Белки получение и химические свойства. Химические свойства белков
§ 9. ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА БЕЛКОВ
Белки – это очень крупные молекулы, по своим размерам они могут уступать только отдельным представителям нуклеиновых кислот и полисахаридам. В таблице 4 представлены молекулярные характеристики некоторые белков.
Таблица 4
Молекулярные характеристики некоторых белков
|
Относитель-ная молекулярная масса |
Число цепей |
Число аминокислотных остатков |
|
|
Рибонуклеаза |
|||
|
Миоглобин |
|||
|
Химотрипсин |
|||
|
Гемоглобин |
|||
|
Глутамат-дегидрогеназа |
В молекулах белков может содержаться самое разное количество аминокислотных остатков - от 50 и до нескольких тысяч; относительные молекулярные массы белков также сильно колеблются - от нескольких тысяч (инсулин, рибонуклеаза) до миллиона (глутаматдегидрогеназа) и более. Число полипептидных цепей в составе белков может составлять от единицы до нескольких десятков и даже тысяч. Так, в состав белка вируса табачной мозаики входит 2120 протомеров.
Зная относительную молекулярную массу белка, можно приблизительно оценить, какое число аминокислотных остатков входит в его состав. Средняя относительная молекулярная масса аминокислот, образующих полипептидную цепь, равна 128. При образовании пептидной связи происходит отщепление молекулы воды, следовательно, средняя относительная масса аминокислотного остатка составит 128 – 18 = 110. Используя эти данные, можно подсчитать, что белок с относительной молекулярной массой 100000 будет состоять приблизительно из 909 аминокислотных остатков.
Электрические свойства белковых молекул
Электрические свойства белков определяются присутствием на их поверхности положительно и отрицательно заряженных аминокислотных остатков. Наличие заряженных группировок белка определяет суммарный заряд белковой молекулы. Если в белках преобладают отрицательно заряженные аминокислоты, то его молекула в нейтральном растворе будет иметь отрицательный заряд, если преобладают положительно заряженные – молекула будет иметь положительный заряд. Суммарный заряд белковой молекулы зависит и от кислотности (рН) среды. При увеличении концентрации ионов водорода (увеличении кислотности) происходит подавление диссоциации карбоксильных групп:
и в то же время увеличивается число протонированных амино-групп;
Таким образом, при увеличении кислотности среды происходит уменьшение на поверхности молекулы белка числа отрицательно заряженных и увеличение числа положительно заряженных групп. Совсем другая картина наблюдается при снижении концентрации ионов водорода и увеличении концентрации гидроксид-ионов. Число диссоциированных карбоксильных групп возрастает
и снижается число протонированных аминогрупп
Итак, изменяя кислотность среды, можно изменить и заряд молекулы белка. При увеличении кислотности среды в молекуле белка снижается число отрицательно заряженных группировок и увеличивается число положительно заряженных, молекула постепенно теряет отрицательный и приобретает положительный заряд. При снижении кислотности раствора наблюдается противоположная картина. Очевидно, что при определенных значениях рН молекула будет электронейтральной, т.е. число положительно заряженных групп будет равно числу отрицательно заряженных групп, и суммарный заряд молекулы будет равен нулю (рис. 14).
Значение рН, при котором суммарный заряд белка равен нулю, называется изоэлектрической точкой и обозначается pI .

Рис. 14. В состоянии изоэлектрической точки суммарный заряд молекулы белка равен нулю
Изоэлектрическая точка для большинства белков находится в области рН от 4,5 до 6,5. Однако есть и исключения. Ниже приведены изоэлектрические точки некоторых белков:
При значениях рН ниже изоэлектрической точки белок несет суммарный положительный заряд, выше – суммарный отрицательный.
В изоэлектрической точке растворимость белка минимальна, так как его молекулы в таком состоянии электронейтральны и между ними нет сил взаимного отталкивания, поэтому они могут «слипаться» за счет водородных и ионных связей, гидрофобных взаимодействий, ван-дер-ваальсовых сил. При значениях рН, отличающихся от рI, молекулы белка будут нести одинаковый заряд - либо положительный, либо отрицательный. В результате этого между молекулами будут существовать силы электростатического отталкивания, препятствующие их «слипанию», растворимость будет выше.
Растворимость белков
Белки бывают растворимые и нерастворимые в воде. Растворимость белков зависит от их структуры, величины рН, солевого состава раствора, температуры и других факторов и определяется природой тех групп, которые находятся на поверхности белковой молекулы. К нерастворимым белкам относятся кератин (волосы, ногти, перья), коллаген (сухожилия), фиброин (щелк, паутина). Многие другие белки растворимы в воде. Растворимость определяется наличием на их поверхности заряженных и полярных группировок (-СОО - , -NH 3 + , -OH и др.). Заряженные и полярные группировки белков притягивают к себе молекулы воды, и вокруг них формируется гидратная оболочка (рис. 15), существование которой обусловливает их растворимость в воде.

Рис. 15. Образование гидратной оболочки вокруг молекулы белка.
На растворимость белка влияет наличие нейтральных солей (Na 2 SO 4 , (NH 4) 2 SO 4 и др.) в растворе. При малых концентрациях солей растворимость белка увеличивается (рис. 16), так как в таких условиях увеличивается степень диссоциации полярных групп и экранируются заряженные группы белковых молекул, тем самым снижается белок-белковое взаимодействие, способствующее образованию агрегатов и выпадению белка в осадок. При высоких концентрациях солей растворимость белка снижается (рис. 16) вследствие разрушения гидратной оболочки, приводящего к агрегации молекул белка.

Рис. 16. Зависимость растворимости белка от концентрации соли
Существуют белки, которые растворяются только в растворах солей и не растворяются в чистой воде, такие белки называют глобулины
. Существуют и другие белки – альбумины
, они в отличие от глобулинов хорошо растворимы в чистой воде.
Растворимость белков зависит и от рН растворов. Как мы уже отмечали, минимальной растворимостью обладают белки в изоэлектрической точке, что объясняется отсутствием электростатического отталкивания между молекулами белка.
При определенных условиях белки могут образовывать гели. При образовании геля молекулы белка формируют густую сеть, внутреннее пространство которой заполнено растворителем. Гели образуют, например, желатина (этот белок используют для приготовления желе) и белки молока при приготовлении простокваши.
На растворимость белка оказывает влияние и температура. При действии высокой температуры многие белки выпадают в осадок вследствие нарушения их структуры, но об этом более подробно поговорим в следующем разделе.
Денатурация белка
Рассмотрим хорошо нам знакомое явление. При нагревании яичного белка происходит постепенное его помутнение, и затем образуется твердый сгусток. Свернувшийся яичный белок – яичный альбумин – после охлаждения оказывается нерастворимым, в то время как до нагревания яичный белок хорошо растворялся в воде. Такие же явления происходят и при нагревании практически всех глобулярных белков. Те изменения, которые произошли при нагревании, называются денатурацией
. Белки в естественном состоянии носят название нативных
белков, а после денатурации - денатурированных
.
При денатурации происходит нарушение нативной кон-формации белков в результате разрыва слабых связей (ион-ных, водородных, гидрофобных взаимодействий). В результате этого процесса могут разрушаться четвертичная, третичная и вторичные структуры белка. Первичная структура при этом сохраняется (рис. 17).

