Millised organismid arendasid esmakordselt närvisüsteemi? Millisel loomal tekkis esmakordselt närvisüsteem? Närvisüsteemi üldised omadused
Evolutsioonis on närvisüsteem läbinud mitmeid arenguetappe, millest said pöördepunktid selle tegevuse kvalitatiivses korraldamises. Need etapid erinevad neuronaalsete moodustiste, sünapside arvu ja tüüpide, nende funktsionaalse spetsialiseerumise tunnuste ja ühiste funktsioonidega omavahel seotud neuronirühmade moodustumise poolest. Närvisüsteemi struktuursel korraldusel on kolm peamist etappi: difuusne, sõlmeline, torukujuline.
Hajus Närvisüsteem on vanim, seda leidub koelenteraatides (hüdras). Sellist närvisüsteemi iseloomustab naaberelementide vaheliste ühenduste paljusus, mis võimaldab ergastusel vabalt levida närvivõrgus igas suunas.
Seda tüüpi närvisüsteem tagab laialdase vahetatavuse ja seeläbi suurema töökindluse, kuid need reaktsioonid on ebatäpsed ja ebamäärased.
Sõlm närvisüsteemi tüüp on tüüpiline ussidele, molluskitele ja vähilaadsetele.
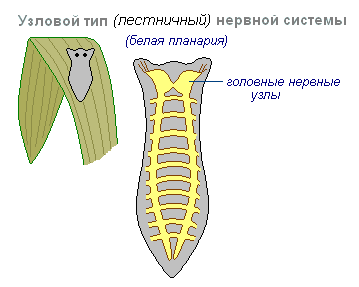
Seda iseloomustab asjaolu, et närvirakkude ühendused on teatud viisil organiseeritud, erutus kulgeb mööda rangelt määratletud radu. Selline närvisüsteemi korraldus osutub haavatavamaks. Ühe sõlme kahjustus põhjustab kogu organismi kui terviku talitlushäireid, kuid selle omadused on kiiremad ja täpsemad.
Torukujuline Närvisüsteem on iseloomulik akordidele, see sisaldab hajusate ja sõlmeliste tüüpide tunnuseid. Kõrgemate loomade närvisüsteem võttis kõik parima: difuusse tüübi kõrge usaldusväärsus, täpsus, lokaalsus, sõlmetüüpi reaktsioonide organiseerimise kiirus.
Närvisüsteemi juhtiv roll
Elusolendite maailma arengu esimeses etapis viidi kõige lihtsamate organismide vaheline interaktsioon läbi ürgse ookeani veekeskkonna, kuhu sisenesid nendest vabanevad keemilised ained. Esimene vanim hulkrakse organismi rakkude vahelise interaktsiooni vorm on keemiline interaktsioon ainevahetusproduktide kaudu, mis sisenevad kehavedelikesse. Sellised ainevahetusproduktid ehk metaboliidid on valkude, süsihappegaasi jne lagunemissaadused. See on mõjude humoraalne edasikandumine, humoraalne korrelatsioonimehhanism või elunditevahelised seosed.
Humoraalset seost iseloomustavad järgmised omadused:
- täpse aadressi puudumine, kuhu verre või muudesse kehavedelikesse sattunud keemiline aine saadetakse;
- kemikaal levib aeglaselt;
- kemikaal toimib väikestes kogustes ja tavaliselt laguneb või eemaldatakse organismist kiiresti.
Humoraalsed seosed on ühised nii looma- kui ka taimemaailmas. Loomamaailma teatud arenguetapis, seoses närvisüsteemi ilmnemisega, kujuneb välja uus, närviline seoste ja regulatsiooni vorm, mis kvalitatiivselt eristab loomamaailma taimemaailmast. Mida kõrgem on looma organismi areng, seda suuremat rolli mängib organite koosmõju närvisüsteemi kaudu, mida nimetatakse refleksiks. Kõrgemates elusorganismides reguleerib närvisüsteem humoraalseid seoseid. Erinevalt humoraalsest ühendusest on närviühendusel täpne suund konkreetsele elundile ja isegi rakkude rühmale; side toimub sadu kordi kiiremini kui kemikaalide leviku kiirus. Humoraalsest ühendusest närviühendusele üleminekuga ei kaasnenud mitte keharakkudevahelise humoraalse sideme hävimine, vaid närvisidemete allutamine ja neurohumoraalsete seoste tekkimine.
