Кембрійський еволюційний вибух. Відкладення кембрійського періоду
Зміст) поява в ранньокембрійських (бл. 540 млн років за еволюційною шкалою) відкладах скам'янілостей представників багатьох підрозділів тваринного царства, на тлі відсутності їх скам'янілостей або скам'янілостей їхніх передбачуваних предків у докембрійських відкладеннях.
Теорія еволюції немає достовірного пояснення цьому «феномену». Теорія креаціонізму бачить у цьому явищі чергове свідчення неспроможності теорії еволюції та свідчення на користь Створення.
Історія відкриття
Першовідкривачем Кембрійського вибуху був англієць Роберт Мерчисон, аристократ за походженням, який під впливом своєї честолюбної дружини вирішив зайнятися наукою. Вивчаючи скам'янілості багатоклітинних стародавніх епох, і верстви порід, в яких вони знаходилися, він зіткнувся з чітко вираженим кордоном, нижче якого були сліди лише найпростіших. одноклітинних організмів- бактерій та водоростей. А шаром вище (у «кембрійських» відкладах) – найбагатша різноманітність біологічних форм. Мерчисон був віруючим християнином і поділяв переконання Ліннея, що «існує рівно стільки видів, скільки їх спочатку створив Творець». У відкритому явленні він побачив доказ дії Божої руки. У 1830-х роках Мерчисон опублікував результати своїх досліджень.
Кембрійський вибух та дарвінізм
Останні відкриття
Упродовж останніх ста років вважалося, що хребетні з'явилися в історії життя пізніше. Однак, у 1999 році в кембрійських породах у Китаї були виявлені скам'янілості риб. що підтверджує, що риби виникають раптово в літописі скам'янілостей разом з іншими типами тварин. Кембрійський вибух став ще гучнішим. Щоб ці повністю сформовані риби з'явилися в Кембрії, передбачуваний предок хребетних має бути відсунутий на мільйони років у докембрійський період, де немає перехідних форм ні для них, ні для всіх основних типів організмів.
У 2006 році в Китаї геологи виявили скам'янілий ембріон тварини, якій, на думку еволюціоністів, 600 мільйонів років. Якщо еволюція справді відбувалася, то сучасні ембріони через сотні мільйонів років еволюції мали б сильно відрізнятися від тих, що були виявлені в Китаї. Проте ембріони, знайдені у Китаї, повністю ідентичні ембріонам сучасних тварин.
У 2008 році в Кембрійських пластах були виявлені викопні медузи, які майже ідентичні сучасним живим видам. Деякі з них мали м'язи, ряд кліток, складні статеві органи та види поведінки (включаючи впізнавання пари та догляд), а також складні очі. Згідно з еволюцією, поки ці медузи так і залишилися такими ж медузами до цього дня, за цей час (близько 500 млн років) мала еволюціонувати практично вся природа - птахи, сосни, крокодили, кенгуру, слони, собаки, квіти, помідори, кити. гекони і т.д.
Див. також
Посилання
| |||||||||||||||||||||||
Ястребов С.А.
(«Хиж», 2016, №10)
Вендська прелюдія
Епоха "Землі-сніжка" закінчилася 635 мільйонів років тому. Почався останній період протерозою - едіакарій (635-542 мільйонів років тому). Тепер нам буде зручніше вести рахунок часу не на мільярди років, а на мільйони - це показує, наскільки події прискорюються. Хоча, можливо, справа просто в тому, що вони до нас ближчі і від них збереглося більше слідів. Раніше едіакарій називали вендом, на честь древніх слов'янських племен - венедів (від них відбулася назва міста Венеція). На жаль, зараз ця гарна назва збереглася лише як не суворий синонім.
Головною подією едіакарію (не можна не додати: з нашої антропоцентричної точки зору) слід назвати появу багатоклітинних тварин. Датувати цю подію нелегко. У палеонтологічному літописі едіакарія вистачає свідчень переходу до багатоклітинності тваринного типу - правда, що вони більш ранні, тим більше спірні («Nature», 2014, 516, 7530, 238-241, див. також статтю Олександра Маркова, http://elementy). ru/novosti_nauki/431720). У другій половині едіакарію удосталь з'являються вендобіонти - великі, до метра довжиною загадкові істоти з плоским дископодібним або листоподібним тілом, що складається з безлічі однотипних «сегментів», що повторюються. «Сегменти» тут поставлені у лапки, бо сегментація вендобіонтів майже напевно не має нічого спільного із сегментацією справжніх багатоклітинних тварин. Сам термін «вендобіонти» вигадав німецький палеонтолог Адольф Зейлахер, який вважав цих істот абсолютно особливою формою життя – гігантськими багатоядерними клітинами («Planetary Systems and the Origins of Life», Cambridge University Press, 2007, 193-209). Дійсно, є підстави вважати, що вендобіонти були в багатьох відношеннях ближчі не до багатоклітинних тварин, а до амеб або грибів (між іншим, і в тих і в інших великі багатоядерні клітини не рідкість). Вони зробили спробу виходу у великий розмір, яка спочатку призвела до успіху, але завершилася невдачею: наприкінці едіакарію вимерли вендобіонти.
З іншого боку, треба враховувати, що вендобіонти були дуже різноманітні. Не факт, що їх взагалі можна хоч у якомусь наближенні вважати єдиною групою. Це скоріше еволюційний рівень. І незважаючи на те, що більшість вендобіонтів ніяких нащадків не залишила, від деяких з них цілком могли безпосередньо відбутися сучасні тварини - наприклад, пластинчасті та гребневики (Evolution and Development, 2011, 13, 5, 408-414). У едіакарському корінні цих еволюційних гілок немає нічого неймовірного.
Найдавніша абсолютно безперечна викопна багатоклітинна тварина називається Kimberella quadrata. Це двосторонньо-симетрична істота довжиною до 15 сантиметрів, що повзала морським дном. Характер змін форми тіла знайдених кімберелл (а знайдено їх багато, у різних частинах світу) разом із відбитками слідів не залишає сумнівів, що вони активно повзали, розтягуючись, стискаючись та згинаючи за допомогою м'язів. Характерні ознакикімберели - витягнуте, але компактне тіло з ногою (м'язистою нижньою поверхнею) і мантією (складкою, що облямовує тулуб). За цими ознаками вона дуже схожа не на когось, а на молюсків («Paleontological Journal», 2009, 43, 601, doi: 10.1134/S003103010906001X). Є думка, що у кімберели була навіть радула - властива молюскам «мова» з хітиновими зубцями, пристосована для зіскребання водоростей (PALAIOS, 2010, 25, 565-575, doi: 10.2110/palo.2009.p09-0). Так чи інакше, це вже за всіма статтями справжня багатоклітинна тварина.
Кімберелла жила 555 мільйонів років тому (Science, 2000, 288, 5467, 841-845). І приблизно в цей же час вперше з'являються численні викопні сліди тварин, які явно активно повзали по дну (Philosophical Transactions of the Royal Society B, 2008, 363, 1496, doi: 10.1098/rstb.2007.2232). Слід зазначити, що «справжні багатоклітинні тварини» - дуже строгий термін; тут достатньо домовитися, що ми називаємо так тварин з м'язами, ротом та кишкою. У вендобіонтів, наскільки можна судити, нічого цього не було. Вони харчувалися в кращому разі мікроскопічними водоростями, а швидше за все - просто речовинами, розчиненими в морській воді (Trends in Ecology & Evolution, 2009, 24, 1, 31-40). Тільки наприкінці едіакарію з'явилися багатоклітинні істоти, здатні активно відшукувати видобуток і захоплювати його великими шматками, щоб переварити всередині. Вендобіонти були перед такими чудовиськами беззахисні - не дивно, що їхнє «золоте століття» на цьому закінчилося. В історії донних угруповань почалася зовсім інша епоха.
«Великий вибух життя»