Рис. 17. Денатурация белка
При денатурации гидрофобные радикалы аминокислот, находящиеся в нативных белках в глубине молекулы, оказываются на поверхности, в результате создаются условия для агрегации. Агрегаты белковых молекул выпадают в осадок. Денатурация сопровождается потерей биологической функции белка.
Денатурация белка может быть вызвана не только повышенной температурой, но и другими факторами. Кислоты и щелочи способны вызвать денатурацию белка: в результате их действия происходит перезарядка ионогенных групп, что приводит к разрыву ионных и водородных связей. Мочевина разрушает водородные связи, следствием этого является потеря белками своей нативной структуры. Денатурирующими агентами являются органические растворители и ионы тяжелых металлов: органические растворители разрушают гидрофобные связи, а ионы тяжелых металлов образуют нерастворимые комплексы с белками.
Наряду с денатурацией существует и обратный процесс – ренатурация. При снятии денатурирующего фактора возможно восстановление исходной нативной структуры. Например, при медленном охлаждении до комнатной температуры раствора восстанавливается нативная структура и биологическая функция трипсина.
Белки могут денатурировать и в клетке при протекании нормальных процессов жизнедеятельности. Совершенно очевидно, что утрата нативной структуры и функции белков – крайне нежелательное событие. В связи с этим следует упомянуть об особых белках – шаперонах . Эти белки способны узнавать частично денатурированные белки и, связываясь с ними, восстанавливать их нативную конформацию. Шапероны также узнают белки, процесс денатурации которых зашел далеко, и транспортируют их в лизосомы, где происходит их расщепление (деградация). Шапероны играют важную роль и в процессе формирования третичной и четвертичной структур во время синтеза белка.
Интересно знать! В настоящее время часто упоминается такое заболевание, как коровье бешенство. Эту болезнь вызывают прионы. Они могут вызывать у животных и человека и другие заболевания, носящие нейродегенеративный характер. Прионы – это инфекционные агенты белковой природы. Прион, попадая в клетку, вызывает изменение конформации своего клеточного аналога, который сам становится прионом. Так возникает заболевание. Прионный белок отличается от клеточного по вторичной структуре. Прионная форма белка имеет в основном b -складчатую структуру, а клеточная – a -спиральную.
Донецкая общеобразовательная школа I – III ступеней № 21
«Белки. Получение белков реакцией поликонденсации аминокислот. Первичная, вторичная и третичная структуры белков. Химические свойства белков: горение, денатурация, гидролиз и цветные реакции. Биохимические функции белков».
Подготовила
учитель химии
учитель – методист
г. Донецк, 2016
«Жизнь – это способ существования белковых тел»
Тема урока. Белки. Получение белков реакцией поликонденсации аминокислот. Первичная, вторичная и третичная структуры белков. Химические свойства белков: горение, денатурация, гидролиз и цветные реакции. Биохимические функции белков.
Цели урока. Ознакомить учащихся с белками как найвысшей степенью развития веществ в природе, которые обусловили появление жизни; показать их строение, свойства и разнообразие биологических функций; расширить понятие о реакции поликонденсации на примере получения белков, информировать школьников о гигиене питания, о сохранении своего здоровья. Развивать у учащихся логическое мышление.
Реактивы и оборудование. Таблица « Первичная, вторичная и третичная структуры белков». Реактивы: HNO3, NaOH, CuSO4, куриный белок, шерстяная нить, химическая посуда.
Метод урока. Информационно – развивающий.
Тип урока. Урок усвоения новых знаний и умений.
Ход урока
І. Организационный момент.
ІІ. Проверка домашнего задания, актуализация и коррекция опорных знаний.
Блицопрос
1. Объясните термин «аминокислота».
2. Назовите функциональные группы, которые входят в состав аминокислот.
3. Номенклатура аминокислот и их изомерия.
4. Почему аминокислоты проявляют амфотерные свойства? Напишите уравнения химических реакций.
5. Благодаря каким свойствам аминокислоты образуют полипептиды. Напишите реакцию поликонденсации аминокислот.
ІІІ. Сообщение темы, цели урока, мотивация учебной деятельности .
IV. Восприятие и первичное осознание нового материала.
Учитель.
«Повсюду, где мы встречаем жизнь, мы находим, что она связана с каким – либо белковым телом» - так написал Ф. Энгельс в своей книге «Анти – Дюринг». Недостаток белка в пище приводит к общему ослаблению организма, у детей – к замедлению умственного и физического развития. На сегодня больше половины человечества не получает с пищей необходимого количества белков. В сутки человеку необходимо 115 г белка, про запас белок не откладывается в отличие от углеводов и жиров, поэтому необходимо следить за своим рационом. Мы знакомы с вами с кератином – белком из которого состоят волосы, ногти, перья, кожа, - он выполняет строительную функцию; знакомы с белком пепсином – он содержится в желудочном соке и способен разрушать другие белки при пищеварении; белок тромбин участвует в свертывании крови; гормон поджелудочной железы - инсулин – регулирует обмен глюкозы; гемоглобин транспортирует О2 ко всем клеткам и тканям организма и т. д.
Откуда же берется это бесконечное многообразие белковых молекул, многообразие их функций и их особая роль в жизненных процессах? Для того, чтобы ответить на этот вопрос обратимся к составу и строению белков.
В состав белков входят атомы?...
Чтобы ответить на этот вопрос проведем разминку. Отгадайте загадки и объясните смысл ответов.
1. Он повсюду и везде:
В камне, в воздухе, в воде.
Он и в утренней росе
И в небес голубизне.
(кислород)
2. Я – самый легкий элемент,
В природе без меня ни шагу.
И с кислородом я в момент
3. В воздухе он главный газ,
Окружает всюду нас.
Угасает жизнь растений
Без него, без удобрений.
В наших клеточках живет
4. Отправились школьники как – то в поход
(К задаче химической это подход).
Ночью костер развели при луне,
Песенки пели о ярком огне.
Отбросьте в сторонку свои сантименты:
Какие горели в огне элементы?
(углерод, водород)
Да, правильно, это главные химические элементы, входящие в состав белка.
Об этих четырех элементах можно сказать словами Шиллера « Четыре элемента, сливаясь вместе, дают жизнь и строят мир».
Белки – это природные полимеры, состоящие из остатков α – аминокислот, соединенных между собой пептидными связями.
В состав белков входит 20 различных аминокислот, отсюда следует огромное многообразие белков при их различных комбинациях. В организме человека насчитывается до 100 000 белков.
Историческая справка.
Первая гипотеза о строении молекулы белка была предложена в 70-х гг. XIX в. Это была уреидная теория строения белка.
В 1903г. немецким ученым была высказана пептидная теория, давшая ключ к тайне строения белка. Фишер предположил, что белки представляют собой полимеры аминокислот, соединенных пептидной связью.
Идея о том, что белки – это полимерные образования, высказывалась еще в 70 – 88 гг. XIX в. , русским ученым. Эта теория получила подтверждение в современных работах.
Уже первое ознакомление с белками дает некоторое представление о чрезвычайно сложном строении их молекул. Получают белки реакцией поликонденсации аминокислот:
https://pandia.ru/text/80/390/images/image007_47.gif" width="16" height="18">H – N – CH2 – C + H – N – CH2 – C →
https://pandia.ru/text/80/390/images/image012_41.gif" height="20 ">
NH2 - CH – C – N – CH – C – N – CH – C - … + nH2O →
⸗ O ⸗ O ⸗ O
→ NH2 – CH – C + NH2 – CH – C + NH2 – CH – C + …
̀ OH ̀ OH ̀ OH
4. Учитель демонстрирует опыт: горение шерстяной нити; ощущается запах паленых перьев – так можно отличить шерсть от тканей других видов.
V. Обобщение и систематизация знаний.
1. Составьте опорный конспект по белкам.
основа жизни ← Белки → полипептиды
(C, H, O, N) ↓ ↓ ↓ \ структуры белка
химичес цветные функции
кие св-ва реакции белка
2. Напишите уравнения реакции образования дипептида из глицина и валина.
VI. Подведение итога урока, домашнее задание.
Выучить §38 с. 178 – 184. Выполнить тестовые задания с. 183.
№1. Белки: пептидная связь, их обнаружение.
Белки – макромолекулы линейных полиамидов, образованных а-аминокислотами в результате реакции поликонденсации в биологических объектах.
Белки – это высокомолекулярные соединения, построенные из аминокислот . В создание белков участвует 20 аминокислот. Они связываются между собой в длинные цепи, которые образуют основу белковой молекулы большой молекулярной массы.
Функции белков в организме
Сочетание своеобразных химических и физических свойств белков обеспечивает именно этому классу органических соединений центральную роль в явлениях жизни.
Белки имеют следующие биологические свойства, или осуществляют следующие основные функции в живых организмах:
1. Каталитическая функция белков. Все биологические катализаторы - ферменты являются белками. В настоящее время охарактеризовано тысячи ферментов, многие из них выделены в кристаллической форме. Почти все ферменты - мощные катализаторы, повышающие скорости реакций, по крайней мере, в миллион раз. Эта функция белков является уникальной, не свойственной другим полимерным молекулам.
2. Питательная (резервная функция белков). Это, прежде всего белки, предназначенные для питания развивающегося зародыша: казеин молока, овальбумин яиц, запасные белки семян растений. Ряд других белков, несомненно, используется в организме в качестве источника аминокислот, которые, в свою очередь, являются предшественниками биологически активных веществ, регулирующих процесс обмена веществ.
3. Транспортная функция белков. Транспорт многих небольших молекул и ионов осуществляется специфическими белками. Например, дыхательная функция крови, а именно перенос кислорода, выполняется молекулами гемоглобина - белка эритроцитов. В транспорте липидов принимают участие альбумины сыворотки крови. Ряд других сывороточных белков образует комплексы с жирами, медью, железом, тироксином, витамином А и другими соединениями, обеспечивая их доставку в соответствующие органы.
4. Защитная функция белков. Основную функцию защиты выполняет иммуннологическая система, которая обеспечивает синтез специфических защитных белков - антител - в ответ на поступление в организм бактерий, токсинов или вирусов (антигенов). Антитела связывают антигены, взаимодействуя с ними, и тем самым нейтрализуют их биологическое действие и сохраняют нормальное состояние организма. Свертывание белка плазмы крови - фибриногена - и образование сгустка крови, предохраняющего от потери крови при ранениях - еще один пример защитной функции белков.
5. Сократительная функция белков. В акте мышечного сокращения и расслабления участвует множество белков. Главную роль в этих процессах играют актин и миозин - специфические белки мышечной ткани. Сократительная функция присуща также и белкам субклеточных структур, что обеспечивает тончайшие процессы жизнедеятельности клеток,
6. Структурная функция белков. Белки с такой функцией занимают первое место среди других белков тела человека. Широко распространены такие структурные белки, как коллаген в соединительной ткани; кератин в волосах, ногтях, коже; эластин - в сосудистых стенках и др.
7. Гормональная (регуляторная) функция белков. Обмен веществ в организме регулируется разнообразными механизмами. В этой регуляцииважное место занимают гормоны, вырабатываемые железами внутреннейсекреции. Ряд гормонов представлен белками, или полипептидами, например гормоны гипофиза, поджелудочной железы и др.
Пептидная связь
Формально образование белковой макромолекулы можно представить как реакцию поликонденсации α-аминокислот.
С химической точки зрения белки - это высокомолекулярные азотсодержащие органические соединения (полиамиды), молекулы которых построены из остатков аминокислот. Мономерами белков служат α-аминокислоты, общим признаком которых является наличие карбоксильной группы -СООН и аминогруппы -NH 2 у второго углеродного атома (α-углеродный атом):
Исходя из результатов изучения продуктов гидролиза белков и выдвинутых А.Я. Данилевским идей о роли пептидных связей -CO-NH- в построении белковой молекулы, немецкий ученый Э.Фишер предложил в начале XX века пептидную теорию строения белков. Согласно этой теории, белки представляют собой линейные полимеры α-аминокислот, связанных пептидной связью - полипептиды:

В каждом пептиде один концевой аминокислотный остаток имеет свободную α-аминогруппу (N-конец), а другой - свободную α-карбоксильную группу (С-конец). Структуру пептидов принято изображать, начиная с N-концевой аминокислоты. При этом аминокислотные остатки обозначаются символами. Например: Ala-Tyr-Leu-Ser-Tyr- - Cys. Этой записью обозначен пептид, в котором N-концевой α-аминокислотой яв ляется аланин, а С-концевой - цистеин. При чтении такой записи окончания названий всех кислот, кроме последних меняются на - "ил": аланил-тирозил-лейцил-серил-тирозил- -цистеин. Длина пептидной цепи в пептидах и белках, встречающихся в организме, колеблется от двух до сотен и тысяч аминокислотных остатков.
№2. Классификация простых белков.
К простым (протеинам) относят белки, дающие при гидролизе только аминокислоты.
Протеиноиды ____простые белки животного происхождения, нерастворимые вводе, растворах солей, разбавленных кислотах и щелочах. Выполняют главным образом опорные функции (например, Коллаген, кератин
протамины – положительно заряженные ядерные белки, с молекулярной массой 10-12 kDa. Примерно на 80% состоят из щелочных аминокислот, что дает им возможность взаимодействовать с нуклеиновыми кислотами посредством ионных связей. Принимают участие в регуляции генной активности. Хорошо растворимы в воде;
гистоны – ядерные белки, играющие важную роль в регуляции генной активности. Они найдены во всех эукариотических клетках, и разделены на 5 классов, различающихся по молекулярной массе и аминокислотному. Молекулярная масса гистонов находится в интервале от 11 до 22 kDa, а различия в аминокислотном составе касаются лизина и аргинина, содержание которых варьирует от 11 до 29% и от 2 до 14% соответственно;
проламины – не растворимы в воде, но растворимы в 70% спирте, особенности хим.строения – много пролина, глутаминовой кислоты нет лизина,
глутелины – растворимы в щелочных растворах,
глобулины – белки, не растворимые в воде и в полунасыщенном растворе сернокислого аммония, но растворимые в водных растворах солей, щелочей и кислот. Молекулярная масса – 90-100 kDa;
альбумины – белки животных и растительных тканей, растворим в воде и солевых растворах. Молекулярнаяя масса равна 69 kDa;
склеропротеины – белки опорных тканей животных
В качестве примеров простых белков могут служить фиброин шелка, яичный сывороточный альбумин, пепсин и др.
№3. Способы выделения и осаждения (очистки) белков.





