
В- Відношення розміру ядра до розміру цитоплазми.
Г- Знижено при підвищеній функціональній активності клітини.
Відповідь: В,Г.
Що правильно для ядерців?
А- Добре видно під час мітозу.
Б- Складаються з гранулярного та фібрилярного компонентів.
В-Гранули ядерця - субодиниці рибосом.
Г- Нитки ядерця - рибонуклеопротеїди
Відповідь: Б,В,Г.
Які перелічені ознаки належать до некрозу?
А- Це генетично запрограмована загибель клітини
Б- На початку апоптозу синтез РНК та білка зростає.
В-руйнуються мембрани
Г-ферменти лізосом виходять у цитоплазму
Д-фрагментація цитоплазми з утворенням апоптичних тіл
Відповідь: В,Г.
Правильно все, крім
1.Функція комплексу Гольджі (вірно все, крім):
А- сортування білків за транспортними бульбашками
Б-глікозилювання білків
В-реутилізація мембран секреторних гранул після екзоцитозу
Г-упаковка секреторного продукту
Д-синтез стероїдних гормонів
2. Мікротрубочки забезпечують (вірно все, крім):
А-організацію внутрішнього простору клітини
Б-підтримка форми клітини
В-поляризацію клітини при розподілі
Г-формують скорочувальний апарат
Д-організацію цитоскелета
Е-транспорт органел
3. До спеціалізованих структур, побудованих на основі цитоскелета, відносяться (вірно все, крім):
А- війки, джгутики
Б- базальна смугастість
В-мікроворсинки
4. Локалізація вій (вірно все, крім):
А- епітелій слизової оболонки повітроносних шляхів
Б-епітелій проксимального відділу нефрону
В- епітелій слизової оболонки репродуктивного тракту жінок
Г- епітелій слизової оболонки сім'явиносних шляхів
5. Локалізація мікроворсинок (вірно все, крім):
А- епітелій слизової оболонки тонкої кишки
Б- епітелій слизової оболонки трахеї
В- епітелій проксимального відділу нефрону
6. Базальна смугастість (вірно все, крім):
А- забезпечує транспорт речовин проти градієнта концентрації
Б- ділянка клітини, де йдуть високоенергоємні процеси
В- ділянку клітини, де відбувається проста дифузія іонів
Г-де відбувається реабсорбція елементів первинної сечі в проксимальному канальці нефрону.
Д- бере участь у концентрації слинного секрету
7. Щіточна облямівка (вірно все, крім):
А-розташовується на апікальній поверхні клітин
Б- збільшує площу всмоктувальної поверхні
В- складається з вій
Г- складається з мікроворсинок
Д-збільшує транспортну поверхню в проксимальних канальцях нефрону
8. Органоїди загального призначення (вірно все, крім):
А-мітохондрії
В-комплекс Гольджі
Г-віски
Д-лізосоми
Е-пероксисоми
Ж-центріолі
З-елементи цитоскелета
9.Функція пероксисом (вірно все, крім):
А-окислення органічного субстрату з утворенням перекису водню
Б-синтез ферменту - каталази
В-утилізація перекису водню
10. Рибосоми (вірно все, крім):
А-при світловій мікроскопії про їх наявність судять по вираженій базофілії цитоплазми
Б- складаються з малої та великої субодиниць
В-утворюються в гранулярній ЕПС
Г- складаються з рРНК та білків
Д-немембранної будови
11.Які органоїди добре розвинені в стероїдпродукуючих клітинах (вірно все, крім):
А-гранулярна ендоплазматична мережа
Б-агранулярна ендоплазматична мережа
В- мітохондрії з трубчастими христами
12.Трофічні включення (вірно все, крім):
А-вуглеводні
Б-слизові
В-білкові
Г-ліпідні
13.Ядерна оболонка (вірно все, крім):
А- складається з одинарної мембрани
Б складається з двох мембран
В-зовні на ній розташовані рибосоми
Г- зсередини з нею пов'язана ядерна платівка
Д- пронизана порами
14. Структурні компоненти ядра (вірно все, крім):
А-нуклеоплазма
Б-нуклеолема
В-мікротрубочки
Г-хроматин
Д-ядра
15. Будова ядерної доби (вірно все, крім):
А-мембранний компонент
Б-хромосомний компонент
В-фібрилярний компонент
Г-гранулярний компонент
16. Ядрішко (вірно все, крім):
А- оточене мембраною
Б-не оточене мембраною
У його організації беруть участь п'ять пар хромосом
Г містить гранулярний і фібрилярний компонент
17. Ядрішко (вірно все, крім):
А-кількість залежить від метаболічної активності клітини
Б- бере участь в утворенні субодиниць рибосом
В- в організації беруть участь 13,14, 15, 21 і 22 хромосоми
Г- в організації беруть участь 7, 8, 10, 11 і 23 хромосоми
Д- складається з трьох компонентів
18. Клітинний центр (вірно все, крім):
А- локалізується поблизу ядра
Б є центром організації веретена поділу
В- складається з двох центріолей
Г-центріолі утворені 9 дуплетами мікротрубочок
Д-центріолі дуплікуються в S періоді інтерфази
19. Мітохондрії (вірно все, крім):
А- наявність христ
Б-здатність ділитися
20. Функції актинових філаментів (вірно все, крім):
А-рух клітини
Б-зміна форми клітини
В- участь в екзо- та ендоцитозі
Г-забезпечують рух вій
Д-входять до складу мікроворсинок
21. Для ядерця вірно все, крім:
А- утворюються в області ядерцевих організаторів (вторинних перетяжок хромосом)
Б-Гранули ядерців виходять у цитоплазму
В-Білки ядерців синтезуються в цитоплазмі
Г-Ядерцева РНК утворюється в цитоплазмі
На відповідність
1. Зіставте періоди інтерфази з процесами, що відбуваються в них:
1. Пресинтетичний А-подвоєння ДНК, збільшення синтезу РНК
2. Синтетичний Б-синтез рРНК, іРНК, тубулінів
3. Постсинтетичний В- зростання клітин, підготовка їх до синтезу ДНК
Відповідь: 1-В; 2-А; 3-Б.
2 .Зіставте фази мітозу з процесами, що відбуваються в них:
1. Профаза А-освіта екваторіальної платівки з хромосом
2. Метафаза Б-освіта нуклеолеми, деспіралізація хромосом,
утворення ядерця, цитотомія
3. Анафаза В-спіралізація хромосом, зникнення ядерця,
фрагментація нуклеолеми
4. Телофаза Г-розбіжність хроматид до протилежних полюсів
Відповідь: 1-В; 2-А; 3-Г; 4-Б.
3. Зміна структури ядра називається (порівняйте):
1.каріолізис А- зменшення розмірів та ущільнення хроматину
2.каріорексіс Б-фрагментація
3.каріопікноз В-розчинення його компонентів
Відповідь: 1-В, 2-Б, 3-А.
4. Характеристика компонентів препарату:
1.хромофобний А-фарбується барвником суданом
2.хромофільний Б-не фарбується барвником
3.суданофільний В-фарбується барвником
 Приватні похідні функції трьох змінних
Приватні похідні функції трьох змінних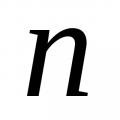 Емпірична функція розподілу За цією вибіркою побудувати емпіричну функцію розподілу
Емпірична функція розподілу За цією вибіркою побудувати емпіричну функцію розподілу Грошей немає, але ви тримаєтеся
Грошей немає, але ви тримаєтеся