Elusolendite arengu järgmisel etapil ilmuvad spetsiaalsed elundid - näärmed, milles toodetakse hormoone, mis moodustuvad kehasse sisenevatest toiduainetest. Närvisüsteemi põhiülesanne on nii üksikute elundite aktiivsuse reguleerimine omavahel kui ka keha kui terviku koostoimes väliskeskkonnaga. Igasugune väliskeskkonna mõju kehale avaldub ennekõike retseptoritele (sensoorsetele organitele) ja toimub väliskeskkonna ja närvisüsteemi poolt põhjustatud muutuste kaudu. Närvisüsteemi arenedes saab selle kõrgeimast osakonnast – ajupoolkeradest – „kogu keha tegevuste juht ja jagaja”.
Närvisüsteemi struktuur
Närvisüsteemi moodustab närvikude, mis koosneb tohutust kogusest neuronid- protsessidega närvirakk.

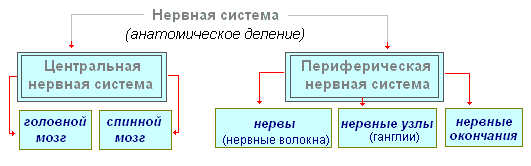
Tavapäraselt jaguneb närvisüsteem kesk- ja perifeerseks.

kesknärvisüsteem hõlmab pea- ja seljaaju ning perifeerne närvisüsteem- nendest ulatuvad närvid.
Aju ja seljaaju on neuronite kogum. Aju ristlõikes eristatakse valget ja halli ainet. Hallaine koosneb närvirakkudest ja valgeaine koosneb närvikiududest, mis on närvirakkude protsessid. Kesknärvisüsteemi erinevates osades on valge ja halli aine paiknemine erinev. Seljaajus paikneb hallollus sees ja valgeaine on väljas, ajus (ajupoolkerad, väikeaju) aga hallollus on väljas, valge aine sees. Aju erinevates osades on valgeaine sees eraldi närvirakkude (halli aine) klastrid - tuumad. Närvirakkude klastrid asuvad ka väljaspool kesknärvisüsteemi. Neid kutsutakse sõlmed ja kuuluvad perifeersesse närvisüsteemi.
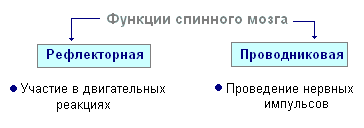
Närvisüsteemi refleksne aktiivsus
Närvisüsteemi põhitegevuse vorm on refleks. Refleks- keha reaktsioon sise- või väliskeskkonna muutustele, mis viiakse läbi kesknärvisüsteemi osalusel vastusena retseptorite ärritusele.
Mis tahes ärrituse korral kandub erutus retseptoritest mööda tsentripetaalseid närvikiude kesknärvisüsteemi, kust see läbi interneuroni piki tsentrifugaalkiude suundub perifeeriasse ühte või teise elundisse, mille aktiivsus muutub. Kogu seda teed läbi kesknärvisüsteemi tööorganini nimetatakse refleksi kaar Tavaliselt moodustuvad kolm neuronit: sensoorne, interkalaarne ja motoorne. Refleks on keeruline toiming, milles osaleb oluliselt suurem arv neuroneid. Ergastus, sattudes kesknärvisüsteemi, levib paljudesse seljaaju osadesse ja jõuab ajju. Paljude neuronite koosmõju tulemusena reageerib organism ärritusele.
Selgroog
Selgroog- umbes 45 cm pikkune, 1 cm läbimõõduga nöör, mis asub seljaaju kanalis, kaetud kolme ajukelmega: kõvakesta, ämblikuvõrkkest ja pehme (vaskulaarne).

Selgroog asub lülisambakanalis ja kujutab endast nööri, mis ülaosas läheb medulla piklikusse ja alumises otsas teise nimmelüli tasemel. Seljaaju koosneb hallainest, mis sisaldab närvirakke, ja valgeainest, mis koosneb närvikiududest. Hallollus paikneb seljaaju sees ja on igast küljest ümbritsetud valge ainega.