Кінець едіакарського періоду є в той же час кордоном двох еонів - протерозою та фанерозою; і ось тут потрібне невелике пояснення. "Фанерозою" буквально означає "явне життя". Це епоха, до якої належить переважна більшість викопних залишків, що вивчаються палеонтологами. Усі попередні часи, включаючи протерозою, архей і катархей, збірно називають криптозоєм – «прихованим життям». Фанерозою ж, у свою чергу, ділиться на три ери, назви яких більшості з нас швидше за все знайомі: палеозою, мезозою та кайнозою. "Палеозою" означає "давнє життя", "мезозою" - "середнє життя", "кайнозою" - "нове життя". Кожна з цих ер поділяється на періоди. Період, з якого починається палеозою (і цим весь фанерозою), називається кембрієм. Як і багато інших геологічних періодів, кембрій отримав своє ім'я з географії: Кембрія - це римська назва Уельсу, кельтської країни на заході Британії. Відповідно дуже поширений синонім криптозою – докембрій.
Щоб правильно бачити перспективу, пам'ятатимемо наступне: весь фанерозою становить (округлено) лише приблизно 1/9 як від часу існування Землі, так і від історії життя на ній. Інші 8/9 - це докембрій. Інша річ, що у фанерозої події дуже згущуються.
У 1845 році великий шотландський геолог Родерік Мурчисон запропонував назвати всі часи до початку кембрію азойською ерою, тобто - буквально - неживою. Довго ця назва не протрималася: вже палеонтологи XIX століття показали, що в товщі докембрійських порід є сліди життя (The Journal of Geology, 1927, 35, 8, 734-742). А зараз ми точно знаємо, що життя було на Землі протягом більшої частини докембрію, і можемо датувати багато докембрійських ключових подій – наприклад, кисневу революцію чи появу багатоклітинності.
Головна відмінність фанерозойського життя від докембрійського - колосальна велика кількість багатоклітинних тварин, переважна більшість яких належить вже до сучасних типів. У кембрії з'являються губки, гребневики, кишковопорожнинні, що стікають, всілякі черв'яки, членистоногі, молюски, брахіоподи, голкошкірі, напівхордові і хордові. Раптова поява цих тварин у палеонтологічному літописі прийнято називати кембрійським вибухом. У більш давніх шарах їх залишків немає (принаймні визначених однозначно і безперечно). Кембрій – це час народження фауни, близької до сучасної. Кембрійський вибух дав такий ефект і стався так швидко, що його часто називають «еволюційним Великим вибухом» - за аналогією з тим самим Великим вибухом, в якому народився Всесвіт.
Ще кембрійський вибух іноді називають «скелетною революцією». Дійсно, багато груп тварин, що з'явилися в цей момент, придбали тверді скелети, причому зовсім різні і на різній основі: наприклад, між спікулами губок, раковинами молюсків і хітиновими панцирями членистоногих немає буквально нічого спільного. Така одночасність не може бути випадковою. Проте «кембрійський вибух» та «скелетна революція» не синоніми. По-перше, тверді скелети були далеко не у всіх кембрійських тварин (наприклад, у перших хордових їх не було). По-друге, і в докембрії іноді трапляються явні скелетні структури - наприклад, комусь належали житлові трубки («Природа», 2006, 2, 37-40). Загалом поняття «кембрійський вибух» – куди більш визначене, і не дивно, що сучасні автори найчастіше говорять саме про нього.
А чи був вибух?
Але питання: чи був кембрійський вибух насправді? Існує думка, що багато сучасних груп тварин з'явилися ще в глибокому докембрії, проте довгий час майже не залишали викопних залишків, а тому були палеонтологічно «невидимі» (Science, 2011, 334, 6059, 1091-1097, doi: 10). .1206375). Причини цього могли бути різними: маленький розмір тварин, відсутність у них твердих скелетів або фізичні умови, що просто не підходять для поховання. Гіпотеза «довгої прихованої докембрійської еволюції» непогано підтримується молекулярною систематикою, тобто порівнянням амінокислотних та нуклеотидних послідовностей білків та генів різних тварин (звичайно, сучасних – з часів кембрію ні білків, ні ДНК не залишилося). Реконструкції, зроблені виключно за молекулярними даними, часто ведуть коріння сучасних типів тварин навіть не в едіакарій, а в попередній період - кріогеній (Systematic Biology, 2013, 62, 1, 93-109). Тоді виходить, що кембрійський вибух - не так еволюційна подія, як артефакт безпеки. На рубежі кембрія еволюційні гілки тварин просто «проявилися», знайшовши тверді скелети і почавши захоронюватися в осадових товщах; а виникли вони набагато раніше.
Однак ретельної перевірки, коли молекулярно-біологічні дані крок за кроком об'єктивно зіставляються з палеонтологічними, гіпотеза «довгої прихованої докембрійської еволюції» не витримує (Current Biology, 2013, 23, 19, 1889-1895). І виходить, що кембрійський вибух – ніякий не артефакт. Більшість великих еволюційних гілок тварин справді виникла в найближчій околиці кордону кембрія (плюс-мінус кілька мільйонів років). Є та математичні моделі, що підтверджують, що занурені в докембрій «стволи» еволюційних дерев сучасних типів тварин мають бути короткими («Philosophical Transactions of the Royal Society B», 2016, 371, 1685, doi: 10.1098/rstb.2015). Час їх існування - лічені мільйони років, можливо, перші десятки мільйонів, але аж ніяк не сотні. Загалом, на Наразіу нас вистачає підстав, щоб вважати гіпотезу «довгою прихованою докембрійською еволюцією» швидше невірною, а кембрійський вибух - реальністю, як це, власне кажучи, і випливає безпосередньо з палеонтологічних даних.
Щоб послабити категоричність, додамо: висновок, який ми щойно зробили, безумовно, має властивість фальсифікованості. Це означає, що можна сформулювати чіткі умови, за яких його спростують. Наприклад, для цього цілком достатньо знайти хоча б одного скорпіона (або сороконіжку, або равлика) кріогенієвого віку. Але поки що такого не було, і ймовірність, що це колись станеться, з кожним роком зменшується.
Причини вибуху
Отже, на початку кембрію унікально швидко виникло багато нових великих еволюційних гілок тварин. Такого більше не траплялося ніколи, ні раніше, ні пізніше. Навіть після катастрофічних масових вимирань (про які мова попереду) тваринний світ відновлювався за рахунок наростання різноманітності великих груп, що вже існували, а не за рахунок появи нових. Саме тому кембрійський вибух обов'язково вимагає пояснення.
Щоправда, «швидко» все ж таки не означає «моментально». Нові групи тварин аж ніяк не з'являються разом у повному складі, як актори після підняття завіси. Кембрійський вибух був хоч і сильно стислим у часі, але все ж таки поступовим; швидкість еволюційних процесів у ньому можна виміряти, і такі дослідження є. Кембрій тривав приблизно 57 мільйонів років (542-485 млн років тому), причому на самому його початку (перші шість мільйонів років) морська фауна ще досить бідна. Нові групи тварин з'являються там справді швидко за мірками історії Землі, але не миттєво.
З чим це було пов'язано? За півтора століття, що минули відтоді, як вчені (і зокрема Чарльз Дарвін) усвідомили загадку кембрійського вибуху, пропонувалися різноманітні пояснення цієї події, від генетичних до космічних. Одна сучасна оглядова стаття на цю тему так і називається - "По той бік кембрійського вибуху: від галактики до геному" ("Gondwana Research", 2014, 25, 3, 881-883, doi: 10.1016/j.gr.2014.01. ). Наприклад, тенденція до масової освіти мінеральних скелетів - знаменита "скелетна революція", вона ж "біомінералізація" - на початку кембрія охопила не тільки різних багатоклітинних тварин, але і одноклітинних еукаріотів, і деякі водорості. Напрошується припущення, що це пов'язано із глобальною зміною хімічного складу зовнішнього середовищатобто в даному випадку морської води. І справді, показано, що на початку кембрію з якихось чисто геологічним причин приблизно втричі зросла концентрація в морській воді кальцію (Ca2+) - іона, який потрібен для створення твердих скелетів як жодної іншої (Geology, 2004, 32, 6, 473-476). Мінеральна основа скелетів тварин – найчастіше карбонат кальцію (раковини молюсків, голки та філіжанки коралових поліпів, спікули губок), а іноді фосфат кальцію (кістки хребетних).
Проблема в тому, що пояснити скелетну революцію не означає пояснити сам кембрійський вибух. Скелетна революція лише забезпечила твердими мінералізованими тканинами ряд тварин, які вже існували на момент її початку. Причому це навіть не всіх їх торкнулося. У тих кембрійських місцезнаходженнях, тип безпеки яких допускає поховання безскелетних істот, відразу ж виявляється, що чимала частина кембрійської фауни була «м'якотілою». Отже, справа не в скелетах. Явище, яке треба пояснити в першу чергу, - це унікальне прискорення еволюції багатоклітинних тварин, що дуже швидко (наприкінці едіакарію - початку кембрію) створило багато нових великих груп, не важливо, скелетних чи ні.
Надалі розповіді ми виходитимемо зі сценарію, який коротко виклав ще на початку 1970-х років американський палеонтолог Стівен Стенлі. Звичайно, палеонтологія - наука, що дуже швидко розвивається; роботи сорокалітньої давності в ній завжди вимагають поправок, і ми ці поправки під час розмови внесемо. Щоправда, насправді це будуть скоріше доповнення. Головна ідея Стенлі винятково добре витримала перевірку часом. Сума фактів, відомих зараз, чудово в неї вписується.
Почнемо спочатку. У дужках зауважимо: вирішити, що саме варто прийняти за «початок», при розборі будь-якого історичного процесу- Завдання непросте, адже причинно-наслідкові ланцюжки можуть тягтися в минуле чи не до нескінченності, збиваючи необережного дослідника з пантелику. У нашому випадку "початком" буде едіакарська біота. Що вона собою являла?
В екології прийнято виділяти організми-средоутворювачі, активність яких визначає структуру цілих угруповань. Такі організми називаються едифікаторами.
Наприклад, в сучасній діброві едифікатором є дуб, в маленькому тихому ставку їм цілком може бути ряска і т. д. 1, 86-93, doi: 10.2307/3515363). На цих «килимах» жили вже знайомі нам вендобіонти. Більшість із них вела прикріплений спосіб життя; як вони харчувалися - не зовсім зрозуміло, але швидше за все - обзорно, всмоктуючи з води розчинені речовини всією поверхнею тіла. Таким способом досі харчуються деякі найпростіші морські, наприклад великі - до 20 сантиметрів! - багатоядерні ксенофіофори, схожі на гігантські амеби. Вендобіонти цілком могли бути близькими до них за способом життя.
Є інші версії. У 1986 році палеонтолог Марк Мак- Менамін припустив, що вендобіонти були екологічними аналогами сучасних погонофор - глибоководних кільчастих черв'яків, позбавлених рота та кишечника. Погонофори живуть в океані на такій глибині, куди сонячне світло не проникає. Зате там зустрічаються гарячі джерела, що виділяють у воду сірководень (H2S). Тіло погонофори набите симбіотичними бактеріями, які окислюють сірководень до сірки і отриману при цьому енергію використовують для фіксації вуглекислоти, як при фотосинтезі. За рахунок цього процесу харчуються і бактерії, і черв'як, в якому вони живуть. Вендобіонтам було простіше: вони часто жили на мілководді, де сонячного світла для фотосинтезу вистачало, і цілком могли харчуватися за рахунок симбіотичних одноклітинних водоростей, що наповнювали їх тіло. Це теж цілком реально, є сучасні черв'яки та молюски, які так і роблять, - правда, для них це джерело живлення додаткове. Але чому б йому не бути основним? Світ вендобіонтів, де ніхто нікого не їв, Мак-Менамін назвав «садом Едіакари», з явною жартівливою алюзією на сад Едема (PALAIOS, 1986, 1, 2, 178-182, doi: 10.2307/3514512). Великий недолік цієї гіпотези в тому, що її поки що важко перевірити; до того ж вона свідомо не може поширюватися на всіх вендобіонтів без винятку - деякі з них жили в морі глибше рівня, куди проникає достатньо світла для фотосинтезу. ). Але, зрештою, у різних умовах вони могли харчуватися по-різному.
Парадокс у тому, що концепція «саду Едіакари» є близькою до істини при будь-якому реалістичному припущенні про спосіб харчування вендобіонтів. Тут уже й не має особливого значення, чи жили в них водорості чи ні. В едіакарском світі дійсно ніхто нікого не їв (не рахуючи одноклітинних об'єктів, але одноклітинні еукаріоти можуть і самі пожирати один одного). Принципово важливо, що в едіакарських співтовариствах до певного моменту не було не тільки хижаків (які харчувалися б іншими тваринами), а й «травоїдних» (які зшкрябали б водорості або якось активно їх виїдали). Таким чином, водоростевим матам ніхто не заважав рости.
Все змінилося, коли підвищення концентрації кисню в морській воді (судячи за геологічними даними, воно йшло поступово протягом усього едіакарію) дозволило деяким багатоклітинним істотам прискорити обмін речовин настільки, щоб почати вести по-справжньому активний образжиття. З'явилися «збирачі врожаю» - великі тварини з руховою системою та ротом, які переміщалися водоростевими матами та виїдали їх значні ділянки. Одним із таких «збирачів» була знайома нам кімберелла. За способом життя і за швидкістю пересування перші едіакарські тварини-водороїди, швидше за все, нагадували сучасних равликів; для нас це виглядає безневинно, але «з погляду» едіакарських жителів поява таких істот була справжньою катастрофою. Водорості мати негайно перестали бути суцільними; тварини не тільки зіскребали їх згори, а й об'їдали знизу, освоївши для цього проникнення в ґрунт (у зоологів прийнято називати такі дії «мінуванням»). Тут дісталося і вендобіонтам, які наприкінці едіакарію просто зникли.
З цього моменту почала працювати загальна закономірність, встановлена екологами вже давно і перевірена у різний спосіб, аж до прямих експериментів: в умовах тиску хижака різноманітність його жертв підвищується в порівнянні з спільнотою, де хижаків немає зовсім (Proceedings of the National Academy of Sciences USA) , 1973, 70, 5, 1486-1489). Якщо раніше донні спільноти насичувалися дуже нечисленними домінуючими видами водоростей, то тепер рівновага впала і почалася бурхлива еволюція. А набір екологічних ніш, доступних тваринам, теж розширювався. З'явилися активні ґрунтоєди, пристосовані до того, щоб постійно жити в норах, пропускаючи донний ґрунт крізь кишечник та витягуючи з нього поживні речовини; так досі живе багато морських черв'яків - песшкірил, наприклад. Черв'яки-ґрунтоїди вперше стали рити в морському дні не тільки горизонтальні, а й вертикальні ходи, викликаючи збагачення ґрунту киснем і тим самим додатково полегшуючи його заселення іншими тваринами. Ці події отримали назву субстратної революції (GSA Today, 2000, 10, 9, 1-7, ftp://rock.geosociety.org/pub/ GSAToday/gt0009.pdf). Таким чином, еволюціонують тварини не тільки займали готові екологічні ніші, А й активно створювали нові, перетворюючи процес на автокаталітичний (самоускоряющийся).
Деякі жителі поверхні дна стали розширювати свої екологічні ніші над бік грунту, а навпаки, у бік водної товщі. В результаті виник зоопланктон - спільнота дрібних тварин, які перебувають зваженими у воді і дрейфують разом з нею. Як правило, представники зоопланктону харчуються, фільтруючи воду і відціджуючи з неї фітопланктон, тобто одноклітинні водорості, які знаходяться в тій же водній товщі (таких на момент кембрійського вибуху було вже скільки завгодно). І справді, в ранньому кембрії в палеонтологічному літописі з'являються перші планктонні фільтратори - жаброногі рачки (Paleobiology, 1997, 23, 2, 247-262). Зяброноги, як і всі ракоподібні, - володарі членистих кінцівок, вихідно призначених для ходіння ґрунтом, тобто дном. Тому немає жодних сумнівів, що ранні етаписвоєї еволюції вони провели на дні, а до планктонного способу життя звернулися лише згодом.
Наслідки появи зоопланктону виявилися глобальними. Справа в тому, що тварини-планктери відфільтровують з води не тільки водорості, а й будь-яку завись, в якій можуть бути хоч якісь поживні речовини. В основному це розпорошені залишки мертвих організмів. Відфільтрувавши завис і всмоктавши з неї корисні молекули, планктери (насамперед цим відрізняються ракоподібні) акуратно «упаковують» у своєму кишечнику інше в щільні грудки - фекальні пелети, які швидко тонуть і вирушають на дно. Пелетне транспортування суспензії найважливіший факторзнижує каламутність води в океані. Таким чином, після появи планктонних фільтраторів вода стала прозорою, світло проникало в неї на більшу глибину, і в ній підвищилася концентрація кисню (частина його раніше витрачалася на окислення тієї ж мертвої суспензії). Перший фактор збільшив глибину зони, в якій можливий фотосинтез, другий покращив умови для придонної фауни. За всіма даними, прозорий насичений киснем фанерозойський океан різко відрізняється від каламутного докембрійського океану (Geobiology, 2009, 7, 1, 1-7). Заодно підвищилася концентрація кисню та у атмосфері. Природно, що у нових умовах різноманітність як рослин, і тварин додатково зросла. Замкнулася ще одна автокаталітична петля.
Наступ хижака
Всі тварини, про які ми досі говорили, були в найширшому значенні «травоїдними». Вони харчувалися або фотосинтезуючими організмами, або на крайній край чиїмось мертвими залишками. При цьому власна біомаса «травоїдних» була цінним (і до якогось моменту абсолютно незатребуваним) ресурсом для тварин, які харчуються іншими тваринами, тобто для хижаків. Спершу ніяких хижаків просто не існувало. Але за наявності таких атрибутів активного життя, як нервова система, мускулатура і ротовий апарат, їхня поява була лише питанням часу. Перші великі хижаки, вже безперечно спеціалізовані на харчуванні іншими багатоклітинними тваринами, з'являються приблизно 520 мільйонів років тому; це динокариди - добре плаваючі істоти, споріднені з членистоногим («Gondwana Research», 2014, 25, 896-909, doi, 10.1016/j.gr.2013.06.001). Найвідоміший представник динокарид - аномалокаріс, струнке сегментоване створення довжиною близько метра зі складними фасетковими очима і потужними членистими навколоротовими кінцівками, які явно служили для захоплення рухомого видобутку. На початку кембрія подібних хижаків немає. «Скелетна революція», безсумнівно, певною мірою стала відповіддю з їхньої поява; зміна хімічного складу морської води лише полегшила її. А поява кістяків, у свою чергу, запустила освоєння нових екологічних ніш. Стівен Стенлі абсолютно правильно писав, що для пояснення кембрійського вибуху цілком вистачає чисто біологічних причин; фактори, що діють на біосферу ззовні, могли вплинути на швидкість того чи іншого процесу, але всі головні події можна пояснити без них. Спалах різноманітності багатоклітинних тварин був природним результатом серії автокаталітичних процесів, запущених появою перших «травоїдних» (на зразок кімберели) і тих, що відбувалися на рівні спільнот, інакше кажучи, екосистем. Поза екологією пояснити кембрійський вибух справді неможливо.
З появою хижаків процес утворення нових життєвих форм став потроху гальмуватися. Репертуар екологічних ніш склався, майже всі вони вже були розподілені та зайняті. Зрозуміло, розширення угруповань тривало і далі - просто повільніше. Наприклад, тільки після закінчення кембрійського періоду з'явилися лопатоногі молюски, що зайняли досить-таки екзотичну нішу хижаків, що риють («Advances in Marine Biology», 2002, 42, 137-236). Але такого розмаху, як на рубежі едіакарію та кембрію, великомасштабна еволюція тварин більше ніколи не досягала.
З погляду подійної історії початком кембрійського вибуху вважатимуться поява перших ефективних водоростоїдів (кімберелла), а кінцем - поява перших ефективних хижаків (аномалокаріс). Кімберелла з'явилася 555 мільйонів років тому, аномалокаріс – 520 мільйонів років тому, інтервал між ними – 35 мільйонів років. Не так вже й швидко.
Виникли на кордоні кембрія майже раптово, не будучи нащадками фауни, що раніше існувала.
Однак інші дослідження, як відносяться до 1970-х років, так і пізніші, зазначали, що складні тварини, подібні до сучасних видів, виникли задовго до початку кембрія. У будь-якому випадку, численні свідчення на користь існування життя задовго до Кембрію зняли з порядку денного питання про «кембрійський вибух», як феномен раптового виникнення життя.
Питання ж про причини та механізми чергового зростання складності та різноманітності форм життя на кордоні кембрію, які не є нащадками зниклих хайнаньської та вендської біот, залишається відкритим. На сьогоднішній день проблематика «кембрійського вибуху» зосереджена на двох ключових питаннях:
- чи справді в ранньому кембрії відбулося «вибухове» зростання різноманітності та складності організмів, і
- що могло спричинити таку швидку еволюцію.
Джерела інформації
Побудова точної хронології подій на кордоні між докембрієм та раннім кембрієм становить значні труднощі. У силу цього опису послідовності та взаємозв'язку тих чи інших явищ у рамках обговорення кембрійського вибуху слід розглядати з певною обережністю.
Крім проблем з датуванням, вивчення подій на кордоні кембрію утруднено нестачею самого палеонтологічного матеріалу. На жаль, що далі від нас досліджуваний період, тим менш доступні вивчення його скам'янілості. Серед іншого причинами цього є:
Останки живих організмів
Поряд із скам'янілостями, кембрійські відкладення містять надзвичайно високу кількість родовищ, що зберегли відбитки м'яких частинтіл різних організмів. Такі відбитки дозволяють детально вивчити тварин, які не зберігаються у вигляді скам'янілостей, а також внутрішній пристрійта функціонування організмів, які зазвичай представлені лише раковинами, шипами, пазурами тощо.
У кембрії найбільш важливими родовищами є: ранньокембрійські
середньокембрійські
та верхньокембрійське родовище
- Орстен (Швеція).
Хоча всі ці родовища чудово зберегли анатомічні деталі організмів, і вони далекі від досконалості. Більшість кембрійської фауни, можливо, взагалі в них не представлена, оскільки родовища формувалися в специфічних умовах (зсувах або вулканічному попелі, що дуже швидко зберігали м'які частини тіл). На додаток до цього, відомі родовища покривають лише обмежений період кембрію і не торкаються найважливішого часу безпосередньо перед його початком. Оскільки поховання, що добре збереглися, взагалі рідкісні, а родовища скам'янілостей - вкрай рідкісні, дуже малоймовірно, що в них представлені всі типи існуючих організмів.
Скам'янілі сліди, залишені живими організмами
Скам'янілі сліди складаються, головним чином, із доріжок та норок, залишених на морському дні. Такі сліди дуже важливі, оскільки вони надають досліднику дані про організми, чиї тіла не збереглися в скам'янілості. Найчастіше вони дозволяють вивчити організми, які стосуються періоду, від якого не збереглися останки тварин, здатних залишити такі сліди. Хоча точне співвіднесення слідів з організмами, що залишили їх, як правило, неможливо, сліди можуть бути найбільш раннім доказом існування щодо складних тварин (подібних, наприклад, дощовим черв'якам).
Геохімічні спостереження
У геологічних породах, що відносяться до нижньої межі кембрію та його початку, відзначені сильні коливання ізотопного складу трьох елементів - стронцію (87 Sr/86 Sr), сірки (34 S/32 S) та вуглецю (13 C/12 C).
- Масове вимирання. Масові вимирання організмів повинні безпосередньо підвищувати частку ізотопу 12 °C у складі відкладень і таким чином зменшувати співвідношення 13 °C/12 °C.
- Вивільнення метану. У вічній мерзлоті і на континентальному шельфі молекули метану, що виробляються бактеріями, утримуються в «клітині» з молекул води, формуючи суміш, звану клатрат метану . Будучи вироблений живими організмами, цей метан збагачений ізотопом 12 C. У міру зростання температури або падіння атмосферного тиску, клатрати розпадаються. Подібний розпад вивільняє в атмосферу запасаний метан, збагачений вуглецем-12. В атмосфері метан перетворюється на діоксид вуглецю і воду, причому діоксид вуглецю взаємодіє з мінералами, формуючи вуглецевмісні породи з надлишком вуглецю-12. В результаті ізотопний склад геологічних відкладень зміщується у бік 12 °C.
Порівняльна анатомія
Кладистика - це спосіб побудови «еволюційного дерева» організмів, найчастіше - шляхом зіставлення їх анатомічного будови. З допомогою такого аналізу можна зіставляти між собою як сучасні, і викопні організми задля встановлення ходу їх еволюції. У ряді випадків можна зробити висновок, що група A повинна була з'явитися раніше груп B і C, оскільки ті мають більше подібності між собою, ніж з A. Сам по собі (без співвіднесення з даними палеонтологічних розкопок) цей метод нічого не говорить про час, коли відбувалися зміни, проте здатний відновлювати послідовності еволюційного розвитку організмів.
Молекулярна філогенетика
Палеонтологічні свідчення
У цьому розділі основні свідоцтва упорядковані за часом формування тих відкладень, у яких їх було виявлено, оскільки датування є центральним питанням щодо кембрійського вибуху. При цьому слід пам'ятати про неоднозначність хронології копалин, що належать до цього періоду.
Огляд знахідок починається з часу, що передує кембрію, і закінчується в ранньому ордовику, оскільки існує думка, що формування основних типів сучасної фауни почалося до і закінчилося після кембрія.
Дані молекулярної філогенетики (1,2 – 0,5 млрд років)
Досі продовжуються дебати щодо хронологічної інтерпретації даних молекулярної філогенетики:
У будь-якому випадку, дані молекулярної філогенетики говорять про те, що формування основних типів тварин було досить тривалим процесом, що далеко виходить за рамки 10 млн. років (близько 543-533 млн. років тому) «кембрійського вибуху».
Докембрійські сліди багатоклітинних
Існують як непрямі, і прямі палеонтологічні свідчення те, що багатоклітинні організми вперше виникли задовго на початок кембрія.
Занепад бактеріальних матів (1,25 млрд років)
Докембрійський строматоліт

