Походження та еволюція наземних рослин. Докази тваринного походження людини
Виникнення одноклітинних та багатоклітинних водоростей, виникнення фотосинтезу: вихід рослин на сушу (псилофіти, мохи, папороті, голонасінні, покритонасінні).
Розвиток рослинного світу відбувалося в 2 етапи і пов'язане з появою нижчих та вищих рослин. За новою систематикою до нижчих відносять водорості (а раніше відносили бактерії, гриби та лишайники. Тепер вони виділені в самостійні царства), а до вищих – мохи, папоротькоподібні, голонасінні та покритонасінні.
В еволюції нижчих організмів виділяються 2 періоди, що істотно різняться між собою організацією клітини. Протягом 1 періоду панували організми, подібні до бактерій і синьо-зелені водоростями. Клітини цих життєвих форм не мали типових органоїдів (мітохондирій, хлоропластів, апарату Гольджі та ін.). Ядро клітини не було обмежене ядерною мембраною (це прокаріотичний тип клітинної організації). 2 період був пов'язаний з переходом нижчих рослин (водоростей) до автотрофного типу харчування та з утворенням клітини з усіма типовими органоїдами (це еукаріотичний тип клітинної організації, який зберігся і на наступних щаблях розвитку рослинного та тваринного світу). Цей період можна назвати періодом панування зелених водоростей, одноклітинних, колоніальних та багатоклітинних. Найпростішими з багатоклітинних є нитчасті водорості (улотрикс), які мають ніякого розгалуження свого тіла. Їх тіло є довгим ланцюжком, що складається з окремих клітин. Інші ж багатоклітинні водорості розчленовані великою кількістювиростів, тому їхнє тіло гілкується (у хари, у фукуса).
Багатоклітинні водорості у зв'язку з їх автотрофною (фотосинтетичною) діяльністю розвивалися у напрямку збільшення поверхні тіла для кращого поглинання поживних речовин з водного середовища та сонячної енергії. У водоростей з'явилася прогресивніша форма розмноження - статеве розмноження, при якому початок новому поколінню дає диплоїдна (2н) зигота, що поєднує в собі спадковість 2-х батьківських форм.
2 Еволюційний етап розвитку рослин необхідно пов'язувати з поступовим переходом їх від водного способу життя до наземного. Первинним наземним організмами виявилися псилофіти, які збереглися як копалин залишків в силурійських і девонських відкладеннях. Будова цих рослин складніша порівняно з водоростями: а) вони мали спеціальні органи прикріплення до субстрату - різоїди; б) стеблоподібні органи з деревиною, оточеною лубом; в) зачатки провідних тканин; г) епідерміс з продихами.
Починаючи з псилофітів, потрібно простежити 2 лінії еволюції вищих рослин, одна з яких представлена мохоподібними, а друга - папоротеподібними, голонасінними та покритонасінними.
Головне, що характеризує мохоподібні, це переважання у циклі їхнього індивідуального розвитку гаметофіту над спорофітом. Гаметофіт – це все зелена рослиназдатне до самостійного харчування. Спорофіт представлений коробочкою (зозулин льон) і повністю залежить у своєму харчуванні від гаметофіту. Домінування у мохів вологолюбного гаметофіта в умовах повітряно-наземного способу життя виявилося нецелеособно, тому мохи стали особливою гілкою еволюції вищих рослин і поки не дали після себе досконалих груп рослин. Цьому сприяв і те що, що гаметофит проти спорофитом мав обідню спадковість (гаплоидный (1н) набір хромосом). Ця лінія в еволюції вищих рослин називається гаметофітною.
Друга лінія еволюції на шляху від псилофітів до покритонасінних є спорофітною, тому що у папоротеподібних, голонасінних та покритонасінних у циклі індивідуального розвитку рослин домінує спорофіт. Він являє собою рослину з коренем, стеблом, листям, органами спороношення (у папороті) або плодоношення (у покритонасінних). Клітини спорофіту мають диплоїдний набір хромосом, т.к. вони розвиваються з диплоїдної зиготи. Гаметофіт сильно редукований та пристосований тільки для утворення чоловічих та жіночих статевих клітин. У квіткових рослин жіночий гаметофіт представлений зародковим мішком, в якому знаходиться яйцеклітина. Чоловічий гаметофіт утворюється під час проростання пилку. Він складається з однієї вегетативної та однієї генеративної клітин. При проростанні пилку із генеративної клітини виникає 2 спермія. Ці 2 чоловічі статеві клітини беруть участь у подвійному заплідненні у покритонасінних. Запліднена яйцеклітина дає початок новому поколінню рослини - спорофіту. Прогрес покритонасінних обумовлений удосконаленням функції розмноження.
|
Висновки:
1. Вивчення геологічного минулого Землі, будови та складу ядра та всіх оболонок, польоти космічних апаратівна Місяць, Венеру, вивчення зірок наближає людину до пізнання етапів розвитку нашої планети та життя на ній.2. Процес еволюції мав природний характер.
3. Рослинний світ різноманітний, це різноманіття є результатом його розвитку протягом тривалого часу. Причина його розвитку - не божественна сила, - а зміна та ускладнення будови рослин під впливом змінних умов довкілля.
Наукові докази: клітинна будова рослин, початок розвитку з однієї заплідненої клітини, необхідність води для життєвих процесів, знаходження відбитків різних рослин, наявність "живих" копалин, вимирання деяких видів та утворення нових.
на тему: «Біоценози та екосистеми»
ВЛАСТИВОСТІ ТА ТИПИ БІОЦЕНОЗІВ
Природні біоценози дуже складні. Вони характеризуються, перш за все, видовою різноманітністю та щільністю популяції.
Видова різноманітність- Число видів живих організмів, що утворюють біоценоз і визначають різні харчові рівні в ньому. Чисельність видових популяцій визначається кількістю особин даного виду на одиниці площі. Деякі види є у співтоваристві домінантними, перевищуючи чисельність інші. Якщо спільноті домінують кілька видів, а щільність інших дуже мала, то розмаїтість низька. Якщо при тому самому видовому складі чисельність кожного з них більш-менш вирівняна, то видова різноманітність висока.
Крім видового складу біоценоз характеризується біомасою та біологічною продуктивністю.
Біомаса- Загальна кількістьорганічної речовини та укладеної в ній енергії всіх особин цієї популяції або всього біоценозу, що припадає на одиницю площі. Біомаса визначається кількістю сухої речовини на 1 га або кількістю енергії (Дж) 1 .