№4. Белки как полиэлектролиты. Изоэлектрическая точка белка.
Белки являются амфотерными полиэлектролитами, т.е. проявляют как кислотные, так и основные свойства. Это обусловлено наличием в молекулах белков аминокислотных радикалов, способных к ионизации, а также свободных α-амино- и α-карбоксильных групп на концах пептидных цепей. Кислотные свойства белку придают кислые аминокислоты (аспарагиновая, глутаминовая), а щелочные свойства - основные аминокислоты (лизин, аргинин, гистидин).
Заряд белковой молекулы зависит от ионизации кислых и основных групп аминокислотных радикалов. В зависимости от соотношения отрицательных и положительных групп молекула белка в целом приобретает суммарный положительный или отрицательный заряд. При подкислении раствора белка степень ионизации анионных групп снижается, а катионных повышается; при подщелачивании - наоборот. При определенном значении рН число положительно и отрицательно заряженных групп становится одинаковым, возникает изоэлектрическое состояние белка (суммарный заряд равен 0). Значение рН, при котором белок находится в изоэлектрическом состоянии, называют изоэлектрической точкой и обозначают pI, аналогично аминокислотам. Для большинства белков pI лежит в пределах 5,5-7,0, что свидетельствует о некотором преобладании в белках кислых аминокислот. Однако есть и щелочные белки, например, сальмин - основной белок из молок семги (pl=12). Кроме того, есть белки, у которых pI имеет очень низкое значение, например, пепсин - фермент желудочного сока (pl=l). В изоэлектрической точке белки очень неустойчивые и легко выпадают в осадок, обладая наименьшей растворимостью.
Если белок не находится в изоэлектрическом состоянии, то в электрическом поле его молекулы будут перемещаться к катоду или аноду, в зависимости от знака суммарного заряда и со скоростью, пропорциональной его величине; в этом заключается сущность метода электрофореза. Этим методом можно разделять белки с различным значением pI.
Белки хотя и обладают свойствами буфера, но емкость их при физиологических значениях рН ограничена. Исключение составляют белки, содержащие много гистидина, так как только радикал гистидина обладает буферными свойствами в интервале рН 6-8. Таких белков очень мало. Например, гемоглобин, содержащий почти 8% гистидина, является мощным внутриклеточным буфером в эритроцитах, поддерживая рН крови на постоянном уровне.
№5. Физико-химические свойства белков.
Белки имеют различные химические, физические и биологические свойства, которые определяются аминокислотным составом и пространственной организацией каждого белка. Химические реакции белков очень разнообразны, они обусловлены наличием NH 2 -, СООН-групп и радикалов различной природы. Это реакции нитрования, ацилирования, алкилирования, этерификации, окисления-восстановления и другие. Белки обладают кислотно-основными, буферными, коллоидными и осмотическими свойствами.
Кислотно-основные свойства белков
Химические свойства. При слабом нагревании водных растворов белков происходит денатурация. При этом образуется осадок.
При нагревании белков с кислотами происходит гидролиз, при этом образуется смесь аминокислот.
Физико-химические свойства белков
Белки имеют высокий молекулярный вес.
Заряд белковой молекулы. Все белки имеют хоть одну свободную -NH и - СООН группы.
Белковые растворы - коллоидные растворы с разными свойствами. Белки бывают кислыми и основными. Кислые белки содержат много глу и асп, у которых есть дополнительные карбоксильные и меньше аминогрупп. В щелочных белках много лиз и арг. Каждая молекула белка в водном растворе окружена гидратной оболочкой, так как у белков за счет аминокислот есть много гидрофильных группировок (-СООН, -ОН, -NH 2 , -SH). В водных растворах белковая молекула имеет заряд. Заряд белка в воде может меняться в зависимости от РН.
Осаждение белков. У белков есть гидратная оболочка, заряд, препятствующий склеиванию. Для осаждения необходимо снять гидратную оболочку и заряд.
1.Гидратация. Процесс гидратации означает связывание белками воды, при этом они проявляют гидрофильные свойства: набухают, их масса и объем увеличивается. Набухание белка сопровождается его частичным растворением. Гидрофильность отдельных белков зависит от их строения. Имеющиеся в составе и расположенные на поверхности белковой макромолекулы гидрофильные амидные (–CO–NH–, пептидная связь), аминные (NH2) и карбоксильные (COOH) группы притягивают к себе молекулы воды, строго ориентируя их на поверхность молекулы. Окружая белковые глобулы гидратная (водная) оболочка препятствует устойчивости растворов белка. В изоэлектрической точке белки обладают наименьшей способностью связывать воду, происходит разрушение гидратной оболочки вокруг белковых молекул, поэтому они соединяются, образуя крупные агрегаты. Агрегация белковых молекул происходит и при их обезвоживании с помощью некоторых органических растворителей, например этило- вого спирта. Это приводит к выпадению белков в осадок. При изменении pH среды макромолекула белка становится заряженной, и его гидратационная способность меняется.
Реакции осаждения делят на два вида.
Высаливание белков: (NH 4)SO 4 - снимается только гидратная оболочка, белок сохраняет все виды своей структуры, все связи, сохраняет нативные свойства. Такие белки можно затем вновь растворить и использовать.
Осаждения с потерей нативных свойств белка - процесс необратимый. С белка снимается гидратная оболочка и заряд, нарушаются различные свойства в белке. Например соли меди, ртути, мышьяка, железа, концентрированные неорганические кислоты - HNO 3 , H 2 SO 4 , HCl, органические кислоты, алкалоиды - танины, йодистая ртуть. Добавление органических растворителей понижает степень гидратации и приводит к осаждению белка. В качестве таких растворителей используют ацетон. Осаждают белки также с помощью солей, например, сульфата аммония. Принцип этого метода основан на том, что при повышении концентрации соли в растворе происходит сжатие ионных атмосфер, образуемых противоионами белка, что способствует сближению их до критического расстояния, на котором межмолекулярные силы ван-дер-ваальсова притяжения перевешивают кулоновские силы отталкивания противоионов. Это приводит к слипанию белковых частиц и их выпадению в осадок.
При кипячении молекулы белков начинают хаотично двигаться, сталкиваются, снимается заряд, уменьшается гидратная оболочка.
Для обнаружения белков в растворе применяются:
цветные реакции;
реакции осаждения.
Методы выделения и очистки белков.
высаливание;
электрофорез;
хроматография: адсорбция, расщепление;
ультрацентрифугирование.
гомогенизация - клетки растираются до однородной массы;
экстракция белков водными или водно-солевыми растворами;
Структурная организация белков.
Первичная структура - определяется последовательностью аминокислот в пептидной цепочке, стабилизируется ковалентными пептидными связями (инсулин, пепсин, химотрипсин).
Вторичная структура - пространственная структура белка. Это либо -спираль, либо -складчатость. Создаются водородные связи.
Третичная структура - глобулярные и фибриллярные белки. Стабилизируют водородные связи, электростатические силы (СОО-, NН3+), гидрофобные силы, сульфидные мостики, определяются первичной структурой. Глобулярные белки - все ферменты, гемоглобин, миоглобин. Фибриллярные белки - коллаген, миозин, актин.
Четвертичная структура - имеется только у некоторых белков. Такие белки построены из нескольких пептидов. Каждый пептид имеет свою первичную, вторичную, третичную структуру, называются протомерами. Несколько протомеров соединяются вместе в одну молекулу. Один протомер не функционирует как белок, а только в соединении с другими протомерами.
Пример: гемоглобин = -глобула + -глобула - переносит О 2 в совокупности, а не по раздельности.
Белок может ренатурировать. Для этого необходимо очень короткое воздействие агентов.

6) Способы обнаружения белков.
Белки – высокомолекулярные биологические полимеры, структурными (мономерными) звеньями которых служат -аминокислоты. Аминокислоты в белках соединены друг с другом пептидной связью,образование которой происходит за счет карбоксильной группы, стоящей у -углеродного атома одной аминокислоты и -аминной группы другой аминокислоты с выделением молекулы воды. Мономерные звенья белков называют остатками аминокислот.
Пептиды, полипептиды и белки отличаются не только количеством, составом но и последовательностью аминокислотных остатков, физико-химическими свойствами и функциями, выполняемыми в организме. Молекулярная масса белков варьирует от 6 тыс. до 1 млн. и более. Химические и физические свойства белков обусловлены химической природой и физико-химическими свойствами радикалов, входящих в них остатков аминокислот. Способы обнаружения и количественного определения белков в биологических объектах и продуктах питания, а также выделения их из тканей и биологических жидкостей основаны на физических и химических свойствах этих соединений.
Белки при взаимодействии с некоторыми химическими веществами дают окрашенные соединения . Образование этих соединений происходит при участии радикалов аминокислот, их специфических групп или пептидных связей. Цветные реакции позволяют установитьналичие белка в биологическом объекте или растворе и доказать присутствиеопределенных аминокислот в белковой молекуле . На основе цветных реакций разработаны некоторые методы количественного определения белков и аминокислот.
Универсальными считают биуретовую и нингидриновую реакции , так как их дают все белки.Ксантопротеиновая реакция, реакция Фоля и др. являются специфическими, так как они обусловлены радикальными группами определенных аминокислот в молекуле белка.
Цветные реакции позволяют установить наличие белка в исследуемом материале и присутствие определенных аминокислот в его молекулах.
Биуретовая реакция . Реакция обусловлена наличием в белках, пептидах, полипептидахпептидных связей , которые в щелочной среде образуют сионами меди (II) комплексные соединения, окрашенные вфиолетовый (с красным или с синим оттенком) цвет . Окраска обусловлена наличием в молекуле не менее двух групп-CO-NH- , связанных непосредственно между собой или при участии атома углерода или азота.
Ионы меди (II) соединяются двумя ионными связями с группами =С─О ˉ и четырьмя координационными связями с атомами азота (=N―).
Итенсивность окраски зависит от количества белка в растворе. Это позволяет использовать данную реакцию для количественного определения белка. Цвет окрашенных растворов зависит от длины полипептидной цепи. Белки дают сине-фиолетовое окрашивание; продукты их гидролиза (поли- и олигопептиды) – красную или розовую окраску. Биуретовую реакцию дают не только белки, пептиды и полипептиды но и биурет (NH 2 -CO-NH-CO-NH 2) , оксамид (NH 2 -CO-CO-NH 2), гистидин.
Образующееся в щелочной среде комплексное соединение меди (II) с пептидными группами имеет следующее строение:


Нингидриновая реакция . В этой реакции растворы белка, полипептидов, пептидов и свободных α-аминокислот при нагревании с нингидрином дают синее, сине-фиолетовое или розово-фиолетовое окрашивание. Окраска в этой реакции развивается за счет α-аминогруппы.