Сучасні строматоліти. Західна Австралія.
Строматоліти становлять важливу частину скам'янілостей, починаючи з часу близько 3 млрд. років тому. Їх розквіт припадає на період 1,25 млрд. років тому, після чого вони стали занепадати (як у загальній чисельності, так і у різноманітності). На початок кембрію таке скорочення становило вже близько 20%.
Найбільш поширеним поясненням занепаду є припущення, що мікроорганізми, що входять до складу бактеріальних матів, стали жертвами поїдання («обгризання строматолітів») з боку інших живих організмів (що має свідчити про існування досить складних хижаків вже близько 1 млрд років тому). Це припущення підтверджується антикореляціями, що спостерігаються, між різноманітністю і чисельністю строматолітів - з одного боку і багатством морської фауни - з іншого. Так, повторний занепад строматолітів припав на пізній ордовик - відразу після чергового «спалаху» різноманітності та чисельності морської фауни. У ході ордовиксько-силурійського та пермсько-тріасового вимирань знову спостерігалося відновлення строматолітів - з наступними занепадами у міру відновлення морської фауни.
Розвиток засобів захисту у акрітархів. Раннє хижацтво (1 млрд років)
Акритархи - це скам'янілі залишки копалин невизначеної природи, як правило - оболонки цист одноклітинних та багатоклітинних водоростей. Вперше вони виявляються у відкладах, що датуються часом 2 млрд. років тому.
Близько 1 млрд років тому відбулося різке зростання у їх чисельності, різноманітності, розмірах, анатомічній складності та, особливо, у кількості та видах колючок. Чисельність архітархів різко скоротилася в ході глобального заледеніння, проте згодом відновилася з досягненням максимальної різноманітності вже в палеозої.
Їхні виключно колючі форми, що належать до періоду 1 млрд років тому, можуть свідчити про існування вже тоді хижаків, досить великих, щоб роздавлювати їх або заковтувати цілком. Інші групи невеликих організмів неопротерозою також мають ті чи інші засоби захисту від хижаків.
Сліди, залишені багатоклітинними організмами (1 млрд років)
В Індії у складі відкладень, датованих 1 млрд років тому, виявлено скам'янілості, які можуть бути слідами організмів, що рухалися через і крізь м'які породи. Знайдені сліди залишено, очевидно, безпосередньо під шаром ціанобактеріальних матів, що покривали морське дно. Дослідники зробили висновок, що сліди зобов'язані своєю появою перистальтиці тришарових багатоклітинних розміром до 5 мм - іншими словами, тваринами, чий діаметр був порівняний з діаметром земляних черв'яків, і можливо, що мали в цілому. Інші дослідники вважають, що ці та подібні до них знахідки старші 600 млн років, були залишені не живими організмами, а фізичними процесами.
Багатоклітинні ембріони з Доушаньто (632-550 млн років)
Однак виявлення в 2007 році ембріонів, оточених складно влаштованою оболонкою (у породах віком 580-550 млн років), свідчить про те, що скам'янілості в Доушаньто - не що інше, як яйця багатоклітинних безхребетних, що покояться. Більше того, стало ясно, що частина акритархів, виявлених у ранніх породах Доушаньто (632 млн років), насправді представляють оболонки таких ембріонів.
Інше викопне з Доушаньто - Vernanimalcula(Від 0,1 до 0,2 мм в діаметрі, вік близько 580 млн років) - розглядається рядом вчених як останки тришарового білатерального організму, що мав загалом, тобто тварини настільки ж складного, як дощові черв'яки або молюски. Незважаючи на сумніви в органічній природі цих копалин, оскільки всі 10 знайдених зразків Vernanimalculaмають однакові розмір та конфігурацію, малоймовірно, щоб подібна одноманітність була результатом неорганічних процесів.
У пізніших відкладах Доушаньто також виявлено різке падіння співвідношення ізотопів вуглецю 13 C/ 12 C. Хоча ця зміна є загальносвітовою, вона не збігається за часом з іншими великими подіями, такими як масові вимирання. Можливе пояснення полягає в ланцюгової реакції» взаємопов'язаної еволюції організмів та змін хімічного складу морської води. Багатоклітинні організми, активно поглинаючи вуглець з води, могли сприяти зростанню концентрації кисню, розчиненого в морській воді, у свою чергу, забезпечуючи появу нових багатоклітинних (таких як Namapoikia).
Едіакарська фауна (610-543 млн років)