Розмір біомаси залежить від особливостей виду, його біології. Наприклад, у швидковідмираючих видів (мікроорганізмів) біомаса невелика в порівнянні з довгоживучими організмами, що накопичують у своїх тканинах велику кількість органічних речовин (дерева, чагарники, великі тварини).
Біологічна продуктивність- Швидкість утворення біомаси в одиницю часу. Це найбільш важливий показникжиттєдіяльності організму, популяції та екосистеми в цілому. Розрізняють первинну продуктивність – утворення органічної речовини автотрофами (рослинами) у процесі фотосинтезу та вторинну – швидкість утворення біомаси гетеротрофами (консументами та редуцентами).
Співвідношення продуктивності та біомаси по-різному у різних організмів. Крім того, у різних екосистемах продуктивність неоднакова. Вона залежить від величини сонячної радіації, ґрунти, клімату. Найнижча біомаса і продуктивність мають пустелі і тундру, найвищою - дощові тропічні ліси. Порівняно із сушею біомаса Світового океану значно нижча, хоча вона займає 71% поверхні планети, що пов'язано з низьким вмістом поживних речовин. У прибережній зоні біомаса значно зростає.
У біоценозах розрізняють два типи трофічної мережі: пасовищну та детритну. У пасовищному типіхарчової мережі енергія йде від рослин до рослиноїдних тварин, а далі до консументів вищого порядку. Травоїдні тварини, незалежно від їх величини та довкілля (наземні, водні, грунтові), пасуться, виїдають зелені рослини і передають енергію на наступні рівні.
Якщо потік енергії починається з мертвих рослинних і тваринних залишків, екскрементів і йде до первинних детритофагів - редуцентів, що частково розкладають органічні речовини, то така трофічна мережа називається детритної,або мережею розкладання. До первинних детрітофагів відносяться мікроорганізми (бактерії, гриби) і дрібні тварини (хробаки, личинки комах).
У наземних біогеоценозах присутні обидва типи трофічної мережі. У водних угрупованнях переважає ланцюг виїдання. І в тому, і в іншому випадку енергія використовується повністю.
Еволюція екосистем
СУКЦЕСІЇ
Усі екосистеми еволюціонують у часі. Послідовна зміна екосистем називається екологічну сукцесію.Сукцесія відбувається головним чином під впливом процесів, що протікають усередині спільноти при взаємодії із навколишнім середовищем.
Первинна сукцесія починається з освоєння середовища, яке раніше була житла: зруйнована гірська порода, скеля, піщана дюна тощо. буд. Тут велика роль перших поселенців: бактерій, ціанобактерій, лишайників, водоростей. Виділяючи продукти життєдіяльності, вони змінюють материнську породу, руйнують її та сприяють ґрунтоутворенню. Відмираючи, первинні живі організми збагачують поверхневий шар органічними речовинами, що дозволяє оселятися іншим організмам. Вони поступово створюють умови для більшого розмаїття організмів. Спільнота рослин та тварин ускладнюється, поки не досягає певної рівноваги із середовищем. Таку спільноту називають клімаксовим.Воно підтримує свою стабільність доти, доки порушується рівновагу. Ліс є стійкий біоценоз - клімаксове співтовариство.
Вторинна сукцесія розвивається дома вже наявного раніше сформованого співтовариства, наприклад, дома пожарища чи занедбаного поля. На згарищі поселяються світлолюбні рослини, під їх пологом розвиваються тіньовитривалі види. Поява рослинності покращує стан ґрунту, на якому починають рости вже інші види, витісняючи перших поселенців. Вторинна сукцесія відбувається в часі і в залежності від ґрунту може бути швидкою або повільною, поки, нарешті, не утворюється клімаксова спільнота.
Озеро, при порушенні в ньому екологічної рівноваги, може перетворитися на луг, а потім і на ліс, характерний для даної кліматичної зони.
Сукцесія призводить до послідовного ускладнення спільноти. Його харчові мережістають дедалі більше розгалуженими, дедалі повніше використовуються ресурси середовища. Зріле співтовариство найбільше пристосовано до умов середовища, популяції видів стабільні і добре відтворюються.
Штучні екосистеми. АГРОЦЕНОЗИ
Агроценоз- штучно створені та підтримувані людиною екосистеми (поля, сіножаті, парки, сади, городи, лісові посадки). Їх виробляють для отримання сільськогосподарської продукції. Агроценози мають погані динамічні якості, малу екологічну надійність, але характеризуються високою врожайністю. Займаючи приблизно 10% площі суші, агроценози щорічно виробляють 2,5 млрд. т сільськогосподарської продукції.
Як правило, в агроценозі культивується один або два види рослин, тому взаємозв'язки організмів не можуть забезпечити сталість такого співтовариства. Дія природного відбору ослаблена людиною. Штучний відбір йде у напрямку збереження організмів із максимальною продуктивністю. Крім сонячної енергії в агроценозі є й інше джерело - мінеральні та органічні добрива, що вносяться людиною. Основна частина поживних речовин постійно виноситься з круговороту як урожай. Таким чином, кругообіг речовин не здійснюється.
В агроценозі, як і в біоценозі, складаються харчові ланцюги. Обов'язковою ланкою цього ланцюга є людина. Причому тут він постає як консумент I порядку, але цьому харчова ланцюг переривається. Агроценози дуже нестійкі і без участі людини існують від 1 року (зернові, овочеві) до 20-25 років (плодово-ягідні).
РОЗВИТОК БІОЛОГІЇ У ДОДАРВІНІВСЬКИЙ ПЕРІОД
Зародження біології як науки пов'язані з діяльністю грецького філософа Аристотеля (IV в. е.). Він намагався побудувати класифікацію організмів на основі анатомічних та фізіологічних досліджень. Йому вдалося описати майже 500 видів тварин, яких він розташував як ускладнення. Вивчаючи ембріональний розвиток тварин, Аристотель виявив велику схожість початкових стадійЕмбріогенез і прийшов до думки про можливість єдності їх походження.
У період із XVI по XVIII ст. йде інтенсивний розвиток описової ботаніки та зоології. Відкриті та описані організми вимагали систематизації та введення єдиної номенклатури. Ця заслуга належить видатному вченому Карлу Ліннею (1707-1778). Він уперше звернув увагу на реальність виду як структурної одиниціживої природи. Він запровадив бінарну номенклатуру виду, встановив ієрархії одиниць систематики (таксонів), описав та систематизував 10 тис. видів рослин та 6 тис. видів тварин, а також мінерали. На свій світогляд К. Лінней був креаціоністом. Він відкидав ідею еволюції, вважаючи, що видів стільки, скільки різних формбуло створено богом на початку. Наприкінці життя К. Лінней все ж погодився з існуванням мінливості в природі, віра в незмінність виду була трохи похитнута.