Очень легко реагируют с нингидрином -аминокислоты. Наряду с ними сине-фиолетовый Руэмана образуют также белки, пептиды, первичные амины, аммиак и некоторые другие соединения. Вторичные амины, например пролин и оксипролин, дают желтую окраску.
Нингидриновую реакцию широко используют для обнаружения и количественного определения аминокислот.
Ксантопротеиновая реакция. Эта реакция указывает на наличие в белках остатков ароматических аминокислот – тирозина, фенилаланина, триптофана. Основана на нитровании бензольного кольца радикалов этих аминокислот с образованием нитросоединений, окрашенных в желтый цвет (греческое «Ксантос» – желтый). На примере тирозина эту реакцию можно описать в виде следующих уравнений.
В щелочной среде нитропроизводные аминокислот образуют соли хиноидной структуры, окрашенные в оранжевый цвет. Ксантопротеиновую реакцию дают бензол и его гомологи, фенол и другие ароматические соединения.

Реакции на аминокислоты, содержащие тиоловую группу в восстановленном или окисленном состоянии (цистеин, цистин).
Реакция Фоля. При кипячении со щелочью от цистеина легко отщепляется сера в виде сероводорода, который в щелочной среде образует сульфид натрия:
В связи с этим реакции определения тиолсодержащих аминокислот в растворе подразделяют на два этапа:
Переход серы из органического состояния в неорганическое
Обнаружение серы в растворе
Для выявления сульфида натрия используют ацетат свинца, который при взаимодействии с гидроксидом натрия превращается в его плюмбит:
Pb(CH 3 COO) 2 + 2NaOH Pb(ONa) 2 + 2CH 3 COOH
В результате взаимодействия ионов серы и свинца образуется сульфид свинца черного или бурого цвета:
Na 2 S + Pb (ONa ) 2 + 2 H 2 O PbS (черный осадок) + 4 NaOH
Для определения серусодержащих аминокислот к исследуемому раствору добавляют равный объем гидроксида натрия и несколько капель раствора ацетата свинца. При интенсивном кипячении в течение 3-5 минут жидкость окрашивается в черный цвет.
Наличие цистина может быть определено с помощью этой реакции, так как цистин легко восстанавливается в цистеин.
Реакция Миллона:
Это реакция на аминокислоту тирозин.
Свободные фенольные гидроксилы молекул тирозина при взаимодействии с солями дают соединения ртутной соли нитропроизводного тирозина, окрашенной в розовато-красный цвет:

Реакция Паули на гистидин и тирозин . Реакция Паули позволяет обнаружить в белке аминокислоты гистидин и тирозин, которые образуют с диазобензолсульфоновой кислотой комплексные соединения вишнево-красного цвета. Диазобензолсульфоновая кислота образуется в реакции диазотирования при взаимодействии сульфаниловой кислоты с нитритом натрия в кислой среде:

К исследуемому раствору прибавляют равный объем кислого раствора сульфаниловой кислоты (приготовленного с использованием соляной кислоты) и двойной объем раствора нитрита натрия, тщательно перемешивают и сразу прибавляют соду (карбонат натрия). После перемешивания смесь окрашивается в вишнево-красный цвет при условии наличия гистидина или тирозина в исследуемом растворе.


Реакция Адамкевича-Гопкинса-Коля (Шульца - Распайля) на триптофан (реакция на индоловую группу). Триптофан реагирует в кислой среде с альдегидами, образуя окрашенные продукты конденсации. Реакция протекает за счет взаимодействия индольного кольца триптофана с альдегидом. Известно, что из глиоксиловой кислоты в присутствии серной кислоты образуется формальдегид:
Р астворы,
содержащие триптофан, в присутствии
глиоксиловой и серной кислот дают
красно-фиолетовое окрашивание.
астворы,
содержащие триптофан, в присутствии
глиоксиловой и серной кислот дают
красно-фиолетовое окрашивание.
Глиоксиловая кислота всегда присутствует в небольшом количестве в ледяной уксусной кислоте. Поэтому реакцию можно проводить, используя уксусную кислоту. При этом к исследуемому раствору добавляют равный объем ледяной (концентрированной) уксусной кислоты и осторожно нагревают до растворения осадка.После охлаждения к смеси осторожно по стенке (во избежание смешивания жидкостей) добавляют объем концентрированной серной кислоты, равный добавленному объему глиоксиловой кислоты. Через 5-10 минут на границе раздела двух слоев наблюдают образование красно-фиолетового кольца. Если перемешать слои, содержимое посуды равномерно окрасится в фиолетовый цвет.
К
 онденсация
триптофана с формальдегидом:
онденсация
триптофана с формальдегидом:
Продукт конденсации окисляется до бис-2-триптофанилкарбинола, который в присутствии минеральных кислот образует соли, окрашенные в сине-фиолетовый цвет:


7) Классификация белков. Способы исследования аминокислотного состава.
Строгой номенклатуры и классификации белков до сих пор не существует. Названия белков дают по случайным признакам, чаще всего принимая во внимание источник выделения белка или же учитывая растворимость его в тех или иных растворителях, форму молекулы и др.
Классификация белков проводится по составу, по форме частиц, по растворимости, по аминокислотному составу, по происхождению и т.д.
1. По составу белки делят на две большие группы: простые и сложные белки.
К простым (протеинам) относят белки, дающие при гидролизе только аминокислоты (протеиноиды, протамины, гистоны, проламины, глутелины, глобулины, альбумины). В качестве примеров простых белков могут служить фиброин шелка, яичный сывороточный альбумин, пепсин и др.
К сложным (к протеидам) относят белки, составленные из простого белка и добавочной (простетической) группы небелковой природы. Группу сложных белков делят на несколько подгрупп в зависимости от характера небелкового компонента:
Металлопротеиды, содержащие в своем составе металлы (Fe, Си, Mg и др.), связанные непосредственно с полипептидной цепью;
Фосфопротеиды - содержат остатки фосфорной кислоты, которые сложноэфирными связями присоединены к молекуле белка по месту гидроксильных групп серина, треонина;
Гликопротеиды - их простетическими группами являются углеводы;
Хромопротеиды - состоят из простого белка и связанного с ним окрашенного небелкового соединения, все хромопротеиды биологически очень активны; в качестве простетических групп в них могут быть производные порфирина, изоаллоксазина и каротина;
Липопротеиды - простетическая группа липиды - триглицериды (жиры) и фосфатиды;
Нуклеопротеиды - белки, состоящие из простого белка и соединенной с ним нуклеиновой кислоты. Эти белки играют колоссальную роль в жизнедеятельности организма и будут рассмотрены ниже. Они входят в состав любой клетки, некоторые нуклеопротеиды существуют в природе в виде особых частиц, обладающих патогенной активностью (вирусы).
2. По форме частиц - белки делят на фибриллярные (нитеподобные) и глобулярные (сферические) (см. стр 30).
3. По растворимости и особенностям аминокислотного состава выделяют следующие группы простых белков:
Протеиноиды - белки опорных тканей (костей, хрящей, связок, сухожилий, волос, ногтей, кожи и т.д.). Это в основном фибриллярные белки с большой молекулярной массой (> 150000 Да), нерастворимые в обычных растворителях: воде, солевых и водно-спиртовых смесях. Они растворяются только в специфических растворителях;
Протамины (простейшие белки) - белки, растворимые в воде и содержащие 80-90% аргинина и ограниченный набор (6-8) других аминокислот, представлены в молоках различных рыб. Вследствие высокого содержания аргинина имеют основные свойства, их молекулярная масса сравнительно мала и примерно равна 4000-12000 Да. Они являются белковым компонентом в составе нуклеопротеидов;
Гистоны - хорошо растворимы в воде и разбавленных растворах кислот (0,1Н), отличаются высоким содержанием аминокислот: аргинина, лизина и гистидина (не менее 30%) и поэтому обладают основными свойствами. Эти белки в значительных количествах содержатся в ядрах клеток в составе нуклеопротеидов и играют важную роль в регуляции обмена нуклеиновых кислот. Молекулярная масса гистонов невелика и равна 11000-24000 Да;
Глобулины - белки, нерастворимые в воде и солевых растворах с концентрацией соли более 7%. Глобулины полностью осаждаются при 50%-ном насыщении раствора сульфатом аммония. Эти белки отличаются высоким содержанием глицина (3,5%), их молекулярная масса > 100000 Да. Глобулины - слабокислые или нейтральные белки (р1=6-7,3);
Альбумины - белки, хорошо растворимые в воде и крепких солевых растворах, причем концентрация соли (NH 4) 2 S0 4 не должна превышать 50 % от насыщения. При более высокой концентрации альбумины высаливаются. По сравнению с глобулинами эти белки содержат глицина в три раза меньше и имеют молекулярную массу, равную 40000-70000 Да. Альбумины имеют избыточный отрицательный заряд и кислые свойства (pl=4,7) из-за большого содержания глутаминовой кислоты;
Проламины - группа растительных белков, содержащаяся в клейковине злаковых растений. Они растворимы только в 60-80%-ном водном растворе этилового спирта. Проламины имеют характерный аминокислотный состав: в них много (20-50%) глутаминовой кислоты и пролина (10-15%), в связи с чем они и получили свое название. Их молекулярная масса более 100000 Да;
Глютелины - растительные белки нерастворимые в воде, растворах солей и этаноле, но растворимы в разбавленных (0,1Н) растворах щелочей и кислот. По аминокислотному составу и молекулярной массе сходны с проламинами, но аргинина содержат больше, а пролина меньше.
Способы исследования аминокислотного состава
Под действием ферментов пищеварительных соков белки расщепляются на аминокислоты. Были сделаны два важных вывода: 1) в состав белков входят аминокислоты; 2) методами гидролиза может быть изучен химический, в частности амнокислотный, состав белков.
Для изучения аминокислотного состава белков пользуются сочетанием кислотного (НСl), щелочного [Ва(ОН) 2 ] и, реже, ферментативного гидролиза или одним из них. Установлено, что при гидролизе чистого белка, не содержащего примесей, освобождаются 20 различных α-аминокислот. Все другие открытые в тканях животных, растений и микроорганизмов аминокислоты (более 300) существуют в природе в свободном состоянии либо в виде коротких пептидов или комплексов с другими органическими веществами.
Первый этап в определении первичной структуры белков заключается в качественной и количественной оценке аминокислотного состава данного индивидуального белка. Необходимо помнить, что для исследования нужно иметь определённое количество чистого белка, без примесей других белков или пептидов.
Кислотный гидролиз белка
Для определения аминокислотного состава необходимо провести разрушение всех пептидных связей в белке. Анализируемый белок гидролизуют в 6 мол/л НС1 при температуре около 110 °С в течение 24 ч. В результате такой обработки разрушаются пептидные связи в белке, а в гидролизате присутствуют только свободные аминокислоты. Кроме того, глутамин и аспарагин гидролизуются до глутаминовой и аспарагиновой кислот (т.е. разрывается амидная связь в радикале и от них отщепляется аминогруппа).
Разделение аминокислот с помощью ионообменной хроматографии
Смесь аминокислот, полученных кислотным гидролизом белков, разделяют в колонке с катионообменной смолой. Такая синтетическая смола содержит прочно связанные с ней отрицательно заряженные группы (например, остатки сульфоновой кислоты -SO 3 -), к которым присоединены ионы Na + (рис. 1-4).
В катионообменник вносят смесь аминокислот в кислой среде (рН 3,0), где аминокислоты в основном представляют катионы, т.е. несут положительный заряд. Положительно заряженные аминокислоты присоединяются к отрицательно заряженным частицам смолы. Чем больше суммарный заряд аминокислоты, тем прочнее её связь со смолой. Так, аминокислоты лизин, аргинин и гистидин наиболее прочно связываются с катионообменником, а аспарагиновая и глутаминовая кислоты - наиболее слабо.
Высвобождение аминокислот из колонки осуществляют вымыванием (элюированием) их буферным раствором с увеличивающейся ионной силой (т.е. с увеличением концентрации NaCl) и рН. При увеличении рН аминокислоты теряют протон, в результате уменьшается их положительный заряд, а следовательно и прочность связи с отрицательно заряженными частицами смолы.
Каждая аминокислота выходит из колонки при определённом значении рН и ионной силы. Собирая с нижнего конца колонки раствор (элюат) в виде небольших порций, можно получить фракции, содержащие отдельные аминокислоты.
(подробнее «гидролиз» см вопрос №10)
8) Химические связи в структуре белка.