Спріггіна

Багатоклітинні копалини тварини едіакарського періоду були вперше виявлені в місцезнаходженні Едіакарські пагорби в Австралії, а потім і у відкладеннях з інших регін: Чарнвудскій ліс (Англія) і півострів Авалон (Канада). Ці копалини мають вік 610-543 млн років (едіакарський період передує кембрійському). Більша їх частина мала розміри в кілька сантиметрів і була істотно більшою за своїх попередників. Багато з цих організмів не мають аналогів з жодним з видів, що жили до або після едіакарського періоду. Висловлювалися припущення, що найбільш «дивні» представники едіакарської фауни мають бути віднесені до окремого царства – «Вендобіонти» (Vendozoa). Саме до них належить чарнія - найдавніша зі знахідок едіакарського періоду (вік - 580 млн років).
Однак деякі едіакарські організми можуть виявитися попередниками пізнішої фауни:
Отвори в раковинах Cloudina. Відбір у системі «хижак-жертва»
У ряді місць до 20% скам'янілостей Cloudinaмістять отвори діаметром від 15 до 400 мкм, залишені хижаками. Деякі Cloudinaбуло пошкоджено кілька разів, що говорить про їхню здатність переживати атаки (хижаки повторно не атакують порожні раковини). Дуже схожі з Cloudinaкопалини Sinotubulites, що виявляються в тих же похованнях, взагалі не містять отворів. Така вибірковість може свідчити про існування вже в едіакарському періоді еволюційного відбору розмірних класів, а також спеціалізації жертв у відповідь на хижацтво, що розглядається як одна з причин вибуху Кембриї.
Зростання різноманітності слідів, залишених організмами (565-543 млн років)
Найбільш ранні едіакарські копалини, що належать до часу 610-600 млн. років тому, містили лише сліди, залишені цвітінням. Близько 565 млн років тому з'являються складніші сліди. Щоб їх залишити, організмам був потрібний шкіряний мішок, а їхня загальна будова - бути складнішою, ніж у кучерявих або плоских черв'яків.
Безпосередньо перед початком кембрія (близько 543 млн. років тому) виникає безліч нових слідів, включаючи вертикальні норки. Diplocraterionі Skolithos), а також сліди можливих членистоногих ( Cruzianaі Rusophycus). Вертикальні норки свідчать про те, що червоподібні тварини набули нової поведінки і, можливо, нові фізичні можливості. Сліди ж Cruzianaі Rusophycusговорять про існування у безпосередніх попередників членистоногих екзоскелетів, хоча можливо і не настільки жорсткого, як згодом.
Кембрійські копалини
Раковинна фауна (543-533 млн років)
Копалини, відомі як «дрібна раковиста фауна» (англ. дрібні скелі fossils), були виявлені в різних частинах світу і датуються кінцем венда (немакіт-далдинський ярус) та першими 10 млн років від початку кембрія (томмотський ярус). До них відноситься дуже різноманітна колекція копалин: голки, склерити (пластинки броні), трубки, археоціати (група губок або близьких до них тварин), а також невеликі раковини, що дуже нагадують брахіопод і равликоподібних молюсків, хоча і дуже маленьких (1-2 мм в довжину).
Раннекембрійські трилобіти та голкошкірі (530 млн років)
Фауна Сіріус Пассет (527 млн років)
Найбільш поширеними копалинами гренландського поховання Сіріус Пассет є членистоногі. Також є кілька організмів з твердими (мінералізованими) частинами тіла: трилобіти, хіоліти, губки, брахіоподи. Повністю відсутні голкошкірі та молюски.
Найбільш дивними організмами Сіріус Пассет були Pambdelurionі Керігмачела. Їх довге тіло, що складається з м'яких сегментів, з парою широких «плавників» на більшості сегментів і парою сегментованих придатків ззаду роблять їх схожими з аномалокаридами. При цьому зовнішні частини верхньої поверхні «плавників» мали рифлені поверхні, які можуть бути зябрами. Під кожним "плавником" є коротка безкісткова нога. Така будова дозволяє пов'язати їх із членистоногими.
Ченцзянська фауна (525-520 млн років)

Haikouichthys - реконструкція

Аномалокаріс - реконструкція

Hallucigenia - реконструкція
Ця фауна описана з кількох місць виявлення скам'янілостей у повіті Ченцзян (міський округ Юйсі, провінція Юньнань, Китай). Найбільш важливим є Maotianshan shale- поховання, в якому дуже добре представлені скам'янілості м'якотілих тварин. Ченцзянська фауна відноситься до періоду 525-520 млн років тому - середині раннього кембрію, на кілька мільйонів років пізніше Sirius Passetі щонайменше на 10 млн років раніше сланців Берджес.
У скам'янілостях виявлені частини тіла найдавніших хордових (тип, до якого належать всі хребетні):
У цих відкладеннях знайдені представники груп, близьких до членистоногих:
Ймовірно, ці організми відносяться до групи. Lobopodia, до якої із сучасних груп належать оніхофори .
Близько половини ченцзянських копалин є членистоногими, частина яких мала твердий, мінералізований екзоскелет, як і більшість пізніших морських членистоногих. Лише 3% організмів мали тверді панцирі (в основному - трилобіти). Тут також виявлені представники багатьох інших типів тварин:
- Приапуліди (риючі морські черв'яки - засадні хижаки);
- Щетинкощелепні (морські безхребетні, що входять до складу планктону);
- Гребеневики (кишковопорожнинні, зовні схожі на медуз);
- Голкошкірі (морські зірки, морські огірки тощо),
- Хіоліти (загадкові тварини, що мали маленькі конічні раковини),
Раннекембрійські ракоподібні (520 млн років)
Сланці Берджес (515 млн років)
Основна стаття: Берджес-Шейл
 Marrella
Marrella


Pikaia - реконструкція
Сланці Берджес - перший з відомих великих поховань кембрійського періоду, відкритий Уолкоттом в 1909 році. Повторний аналіз копалин, виконаний Віттінтоном та його колегами в 1970-х, ліг в основу книги Гулда « Дивовижне життя», яка відкрила кембрійський вибух для широкої публіки.
Серед копалин сланців Берджес найбільш поширені членистоногі, проте багато з них незвичайні і важко піддаються класифікації:

Opabinia - реконструкція

Wiwaxia - реконструкція
Крім того, у похованні представлені зразки екзотичних організмів:
Виникнення нових екосистем та типів після кембрію
Через велике вимирання на кордоні кембрія та ордовика типові для палеозою морські екосистеми сформувалися лише в ході подальшого відновлення морської фауни. Найбільш ранні копалини, що належать до мшанок, також вперше виявляються в ордовицькому періоді - набагато пізніше «кембрійського вибуху».
Висновки
Тривалий процес виникнення багатоклітинних
За часів Дарвіна все, що було відомо про викопні останки, дозволяло припустити, що основні типи багатоклітинних виникли і сформувалися протягом декількох мільйонів років - від раннього до середнього кембрію. Аж до 1980-х років ці уявлення ще залишалися чинними.
Проте останні знахідки свідчать, що щонайменше деякі тришарові билатеральные організми існували на початок кембрія: Kimberellaможна розглядати як ранніх молюсків, а подряпини на скельних породах біля цих копалин дозволяє припустити молюскоподібний метод харчування (555 млн. років тому). Якщо ж припустити, що Vernanimalculaмала тришаровий білатеральний цілому, це відсуває виникнення складних тварин ще на 25-50 млн років тому. Виявлення отворів в оболонці Cloudinaтакож дозволяє припустити наявність розвинених хижаків наприкінці едіакарського періоду. Крім того, деякі сліди у копалин, що належать до середини едіакарського періоду (близько 565 млн років тому), могли бути залишені тваринами, складнішими, ніж плоскі червиі мають шкіряний мішок.
Задовго до цього тривалий захід строматолітів (почався близько 1,25 млрд років тому), говорить про раннє виникнення тварин, досить складних для їх «обгризання». Зростання ж достатку та різноманітності шипів у акритархів у цей час призводить до висновку, що вже тоді існували хижаки, досить великі для того, щоб такий захист був необхідний. На іншому кінці тимчасової шкали, що належить до кембрійського вибуху, слід зазначити відсутність низки основних типів нинішньої фауни до кінця кембрія, а типових палеозойських екосистем - до ордовика .
Таким чином, на сьогодні спростовано точку зору, згідно з якою тварини «сучасного» рівня складності (порівняного з безхребетними, що нині живуть) виникли протягом усього декількох мільйонів років раннього кембрію. Тим не менш, переважна частина сучасних типів вперше з'явилася в кембрії (за винятком молюсків, голкошкірих та членистоногих, можливо, що виникли в едіакарський період). Крім того, вибухове зростання таксономічної різноманітності також спостерігалося на початку кембрію.
«Вибух» таксономічного розмаїття в ранньому кембрії
«Таксономічне розмаїття» означає кількість організмів, які значно різняться за своєю будовою. У той же час «морфологічна різноманітність» означає загальну кількість видів і нічого не говорить про кількість базових «дизайнів» (можливо безліч варіацій невеликої кількості основних типів анатомічної будови). Немає сумнівів, що саме таксономічна різноманітність різко зросла в ранньому кембрії і зберігалася на цьому рівні протягом усього періоду - ми можемо виявити сучасних тварин (таких як ракоподібні, голкошкірі, і риби) майже в один і той же час, а найчастіше - і у загальних похованнях з організмами типу аномалокарису та Halkieria, Які вважаються «дядями» або «пра-дядями» сучасних видів.
При ближчому вивченні виявляється інший сюрприз - деякі сучасно виглядають тварини, наприклад ранньокембрійські ракоподібні, трилобіти і голкошкірі, знаходяться в більш ранніх відкладеннях, ніж деякі «дядьки» або «пра-дядьки» груп, що нині живуть, не залишили прямих нащадків. Це може бути результатом розривів та варіацій формування викопних відкладень, або означати, що предки сучасних організмівеволюціонували в різні часи і, можливо, з різною швидкістю.
Можливі причини вибуху
Незважаючи на те, що досить складні тришарові тварини існували до (а можливо, задовго до) кембрію, еволюційний розвиток в ранньому кембрії видається виключно швидким. Було багато спроб пояснити причини такого «вибухового» розвитку.
Зміни довкілля
Зростання концентрації кисню
Найраніша атмосфера Землі взагалі містила вільного кисню. Той кисень, яким дихають сучасні тварини - як у повітрі, і розчинений у воді - є продуктом мільярдів років фотосинтезу , переважно - мікроорганізмів (таких як ціанобактерії). Приблизно 2,5 мільярда років тому концентрація кисню в атмосфері різко зросла. До цього часу весь кисень, що виробляється мікроорганізмами, повністю витрачався на окислення елементів з високою спорідненістю до кисню, таких як залізо. Поки не відбулося їхнє повне зв'язування на суші та у верхніх шарах океану, в атмосфері існували лише локальні «кисневі оази».
Нестача кисню могла тривалий час перешкоджати розвитку великих складних організмів. Проблема полягає в тому, що кількість кисню, яке тварина може абсорбувати з довкілля, обмежена площею поверхні (легких і зябер у найбільш складних тварин; шкіри - у більш простих). Кількість кисню, необхідне життєдіяльності, визначається масою і обсягом організму, які зі збільшенням розмірів зростають швидше, ніж площа. Зростання концентрації кисню в повітрі і у воді могло послаблювати або зовсім усувати це обмеження.
Слід зазначити, що достатньо кисню існування великих вендобионтов було вже у эдиакарский період. Однак подальше зростання концентрації кисню (між едіакарським та кембрійським періодами) могло надати організмам додаткову енергію для виробництва речовин (таких як колаген), необхідних для розвитку принципово більш складних структур тіла, у тому числі - використовуваних для хижацтва та захисту від нього.
Земля-сніжок (Snowball Earth)
Існують численні докази того, що в пізньому неопротерозої (включаючи ранній едіакарскій період) Земля піддавалася глобальному заледеніння, в ході якого більшість її була покрита льодом, а температура поверхні була близька до точки замерзання навіть на екваторі. Деякі дослідники вказують, що ця обставина може бути тісно пов'язана з кембрійським вибухом, оскільки ранні з відомих копалин відносяться до періоду незабаром після кінця останнього повного заледеніння.
Однак досить важко вказати причинно-наслідковий зв'язок таких катастроф із подальшим зростанням розмірів та складності організмів. Можливо, низькі температуризбільшували концентрацію кисню в океані - його розчинність у морській воді зростає майже вдвічі при падінні температури з 30 до 0 °C.
Флуктуація ізотопного складу вуглецю
У відкладеннях на межі едіакарського та кембрійського періодів спостерігається дуже різке зниження, а слідом за ним – надзвичайно сильні коливання співвідношення ізотопів вуглецю 13 C/12 C протягом усього раннього кембрію.
Багато вчених припускали, що вихідне падіння пов'язане з масовим вимиранням безпосередньо перед початком кембрію. . Можна також припустити, що вимирання саме стало наслідком розпаду клатратів метану, що передував. Широко відомо, що емісія метану та подальше насичення атмосфери діоксидом вуглецю викликає глобальний парниковий ефект, що супроводжується різними екологічними катастрофами. Подібна картина (різке падіння співвідношення 13 C/12 C з наступними коливаннями) спостерігалася у тріасі, коли життя відновлювалося після масового Пермського вимирання.
Однак досить важко пояснити, як масове вимирання могло викликати різке зростання таксономічного та морфологічного розмаїття. Хоча масові вимирання, такі як пермське і крейда-палеогенове, призводили до подальшого зростання чисельності. окремих видіввід несуттєвої до «домінуючої», проте в обох випадках екологічні ніші заміщалися хоча й іншими, але складними організмами. При цьому стрибкоподібного зростання таксономічного чи морфологічного розмаїття у новій екосистемі не спостерігалося.
Ряд дослідників припускав, що кожне короткочасне зниження частки 13 C/12 C в ранньому кембрії представляє вивільнення метану, яке завдяки викликаному ним невеликому парниковому ефекту та підвищенню температури, призводило до зростання морфологічного розмаїття. Але ця гіпотеза не пояснює різкого збільшення таксономічного розмаїття на початку кембрія.
Пояснення з урахуванням розвитку організмів
В основі низки теорій лежить та ідея, що щодо малі зміни у способі, яким тварини розвиваються з ембріона у дорослий організм, можуть призвести до різких змін форми тіла.
Виникнення системи білатерального розвитку
Hox-гени у різних груп тварин настільки схожі, що, наприклад, можна трансплантувати людський ген «формування очей» в ембріон дрозофіли, що призведе до формування ока - але це буде око дрозофіли завдяки активації відповідних «робочих» генів. Звідси видно, що наявність подібного набору Hox-генів зовсім не означає анатомічної подібності організмів (одні і ті ж Hox-гени можуть керувати формуванням різних структур, як очі людини і комахи). Тому виникнення подібної системи могло спричинити різке зростання різноманітності – як морфологічної, так і таксономічної.
Оскільки одні й самі Hox-гени управляють диференціацією всіх відомих билатеральных організмів, еволюційні лінії останніх мали розійтися доти, як вони почали утворюватися якісь спеціалізовані органи. Таким чином, «останній загальний предок» всіх білатеральних організмів повинен був бути невеликим, анатомічно простим і, найімовірніше, схильним повному розкладаннюбез збереження в скам'янілості. Ця обставина робить його виявлення вкрай малоймовірним. Однак цілий ряд вендобіонтів (наприклад, кімберелла, сприггіна або Arkarua), можливо, мав білатеральну будову тіла (на думку ряду вчених, це не так - симетрія вендобіонтів не білатеральна, а ковзна, що принципово відрізняє їх від більшості інших організмів). Таким чином, подібна системарозвитку могла виникнути щонайменше за кілька десятків мільйонів років до кембрійського вибуху. І тут для його пояснення необхідні якісь додаткові причини.
Розвиток статевого розмноження
Організми, які використовують статеве розмноження, змінюються дуже слабко. У більшості організмів, що мають статеве розмноження, нащадок отримує приблизно по 50% своїх генів від кожного з батьків. Це означає, що навіть невелике зростання складності геному здатне породити безліч варіацій будови та форми тіла. Більшість біологічної складності, ймовірно, виникає завдяки дії щодо простих правил на великій кількості клітин, що функціонують як клітинні автомати (прикладом такого ефекту служить конвіївська гра «Життя», де складні форми та складна поведінка демонструються клітинами, що діють за виключно простими правилами). Можлива поява статевого розмноження або його суттєвий розвиток у період кембрійського вибуху для дуже примітивних і схожих істот може означати те, що існувала можливість їхнього міжвидового та далекого схрещування. Це різко збільшувало мінливість. Лише з розвитком геному з'являються по-справжньому ізольовані види, що не схрещуються з іншими. Приклад сучасних істот такого роду - корали.
Колія розвитку
Деякі вчені припускають, що в міру ускладнення організмів на еволюційні зміни загальної будовитіла накладаються вторинні зміни у бік кращої спеціалізації його частин, що склалися. Це знижує можливість проходження природного відбору новими класами організмів - через конкуренцію з «удосконаленими» предками. Через війну, зі складання загального (на рівні таксономічного класу) будівлі, формується «колію розвитку», а просторова структура тіла «заморожується». Відповідно формування нових класів відбувається «легше» на ранніх стадіях еволюції основних скарбів, а їх подальша еволюція йде на нижчих таксономічних рівнях. Згодом автор цієї ідеї вказував, що таке заморожування не є основним поясненням кембрійського вибуху.
Скам'янілості, які б підтвердити цю ідею, неоднозначні. Відзначено, що варіації організмів одного класу найчастіше найбільші на перших стадіях розвитку скарби. Наприклад, деякі кембрійські трилобіти сильно варіювали за кількістю грудних сегментів, причому згодом подібна різноманітність суттєво знизилася. Однак виявлено, що зразки силурійських трилобітів мають таку ж високу варіативність будівлі, що і ранньокембрійські. Дослідники припустили, що загальне зниження різноманітності пов'язане з екологічними чи функціональними обмеженнями. Наприклад, очікується меншої варіативності числа сегментів після того, як у трилобітів (що нагадували сучасну мокрицю) сформувалася опукла будова тіла, що є ефективним способом його захисту.
Екологічні пояснення
Такі пояснення зосереджені взаємодії між різними видами організмів. Деякі з подібних гіпотез мають справу із змінами харчових кіл; інші розглядають гонку озброєнь між хижаками та жертвами, що могла викликати еволюцію жорстких частин тіла у ранньому кембрії; Ще кілька гіпотез зосереджено більш загальних механізмах коеволюції (найвідоміший пізніший приклад коеволюції квіткових рослин із комахами-запилювачами).
«Гонка озброєнь» між хижаками та жертвами
Хижацтво за визначенням передбачає загибель жертви, внаслідок чого воно стає сильним чинником та прискорювачем природного відбору. Тиск на жертви у напрямі кращої адаптації має бути сильнішим, ніж на хижаків – оскільки, на відміну від жертви, вони мають шанс зробити нову спробу (ця асиметрія відома як принцип «життя проти обіду» – хижак ризикує втратою лише обіду, в той час як жертва ризикує життям).
Однак є свідчення (наприклад - скам'янілості колючих акрітархів, а також отвори, виконані в оболонці клаудінід) того, що хижацтво було присутнє задовго до початку кембрію. Тому малоймовірно, що воно саме по собі стало причиною кембрійського вибуху, хоча й мало сильний вплив на анатомічні форми організмів, що виникли при цьому.
Поява фітофагів
Стенлі (1973) припустив, що поява 700 млн. років тому найпростіших (одноклітинних еукаріотів), що «обголодують» мікробні мати, вкрай розширило харчові ланцюги і мало призвести до зростання різноманітності організмів. Однак сьогодні відомо, що «обгладування» виникло понад 1 млрд років тому, а згасання строматолітів почалося близько 1,25 млрд років тому - задовго до «вибуху».
Зростання розмірів та різноманітності планктону
Геохімічні спостереження чітко свідчать, що Загальна масапланктону стала порівнянною з нинішньою вже в ранньому протерозої. Проте до кембрія планктон не вносив суттєвого внеску у харчування глибоководних організмів, оскільки їхні тіла були надто малі для швидкого занурення на морське дно. Мікроскопічний планктон поїдався іншим планктоном або руйнувався хімічними процесами у верхніх шарах моря задовго до проникнення в глибоководні шари, де міг би стати їжею для нектону та бентосу (плаваючі організми та мешканці морського дна відповідно).
У складі ж ранніх кембрійських копалин виявили мезозоопланктон (планктон середніх розмірів, помітний неозброєним оком), який міг відфільтровувати мікроскопічний планктон (головним чином, фітопланктон - планктонну «рослинність»). Новий мезозоопланктон міг служити джерелом останків, а також виділяти екскременти у формі капсул, досить великих для швидкого занурення - вони могли бути їжею для нектону та бентосу, викликаючи зростання їх розмірів та різноманітності. Якщо ж частинки органіки досягали морського дна, в результаті подальшого поховання вони мали підвищувати концентрацію кисню у воді при одночасному зниженні концентрації вільного вуглецю. Іншими словами, поява мезозоопланктону збагатило глибокі ділянки океану як їжею, так і киснем, і, тим самим, уможливило появу та еволюцію більших і різноманітних мешканців морських глибин.
Нарешті, виникнення серед мезозоопланктону фітофагів могло сформувати додаткову екологічну нішу для великих мезозоопланктонних хижаків, чиї тіла, поринаючи в море, вели до подальшого його збагачення їжею та киснем. Можливо, першими хижаками серед мезозоопланктону були личинки донних тварин, чия подальша еволюція стала результатом загального зростання хижацтва у морях едіакарського періоду.
Безліч порожніх ніш
Джеймс Валентайн (James W. Valentine) у кількох роботах зробив такі припущення: різкі зміни у будові тіла є «скрутними»; зміни мають набагато більше шансів на існування, якщо вони зустрічають слабку (або зовсім не зустрічають) конкуренцію за екологічну нішу, на яку вони націлені. Останнє необхідно, щоб новий типорганізмів мав достатньо часу для адаптації до своєї нової ролі.
Ця обставина повинна призводити до того, що реалізація основних еволюційних змін набагато вірогідніша на початкових стадіях формування екосистеми - через те, що подальша диверсифікація заповнює майже всі екологічні ніші. Надалі, незважаючи на те, що нові типи організмів продовжують виникати, нестача порожніх ніш перешкоджає їх поширенню в екосистемі.
Модель Валентайна добре пояснює факт унікальності кембрійського вибуху – чому він трапився лише один раз і чому його тривалість була обмежена.
Примітки
- Darwin, C. On Origin of Species by Natural Selection. - London, United Kingdom: Murray, 1859. - P. 315-316.
- Walcott, C. D. Cambrian Geology and Paleontology // Smithsonian Miscellaneous Collections. – 1914. – Т. 57. – P. 14.
- Whittington, H.B.; Geological Survey of Canada. Origins and early evolution of predation // The Burgess Shale. - Yale University Press, 1985.
- Gould, SJ. Wonderful Life: The Burgess Shale and the Nature of History. - Нью-Йорк: W.W. Norton & Company, 1989. – ISBN 0-393-02705-8
- McNamara, KJ. Dating the Origin of Animals // Science. – 1996-12-20. – Т. 274. – № 5295. – С. 1993–1997.
- Awramik, S. M. Precambrian columnar stromatolite diversity: Reflection of metazoan appearance // Science. – 1971-11-19. – Т. 174. – № 4011. – С. 825–827.
- Fedonkin, M. A.; Waggoner, B. Late Precambrian fossil Kimberella є mollusc-like bilaterian organism // Nature. – 1997. – Т. 388. – С. 868–871.
- Єськов, До.Чернетки Господа Бога // Знання сила. - 2001. - № 6.
- Jago, J. B., Haines, P. W.Останній radiometric dating деяких Cambrian років в південному Australia: relevance to Cambrian time scale // Revista Española de Paleontología. – 1998. – С. 115–22.
- Єськов К. Ю.Дивовижна палеонтологія: Історія Землі та життя на ній. – М.: Видавництво НЦ ЕНАС, 2007. – 540 с. – (Про що замовчали підручники). - ISBN 978-5-93196-711-0
- Gehling, J.; Jensen, S.; Droser M.; Myrow P,.; Narbonne, G. Burrowing below the basal Cambrian GSSP, Fortune Head, Newfoundland // Geological Magazine. – 2001-03. – Т. 138. – № 2. – С. 213–218.
- Benton, MJ; Wills, M.A.; Hitchin, R. Quality of the fossil record through time // Nature. – 2000. – Т. 403. – № 6769. – С. 534–537.
- Butterfield, N. J. Exceptional Fossil Preservation and Cambrian Explosion // . – 2003. – Т. 43. – № 1. – С. 166–177.
- Cowen, R. History of Life. - Blackwell Science.
- Morris, S. C. The Burgess Shale (Middle Cambrian) Fauna // Annual Review of Ecology and Systematics. – 1979. – Т. 10. – № 1. – С. 327–349.
- Yochelson, E. L Discovery, Collection, та Зображення з середини Cambrian Burgess Shale Biota by Charles Doolittle Walcott // Процеси з American Philosophical Society. – 1996. – Т. 140. – № 4. – С. 469–545.
- Butterfield, NJ. Ecology and evolution of Cambrian plankton // Columbia University Press Ecology of Cambrian Radiation. - New York, 2001. - С. 200-216.
- Signor, P. W. Sampling bias, gradual extinction patterns and catastrophes in the fossil record // Geological Society of America Geological implications of impacts of asteroids and comets on the earth. – 1982. – С. 291-296.
- Seilacher, A. How valid is Cruziana Stratigraphy? // International Journal of Earth Sciences. – 1994. – Т. 83. – № 4. – С. 752–758.
- Magaritz, M.; Holser, W.T.; Kirschvink, JL. Carbon-isotope events across the Precambrian/Cambrian boundary on the Siberian Platform // Nature. – 1986. – Т. 320. – № 6059. – С. 258–259.
- Kaufman, AJ, Knoll AH.Неопротерозійні variations в C-isotopic composition of seawater: stratigraphic and biogeochemical implications // Elsevier Precambrian Research. – 1995. – Т. 73. – С. 27-49.
- Derry, L.A.; Brasier, M.D.; Corfield, R.M.; Rozanov, A.Yu.; Журавлев, А. Ю Sr and C isotopes в Lower Cambrian carbonates від Siberian craton: Paleoenvironmental record під час 'Cambrian explosion' // Elsevier Earth and Planetary Science Letters. – 1994. – Т. 128. – С. 671-681.
- Rothman, D.H.; Hayes, J.M.; Summons, R. E. Dynamics of the Neoproterozoic carbon cycle // The National Academy of SciencesПроцедури Національної академії наук США. – 2003-07-08. – Т. 100. – № 14. – С. 8124–8129.
- Ripperdan, R. L. Global Variations in Carbon Isotope Composition Latest Neoproterozoic and Earliest Cambrian // Annual Review of Earth and Planetary Sciences. – 1994-05. – Т. 22. – С. 385-417.
- Marshall, CR. Explaining the Cambrian “Explosion” of Animals // Annual Review of Earth Planetary Sciences. – 2006. – Т. 34. – С. 355-384.
- Hug, L.A.; Roger, AJ. The Impact of Fossils and Taxon Sampling on Ancient Molecular Dating Analyses // . – 2007. – Т. 24(8). – С. 1889-1897.
- Wray, G.A.; Levinton, J.S.; Shapiro, L. H.(((Назва))) // Science. – 1996-10. – Т. 274. – № 5287. – С. 568–573.
- Ayala, FJ; Ржецький, А.; Ayala, FJ.(((Назва))) // . – 1998-01. – Т. 95. – № 2. – С. 606-611.
- Aris-Brosou, S.; Yang, Z. Bayesian Models of Episodic Evolution Support a Late Precambrian Explosive Diversification of the Metazoa // Molecular Biology and Evolution. – 2003-08. – Т. 20. – № 12. – С. 1947-1954.
- Peterson, KJ; Lyons, J.B.; Nowak, K.S.; Takacs, C.M.; Wargo, MJ; McPeek, MJ.Використання metazoan divergence times with molecular clock // Відповідь Національної академії наук (США). – 2004-04. – Т. 101. – № 17. – С. 6536-6541.
- Blair, J.E.; Hedges, S. B. Molecular Clocks Do Not Support the Cambrian Explosion // Molecular Biology and Evolution. – 2004-11. – Т. 22. – № 3. – С. 387-390.
- Bengtson, S. Origins and early evolution of predation // The fossil record of predation. Paleontological Society Papers 8 / Kowalewski, M., і Kelley, P.H.. - New York: The Paleontological Society, 2002. - С. 289-317.
- Sheehan, PM; Harris, M. T. Microbialite resurgence after the Late Ordovician extinction // Nature. – 2004. – Т. 430. – С. 75–78.
- Stanley Predation defeats competition on the seafloor // Paleobiology. – 2008. – Т. 34. – С. 1.
- Seilacher, A.; Bose, P.K.; Pflüger, F. Animals Більше Than 1 Billion Years Ago: Trace Fossil Evidence from India // Science. – 1998. – Т. 282. – № 5386. – С. 80–83.
- Jensen, S. The Proterozoic and Earliest Cambrian Trace Fossil Record; Patterns, Problems and Perspectives // Integrative and Comparative Biology. – 2003. – Т. 43. – № 1. – С. 219–228.
- Наймарк, ОленаТаємниця ембріонів Доушаньтуо розкрита. Науково-популярний проект «Елементи» (12 квітня 2007 року). Архівовано з першоджерела 24 серпня 2011 року. Перевірено 21 травня 2008 року.
Статистичний аналіз відвідуваності нашого сайту переконливо свідчить про те, що найбільшим успіхомкористуються науково-популярні статті, причому подібні матеріали проглядаються стабільно протягом тривалого терміну. Ось я і вирішив знову попрацювати в цьому руслі, висвітлюючи тему, яка хвилює мене давно.
Питання про те як міг виглядати світ на початку свого шляху, хвилювало людські уми з незапам'ятних часів. Довгий час загадки минулих епох залишалися таємницями за сімома печатками, і за відсутністю наукового знання людство творило міфи, нагадуючи оточеного казковими фантазіями дитини в колисці. З розвитком наукового світогляду загадки нікуди не зникли, вони лише набули конкретної форми. Однією із таких загадок є кембрійський вибух.
Відомо, що вік геологічних порід можна визначити стратиграфічно: молодші верстви, як правило, лежать вище за давні. За скам'янілими залишками вимерлих організмів, що насичують породи, можна відтворити картину поступової еволюціїжиття на землі. Однак поступовість ця раптово обривається на глибинах, що відповідають верствам кембрійського періоду «541,0 ± 1,0 млн років тому – 485,4 ± 1,9 млн років тому). Тут виявляються майже всі сучасні типи багатоклітинних тварин, а далі – ніби порожнеча, таємничий докембрій, наче відбувся акт створення… Цей феномен отримав у науці назву кембрійського вибуху.
 Кембрій
Кембрій Втім, насправді відбитки найдавніших тварин вченими виявлялися, проте тривалий час їх відносили до кембрійських верств і ототожнювали з групами таксономічними організмів, що збереглися до наших днів. Зіткнулися з подібними артефактами і німецькі геологи в Намібії Південної Африки в 1908 р., і Р. Спріг в Едіакарі Південної Австралії, на початку 30-х років. XX ст., що випустив 1947 р. роботу «Раннекембрійські медузи хребта Фліндерс Південної Австралії». Уникнути спокуси пояснити невідоме давно звичним – для цього досліднику потрібна певна мужність. Лише 1952 р. академік АН СРСР Б.С. Соколов встановив існування венда – періоду, що передував кембрію. «…вперше зайняла своє справжнє геохронологічне становище і так звана едіакарська фауна безскелетних Metazoa, що спочатку вважалася кембрійською… Вендський період (венд) названий на ім'я найдавнішого слов'янського племені вендів (або венедів), що жили на південь від Балтійського моря.
 Борис Сергійович Соколов
Борис Сергійович Соколов У зв'язку з цим не можу втриматися, не розповісти про курйозний випадок, коли у віці 5 років виграв у суперечці кандидата геологічних наук з цього питання. Зібралися в нас в черговий раз вдома гості, з якоїсь родинної урочистості. Завершувалась зовсім інша епоха в житті країни, і здається тепер сном благополучний радянський Баку зі столами, повними фруктами та осетровою ікрою, з регулярними візитами один до одного дружної юрби родичів. З двоюрідним братом під керівництвом старшої двоюрідної сестри ми того вечора грали в космонавтів, подорожуючи з зав'язаними очима по підвіконнях і шафах, представляючи їх іншими планетами. І була в мене в дитинстві книжка з палеонтології «Живе минуле Землі», з геохронологічною таблицею, до якої був включений вендський період… Дядько, тоді – кандидат геологічних наук, запитав мене: який період – найперший, і я відповів що венд. А дядько відповідає: ні, кембрій. Я з ним не погодився. Через деякий час, порившись у якійсь літературі, дядько визнав, що з'явилися нові наукові дані, викликавши чималі веселощі моїх батьків.