Автором першої еволюційної теоріїбув французький біолог Жан-Батіст Ламарк (1744-1829). Ламарк увічнив своє ім'я, ввівши термін «біологія», створивши систему тваринного світу, де вперше розділив тварин на «хребетних» та «безхребетних». Ламарк вперше створив цілісну концепцію розвитку природи та сформулював три закони змінності організмів.
1. Закон прямого пристосування. Пристосувальні зміни рослин та нижчих тварин відбуваються під прямим впливом довкілля. Пристосування виникають за рахунок подразливості.
2. Закон вправи та невправи органів. На тваринах з центральною нервовою системоюсередовище надає опосередкований вплив. Тривалий вплив середовища викликає у тварин звички, пов'язані з найчастішим вживанням органів. Посилена його вправа призводить до поступового розвитку цього органу та закріплення змін.
3. Закон «успадкування набутих ознак», згідно з яким корисні зміни передаються та закріплюються в потомстві. Цей процес має поступовий характер.
Неперевершеним авторитетом в XIX ст. в галузі палеонтології та порівняльної анатомії був французький зоолог Жорж Кюв'є (1769-1832). Він був одним із реформаторів порівняльної анатомії та систематики тварин, запровадив поняття «тип» у зоології. Ґрунтуючись на багатому фактичному матеріалі, Кюв'є встановив «принцип кореляції частин тіла», на базі якого реконструював будову вимерлих форм тварин. На свої погляди він був креаціоністом і стояв на позиціях незмінності видів, а наявність пристосувальних ознак у тварин розглядав як свідчення спочатку встановленої гармонії в природі. Причини зміни копалин фаун Ж. Кюв'є бачив у катастрофах, що відбувалися на Землі. За його теорією, після кожної катастрофи відбувалося повторне створення органічного світу.
ОСНОВНІ ПОЛОЖЕННЯ ТЕОРІЇ Ч. ДАРВІНА
Честь створення наукової теорії еволюції належить Чарлзу Дарвіну (1809-1882) - англійському натуралісту. Історичною заслугою Дарвіна є встановлення самого факту еволюції, а розтин основних причин і рушійних сил її. Він ввів термін «природний відбір» і довів, що основою природного відбору та еволюції є спадкова мінливість організмів. Результатом його багаторічної роботи стала книга «Походження видів шляхом природного відбору» (1859). У 1871 р. виходить у світ його інший велика праця«Походження людини та статевий добір».
Основними рушійними силами еволюціїЧ. Дарвін назвав спадкову мінливість, боротьбу за існуванняі природний відбір.Вихідним становищем вчення Дарвіна було його твердження про мінливість організмів. Він виділив групову, чи певну, мінливість, яка успадковується і у прямої залежності від чинників середовища. Другий тип мінливості - індивідуальна, чи невизначена, що виникає в окремих організмів у результаті невизначених впливів середовища на кожну особину і успадковується. Саме ця мінливість є основою різноманітності особин.
Спостерігаючи та аналізуючи одну з основних властивостей всього живого - здатність до необмеженого розмноження, Дарвін зробив висновок про існування фактора, що перешкоджає перенаселенню та обмежує чисельність особин. Висновок:інтенсивність розмноження, а також обмеженість природних ресурсівта засобів життя призводять до боротьби за існування.
Наявність спектру мінливості в організмів, їх неоднорідність та боротьба за існування призводять до виживання найбільш пристосованих та знищення менш пристосованих особин. Висновок:у природі йде природний відбір, який сприяє накопиченню корисних ознак, передачі та закріпленню їх у потомстві. Ідея природного відбору виникла у Дарвіна внаслідок спостережень за штучним відбором та селекцією тварин. По Дарвіну, результатом природного відбору у природі стали:
1) виникнення пристроїв;
2) змінність, еволюція організмів;
3) утворення нових видів. Видоутворення йде з урахуванням дивергенції ознак.
Дивергенція- розходження ознак у межах виду, що виникає під впливом природного добору. Найбільші переваги виживання мають особини з крайніми ознаками, тоді як особини з середніми, подібними, ознаками гинуть у боротьбі існування. Організми з ознаками, що ухиляються, можуть стати родоначальниками нових підвидів і видів. Причиною дивергенції ознак є наявність невизначеної мінливості, внутрішньовидова конкуренція та різноспрямований характер дії природного відбору.
Теорія видоутворення Дарвіна називається монофілетичною – походження видів від загального родоначальника, вихідного виду. Ч. Дарвін довів історичний розвитокживої природи, пояснив шляхи видоутворення, обґрунтував формування пристроїв та їх відносний характер, визначив причини та рушійні силиеволюції.
ДОВІДКИ ЕВОЛЮЦІЇ
Біологічна еволюція- історичний процесрозвитку органічного світу, що супроводжується змінами організмів, вимиранням одних та появою інших. Сучасна наукамає у своєму розпорядженні багато фактів, що свідчать про еволюційні процеси.
Ембріологічні докази еволюції.
У першій половині ХІХ ст. отримує розвиток теорія «зародкової схожості». Російський учений Карл Бер (1792-1876) встановив, що на ранніх стадіяхрозвитку ембріонів виявляється велика схожість між різними видами в межах типу.
Роботи Ф. Мюллера та Е. Геккеля дозволили їм сформулювати біогенетичний закон:«онтогенез є коротке та швидке повторення філогенезу». Пізніше трактування біогенетичного закону було розвинуто і уточнено А. Н. Северцовим: «в онтогенезі повторюються ембріональні стадії предків». Найбільше подібність мають зародки ранніх стадіях розвитку. Загальні ознаки типу формуються під час ембріогенезу раніше, ніж спеціальні. Так, всі ембріони хребетних на I стадії мають зяброві щілини та двокамерне серце. На середніх стадіях з'являються особливості, характерні кожного класу, і лише пізніших формуються особливості виду. Порівняльно-анатомічні та морфологічні докази еволюції.
Доказом єдності походження є клітинна будова організмів, єдиний план будови органів та їх еволюційні зміни.