9) Понятие об иерархии и структурной организации белков. (см. вопрос №12)
10) Гидролиз белка. Химизм реакции (ступенчатость, катализаторы, реагенты, условия протекания реакции) – полное описание гидролиза.
11) Химические превращения белков.
Денатурация и ренатурация
При нагревании растворов белков до 60-80% или при действии реагентов, разрушающих нековалентные связи в белках, происходит разрушение третичной (четвертичной) и вторичной структуры белковой молекулы, она принимает в большей или меньшей степени форму беспорядочного случайного клубка. Этот процесс называют денатурацией. В качестве денатурирующих реагентов могут быть кислоты, щелочи, спирты, фенолы, мочевина, гуанидинхлорид и др. Сущность их действия в том, что они образуют водородные связи с =NH и =СО - группами пептидного остова и с кислотными группами радикалов аминокислот, подменяя собственные внутримолекулярные водородные связи в белке вследствие чего вторичная и третичная структуры изменяются. При денатурации падает растворимость белка, он "свертывается" (например, при варке куриного яйца), утрачивается биологическая активность белка. На этом основано, например, применение водного раствора карболовой кислоты (фенола) в качестве антисептика. В определенных условиях при медленном охлаждении раствора денатурированного белка происходит ренатурация - восстановление исходной (нативной) конформации. Это подтверждает тот факт, что характер укладки пептидной цепи определяется первичной структурой.
Процесс денатурации отдельной белковой молекулы, приводящий к распаду её «жёсткой» трёхмерной структуры, иногда называют плавлением молекулы. Практически любое заметное изменение внешних условий, например, нагревание или существенное изменение pH приводит к последовательному нарушению четвертичной, третичной и вторичной структур белка. Обычно денатурация вызывается повышением температуры, действием сильных кислот и щелочей, солей тяжелых металлов, некоторых растворителей (спирт), радиации и др.
Денатурация часто приводит к тому, что в коллоидном растворе белковых молекул происходит процесс агрегации частиц белка в более крупные. Визуально это выглядит, например, как образование «белка» при жарке яиц.
Ренатурация - процесс, обратный денатурации, при котором белки возвращают свою природную структуру. Нужно отметить, что не все белки способны ренатурировать; у большинства белков денатурация необратима. Если при денатурации белка физико-химические изменения связаны с переходом полипептидной цепи из плотно упакованного (упорядоченного) состояния в беспорядочное, то при ренатурации проявляется способность белков к самоорганизации, путь которой предопределён последовательностью аминокислот в полипептидной цепи, то есть её первичной структурой, детерминированной наследственной информацией. В живых клетках данная информация, вероятно, является решающей для преобразования неупорядоченной полипептидной цепи во время или после её биосинтеза на рибосоме в структуру нативной молекулы белка. При нагревании двухцепочечных молекул ДНК до температуры около 100°C водородные связи между основаниями разрываются, и комплементарные цепи расходятся - ДНК денатурирует. Однако при медленном охлаждении комплементарные цепи могут вновь соединяться в регулярную двойную спираль. Эта способность ДНК к ренатурации используется для получения искусственных гибридных молекул ДНК.
Природные белковые тела наделены определенной, строго заданной пространственной конфигурацией и обладают рядом характерных физико-химических и биологических свойств при физиологических значениях температуры и рН среды. Под влиянием различных физических и химических факторов белки подвергаются свертыванию и выпадают в осадок, теряя нативные свойства. Таким образом, под денатурацией следует понимать нарушение общего плана уникальной структуры нативной молекулы белка, преимущественно ее третичной структуры, приводящее к потере характерных для нее свойств (растворимость, электрофоретическая подвижность, биологическая активность и т.д.). Большинство белков денатурирует при нагревании их растворов выше 50–60°С.
Внешние проявления денатурации сводятся к потере растворимости, особенно в изоэлектрической точке, повышению вязкости белковых растворов, увеличению количества свободных функциональных SH-групп и изменению характера рассеивания рентгеновских лучей. Наиболее характерным признаком денатурации является резкое снижение или полная потеря белком его биологической активности (каталитической, антигенной или гормональной). При денатурации белка, вызванной 8М мочевиной или другим агентом, разрушаются в основном нековалентные связи (в частности, гидрофобные взаимодействия и водородные связи). Дисульфидные связи в присутствии восстанавливающего агента меркаптоэтанола разрываются, в то время как пептидные связи самого остова полипептидной цепи не затрагиваются. В этих условиях развертываются глобулы нативных белковых молекул и образуются случайные и беспорядочные структуры (рис.)

Денатурация белковой молекулы (схема).
а - исходное состояние; б - начинающееся обратимое нарушение молекулярной структуры; в - необратимое развертывание полипептидной цепи.

Денатурация и ренатурация рибонуклеазы (по Анфинсену).
а - развертывание (мочевина + меркаптоэтанол); б - повторное свертывание.