Але повернемося до Кембрійського вибуху. З т.зр. стратиграфічній відповісти про причини вибухового збільшення кількості знахідок, що спостерігається, з переходом від докембрійських товщ до кембрійських – не так вже й складно. У кембрії з'являються скелетні організми: раковини, панцирі, шипи – все це чудово зберігається у скам'янілому вигляді. Докембрійські форми – м'якотілі та безскелетні, потрібен особливий збіг обставин, щоб їхні відбитки залишили пам'ять у літописі нашої планети, і саме тому тривалий час про них було невідомо. Тому тепер історія органічного світу Землі поділяється на 2 великі еони: фанерозою – ера явного життя і криптозою – ера прихованого життя. Але що стало справжньою причиною т.зв. «скелетної революції?»
На цей рахунок є дуже різноманітні версії, які завжди можуть ознайомитися з ними в мережі Інтернет. Довгий час у вітчизняній літературі домінувала гіпотеза про те, що збільшення за геологічними причинами концентрації в морській воді іонів Са2+, необхідних для побудови захисних систем, Таких як раковини, панцирі, і т.д. - Причина. Однак ще в далекому вже тепер 2007 р. мені спала на думку у формі інтуїтивних візуальних образів інша версія, що представляється більш простою і логічною (дотримуючись принципу бритви Оккама про те, що найкоротший шлях до істини – прямою). Згодом стало ясно, що багато вчених, які займаються проблемою, пішли тим самим шляхом, і нині ця гіпотеза згадується навіть у вітчизняних шкільних підручниках біології.
«…вже вивчено тисячі екземплярів представників вендської біоти, і на жодному з них не виявлено пошкоджень або слідів укусів. Це означає, що у вендській екосистемі практично були відсутні біотурбатори, макроскопічні трупоїди та хижаки, що подрібнювали їжу. Відмерла органіка піддавалася лише мікробного розкладання». До речі, 2007 р. навіть ці факти мені не знали.
Отже, відносини хижак-жертва серед багатоклітинних з'являються лише в кембрії (хоча є дані, і це закономірно, що вони зароджувалися в едіакарі (альтернативна назва вендського періоду). «Зі збільшенням кількості кисню (в результаті мільйонів років фотосинтетичної активності водоростей – прим. авт) .), одні тварини почали споживати інших, провокуючи кембрійський вибух через ескалацію «перегони озброєнь» хижаків і жертв.» Тут мається на увазі що виникнення кисневого дихання уможливило збільшення швидкості обміну речовин і енергії, і тут моя наукова інтуїція намацує деякий причин. провальник: виникнення антагоністичних екологічних відносин хижак-жертва визначається прогресивним, з точки зору ефективності використання ресурсів, ароморфозом (ароморфоз – прогресивний еволюційний стрибок) – виникненням кисневого дихання, якщо згадати таке поняття В.І.Вернадського як тиск життя безперервним прагненням розумно тиснути біомасу в процесі розмноження з одного боку і обмежується наявними ресурсами з іншого, то логічно припустити, що відносини хижак-жертва виникли як наслідок описаних В.І. Вернадських принципів. Коротше кажучи, одного разу їх стало надто багато, а їхня зброя і те, чим вони захищалися одна від одної, добре перетворюється на камінь.
Існує гіпотеза, згідно з якою вендські тварини могли харчуватися за рахунок мікроскопічних водоростей-симбіонтів, що живуть у їх тілі, які в свою чергу споживали сонячну енергію в процесі фотосинтезу, адже багато представників едіакарської біоти, схоже, були мешканцями заливаються сонячними променями мілководій. Як додатковий такий тип харчування зустрічається і у сучасних тварин. У вендський період цей спосіб харчування міг бути основним. Світ вендобіонтів, де ніхто нікого не їв, Мак-Менамін назвав "садом Едіакари", з явною алюзією на садок Едема. Подібна алюзія виникала і в мене.
Яким був світ венда? Доба була на 3 години коротшою, а рік налічував 420 днів. Інші материки, інші океани… Докембрійський світ яскраво описують Я.Є. Малаховська та А.Ю. Іванцов ... «Дно великих мілководних морів і низовини суші покривали килими бактеріальних матів, де-не-де колихали ліси стрічковоподібних водоростей». «Це було велике зледеніння: за оцінками вчених, тоді в морі льоди запливали навіть у тропічну зону, а суша була вкрита ледь не повністю. Після закінчення льодовикової епохи мікроорганізмів і водоростей, що поверталися на мілководдя, вже включали багатоклітинних тварин. Серед цих м'якотілих істот зустрічалися гіганти, що досягали в довжину півтора метра, і зовсім малюки, не більше 2-3 мм. Одні плавали чи ширяли в товщі води, інші жили на дні: прикріплювалися до нього, вільно лежали чи повзали» . «Незважаючи на сторічну історію вивчення вендських відбитків, до останнього часу не було задовільної відповіді на питання, ким були вендські організми – рослинами, тваринами, грибами чи належали іншому царству, яке не дожило до наших днів. Більшість дослідників вважає їх багатоклітинними тваринами, можливо, лише через зовнішню схожість відбитків з деякими безхребетними».