Гомологічні органимають подібний план будівлі та спільність походження, виконують як однакові, і різні функції. Гомологічні органи дозволяють довести історичну спорідненість різних видів. Первинна морфологічна подібність замінюється, різного ступеня, відмінностями, набутими під час дивергенції Типовим прикладомгомологічних органів є кінцівки хребетних, що мають загальний планбудови незалежно від виконуваних функцій.
Деякі органи рослин морфологічно розвиваються з листових зачатків і є видозміненим листям (усики, колючки, тичинки).
Аналогічні органи- Вторинне, не успадковане від загальних предків, морфологічна схожість в організмів різних систематичних груп. Аналогічні органи подібні до виконуваних функцій і розвиваються в процесі конвергенції.Вони свідчать про однотипні пристрої, що виникають у ході еволюції в однакових умовах середовища в результаті природного відбору. Наприклад, аналогічні органи тварин – крила метелика та птиці. Це пристосування до польоту у метеликів розвинулося з хітинового покриву, а птахів - з внутрішнього скелета передніх кінцівок і пір'яного покриву. Філогенетично ці органи формувалися по-різному, але виконують однакову функцію – служать для польоту тварини. Іноді аналогічні органи набувають разючої подібності, як, наприклад, очі головоногих молюсків та наземних хребетних. Вони мають однаковий загальний план будови, схожі структурні елементи, хоч і розвиваються з різних зачатків в онтогенезі і ніяк не пов'язані між собою. Подібність пояснюється лише фізичною природою світла.
Прикладом аналогічних органів є колючки рослин, які захищають їхню відмінність від поїдання тваринами. Колючки можуть розвиватися з листя (барбарис), прилистків (біла акація), пагона (глоду), кори (ожина). Вони подібні лише зовні і за функціями, що виконуються.
Рудиментарні органи- Порівняно спрощені або недорозвинені структури, що втратили своє початкове призначення. Вони закладаються під час ембріонального розвитку, але не розвиваються. Іноді рудименти беруть він інші функції проти гомологічними органами інших організмів. Так, рудимент апендикс людини виконує функцію лімфотворення, на відміну від гомологічного органу – сліпої кишки травоїдних. Рудименти тазового пояса кита та кінцівок пітона підтверджують факт походження китів від наземних чотирилапих, а пітонів – від предків із розвиненими кінцівками.
Атавізм -явище повернення до предковим формам, що спостерігається в окремих особин. Наприклад, зеброїдна забарвлення лошат, багатососковість у людини.
Біогеографічні докази еволюції
Вивчення флори та фауни різних материків дозволяє відновити загальний перебіг еволюційного процесу та виділити кілька зоогеографічних зон, що мають подібні наземні тварини.
1. Голарктична область, яка поєднує Палеарктичну (Євразія) та Неоарктичну (Північна Америка) області. 2. Неотропічна область (Південна Америка). 3. Ефіопська область (Африка). 4. Індо-Малайська область (Індокитай, Малайзія, Філіппіни). 5. Австралійська область. У кожній із перерахованих областей спостерігається велика схожість тваринного та рослинного світу. Одна область від інших відрізняється від певних ендемічних груп.
Ендеміки- види, пологи, сімейства рослин чи тварин, поширення яких обмежено невеликий географічною областю, тобто це специфічна для цієї галузі флора або фауна. Розвиток ендемії найчастіше пов'язаний із географічною ізоляцією. Наприклад, найбільш раннє відділення Австралії від південного материкаГондвани (понад 120 млн років) призвело до самостійного розвитку низки тварин. Не відчуваючи тиску з боку хижаків, які відсутні в Австралії, тут збереглися однопрохідні ссавці-перозвірі: качконіс та єхидна; сумчасті: кенгуру, коала.
Флора та фауна Палеарктичної та Неоарктичної областей, навпаки, подібні між собою. Наприклад, близькими є американські та європейські клени, ясені, сосни, ялини. З тварин такі ссавці, як лосі, куниці, норки, білі ведмеді, мешкають у Північній Америці та Євразії. Американському бізону відповідає споріднений вигляд – європейський зубр. Подібна спорідненість свідчить про тривалу єдність двох материків.
Палеонтологічні докази еволюції
Палеонтологія вивчає викопні організми та дозволяє встановити історичний процес та причини зміни органічного світу. На основі палеонтологічних знахідок складено історію розвитку органічного світу.
Викопні перехідні форми -форми організмів, що поєднують ознаки більш давніх та молодих груп. Вони допомагають відновити філогенез окремих груп. Представники: археоптерикс – перехідна форма між рептиліями та птахами; іноземців - перехідна форма між рептиліями і ссавцями; псилофіти - перехідна форма між водоростями та наземними рослинами.
Палеонтологічні рядискладаються з копалин форм і відбивають перебіг філогенезу (історичного розвитку) виду. Такі ряди існують для коней, слонів, носорогів. Перший палеонтологічний ряд коней було складено В. О. Ковалевським (1842-1883).
Релікти- види рослин чи тварин, що збереглися від давніх зниклих організмів. Їх характерні ознаки вимерлих груп минулих епох. Вивчення реліктових форм дозволяє відновити вигляд зниклих організмів, припустити умови їх проживання та спосіб життя. Гаттерія - представник давніх примітивних плазунів. Такі плазуни мешкали в юрі та крейдяному періоді. Кістепера риба латимерія відома з раннього девону. Ці тварини дали початок наземним хребетним. Гінкго є найбільш примітивною формою голонасінних. Листя велике, віялоподібне, рослини листопадні.
Порівняння сучасних примітивних та прогресивних форм дає можливість відновити деякі риси передбачуваних предків прогресивної форми, проаналізувати перебіг еволюційного процесу.
Реферат на тему: «Біоценози та екосистеми» ВЛАСТИВОСТІ І ТИПИ БІОЦЕНОЗІВ Природні біоценози дуже складні. Вони характеризуються, перш за все, видовою різноманітністю та щільністю популяції. Видове розмаїття - число видів живихІІ. Ембріологічні докази (ембріологія вивчає зародковий розвиток організму).
1. Подібність зародків.
а) Будова зародка хордових послідовно нагадує тіло тварин інших типів:
· Яйцеклітина - найпростіших;
· Гаструла - кишковопорожнинних;
· Представників підтипу Безчерепні.
б) Це свідчить про спільність походження всіх хордових.
2. Розбіжність ознак зародків (ембріональна дивергенція).
а) З розвитком риси подібності між зародками різних видівслабшають.
б) Спочатку з'являються ознаки роду, та був виду.
· Початкова подібність у будові голови у дитини та дитинчата мавп поступово зникає.