1. Гидролиз белков: H+
[− NH2─CH─ CO─NH─CH─CO − ]n +2nH2O → n NH2 − CH − COOH + n NH2 ─ CH ─ COOH
│ │ │ │
Аминокислота 1 аминокислота 2
2. Осаждение белков:
а) обратимое
Белок в растворе ↔ осадок белка. Происходит под действием растворов солей Na+, K+
б) необратимое (денатурация)
При денатурации под действием внешних факторов (температура; механическое воздействие – давление, растирание, встряхивание, ультразвук; действия химических агентов – кислот, щелочей и др.) происходит изменение вторичной, третичной и четвертичной структур белковой макромолекулы, т.е её нативной пространственной структуры. Первичная структура, а, следовательно, и химический состав белка не меняются.
При денатурации изменяются физические свойства белков: снижается растворимость, теряется биологическая активность. В тоже время увеличивается активность некоторых химических групп, облегчается воздействие на белки протеолитических ферментов, а, следовательно, он легче гидролизуется.
Например, альбумин - яичный белок - при температуре 60-70° осаждается из раствора (свертывается), теряя способность растворяться в воде.
Схема процесса денатурации белка (разрушение третичной и вторичной структур белковых молекул)
3. Горение белков
Белки горят с образованием азота, углекислого газа, воды, а также некоторых других веществ. Горение сопровождается характерным запахом жженых перьев
4. Цветные (качественные) реакции на белки:
а) ксантопротеиновая реакция (на остатки аминокислот, содержащих бензольные кольца):
Белок + HNO3 (конц.) → желтое окрашивание
б) биуретовая реакция (на пептидные связи):
Белок + CuSO4 (насыщ) + NaOH (конц) → ярко-фиолетовое окрашивание
в) цистеиновая реакция (на остатки аминокислот, содержащих серу):
Белок + NaOH + Pb(CH3COO)2 → Черное окрашивание
Белки являются основой всего живого на Земле и выполняют в организмах многообразные функции.
Высаливание белков
Высаливанием называется процесс выделения белков из водных растворов нейтральными растворами концентрированных солей щелочных и щелочноземельных металлов. При добавлении больших концентраций солей к раствору белка происходит дегидратация белковых частиц и снятие заряда, при этом белки выпадают в осадок. Степень выпадения белков в осадок зависит от ионной силы раствора осадителя, размера частиц белковой молекулы, величины ее заряда, гидрофильности. Разные белки осаждаются при различных концентрациях солей. Поэтому в осадках, полученных путем постепенного повышения концентрации солей, отдельные белки находятся в различных фракциях. Высаливание белков является обратимым процессом, и после удаления соли белок вновь приобретает природные свойства. Поэтому высаливанием пользуются в клинической практике при разделении белков сыворотки крови, а также при изолировании, очистке различных белков.
Добавляемые анионы и катионы разрушают гидратную белковую оболочку белков, являющуюся одним из факторов устойчивости белковых растворов. Чаще всего применяются растворы сульфатов Na и аммония. Многие белки отличаются по размеру гидратной оболочки и величине заряда. Для каждого белка есть своя зона высаливания. После удаления высаливающего агента белок сохраняет свою биологическую активность и физико-химические свойства. В клинической практике применяется метод высаливания для разделения глобулинов (при добавлении 50% раствора сульфата аммония (NH4)2SO4 выпадает осадок) и альбуминов (при добавлении 100% раствора сульфата аммония (NH4)2SO4 выпадает осадок).
На величину высаливания оказывают влияние:
1) природа и концентрация соли;
2) рН-среды;
3) температура.
Главную роль при этом играют валентности ионов.
12) Особенности организации первичной, вторичной, третичной структуры белка.
В настоящее время экспериментально доказано существование четырёх уровней структурной организации белковой молекулы: первичной, вторичной, третичной и четвертичной структуры.
Аминокислотный состав и пространственная организация каждого белка определяют его физико-химические свойства. Белки обладают кислотно-основными, буферными, коллоидными и осмотическими свойствами.
Белки как амфотерные макромолекулы
Белки являются амфотерными полиэлектролитами, т.е. сочетают в себе, подобно аминокислотам, кислотные и основные свойства. Однако природа групп, придающих амфотерные свойства белкам, далеко не та же, что у аминокислот. Кислотно-основные свойства аминокислот обусловлены прежде всего наличием α-амино- и α-карбоксильной групп (кислотно-основная пара). В молекулах белков эти группы участвуют в образовании пептидных связей, а амфотерность белкам придают кислотно-основные группы боковых радикалов аминокислот, входящих в белок. Разумеется, в каждой молекуле нативного белка (полипептидной цепи) имеется как минимум по одной концевой α-амино- и α-карбоксильной группе (если у белка только третичная структура). У белка с четвертичной структурой число концевых групп -NН 2 и -СООН равно числу субъединиц, или протомеров. Однако столь незначительное число этих групп не может объяснить амфотерность макромолекул белка. Поскольку большая часть полярных групп находится на поверхности глобулярных белков, то именно они определяют кислотно-основные свойства и заряд белковой молекулы. Кислотные свойства белку придают кислые аминокислоты (аспарагиновая, глутаминовая и аминолимонная), а щелочные свойства - основные аминокислоты (лизин, аргинин, гистидин). Чем больше кислых аминокислот содержится в белке, тем ярче выражены его кислотные свойства, и чем больше входит в состав белка основных аминокислот, тем сильнее проявляются его основные свойства. Слабая диссоциация SН-группы цистеина и фенольной группы тирозина (их можно рассматривать как слабые кислоты) почти не влияет на амфотерность белков.
Буферные свойства . Белки хотя и обладают свойствами буфера, но емкость их при физиологических значениях рН ограничена. Исключение составляют белки, содержащие много гистидина, так как только боковая группа гистидина обладает буферными свойствами в интервале значений рН, близких к физиологическим. Таких белков очень мало. Гемоглобин чуть ли не единственный белок, содержащий до 8% гистидина, является мощным внутриклеточным буфером в эритроцитах, поддерживая рН крови на постоянном уровне.
Заряд белковой молекулы зависит от содержания в ней кислых и основных аминокислот, а точнее, от ионизации кислых и основных групп бокового радикала этих аминокислот. Диссоциация СООН-групп кислых аминокислот вызывает появление отрицательного заряда на поверхности белка, а боковые радикалы щелочных аминокислот несут положительный заряд (за счет присоединения Н + к основным группам). В нативной молекуле белка заряды распределяются асимметрично в зависимости от укладки полипептидной цепи в пространстве. Если в белке кислые аминокислоты преобладают над основными, то в целом молекула белка электроотрицательна, т. е. является полианионом, и наоборот, если преобладают основные аминокислоты, то она заряжена положительно, т. е. ведет себя как поликатион.
Суммарный заряд белковой молекулы, естественно, зависит от рН среды: в кислой среде он положителен, в щелочной отрицателен. То значение рН, при котором белок имеет суммарный нулевой заряд, называется изоэлектрической точкой данного белка. В этой точке белок не обладает подвижностью в электрическом поле. Изоэлектрическая точка каждого белка определяется соотношением кислых и основных групп боковых радикалов аминокислот: чем выше соотношение кислые/основные аминокислоты в белке, тем ниже его изоэлектрическая точка. У кислых белков рН 1 < 7, у нейтральных рН 1 около 7, а у основных рН 1 > 7. При значениях рН среды ниже его изоэлектрической точки белок будет нести положительный заряд, а выше - отрицательный заряд. Усредненная изоэлектрическая точка всех белков цитоплазмы лежит в пределах 5,5. Следовательно, при физиологическом значении рН (около 7,0 - 7,4) клеточные белки имеют общий отрицательный заряд. Избыток отрицательных зарядов белков внутри клетки уравновешивается, как уже говорилось, неорганическими катионами.
Знание изоэлектрической точки очень важно для понимания стабильности белков в растворах, так как в изоэлектрическом состоянии белки наименее устойчивы. Незаряженные частицы белка могут слипаться друг с другом и выпадать в осадок.
Коллоидные и осмотические свойства белков
Поведение белков в растворах имеет некоторые особенности. Обычные коллоидные растворы устойчивы только в присутствии стабилизатора, который препятствует осаждению коллоидов, располагаясь на границе раздела "растворенное вещество - растворитель".
Водные растворы белков являются устойчивыми и равновесными, они со временем не выпадают в осадок (не коагулируют) и не требуют присутствия стабилизаторов. Белковые растворы гомогенны и, в сущности, их можно отнести к истинным растворам. Однако высокая молекулярная масса белков придает их растворам многие свойства коллоидных систем:
- характерные оптические свойства (опалесценция растворов и способность их рассеивать лучи видимого света)
[показать]
.
Оптические свойства белков . Растворы белков, особенно концентрированные, обладают характерной опалесценцией. При боковом освещении раствора белка лучи света в нем становятся видимыми и образуют светящийся конус или полосу - эффект Тиндаля (в сильно разбавленных растворах белка не видна опалесценция и почти отсутствует светящийся конус Тиндаля). Объясняется этот светорассеивающий эффект дифракцией лучей света частицами белка в растворе. Считается, что в протоплазме клетки белок находится в виде коллоидного раствора - золя. Способность белков и других биологических молекул (нуклеиновых кислот, полисахаридов и т. д.) рассеивать свет используется при микроскопическом изучении клеточных структур: в темном поле микроскопа коллоидные частицы видны как светлые вкрапления в цитоплазме.
Светорассеивающую способность белков и других высокомолекулярных веществ используют для их количественного определения методом нефелометрии, сравнивая интенсивность светорассеивания взвешенными частицами исследуемого и стандартного золя.
- малая скорость диффузии
[показать]
.
Малая скорость диффузии . Диффузией называется самопроизвольное перемещение молекул растворенных веществ вследствие градиента концентраций (от зон с высокой концентрацией к зонам с низкой концентрацией). Белки имеют ограниченную скорость диффузии в сравнении с обычными молекулами и ионами, которые перемещаются в сотни и тысячи раз быстрее, чем белки. Скорость диффузии белков больше зависит от формы их молекул, чем от молекулярной массы. Глобулярные белки в водных растворах подвижнее фибриллярных белков.
Диффузия белков имеет важное значение для нормального функционирования клетки. Синтез белков в любом участке клетки (там, где имеются рибосомы) мог бы привести при отсутствии диффузии к скоплению белков в месте их образования. Внутриклеточное распределение белков происходит путем диффузии. Поскольку скорость диффузии белков невысока, она ограничивает скорость процессов, зависящих от функции диффундирующего белка в соответствующем участке клетки.
- неспособность проникать через полупроницаемые мембраны
[показать]
.
Осмотические свойства белков . Белки из-за высокой молекулярной массы не могут диффундировать через полупроницаемую мембрану, тогда как низкомолекулярные вещества легко проходят через такие мембраны. Это свойство белков используют в практике для очистки их растворов от низкомолекулярных примесей. Такой процесс называется диализом.
Неспособность белков диффундировать через полупроницаемые мембраны вызывает явление осмоса, т. е. перемещение молекул воды через полупроницаемую мембрану в раствор белка. Если раствор белка отделить от воды целлофановой мембраной, то, стремясь к достижению равновесия, молекулы воды диффундируют в раствор белка. Однако перемещение воды в пространство, где находится белок, повышает в нем гидростатическое давление (давление столба воды), которое препятствует дальнейшей диффузии молекул воды к белку.
То давление, или сила, которое следует приложить, чтобы остановить осмотический ток воды, называется осмотическим давлением. Осмотическое давление в очень разбавленных растворах белка пропорционально молярной концентрации белка и абсолютной температуре.
Биологические мембраны также непроницаемы для белка, поэтому осмотическое давление, создаваемое белком, зависит от концентрации его внутри и вне клетки. Осмотическое давление, обусловленное белком, называют также онкотическим давлением.
- высокая вязкость растворов
[показать]
.
Высокая вязкость растворов белка . Высокая вязкость характерна не только для растворов белка, но вообще для растворов высокомолекулярных соединений. С увеличением концентрации белка вязкость раствора повышается, поскольку повышаются силы сцепления между молекулами белка. Вязкость зависит от формы молекул. Растворы фибриллярных белков всегда более вязки, чем растворы глобулярных белков. На вязкость растворов сильно влияют температура и присутствие электролитов. С повышением температуры вязкость растворов белка снижается. Добавки некоторых солей, например кальция, повышают вязкость, способствуя сцеплению молекул с помощью кальциевых мостикoв. Иногда вязкость белкового раствора увеличивается настолько, что он теряет текучесть и переходит в гелеобразное состояние.