Чарнії (Сharnia masoni)вели прикріплений спосіб життя.


Трибрахідіум (Tribrachidium heraldicum)нерухомо сидів на дні, тільки трохи ворушилися вії на зовнішній поверхні. Вії вловлювали з води дрібні органічні частинки і переганяли їх від периферії в центр до рота, можливо, навіть до трьох рот.


Дікінсонії (Dickinsonia lissa, Dickinsonia cf. tenuis, Dickinsonia ladoa)повзали. Найбільша, 1,5 м довжиною, походить з Едіакар.


Кімберелла (Kimberella quadrata)чимось нагадувала молюсків.

Єргія (Yorgia waggoneri), черевна сторона тіла якої була вкрита чимось схожим на миготливий епітелій, опустившись на дно, виїдала під собою ділянку субстрату (війки епітелію захоплювали та переміщали до рота органічні частки). Потім вона перепливала на інше місце… Після того, як ділянка морського дна, на якій паслися тварини, була засипана осадом, їх сліди змогли зберегтися завдяки щільній органічній плівці, на якій вони були «вигравіювані».


Вентогірус (Ventogyrus chistyakovi)ймовірно плавав.

Перелічувати всіх у науково-популярній статті немає сенсу…
Будучи підлітком у пострадянському Баку, я бачив найкрасивіший сон у своєму житті. Мої п'яти пестили води нічного доісторичного моря. На дні серед скель - пісок і флюоресцирующие бузковим, зеленим, бірюзовим губки і віддалено нагадують коралові поліпи кущики ... Вантаж сотень мільйонів років жорстокої еволюції і людської історії, що послідувала за нею - все далеко попереду і нагорі ... Там же, на березі вендського моря легкість та безтурботність.

Потім приплив кембрійський трилобіт. Я торкнувся його панцира.

Наставав кембрій… Я прокинувся.