3. Біогенетичний закон Геккеля-Мюллера: кожна особина в індивідуальному розвитку (онтогенезі) коротко і стисло повторює історію розвитку свого виду (філогенез).
а) Приклади у тварин:
· Судини зародків сухопутних хребетних схожі на судини риб;
· У людського зародка є зяброві щілини.
· Гусениці метелика та личинки жуків подібні за будовою з кільчастими хробаками.
· Пуголовки земноводних подібні з рибами.
б) Приклади у рослин:
· Ниркові лусочки у нирці рослин розвиваються як листя.
· Пелюстки бутонів спочатку зелені, а потім набувають властивого їм забарвлення.
· Зі суперечки моху спочатку з'являється зелена нитка, схожа на нитчасту водорість (предросток).
в) Поправки до біогенетичного закону.
· У зародків повторення філогенезу може порушуватися через пристосування до умов життя в онтогенезі. З'являються: зародкові оболонки, жовтковий мішок у ікринки риб, зовнішні зябра у пуголовка, кокон у шовкопряда.
· Онтогенез не повністю відображає філогенез за рахунок появи мутацій, що змінюють хід розвитку зародка (у зародка змії закладаються відразу всі хребці, тобто їх кількість не збільшується поступово; у птахів випала п'ятипала стадія розвитку кінцівки, у зародка закладаються 4 пальці, а не 5, виростають у крилі тільки 3 пальця).
· В онтогенезі відбувається повторення зародкових стадій розвитку, а не дорослих форм (Ланцетник повторює в онтогенезі загальні стадії з вільно личинкою, що плаває асцидії, а не з її дорослою, закріпленою формою).
г) Сучасні уявлення про біогенетичний закон.
· Северцов показав, що з допомогою змін у розвитку можуть: випадати деякі стадії розвитку зародка; виникати зміни органів зародка, яких у предків; виникати нові види; виявлятися нові ознаки (наприклад, хвостаті (тритони) та безхвості (жаби) амфібії походять від одного предка: личинка тритону довга, тому що має багато хребців, у личинки жаби кількість хребців зменшилася за рахунок мутації; у ембріона ящірки менше числохребців, ніж у ембріона змії, за рахунок мутацій розвитку).
ІІІ. Біогеографічні докази (Біогеографія вивчає розподіл тварин та рослин на Землі).
1. Існує 5 зоогеографічних зон, які не різняться за класами та типами тварин:
а) Голарктична;
б) Індо-Малазійська;
в) Ефіопська;
г) Австралійська;
д) Неотропічна зона.
2. Зони розрізняються за сімействами, загонами та пологами.
а) В Австралії всі ссавці сумчасті.
б) У Новій Зеландії живе єдиний представник загону дзьобоголових ящерів – гаттерію.
в) Існують американські та європейські види клена, ясена, сосни.
3. Причини подібності та відмінності фауни та флори.
а) Ізоляція ареалів.
· Якщо ізоляція відбулася нещодавно, то подібності більше, ніж відмінностей: Беренгова протока утворилася нещодавно, тому фауна Азії мало відрізняється від фауни Америки; Північна та Південна Америкаоб'єдналися нещодавно, тому їхні фауни є різними; Австралія відокремилася з інших континентів давно, тому має своєрідну флору і фауну, еволюція йшла повільно, оскільки Австралія щодо невелика; своєрідні фауна та флора островів та замкнутих водойм.
4. Сучасний географічний розподіл тварин та рослин можна пояснити лише з еволюційної точки зору.
IV. Палеонтологічні (Палеонтологія вивчає викопні організми, умови їхнього життя та поховання).
1. Зміна фауни та флори на Землі.
а) У найдавніших пластах виявлені лише безхребетні.
б) Чим молодший пласт, тим ближчі залишки до сучасних видів.
в) За допомогою палеонтологічних знахідок вдалося встановити філогенетичні ряди та перехідні форми.
2. Викопні перехідні форми- Форми організмів, що поєднують ознаки більш давніх і молодих форм.
а) Звірозубі рептилії виявлені на Північній Двіні (рід Іноземцевія). Мали подібність до ссавців у будові наступних органів: черепа; хребта; кінцівок розташованих не з боків тулуба, як у рептилій, а під тулубом, як у ссавців; зубів, диференційованих на ікла, різці та корінні.
б) Археоптерикс– перехідна форма між птахами та рептиліями, виявлена у шарах Юрського періоду(150 млн років тому).
· Ознаки птахів: задні кінцівки з цівкою, крила та пір'я, зовнішня схожість.
· Ознаки рептилій: довгий хвіст, що складається з хребців; черевні ребра; наявність зубів; пазурі на передній кінцівці.
· Погано літав з таких причин: грудина була без кіля, тобто. грудні м'язи були слабкими; хребет і ребра були жорсткою опорою, як у птахів.
в) Псилофіти– перехідна форма між водоростями та наземними рослинами.
· Відбулися від зелених водоростей.
· Від псилофітів походять вищі спорові судинні рослини - плауни, хвощі, папороті.
· З'явилися в силурі, а поширилися у девоні.
· Відмінності від водоростей та вищих спорових: псилофіти - трав'янисті та дерев'янисті рослини, що ростуть по берегах морів; мали розгалужене стебло з лусочками; шкірка мала продихи; підземне стебло нагадувало кореневища з ризоидами; стебло було диференційовано на провідні, покривні та механічні тканини.
3. Філогенетичні ряди– ряди деяких форм, які послідовно змінювали один одного в ході еволюції (філогенезу).
а) В.О. Ковалевський відновив еволюцію коня, збудувавши її філогенетичний ряд.
· Еогіппус, що жив у палеоген, був розміром з лисицю, мав чотирипалу передню кінцівку та трипалу задню. Зуби були горбкуваті (ознака всеїдності).
· У неогені клімат став більш посушливим, змінилася рослинність, еогіппус еволюціонував через ряд форм: еогіппус, мерігіппус, гіппаріон, сучасний кінь.
· Ознаки еогіппусу змінилися: ноги подовжилися; кіготь перетворився на копита; скоротилася поверхня опори, тож кількість пальців зменшилася до одного; швидкий біг призвів до зміцнення хребта; перехід на грубі корми призвів до утворення складчастих зубів.
Перехідні (проміжні) форми- організми, які поєднують у своїй будові ознаки двох великих систематичних груп.