- способность к образованию гелей
[показать]
.
Способность белков к образованию гелей . Взаимодействие между макромолекулами белка в растворе может привести к образованию структурных сеток, внутри которых находятся захваченные молекулы воды. Такие структурированные системы называются гелями или студнями. Считается, что белок протоплазмы клетки может переходить в гелеобразное состояние. Характерный пример - тело медузы является как бы живым студнем, содержание воды в котором до 90%.
Гелеобразование легче протекает в растворах фибриллярных белков; их палочковидная форма способствует лучшему контакту концов макромолекул. Это хорошо известно из бытовой практики. Пищевые студни готовят из продуктов (кости, хрящи, мясо), содержащих в большом количестве фибриллярные белки.
В процессе жизнедеятельности организма гелеобразное состояние белковых структур имеет важное физиологическое значение. Коллагеновые белки костей, сухожилий, хрящей, кожи и т. д. обладают высокой прочностью, упругостью и эластичностью, потому что находятся в гелеобразном состоянии. Отложение минеральных солей при старении снижает их упругость и эластичность. В гелеобразном или студнеобразном виде находится в мышечных клетках актомиозин, выполняющий сократительную функцию.
В живой клетке происходят процессы, напоминающие переход золь - гель. Протоплазма клетки представляет собой золеподобную вязкую жидкость, в которой обнаруживаются островки гелеподобных структур.
Гидратация белков и факторы, влияющие на их растворимость
Белки - гидрофильные вещества. Если растворять сухой белок в воде, то сначала он, как всякое гидрофильное высокомолекулярное соединение, набухает, а затем молекулы белка начинают постепенно переходить в раствор. При набухании молекулы воды проникают в белок и связываются с его полярными группами. Плотная упаковка полипептидных цепей разрыхляется. Набухший белок можно считать как бы обратным раствором, т. е. раствором молекул воды в высокомолекулярном веществе - белке. Дальнейшее поглощение воды приводит к отрыву молекул белка от общей массы и растворению. Но набухание не всегда ведет к растворению; некоторые белки, например коллаген, так и остаются в набухшем виде, поглотив большое количество воды.
Растворение связано с гидратацией белков, т. е. связыванием молекул воды с белками. Гидратная вода так прочно связана с макромолекулой белка, что отделить ее удается с большим трудом. Это говорит не о простой адсорбции, а об электростатическом связывании молекул воды с полярными группами боковых радикалов кислых аминокислот, несущих отрицательный заряд, и основных аминокислот, несущих положительный заряд.
Однако часть гидратной воды связывается пептидными группами, которые образуют с молекулами воды водородные связи. Например, полипептиды с неполярными боковыми группами тоже набухают, т. е. связывают воду. Так, большое количество воды связывает коллаген, хотя этот белок содержит преимущественно неполярные аминокислоты. Вода, связываясь с пептидными группами, раздвигает вытянутые полипептидные цепи. Однако межцепочечные связи (мостики) не дают молекулам белка отрываться друг от друга и переходить в раствор. При нагревании сырья, содержащего коллаген, межцепочечные мостики в коллагеновых волокнах разрываются и освобожденные полипептидные цепи переходят в раствор. Эта фракция частично гидролизованного растворимого коллагена называется желатиной. Желатина по химическому составу близка к коллагену, легко набухает и растворяется в воде, образуя вязкие жидкости. Характерным свойством желатины является способность к гелеобразованию. Водные растворы желатины широко используются в лечебной практике как плазмозамещающее и кровоостанавливающее средство, а способность к гелеобразованию - при изготовлении капсул в фармацевтической практике.
Факторы, влияющие на растворимость белков . Растворимость разных белков колеблется в широких пределах. Она определяется их аминокислотным составом (полярные аминокислоты придают большую растворимость, чем неполярные), особенностями организации (глобулярные белки, как правило, лучше растворимы, чем фибриллярные) и свойствами растворителя. Например, растительные белки - проламины - растворяются в 60-80%-ном спирте, альбумины - в воде и в слабых растворах солей, а коллаген и кератины нерастворимы в большинстве растворителей.
Стабильность растворам белков придают заряд белковой молекулы и гидратная оболочка. Каждая макромолекула индивидуального белка имеет суммарный заряд одного знака, что препятствует их склеиванию в растворе и выпадению в осадок. Все, что способствует сохранению заряда и гидратной оболочки, облегчает растворимость белка и его устойчивость в растворе. Между зарядом белка (или числом полярных аминокислот в нем) и гидратацией существует тесная связь: чем больше полярных аминокислот в белке, тем больше связывается воды (в расчете на 1 г белка). Гидратная оболочка белка иногда достигает больших размеров, и гидратная вода может составлять до 1/5 его массы.
Правда, некоторые белки гидратируются сильнее, а растворяются хуже. Например, коллаген связывает воды больше, чем многие хорошо растворимые глобулярные белки, но не растворяется. Его растворимости мешают структурные особенности - поперечные связи между полипептидными цепями. Иногда разноименно заряженные группы белка образуют много ионных (солевых) связей внутри молекулы белка или между молекулами белков, что мешает образованию связей между молекулами воды и заряженными группами белков. Наблюдается парадоксальное явление: в белке много анионных или катионных групп, а растворимость его в воде низкая. Межмолекулярные солевые мостики вызывают склеивание молекул белка и их выпадение в осадок.
Какие же факторы среды влияют на растворимость белков и их стабильность в растворах?
- Влияние нейтральных солей
[показать]
.
Нейтральные соли в небольших концентрациях повышают растворимость даже тех белков, которые нерастворимы в чистой воде (например, эвглобулины). Это объясняется тем, что ионы солей, взаимодействуя с противоположно заряженными группами молекул белков, разрушают солевые мостики между молекулами белков. Повышение концентрации солей (увеличение ионной силы раствора) оказывает обратное действие (см. ниже - высаливание).
- Влияние рН среды
[показать]
.
рН среды влияет на заряд белка, а следовательно, на его растворимость. Наименее устойчив белок в изоэлектрическом состоянии, т. е. когда его суммарный заряд равен нулю. Снятие заряда позволяет молекулам белка легко сближаться, склеиваться и выпадать в осадок. Значит, растворимость и устойчивость белка будут минимальны при рН, соответствующем изоэлектрической точке белка.
- Влияние температуры
[показать]
.
Строгой зависимости между температурой и характером растворимости белков не имеется. Одни белки (глобулины, пепсин, фосфорилаза мышц) в водных или солевых растворах с повышением температуры растворяются лучше; другие (альдолаза мышц, гемоглобин и т.д.) хуже.
- Влияние разнозаряженного белка
[показать]
.
Если в раствор белка, являющегося полианионом (кислый белок), добавить белок, являющийся поликатионом (основной белок), то они образуют агрегаты. При этом устойчивость вследствие нейтрализации зарядов теряется и белки выпадают в осадок. Иногда эту особенность используют для выделения нужного белка из смеси белков.
Высаливание
Растворы нейтральных солей широко используются не только для повышения растворимости белка, например при выделении его из биологического материала, но и для избирательного осаждения разных белков, т. е. их фракционирования. Процесс осаждения белков нейтральными солевыми растворами называется высаливанием. Характерной особенностью белков, полученных высаливанием, является сохранение ими нативных биологических свойств после удаления соли.
Механизм высаливания состоит в том, что добавляемые анионы и катионы солевого раствора снимают гидратную оболочку белков, являющуюся одним из факторов его устойчивости. Возможно, одновременно происходит и нейтрализация зарядов белка ионами соли, что также способствует осаждению белков.
Способность к высаливанию наиболее выражена у анионов солей. По силе высаливающего действия анионы и катионы располагаются в следующие ряды:
- SO 4 2- > С 6 Н 5 О 7 3- > СН 3 СОО - > Сl - > NO 3 - > Вr - > I - > CNS -
- Li + >Na + > К + > Pb + > Сs +
Эти ряды называются лиотропными.
Сильным высаливающим эффектом в этом ряду обладают сульфаты. На практике для высаливания белков чаще всего применяют сульфат натрия и аммония. Кроме солей белки осаждают органическими водоотнимающими средствами (этанол, ацетон, метанол и др.). Фактически это то же высаливание.
Высаливание широко используют для разделения и очистки белков, поскольку многие белки различаются по размеру гидратной оболочки и величине зарядов. Для каждого из них имеется своя зона высаливания, т. е. концентрация соли, позволяющая дегидратировать и осадить белок. После удаления высаливающего агента белок сохраняет все свои природные свойства и функции.
Денатурация (денативация) и ренатурация (ренативация)
При действии различных веществ, нарушающих высшие уровни организации белковой молекулы (вторичную, третичную, четвертичную) с сохранением первичной структуры, белок теряет свои нативные физико-химические и, главное, биологические свойства. Это явление называется денатурацией (денативацией). Оно характерно только для молекул, имеющих сложную пространственную организацию. Синтетические и природные пептиды не способны к денатурации.
При денатурации разрываются связи, стабилизирующие четвертичную, третичную и даже вторичную структуры. Полипептидная цепь разворачивается и находится в растворе или в развернутом виде, или в виде беспорядочного клубка. При этом теряется гидратная оболочка и белок выпадает в осадок. Однако осажденный денатурированный белок отличается от того же белка, осажденного путем высаливания, так как в первом случае он утрачивает нативные свойства, а во втором сохраняет. Это указывает на то, что механизм действия веществ, вызывающих денатурацию и высаливание, разный. При высаливании сохраняется нативная структура белка, а при денатурации разрушается.
Денатурирующие факторы делятся на
- физические
[показать]
.
К физическим факторам относятся: температура, давление, механическое воздействие, ультразвуковое и ионизирующее излучение.
Тепловая денатурация белков является наиболее изученным процессом. Она считалась одним из характерных признаков белков. Давно известно, что при нагревании белок свертывается (коагулирует) и выпадает в осадок. Большинство белков термолабильны, однако известны белки, очень устойчивые к нагреванию. Например, трипсин, химотрипсин, лизоцим, некоторые белки биологических мембран. Особой устойчивостью к температуре отличаются белки бактерий, обитающих в горячих источниках. Очевидно, у термостабильных белков тепловое движение полипептидных цепей, вызванное нагреванием, недостаточно для разрыва внутренних связей молекул белка. В изоэлектрической точке белки легче подвергаются тепловой денатурации. Этот прием используется в практической работе. Некоторые белки, наоборот, денатурируют при низкой температуре.
- химические
[показать]
.
К химическим факторам, вызывающим денатурацию, относятся: кислоты и щелочи, органические растворители (спирт, ацетон), детергенты (моющие средства), некоторые амиды (мочевина, соли гуанидина и т. д.), алкалоиды, тяжелые металлы (соли ртути, меди, бария, цинка, кадмия и т. д.). Механизм денатурирующего действия химических веществ зависит от их физико-химических свойств.
Кислоты и щелочи широко используются в качестве осадителей белков. Многие белки денатурируются при крайних значениях рН - ниже 2 или выше 10-11. Но некоторые белки устойчивы к действию кислот и щелочей. Например, гистоны и протамины не денатурируются даже при рН 2 или рН 10. Крепкие растворы этанола, ацетон тоже оказывают денатурирующее влияние на белки, хотя для некоторых белков эти органические растворители используются как высаливающие агенты.
Тяжелые металлы, алкалоиды издавна применяются как осадители; они образуют прочные связи с полярными группами белков и тем самым разрывают систему водородных и ионных связей.
Особо следует остановиться на мочевине и солях гуанидина, которые в больших коцентрациях (для мочевины 8 моль/л, для гуанидина гидрохлорида 2 моль/л) конкурируют пептидными группами за образование водородных связей. В результате происходит диссоциация на субъединицы у белков с четвертичной структурой, а затем и разворачивание полипептидных цепей. Это свойство мочевины настолько ярко, что его широко используют для доказательства наличия четвертичной структуры белка и значения его структурной организации в осуществлении физиологической функции.
Свойства денатурированных белков . Наиболее типичными для денатурированных белков являются следующие признаки.