Література:
- Малаховська Я. Є., Іванцов А. Ю. Барвистий ілюстрований атлас найдавніших м'якотілих тварин вендського періоду // Архангельськ, вид-во ПІН РАН: 2003. 48 с.
- Соколов Б. З. Нариси становлення венда // М.: КМК Лтд., 1997. 157 з.
- Mario Aguilera. Dawn of Carnivores Explains Animal Boom in Distant Past // UC San Diego News Center, July 30, 2013.
- Ястребов С. А. Кембрійський вибух // Хімія та Життя, 2016 №10.
- Mark A. S. McMenamin. The Garden of Ediacara // PALAIOS, Vol. 1, No. 2 (Apr., 1986), pp. 178-182
У форматі відео:
Про кембрій:
Усі випуски «Еволюції»:
Дарвін стверджував, що розвиток будь-якого виду зі свого предка є тривалим та поступовим процесом зміни, що проходить через незліченну кількість проміжних форм.
Він усвідомлював, що якщо його теорія вірна, то мали існувати тисячі цих проміжних форм. Більше того, він усвідомлював, що від існування цих форм якраз і залежала міцність його теорії.
Так, Дарвін писав, що «між усіма видами, що живуть і вимерли, мало бути немислима кількість проміжних і перехідних зв'язків. Але, без сумніву, якщо ця теорія вірна, такі існували нашій Землі».
Однак чому ж тоді, запитував він, висловлюючи свої власні сумніви, «ми не знаходимо їх без рахунка у відкладах земної кори?». Він болісно усвідомлював нестачу подібних копалин у геологічних пластах, але обманював себе та своїх читачів: «Відповідь переважно у цьому, що дані менш повні, як прийнято думати».
Тим не менш, цей факт не давав йому спокою, і він навіть присвятив йому цілий розділ у своїй книзі, розмірковуючи в ній на тему «неповноти геологічних даних».
Незважаючи на його вагому аргументацію, він явно, як і раніше, відчував деяку незручність з приводу цієї ситуації, якщо вважав за необхідне висловити в пресі свою впевненість у тому, що в «майбутні віки… будуть виявлені численні копалини зв'язку».
Перебуваючи у захваті від теорії та у впевненості, що, охопивши більше геологічних пластів, що містять залишки копалин, вони успішно усунуть цю «неповноту», геологи та палеонтологи (вчені, які вивчають копалини) доклали титанічних зусиль, щоб заповнити прогалини у викопних даних.
Як не дивно, якщо врахувати ті величезні ресурси, які задіялися для вирішення завдання протягом багатьох років, але ці зусилля не дали результату. Професор Гулд розповів, що «надзвичайна рідкість перехідних форм у викопній історії продовжує оберігатися як професійний секрет палеонтології».
У 1978 році колега Гулда, професор Найлс Елдрідж, зізнався в інтерв'ю, що «нікому не вдавалося знайти якихось „проміжних“ істот: серед викопних свідчень не виявляється жодних „відсутніх зв'язків“, і багато вчених тепер все більше схиляються до переконання, що ці перехідні форми будь-коли існували».
Професор Стівен Стенлі пише: «Насправді в копалині історії немає жодного переконливо підтвердженого випадку переходу одного виду в інший. Крім того, види існували разюче тривалі періоди часу». Нікому, наприклад, не вдавалося знайти копалини жирафа з шиєю середнього розміру.
Якщо викопна історія відмовляється демонструвати очікувані зв'язки, що вона демонструє?І що ж вона доводить?
Викопна історія
Викопна історія в тому вигляді, як вона нам відома, починається в період, який називається геологами кембрійським, який, за їх підрахунками, був приблизно 590 мільйонів років тому. Декілька крихітних скам'янілих останків було виявлено в породах більш раннього часу: кілька бактерій і кілька дуже незвичайних істот, не схожих ні на що з того, що було знайдено до або після - едіакарська фауна, вік якої налічує близько 565 мільйонів років.
Але всі вони, мабуть, вимерли невдовзі після цього. Складається враження, що у книзі життя було написано кілька тренувальних вправ, перекреслених потім жирною лінією: з цього моменту почалася реальна еволюція - чи, по крайнього заходу, почалося щось.
І це щось мало драматичний характер: щодо тваринного царства, то все з'явилося одночасно. Настільки раптовою і загадковою була поява різноманітності форм життя в той час, що вчені, як ми бачили, говорять про кембрійський вибух, який, за їхніми даними, стався близько 530 мільйонів років тому.
Найдивовижнішим відкриттям стало те, що тоді зародилися тварини всіх відомих форм - викопні чи нині. У цей період життя обрало свої основні форми і більше їх не змінювало.
Більше того, хоча вважають, що повністю період кембрію тривав близько 85 мільйонів років, але фактична поява всіх цих нових форм, ймовірно, відбулася приблизно за 10 мільйонів років або менше.
Іншими словами, історія життя на Землі виявляє близько 2 відсотків творчості та 98 відсотків подальшого розвитку.
Спрощена схема класифікації тварин НИНІ ЖИВІ ОРГАНІЗМИ| Тваринне царство| ФІЛЮМ/Eumetazoa(Справжні багатоклітинні)| ТИП/Chordata (хордові)| ПІДТИП/Vertebrata (хребетні)| КЛАС/Mammalia (ссавці)| ОТРЯД/Carnivora (грецькі)| Сімейство/Felidae (котячі)| РІД/Felis (кішки)| ВИД/catus > Felis catus (домашня кішка)Саме за своєю будовою вперше було класифіковано всі живі істоти. Була розроблена складна система, яка поділяє всі форми життя на два величезні царства - тваринне царство та рослинне царство. Вони, своєю чергою, поділяються спочатку на филюмы (від грецького слова «плем'я»), та був на дедалі дрібніші одиниці, до видів і підвидів.
Тварина царство зазвичай ділять на тридцять сім філумів. Всі ці філюми виникли під час кембрію. З того часу еволюція йшла лише по лінії видозміни основного плану. Крім того, немає жодних свідчень будь-якого попереднього їх розвитку. Немає жодних даних про те, що вони «еволюціонували» у дарвінівському розумінні цього терміна. Всі вони з'явилися в викопній історії в готовому вигляді - повністю сформованими істотами зі своїми виразними ознаками.
Вчені перебувають у здивуванні. Звертаючи нашу увагу на той факт, що «всі еволюційні зміни з часу кембрія були лише варіаціями на ті самі базові теми», професор Джеффрі Левінтоп з Нью-Йоркського університету запитує: «Чому ж давні форми такі стійкі?» Він не має відповіді.
Що вельми випливає з геологічних даних, так це те, що така стабільність є нормою. Викопні форми тварин або рослин з'являються, існують та розвиваються мільйони років, а потім зникають – проте будова їх мало змінюється.
Якщо й спостерігаються якісь зміни, то вони мають поступовий характер і обмежуються переважно розмірами: збільшується вся тварина чи рослина – чи окремі її ознаки. Не спостерігається того, щоб одна форма змінювалася на іншу, навіть відносно близьку: миша ніколи не еволюціонувала в щура; горобець ніколи не ставав дроздом.
До того ж подібні зміни мають, зважаючи на все, досить виборчий характер. Величезна кількість і до сьогодні істот, що мешкають на Землі, не зазнали жодних значних змін у своїй будові за весь тривалий час свого існування. Це йде врозріз з усіма очікуваннями Дарвіна.
Устриці та двостулкові молюски зараз мають раковини: вони з'явилися вперше близько 400 мільйонів років тому. Целакант і риби, що дихають, мешкають на Землі без будь-яких істотних змін вже близько 300 мільйонів років. Акули зберігають свій нинішній вигляд уже 150 мільйонів років. Осетр, каймана черепаха, алігатори та тапіри - всі ці види демонструють завидну стабільність форми вже понад 100 мільйонів років.
Сучасні опосуми відрізняються від тих, що мешкали 65 мільйонів років тому, тільки в незначних рисах. Перша черепаха мала той самий панцир, що й сьогодні; перші змії майже нічим не відрізняються від сучасних змій; кажани теж мало змінилися, як і і жаби і саламандры.
Що ж у такому разі еволюція зупинилася? Чи діє якийсь інший механізм чи чинник?
Приклад, який часто використовується для демонстрації еволюції, - кінь. Передбачається, що вона почалася з маленького чотирипалого гіракотерію, який жив 55 мільйонів років тому і розвинувся в сучасну Equus,що живе вже близько 3 мільйонів років. Усюди можна спостерігати елегантні та переконливі схеми та музейні експозиції, що зображують поступальну еволюцію коня. Вони майстерно демонструють, як пальці поступово звелися до одного, як помітно збільшився розмір тварини та як із зміною раціону змінилися зуби.
Однак тепер експерти, як правило, визнають, що ця лінія повільного, але вірного перетворення тварини розміром із собаку на сьогоднішнього великого коня є «переважно апокрифічним». Проблема в тому - і це звичайна проблема реконструкції еволюції за викопними даними, - що є безліч прогалин між різними видами викопного коня, які включені до цього ряду.
Починаючи з першого виду, гіракотерія,чий власний предок залишається загадкою, невідомо жодного зв'язку з передбачуваним «другим» конем і так далі.
Те, що ми маємо, не є лінією розвитку, це не є навіть генеалогічним деревом, що приводить до сучасної Equus,але є величезний кущ, у якого очевидні лише кінчики численних гілок, а всяке питання щодо існування його стовбура залишається відкритим.
У будь-який окремо взятий період часу існувало кілька видів коня - одні з чотирма пальцями, інші з меншим їх числом, одні з великими зубами, інші з маленькими. Коні також спочатку збільшились у розмірах, потім зменшилися, а потім знову збільшилися. І як постійне джерело роздратування - відсутність видів, що об'єднуються.
Нарешті, ми також повинні визнати, що передбачуваний кінь-предок не так сильно відрізняється від сучасного коня. Крім кількох другорядних змін щодо ступнів і зубів і збільшення розміру, мало істотно змінилося.
Це дуже незначна відмінність, яка подається як доказ еволюції, навіть якщо вона і відповідає дійсності, навряд чи вражає на тлі тих 52 мільйонів років, які пішли на це. Говорячи прямо, розглядати цю псевдопослідовність як доказ еволюції - це більшою мірою акт віри, ніж науковий факт.
Раптове походження видів
Викопну історію характеризують два моменти. Перше, як ми вже бачили, стабільність рослинних чи тваринних форм, коли вони вже з'явилися. Друге - раптовість, з якою ці форми виникають і, власне кажучи, надалі зникають.
Точність викопної історії
Загальна кількість наземних хребетних 43
Загальна кількість зафіксованих у викопній історії 42
Таким чином, відсоток виявлених копалин 97,7%
Загальна кількість сімейств, що живуть наземних хребетних 329
Загальна кількість зафіксованих у викопній історії 261
Таким чином, відсоток виявлених копалин 79,3%
Ми можемо дійти невтішного висновку, що викопна історія дає точну статистичну картину форм життя, які існували Землі. Отже, апелювання до неповноти викопних даних, як спосіб пояснення прогалин, не дуже переконливо.
Нові форми виникають у викопній історії, не маючи очевидних предків; так само несподівано вони зникають, не залишаючи якихось очевидних нащадків. Можна сміливо сказати, що майже копалини свідчення є історію величезної ланцюжка творів, об'єднаних лише вибором форми, а чи не еволюційними зв'язками.
Професор Гулд так підсумовує ситуацію: «У кожному окремому районі вид не виникає поступово шляхом планомірної трансформації його предків; він з'являється раптом і одразу і „повністю сформованим“».
Ми можемо спостерігати цей процес майже повсюдно. Коли, скажімо, близько 450 мільйонів років тому з'явилися перші копалини наземні рослини, то вони виникли без ознак попереднього розвитку. І проте навіть у ту ранню епохує всі основні різновиди.
Відповідно до теорії еволюції цього не може бути - якщо тільки ми не припустимо, що жодназ очікуваних сполучних форм не фосилізувалася, тобто. не перетворилася на скам'янілість. Що є досить малоймовірним.
Те саме з квітучими рослинами: хоча період, що передував їх появі, відрізняється великою різноманітністю копалин, не було знайдено жодних форм, які могли б бути їхніми предками. Їхнє походження також залишається незрозумілим.
Та сама аномалія виявляється і в тваринному царстві. Риби з хребтом та мозком вперше з'явилися близько 450 мільйонів років тому. Їхні прямі предки невідомі. І додатковим ударом по еволюційній теорії виявляється те, що у цих перших безщелепних риб, що мали панцир, був частково кістковий скелет.
Зазвичай викладена картина еволюції хрящового скелета (як акул і скатів) в кістковий скелет є, відверто кажучи, невірною. Насправді ці риби, що не мають кісткового скелета, з'являються в викопній історії на 75 мільйонів років пізніше.
Крім того, істотним етапом у передбачуваній еволюції риб було розвиток щелеп. Однак перша щелепна риба в викопній історії з'явилася раптово, при цьому неможливо вказати на будь-яку ранню безщелепну рибу як на джерело її майбутньої еволюції.
Ще одна дивина: міноги – безщелепні риби – чудово існують і досі. Якщо щелепи давали таку еволюційну перевагу, то чому тоді не вимерли ці риби? Не менш загадковим є і розвиток амфібій - водних тварин, здатних при цьому дихати повітрям і жити на суші. Як пояснює у своїй книзі «За межею природного відбору» д-р Роберт Вессон:
«Етапи, на яких риби дали життя земноводним, невідомі… найперші сухопутні тварини з'являються з чотирма добре розвиненими кінцівками, плечовим та тазовим поясом, ребрами та чітко вираженою головою… Через кілька мільйонів років, понад 320 мільйонів років тому, у викопній історії несподівано з'являється дюжина загонів земноводних, причому жоден, мабуть, не є предком якогось іншого».
Ссавці демонструють ту ж раптовість і стрімкість розвитку. Найбільш ранні ссавці були маленькими тваринами, що вели потайливий спосіб життя в еру динозаврів - 100 або більше мільйонів років тому. Потім, після загадкового і все ще не поясненого вимирання останніх (близько 65 мільйонів років тому), у викопній історії в один і той же час – близько 55 мільйонів років тому – з'являється дюжина із зайвим груп ссавців.
Серед копалин цього періоду знаходять скам'янілі зразки ведмедів, левів і кажанів, що мають сучасний вигляд. І що ще більше ускладнює картину – вони з'являються не в одному якомусь районі, а одночасно в Азії, Південній Америці та Південній Африці. На додачу, до всього цього, немає впевненості, що дрібні ссавці епохи динозаврів і справді були предками пізніших ссавців.
Вся викопна історія рясніє пробілами та загадками. Невідомо, наприклад, жодних копалин між першими хребетними і примітивними істотами раннього періоду - хордовими, - яких вважають предками хребетних.
Існуючі сьогодні земноводні разюче відрізняються від перших відомих амфібій: між цими древніми і пізнішими формами в викопній історії сяє прогалина в 100 мільйонів років. Як видається, дарвінівська теорія еволюції буквально на очах розсипається на порох. Ймовірно, якось можна врятувати дарвінівську ідею «природного відбору», але лише у видозміненій формі.
Зрозуміло, що немає свідчень розвитку будь-яких нових форм рослин чи тварин. Лише коли жива форма з'явилася, тоді тільки можливо грає свою роль природний відбір. Але він працює тільки на тому, що вже існує.
Не лише вчені, а й студенти коледжів та університетів проводять селекційні експерименти на плодовій мушці – дрозофілі. Їм пояснюють, що демонструють наочний доказ еволюції. Вони створюють мутації виду, дають їй очі різного забарвлення, ніжку, що росте з голови, або, можливо, подвійний торакс.
Можливо, їм навіть вдається виростити мушку із чотирма крилами замість звичайних двох. Однак ці зміни - лише модифікація вже існуючих видових ознак мушки: чотири крила, наприклад, не більше ніж подвоєння початкових двох. Ніколи не вдавалося створити якийсь новий внутрішній орган, як не вдавалося перетворити плодову мушку на щось, що нагадує бджолу чи метелика. Неможливо навіть перетворити її на інший вид мухи.
Як і завжди, вона залишається представником роду Drosophila.«Природний відбір, можливо, і пояснює походження адаптаційних змін, але не може пояснити походження видів». І це обмежене застосування стикається з проблемами.
Як, наприклад, природний відбір здатний пояснити той факт, що люди – єдиний вид живих істот – мають різні групи крові? Як він здатний пояснити те, що один із найраніших відомих науці викопних видів - трилобіт кембрійського періоду - має око з таким складним пристроєм і настільки ефективний, що не був перевершений ніяким пізнішим представником його філюму? І як могли еволюціонувати пір'я? Д-р Барбара Сталь, автор академічної праці з еволюції, зізнається: «Як виникли, імовірно з луски рептилій, - аналізу не піддається».
Вже на початку Дарвін розумів, що зіткнувся з глибокими проблемами. Розвиток складних органів, наприклад, до краю підривало його теорію. Бо доти, доки такий орган не почав функціонувати, за якою потребою мав заохочувати його розвиток до природного відбору? Як запитує професор Гулд: «Яка користь від недосконалих стадій зачатків, що дають перевагу структур? Яка користь від півщелепи чи півкрила?» Чи, можливо, від півочі? Те саме питання виникло десь і у свідомості Дарвіна. У 1860 році він зізнався колезі: «Око до цього дня приводить мене в холодне тремтіння». І не дивно.
Пропонована еволюція хребетних. На цій схемі представлено різноманіття груп хребетних, що поширилися з часів. Пунктирні лінії позначають відсутні ланки, яких вимагає - для того, щоб зв'язати між собою ці групи - еволюційна теорія. У викопній історії ці ланки не виявлено.
Останнім прикладом - якщо хочете, доказом - того, що природний відбір (якщо він і справді реально діє механізм змін) вимагає більшого розуміння, є факт, що стосується фізіологічних відправлень у лінивця, який наводить д-р Вессон:
«Замість того, щоб відразу ж справляти потребу, подібно до інших мешканців дерев, лінивець зберігає свої фекалії протягом тижня або більше, що нелегко для тварини, яка харчується грубою рослинною їжею. Після чого він спускається на землю, яку в інших випадках не ступає, випорожнюється і закопує екскременти.
Передбачається, що ця пов'язана з неабиякою небезпекою звичка має ту еволюційну перевагу, що тим самим відбувається удобрення дерев'яного будинку. Тобто низка випадкових мутацій призвела до того, що у лінивця розвинулася не схожа на нього звичка при відправленні фізіологічних потреб і що це настільки покращило якість листя облюбованого ним дерева, що викликало появу в нього більш численних нащадків, ніж у лінивців, які випорожнювались прямо на деревах ... »
Або в еволюції є інші форми чи способи «природного відбору», про які ми поки що навіть не здогадуємося, або для пояснення раптового розкиду в викопній історії необхідно використовувати щось зовсім інше - можливо, космічне почуття гумору?
Неправильна еволюція
Про проблеми з копалинами було відомо від початку. Протягом століття або трохи довше вчені просто сподівалися на те, що проблеми мають тимчасовий характер, що будуть зроблені відкриття, які заповнять прогалини. Або, можливо, буде знайдено доказ того, що причина цих прогалин не в проблемах з еволюцією, а в нерегулярності геологічного процесу.
Зрештою, однак, терпіння стало вичерпуватися. Згода в науковому світі була порушена в 1972 році, коли Стівен Джей Гулд і Найлс Елдрідж представили на конференції з еволюції спільну доповідь, що мала революційний характер. Їхня доповідь прямо спростовувала дарвінівську теорію.
Вони висловили твердження, що, хоча викопні дані, безумовно, аж ніяк не задовільні, раптові появи нових видів, що спостерігаються, не є свідченням неповноти викопних даних, - навпаки, вони відображають реальність. Походження видів могло бути не поступовим еволюційним процесом, а процесом, у якому тривалі періоди стабільності зрідка перемежовувалися раптовими масштабними змінами живих формах. За допомогою цього аргументу Гулд і Елдрідж могли пояснити відсутність «ланок, що бракують»: вони стверджували, що їх просто не було.
Як би добре ця ідея не пояснювала, можливо, викопну історію, вона, як і раніше, базується на уявленні про те, що розвиток життя носить безладний, випадковий характер. Однак може бути продемонстровано, що еволюція, хоч би яким чином вона відбувалася, навряд чи була випадковим процесом.
Програми розвитку для рослинних та тваринних форм містяться в генетичному коді. Цей код дуже складний, а кількість варіацій, які могли б бути задіяні, величезна. Чи міг цей код еволюціонувати випадково? Просте знайомство із цифрами показує, що цього не могло бути. Якби, наприклад, мавпа сиділа за машинкою, щосекунди стукаючи навмання по клавішах, то скільки б знадобилося часу, щоб у мавпи - випадково - вийшло осмислене слово з дванадцяти букв? Для цього їй знадобилося б майже 17 мільйонів років.
Скільки б часу знадобилося все тій же мавпі, щоб у неї - випадково - вийшла осмислена пропозиція зі 100 букв - ланцюжок знаків, куди менш складний, ніж генетичний код? Імовірність цього настільки низька, що шанси проти неї перевищують загальну кількість атомів у всьому Всесвіті. Фактично слід говорити про неможливість того, щоб випадковим чином могла вийти осмислена послідовність зі 100 символів. Залишається зробити висновок, що так само неможливо і те, щоб випадково міг вийти складний генетичний код життя, як того вимагає теорія еволюції.
Астроном Фред Хойл, з властивою йому влучністю писав, що ймовірність випадкового створення вищих форм життя подібна до ймовірності того, щоб «торнадо, що проноситься по смітнику, міг зібрати „Боїнг-747“».
А в такому випадку, якщо генетичний код не створений випадковим процесом, тоді він, мабуть, створений невипадковим процесом. До чого могла б привести нас ця думка?
Спрямована еволюція
У 1991 році книга Вессона "За межею природного відбору" стала новим і потужним викликом, кинутим офіційній науці. Він відкинув прихильність до дарвінівської еволюції як «поточку стародавньої мрії про Всесвіт, уподібнений величезному годинниковому механізму». Вессон вказує, що не можна розглядати будь-яку тварину окремо.
Він пропонує нам глянути ширше: «Організми еволюціонують як частину спільності, тобто як екосистема... яка неминуче еволюціонує спільно. Швидше треба говорити не про походження видів, а про розвиток екосистем…»
Проводячи воістину радикальний перегляд, Вессон пропонує застосувати до еволюції висновки теорії хаосу, щоб зрозуміти зміст усіх тих дивовижних і дивних явищ, які ми спостерігаємо як у викопних даних, і у нині існуючих організмах.
5. Викопні
6. Еквус, або кінь справжній.
З книги М. Бейджента „Заборонена археологія”.
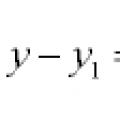 Рівняння прямих та кривих на площині
Рівняння прямих та кривих на площині Узагальнене однорідне рівняння узагальнено однорідне рівняння другого порядку
Узагальнене однорідне рівняння узагальнено однорідне рівняння другого порядку Завдяки кома Завдяки тому що в них
Завдяки кома Завдяки тому що в них