Перехідні форми характеризуються наявністю древніших і примітивних (у сенсі первинних) характеристик, ніж пізніші форми, але, водночас, наявністю більш прогресивних (у сенсі пізніших) характеристик, ніж предки. Як правило, термін «перехідні форми» вживають по відношенню до копалин, хоча проміжні види зовсім не обов'язково повинні вмирати.
Перехідні форми використовують як один із доказів існування біологічної еволюції.
Історія поняття
У 1859 р., коли було видано роботу Ч. Дарвіна «Походження видів», кількість викопних залишків була вкрай малою, науці були відомі перехідні форми. Дарвін описав відсутність проміжних форм «як найбільш очевидне і тяжке заперечення, яке може бути проти теорії», але пояснив це крайньою неповнотою геологічного літопису. Він наголошував на обмеженій кількості доступних колекцій у той час, в той же час описав наявну інформацію про наявні копалини з точки зору еволюції та дії природного відбору. Тільки через два роки, в 1961 році. Було знайдено археоптерикс, який представляв класичну перехідну форму між плазунами та птахами. Його знахідках стала не тільки підтвердженням теорії Дарвіна, а також знаковим фактом, що підтверджує реальність існування біологічної еволюції. З того часу було знайдено велику кількість копалин, які показують, що всі класи хребетних тварин є спорідненими між собою, причому більшість із них — через перехідні форми.
Зі збільшенням відомостей про таксономічну різноманітність судинних рослин на початку ХХ ст., розпочалися дослідження з пошуку їх можливого предка. У 1917 р. Роберт Кідстон та Вільям Генрі Ленд виявили залишки дуже примітивної рослини біля селища. Rhyniaв Шотландії. Ця рослина була названа Rhynia.Вона поєднує ознаки зелених водоростей і судинних рослин.
Трактування поняття
Перехідні форми між двома групами організмів не обов'язково нащадками однієї групи та предком іншої. За копалинами, як правило, неможливо точно встановити, чи є певний організм предком іншого. Крім того, можливість знайти в палеонтологічному літописі прямого предка певної форминадзвичайно мала. Набагато більша можливість виявити щодо близьких родичів цього предка, які схожі з ним за будовою. Тому будь-яка перехідна форма автоматично інтерпретується як бічна гілка еволюції, а чи не «ділянка філогенетичного ствола».
Перехідні форми та таксономія
Еволюційна таксономія залишалася домінуючою формою таксономії протягом ХХ ст. Виділення таксонів виходить з різних ознаках, унаслідок чого таксони зображують як гілок розгалуженого еволюційного дерева. Перехідні форми розглядаються як «падаючі» між різними групами в плані анатомії, вони суміш характеристик від внутрішньої та зовнішньої поклажі, що нещодавно розділилася.
З розвитком кладистики у 1990-х pp. Взаємозв'язки зазвичай зображують у вигляді кладограми, що ілюструє дихотомічне розгалуження еволюційних ліній. Тому в кладистиці перехідні форми розглядаються як більш ранні гілки дерева, де ще не розвинулися не всі риси, характерні для відомих нащадків на цій гілці. Такі ранні представники групи зазвичай називають основним таксоном (англ. Basal taxa)чи сестринським таксоном (англ. Sister taxa),залежно від того, чи належить викопний організмдо цієї поклажі чи ні.
Проблеми виявлення та інтерпретації
Відсутність перехідних форм між багатьма групами організмів є предметом критики креаціоністів. Однак далеко не кожна перехідна форма існує у вигляді скам'янілостей через принципову неповноту палеонтологічного літопису. Неповнота викликана особливостями процесу фосілізації, тобто переходу до скам'янілого стану. Для утворення скам'янілості необхідно, щоб організм, який загинув, був похований під великим шаром осадових порід. Через дуже повільну швидкість осадконакопичення на суші, сухопутні видирідко переходять у скам'янілий стан і зберігаються. Крім того, рідко вдається виявити види, що живуть у глибинах океану через рідкісні випадки підняття на поверхню великих масивів дна. Таким чином, більшість відомих копалин (а, відповідно, і перехідних форм) - це або види, що мешкають на мілководді, в морях і річках, або наземні види, які ведуть напівводний спосіб життя, або живуть у берегової лінії. До згаданих вище проблем слід додати надзвичайно малу (у масштабах планети) кількість палеонтологів, які здійснюють розкопки.
Перехідні форми, як правило, не живуть на великих територіяхі не існують протягом великого часу, інакше вони були б персистентними. Цей факт також знижує ймовірність фосилізації та подальшого виявлення перехідних форм.
Тому можливість виявлення проміжних форм надзвичайно мала.
Приклади серед тварин
Найдавнішими представниками земноводних вважають їхтіостеги. Їх вважають перехідною ланкою між кистеперими рибами та земноводними. Незважаючи на те, що в їхтіостеги була п'ятипала кінцівка, адаптована до життя на суші, значну частину життя вони проводили як риби, мали хвостовий плавець, бічну лінію та деякі інші ознаки риб.
Батрахозаври, що існували в кам'яновугільний і пермський періоди, розглядають як перехідну форму між земноводними та плазунами. Батрахозаври, хоч і проводили життя у дорослій стадії на суші (подібно до плазунів), були тісно пов'язані з водоймами і зберегли ряд ознак, властивих земноводним, зокрема, відкладання ікри та розвиток личинок у воді, наявність зябер тощо.
Виявлено велику кількість плазунів, які виробили здатність літати, частина з них мала пір'я, тому їх розглядають як перехідні форми між плазунами та птахами. Найбільш відомий археоптерикс. Він був розміром приблизно із сучасну ворону. Формою тіла, будовою кінцівок і наявністю оперення подібний до сучасних птахів, можливо, літав. Спільним з плазунами була особлива будова таза і ребер, наявність дзьоба з конічними зубами, по три вільні пальці на крилах, подвійновигнуті хребці, довгий хвіст з 20-21 хребця, кістки могло не пневматизуватися, грудна кістка без кіля. Інші відомі перехідні форми між плазунами та птахами — протоавіс, конфуціусорніс.
Велика кількість викопних форм звіроподібних плазунів (синапсиди, терапсид, пелікозаврів, різних динозаврових та ін.), знайдених у багатьох районах земної кулі, існували в юрський та крейдяний періоди, що поєднують ознаки плазунів та ссавців, розкривають можливі напрямкита способи становлення різних групчотирилапих, зокрема ссавців. Наприклад, звіроподібний плазун із групи терапсид — ліценопс. (Lycaenops)по розвитку кісток ротової порожнини, диференціюванням зубів на ікла, різці, по різцеві зуби та рядом інших ознак будови тіла нагадує хижих ссавців, хоча за іншими ознаками та способом життя це були справжні плазуни.