- Увеличение числа реактивных или функциональных групп по сравнению с нативной молекулой белка (функциональными группами называются группы боковых радикалов аминокислот: СООН, NН 2 , SН, ОН). Часть этих групп обычно находится внутри молекулы белка и не выявляется специальными реагентами. Развертывание полипептидной цепи при денатурации позволяет обнаружить эти дополнительные, или скрытые, группы.
- Уменьшение растворимости и осаждение белка (связано с потерей гидратной оболочки, развертыванием молекулы белка с "обнажением" гидрофобных радикалов и нейтрализацией зарядов полярных групп).
- Изменение конфигурации молекулы белка.
- Потеря биологической активности, вызванная нарушением нативной структурной организации молекулы.
- Более легкое расщепление протеолитическими ферментами по сравнению с нативным белком переход компактной нативной структуры в развернутую рыхлую форму облегчает доступ ферментов к пептидным связям белка, которые они разрушают.
Последнее качество денатурированного белка широко известно. Термическая или иная обработка продуктов, содержащих белки (главным образом мясные), способствует лучшему перевариванию их с помощью протеолитических ферментов желудочно-кишечного тракта. В желудке человека и животных вырабатывается природный денатурирующий агент - соляная кислота, которая, денатурируя белки, помогает их расщеплению ферментами. Однако наличие соляной кислоты и протеолитических ферментов не позволяет применять белковые лекарственные препараты через рот, ибо они денатурируются и тут же расщепляются, теряя биологическую активность.
Заметим также, что денатурирующие вещества, осаждающие белки, используются в биохимической практике с иными целями, чем высаливающие. Высаливание как прием применяется для выделения какого-то белка или группы белков, а денатурация для освобождения от белка смеси каких-либо веществ. Удаляя белок, можно получить безбелковый раствор или устранить действие этого белка.
Долго считалось, что денатурация необратима. Однако в некоторых случаях удаление денатурирующего агента (такие опыты были сделаны при использовании мочевины) восстанавливает биологическую активность белка. Процесс восстановления физико-химических и биологических свойств денатурированного белка называется ренатурацией или ренативацией. Если денатурированный белок (после удаления денатурирующих веществ) вновь самоорганизуется в исходную структуру, то восстанавливается и его биологическая активность.
| Страница 4 | всего страниц: 7 |
Форма белковой молекулы . Исследования нативной конформации белковых молекул показали, что эти частицы в большинстве случаев имеют более или менее асимметричную форму. В зависимости от степени асимметрии, т. е. соотношения между длинной (b) и короткой (а) осями белковой молекулы различают глобулярные (шаровидные) и фибриллярные (нитевидные) белки.
Глобулярными являются белковые молекулы, у которых свертывание полипептидных цепочек привело к образованию сферической структуры. Среди них встречаются строго шаровидные, эллипсовидные и палочкообразные. Они различаются по степени асимметрии. Например, яичный альбумин имеет b/а = 3, глиадин пшеницы - 11, а зеин кукурузы - 20. Многие белки в живой природе являются глобулярными.
Фибриллярные белки образуют длинные высокоасимметричные нити. Многие из них выполняют структурную или механическую функцию. Таковы коллаген (b/а — 200), кератины, фиброин.
Белкам каждой из групп присущи свои характерные свойства. Многие глобулярные белки растворимы в воде и разбавленных солевых растворах. Растворимым фибриллярным белкам свойственны очень вязкие растворы. Глобулярные белки, как правило, обладают хорошей биологической ценностью - усваиваются в процессе пищеварения, в то время как многие фибриллярные белки - нет.
Между глобулярными и фибриллярными белками отсутствует четкая граница. Ряд белков занимает промежуточное положение и сочетает в себе признаки как глобулярных, так и фибриллярных. К таким белкам относятся, например, миозин мышц (b/а = 75) и фибриноген крови (b/а = 18). Миозин имеет палочковидную форму, сходную с формой фибриллярных белков, однако, подобно глобулярным белкам, он растворим в солевых растворах. Растворы миозина и фибриногена вязкие. Эти белки усваиваются в процессе пищеварения. В то же время актин - глобулярный белок мышц - не усваивается.
Денатурация белка . Нативная конформация белковых молекул не является жесткой, она довольно лабильна (лат. «labilis» - скользящий) и при ряде воздействий может серьезно нарушаться. Нарушение нативной конформации белка, сопровождающееся изменением его нативных свойств без разрыва пептидных связей, называется денатурацией (лат. «denaturare » - лишать природных свойств) белка.
Денатурация белков может быть вызвана различными при-чинами, приводящими к нарушению слабых взаимодействий, а также к разрыву дисульфидных связей, стабилизирующих их нативную структуру.
Нагревание большинства белков до температуры выше 50°С, а также ультрафиолетовое и другие виды высокоэнергетического облучения усиливают колебания атомов полипептидной цепи, что приводит к нарушению в них различных связей. Денатурацию белка способно вызвать даже механическое встряхивание.
Денатурация белков также происходит вследствие химического воздействия. Сильные кислоты или щелочи влияют на ионизацию кислотных и основных групп, вызывая нарушение ионных и некоторых водородных связей в молекулах белков. Мочевина (H 2 N-CO-NH 2) и органические растворители - спирты, фенолы и др. - нарушают систему водородных связей и ослабляют в белковых молекулах гидрофобные взаимодействия (мочевина - за счет нарушения структуры воды, органические растворители - вследствие установления контактов с неполярными радикалами аминокислот). Меркаптоэтанол разрушает в белках дисульфидные связи. Ионы тяжелых металлов нарушают слабые взаимодействия.
При денатурации происходит изменение свойств белка и, в первую очередь, уменьшение его растворимости. Например, при кипячении белки коагулируют и выпадают из растворов в осадок в виде сгустков (как при варке куриного яйца). Осаждение белков из растворов происходит также под воздействием белковых осадителей, в качестве которых применяют трихлоруксусную кислоту, реактив Барнштейна (смесь гидроксида натрия с сульфатом меди), раствор таннина и др.
При денатурации уменьшается водопоглотительная способность белка, т. е. его способность к набуханию; могут появляться новые химические группы, например: при воздействии мер каптоэтанола - SH-группы. В результате денатурации белок теряет свою биологическую активность.
Хотя первичная структура белка при денатурации не нарушается, изменения являются необратимыми. Однако, например, при постепенном удалении мочевины методом диализа из раствора денатурированного белка происходит его ренатурация: нативная структура белка восстанавливается, а вместе с ней, в той или иной степени, - и его нативные свойства. Такая денатурация называется обратимой .
Необратимая денатурация белков происходит в процессе старения организмов. Поэтому, например, семена растений, даже при оптимальных условиях хранения, постепенно теряют свою всхожесть.
Денатурация белков имеет место при выпечке хлеба, сушке макарон, овощей, в ходе приготовления пищи и т. д. В результате повышается биологическая ценность этих белков, так как в процессе пищеварения легче усваиваются денатурированные (частично разрушенные) белки.
Изоэлектрическая точка белка . В белках содержатся раз-личные основные и кислотные группы, которые обладают способностью к ионизации. В сильнокислой среде активно протонируются основные группировки (аминогруппы и др.), и молекулы белка приобретают суммарный положительный заряд, а в сильнощелочной среде - легко диссоциируют карбоксильные группы, и молекулы белка приобретают суммарный отрицательный заряд.
Источниками положительного заряда в белках выступают боковые радикалы остатков лизина, аргинина и гистидина, а-аминогруппа остатка N-концевой аминокислоты. Источники отрицательного заряда - боковые радикалы остатков аспарагиновой и глутаминовой кислот, а-карбоксильная группа остатка С-концевой аминокислоты.
При определенном значении рН среды наблюдается равенство положительных и отрицательных зарядов на поверхности белковой молекулы, т. е. ее суммарный электрический заряд оказывается равным нулю. Такое значение рН раствора, при котором молекула белка электронейтральна, называют изоэлектрической точкой белка (pi).
Изоэлектрические точки являются характерными константами белков. Они определяются их аминокислотным составом и структурой: количеством и расположением остатков кислых и основных аминокислот в полипептидных цепях. Изоэлектрические точки белков, в которых преобладают остатки кислых аминокислот, располагаются в области рН<7, а белков, в которых преобладают остатки основных аминокислот - в области рН>7. Изоэлектрические точки большинства белков находятся в слабокислой среде.
В изоэлектрическом состоянии растворы белков обладают минимальной вязкостью. Это связано с изменением формы белковой молекулы. В изоэлектрической точке разноименно заряженные группы притягиваются друг к другу, и белки закручиваются в клубки. При смещении рН от изоэлектрической точки одноименно заряженные группы отталкиваются, и молекулы белка развертываются. В развернутом состоянии белковые молекулы придают растворам более высокую вязкость, чем свернутые в клубки.
В изоэлектрической точке белки обладают минимальной растворимостью и могут легко выпадать в осадок.
Однако осаждения белков в изоэлектрической точке все же не происходит. Этому препятствуют структурированные молекулы воды, удерживающие на поверхности белковых глобул значительную часть гидрофобных аминокислотных радикалов.
Осадить белки можно с помощью органических растворителей (спирта, ацетона), нарушающих систему гидрофобных контактов в молекулах белка, а также высоких концентраций солей (методом высаливания), уменьшающих гидратацию белковых глобул. В последнем случае часть воды идет на растворение соли и перестает участвовать в растворении белка. Такой раствор за недостатком растворителя становится пересыщенным, что влечет за собой выпадение части его в осадок. Белковые молекулы начинают слипаться и, образуя все более крупные частицы, постепенно осаждаться из раствора.
Оптические свойства белка . Растворы белков обладают оптической активностью, т. е. способностью вращать плоскость поляризации света. Это свойство белков обусловлено наличием в их молекулах элементов асимметрии - асимметрических атомов углерода и правозакрученной а-спирали.
При денатурации белка происходит изменение его оптических свойств, что связано с разрушением а-спирали. Оптические свойства полностью денатурированных белков зависят только от наличия в них асимметрических атомов углерода.
По разнице в проявлении белком оптических свойств до и после денатурации можно определить степень его спирализации.
Качественные реакции на белки . Для белков характерны цветные реакции, обусловленные наличием в них тех или иных химических группировок. Эти реакции часто используются для обнаружения белков.
При добавлении к белковому раствору сульфата меди и щелочи появляется сиреневое окрашивание, связанное с образованием комплексов ионов меди с пептидными группами белка. Поскольку эту реакцию дает биурет (H 2 N-CO-NH-CO-NH 2), она получила название биуретовой. Ее часто используют для количественного определения белка, наряду с методом И. Кьельдаля, так как интенсивность возникающей окраски пропорциональна концентрации белка в растворе.
При нагревании растворов белков с концентрированной азотной кислотой появляется желтое окрашивание, обусловленное образованием нитропроизводных ароматических аминокислот. Эту реакцию называют ксантопротеиновой (греч. «ксантос» - желтый).
Многие белковые растворы при нагревании вступают в реакцию с азотнокислым раствором ртути, которая образует с фенолами и их производными комплексные соединения малинового цвета. Это качественная реакция Миллона на тирозин.
В результате нагревания большинства белковых растворов с уксуснокислым свинцом в щелочной среде выпадает черный осадок сульфида свинца. Данная реакция используется для обнаружения серосодержащих аминокислот и называется реакцией Фоля.
 Материк евразия краткое описание для детей Местность евразии
Материк евразия краткое описание для детей Местность евразии Исследовательская работа " Проблемы молодежи" творческая работа учащихся по немецкому языку (9 класс) на тему Топик по немецкому проблемы молодежи
Исследовательская работа " Проблемы молодежи" творческая работа учащихся по немецкому языку (9 класс) на тему Топик по немецкому проблемы молодежи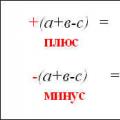 Восприятие учебной информации, психофизиологические особенности Психологические условия развития восприятия учебного материала учащимися
Восприятие учебной информации, психофизиологические особенности Психологические условия развития восприятия учебного материала учащимися