Однією з форм, що збереглася у викопному стані є амбулоцетус Ambulocetus natans(«ходячий кит») - перехідна форма між наземними ссавцями та китоподібними, які є вторинноводними формами. Зовні тварина нагадувала щось середнє між крокодилом та дельфіном. Шкіра має частково редуковану вовну. Тварина мала лапи з перетинками; хвіст та кінцівки пристосовані як допоміжні органи пересування у воді.
Приклади серед рослин
Перші наземні рослини з класу рініопсид, сімей ринієвих і псилофітових, що жили в силурі-девоні, поєднували ознаки зелених водоростей та примітивних форм вищих рослин. Їхнє тіло було безлистим, циліндричний осьовий орган — тілом у верхній частині дихотомічно розгалуженим на верхівках із спорангіями. Функцію мінерального харчування рініопсид виконували ризоїди.
Викопні форми насіннєвих папоротей, які процвітали наприкінці девону, поєднують у собі ознаки папоротей та голонасінних. Вони утворювали не тільки суперечки (як папороті), а й насіння (як насіння рослини). Тканина їх стебел, що проводить, за будовою нагадує деревину голонасінних (саговників).
Інший попередник насіннєвих рослин був ідентифікований із відкладень середнього девону. Рункарія (Runcaria heinzelinii)існувала близько 20 млн. років тому. Це була невелика рослина із радіальною симетрією; мала спорангій, оточений інтегументом та плюскою. Рункарія демонструє шлях еволюції рослин від спорових до насіннєвих.
Перехідні форми в еволюції людини
У наш час знайдено велику кількість копалин, які розкривають еволюційний шлях людини розумної від її людиноподібних предків. До форм, які більшою чи меншою мірою можна віднести до перехідних, належать: сахелантропа, ардипітека, австралопітеки (африканський, афарський та інші), людина вміла, людина працююча, людина прямоходяча, людина-попередник, гейдельберзька людина і кроманьйонці.
Серед згаданих форм значної уваги заслуговують австралопітеки. Австралопитек афарський з погляду еволюції перебуває між сучасними двоногими людьми та його чотирилапими древніми предками. Велика кількість рис скелета цього австралопітеку чітко відображають двоногость, причому настільки, що деякі дослідники вважають, що ця властивість виникла задовго до появи австралопітеку афарського. Серед загальних рис анатомії його таз набагато більше схожий на цих кісток у людини, ніж у мавп. Краї клубових кісток коротші і ширші, крижова кістка широка і розташована безпосередньо позаду кульшового суглоба. Існує явне свідчення про існування місць кріплення для м'язів-розгиначів коліна, що передбачає вертикальне положення цього організму. У той час, як таз австралопітеку не зовсім як у людини (помітно ширше, з орієнтацією краю клубових кісток назовні), ці особливості вказують на важливу перебудову, пов'язану з ходінням на двох ногах. Стегна кістки утворює кут у напрямку коліна. Ця риса дозволяє нозі розміщуватися ближче до середньої лінії тіла і є свідченням звичний характер пересування на двох ногах. У наш час людина розумна, орангутани та коати мають такі самі риси. Ноги австралопітека мали великі пальціщо робить практично неможливим захоплення стопою гілок дерев. Крім особливостей локомоції, в австралопітеку був також значно більше мозок, ніж у сучасних шимпанзе і зуби були значно більше подібними до зубів сучасної людини, ніж до мавп.
Філогенетичні ряди
Філогенетичні ряди - ряди копалин форм, пов'язані між собою в процесі еволюції і відображають поступові зміни їхнього історичного розвитку.
Були досліджені російським ученим А. Ковалевським та англійським Дж. Сімпсоном. Вони показали, що сучасні однопалі копитні походять від стародавніх дрібних всеїдних тварин. Аналіз копалин коней допоміг встановити поступовість процесу еволюції в межах цієї групи тварин, зокрема, як змінюючись у часі, викопні форми набували все більшої схожості з сучасними кіньми. Порівнюючи еоценового єогіпусу з сучасним конем, важко довести їхню філогенетичну спорідненість. Проте наявність низки перехідних форм, які послідовно змінювали одне одного великих просторах Євразії та Північної Америки, дозволила відновити філогенетичний ряд коней і встановити напрямок їх еволюційних змін. Він складається з ряду наступних форм(у спрощеному вигляді): Phenacodus — Eohippus — Miohippus — Parahippus — Pliohippus — Equus.
Гільгендорф (1866) описав палеонтологічний ряд черевоногих молюсків із міоценових відкладень, що накопичилися протягом двох мільйонів років в озерних відкладах Штейнгеймського басейну (Вюртемберг, Німеччина). Було виявлено в послідовних шарах 29 різних форм, що належать до ряду планорбіс (Planorbis).Стародавні молюски мали раковину як спіраї, а пізніші — як турбоспіралі. Ряд мав два відгалуження. Передбачається, що зміна форми черепашки була викликана підвищенням температури та збільшенням вмісту карбонату кальцію внаслідок гарячих вулканічних джерел.
Таким чином, філогенетичні ряди є історичною послідовністю перехідних форм.
В даний час відомі філогенетичні ряди для амонітів (Вааген, 1869), черевоногих молюсків роду живородок (Viviparus)(Неймайром, 1875), носорогів, слонів, верблюдів, парнокопитних та інших тварин.
Подібність зародків. Біогенетичний закон
Вивчення ембріонального та постембріонального розвитку тварин дозволило знайти загальні рисиу цих процесах і сформулювати закон зародкової подібності (К. Бер) та біогенетичний закон (Ф. Мюллер та Е. Геккель), що мають величезне значення для розуміння еволюції.
Усі багатоклітинні організми розвиваються із заплідненого яйця. Процеси розвитку зародків у тварин, що належать до одного типу, багато в чому подібні. У всіх хордових тварин в ембріональному періоді закладається осьовий скелет – хорда, виникає нервова трубка. План будови хордових тварин також однаковий. На ранніх стадіях розвитку зародки хребетних надзвичайно подібні (рис. 24).
Ці факти підтверджують справедливість сформульованого К. Бером закону зародкової подібності: «Ембріони виявляють, вже починаючи з ранніх стадій, відома загальна подібність у межах типу». Подібність зародків є свідченням спільності їх походження. Надалі у будові зародків виявляються ознаки класу, роду, виду і, нарешті, ознаки, притаманні даної особини. Розбіжність ознак зародків у розвитку називається ембріональної дивергенцією і відбиває еволюцію тієї чи іншої систематичної групи тварин.
Велика схожість зародків ранніх стадіях розвитку та поява відмінностей більш пізніх стадіях має пояснення. Вивчення ембріональної мінливості показує, що мінливі стадії розвитку. Мутаційний процес зачіпає і гени, що зумовлюють особливості будови та обміну речовин у наймолодших ембріонів. Але структури, що у ранніх ембріонів (давні ознаки, властиві далеким предкам), грають дуже важливу рольу процесах подальшого розвитку. Зміни на ранніх стадіях зазвичай призводять до недорозвинення та загибелі. Навпаки, зміни на пізніх стадіях можуть бути сприятливими для організму, тому підхоплюються природним відбором.
Поява в ембріональному періоді розвитку сучасних тварин ознак, властивих далеким предкам, відбиває еволюційні перетворення на будову органів.
У своєму розвитку організм проходить одноклітинну стадію (стадія зиготи), що можна розглядати як повторення філогенетичної стадії первісної амеби. У всіх хребетних, включаючи вищих представників, закладається хорда, яка далі заміщається хребтом, а в їхніх предків, якщо судити по ланцетнику, хорда залишалася все життя.
У ході ембріонального розвитку птахів та ссавців, включаючи людину, з'являються зяброві щілини в глотці та відповідні їм перегородки. Факт закладки частин зябрового апарату у зародків наземних хребетних пояснюється їх походженням від рибоподібних предків, що дихали зябрами. Будова серця людського зародка нагадує у період будова цього органу в риб.
Подібні приклади вказують на глибокий зв'язок між індивідуальним розвитком організмів та їх історичним розвитком. Цей зв'язок знайшов свій вираз у біогенетичному законі, сформульованому Ф. Мюллером та Е. Геккелем у XIX ст.: онтогенез (індивідуальний розвиток) кожної особини є коротке та швидке повторення філогенезу (історичний розвиток) виду, до якого ця особина відноситься.
Біогенетичний закон зіграв визначну роль розвитку еволюційних ідей. Великий внесок у поглиблення уявлень про еволюційну роль ембріональних перетворень належить А. Н. Северцову. Він встановив, що у індивідуальному розвитку повторюються ознаки не дорослих предків, які зародків.
Філогенезрозглядається тепер як зміна послідовностей низки дорослих форм, бо як історичний ряд відібраних природним відбором онтогенезів. Піддаються відбору завжди цілі онтогенези, і такі, які, попри вплив несприятливих чинників середовища, виживають усім стадіях розвитку, залишаючи життєздатне потомство. Таким чином, основу філогенезу складають зміни, що відбуваються в онтогенезі окремих особин.
Палеонтологічні докази. Зіставлення викопних залишків із земних пластів різних геологічних епох переконливо свідчить про зміну органічного світу у часі. Дані палеонтології дають великий матеріалпро наступні зв'язки між різними систематичними групами. В одних випадках вдалося встановити перехідні форми, в інших – філогенетичні ряди, тобто ряди видів, які послідовно змінюють один одного.
Викопні перехідні форми:
а) археоптерикс- перехідна форма між птахами та рептиліями, виявлена в шарах Юрського періоду (150 млн років тому). Ознаки птахів: задні кінцівки з цівкою, наявність пір'я, зовнішня схожість, крила. Ознаки рептилій: довгий хвіст, що складається з хребців, черевні ребра, наявність зубів, кістки на передній кінцівці;
Б) псилофіти- перехідна форма між водоростями та наземними рослинами.
Філогенетичні лави. В. О. Ковальський відновив еволюцію коня, побудувавши її ряд філогенезу (рис. 25).

Еволюція коня охоплює досить великий проміжок часу. Найдавніший предокконя відноситься до початку третинного періоду, тоді як сучасний кінь - до четвертинного періоду. Види роду Евкус були маленькими лісовими тваринами заввишки 30 см. Вони мали по чотири пальці на ногах, що полегшувало ходіння та біг по топкому ґрунту лісових боліт. Судячи з зубів, ці тварини харчувалися м'якою рослинною їжею. Належать вони до нижнього еоцену Північної Америки. За цією формою йдуть середньоеоценові орогіппуси, у яких на передніх ногах були розвинені ще чотири пальці. У середньому еоцені з'являється епігіпус, у якого четвертий палець зменшений. В олігоцені жив нащадок попередніх форм – мезогіппус. У нього на ногах зберігаються лише три пальці, причому середній палець розвинений помітно сильніше за інших. Зростання тварин сягає 45 див.
Починають з'являтися зміни у зубній системі. Горбкові передні зуби еогіппусів, пристосовані до м'якої рослинної їжі, перетворюються на зуби з борозенками. Еволюція торкається і корінних зубів, вони стають більш пристосованими до грубої степової рослинної їжі. У верхньому олігоцені мезогіппус поступається місцем цілої низки форм: міогшгаус, а в нижньому міоцені - пара-гіппус. Парагіппус є родоначальником наступного етапу кінського ряду – мерихіппуса. Мерихіппуси були, безсумнівно, жителями відкритих просторів, причому у різних видів цього йшов процес укорочення бічних пальців: в одних видів пальці були довшими, в інших - коротшими, наближаючись до останньому випадкудо швидконогих однопалих коней.
Зрештою, у пліогіппусу, що жив у пліоцені, цей процес закінчується утворенням нової форми, стародавнього однопалого коня - плезіпуса. За формою та розмірами останній близький до сучасного, відомого з плейстоцену коня.
Виникнувши в Америці, сучасна формаконя потім проникає в Євразію в числі кількох видів. Зрештою, всі американські коні вимерли, а європейські вціліли і потім вдруге потрапили в Америку. Цього разу вони були завезені сюди європейцями. початку XVIв. Таким чином, еволюція коней переконливо показує процес еволюції, що веде до виникнення нових видів шляхом перетворення їхніх предків.
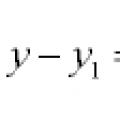 Рівняння прямих та кривих на площині
Рівняння прямих та кривих на площині Узагальнене однорідне рівняння узагальнено однорідне рівняння другого порядку
Узагальнене однорідне рівняння узагальнено однорідне рівняння другого порядку Завдяки кома Завдяки тому що в них
Завдяки кома Завдяки тому що в них