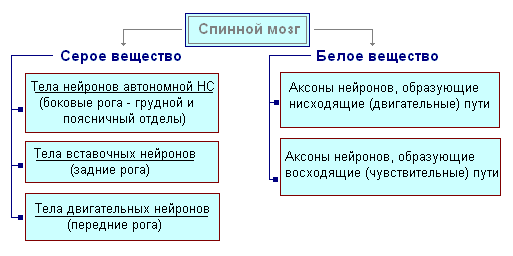
Läbilõikes meenutab hallollus tähte H. See eristab eesmist ja tagumist sarve, samuti ühendavat ristlatti, mille keskel on seljaaju kitsas kanal, mis sisaldab tserebrospinaalvedelikku. Rindkere piirkonnas on külgmised sarved. Need sisaldavad siseorganeid innerveerivate neuronite kehasid. Seljaaju valge aine moodustub närviprotsessides. Lühikesed protsessid ühendavad seljaaju osi ja pikad moodustavad ajuga kahepoolsete ühenduste juhtiva aparatuuri.

Seljaajus on kaks paksenemist – emakakaela- ja nimmeosa, millest närvid ulatuvad ülemistele ja alajäsemetele. Seljaajust tekib 31 paari seljaajunärve. Iga närv algab seljaajust kahe juurega - eesmine ja tagumine. Tagumised juured - tundlik koosnevad tsentripetaalsete neuronite protsessidest. Nende kehad asuvad seljaaju ganglionides. eesmised juured - mootor- on seljaaju hallaines paiknevate tsentrifugaalneuronite protsessid. Eesmise ja tagumise juurte ühinemise tulemusena moodustub segatud seljaaju närv. Seljaaju sisaldab keskusi, mis reguleerivad lihtsamaid refleksi toiminguid. Seljaaju peamised funktsioonid on refleksi aktiivsus ja erutuse juhtimine.
Inimese seljaajus on refleksikeskused üla- ja alajäsemete lihaste, higistamise ja urineerimise jaoks. Ergastuse funktsioon seisneb selles, et ajust kõikidesse kehapiirkondadesse ja tagasi suunduvad impulsid läbivad seljaaju. Tsentrifugaalimpulsid elunditest (nahk, lihased) edastatakse tõusvate radade kaudu ajju. Mööda laskuvaid radu kanduvad tsentrifugaalimpulsid ajust seljaajusse, sealt perifeeriasse, organitesse. Kui teed on kahjustatud, kaob tundlikkus erinevates kehaosades, rikutakse vabatahtlikke lihaste kontraktsioone ja liikumisvõimet.
Selgroogsete aju evolutsioon
Kesknärvisüsteemi moodustumine neuraaltoru kujul ilmneb esmalt akordidena. U madalamad akordid neuraaltoru püsib kogu elu, kõrgemale- selgroogsed - embrüonaalses staadiumis moodustub seljapoolele närviplaat, mis vajub naha alla ja voldib toruks. Embrüonaalses arengustaadiumis moodustub neuraaltoru eesmises osas kolm turset - kolm ajuvesiikulit, millest arenevad ajuosad: eesmine vesiikul annab eesaju ja vaheaju, keskmine vesiikul muutub keskajuks, tagumine vesiikul moodustab väikeaju ja pikliku medulla. Need viis ajupiirkonda on iseloomulikud kõigile selgroogsetele.
Sest madalamad selgroogsed- kalad ja kahepaiksed – mida iseloomustab keskaju ülekaal teistest osadest. U kahepaiksed Eesaju suureneb mõnevõrra ja poolkerade katusesse moodustub õhuke närvirakkude kiht - esmane medullaarne võlv, iidne ajukoor. U roomajad Närvirakkude kuhjumise tõttu suureneb eesaju oluliselt. Suurema osa poolkerade katusest on hõivatud iidse ajukoorega. Esimest korda ilmub roomajatel uue ajukoore alge. Eesaju poolkerad hiilivad teistele osadele, mille tagajärjel tekib vahekeha piirkonnas painutus. Alates iidsetest roomajatest said ajupoolkerad aju suurimaks osaks.
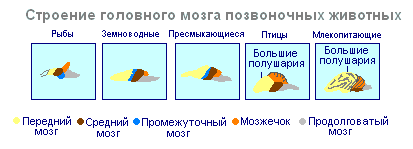
Aju ehituses linnud ja roomajad palju ühist. Aju katusel asub esmane ajukoor, keskaju on hästi arenenud. Lindudel aga võrreldes roomajatega suureneb aju kogumass ja eesaju suhteline suurus. Väikeaju on suur ja volditud struktuuriga. U imetajad eesaju saavutab oma suurima suuruse ja keerukuse. Suurem osa ajuainest koosneb neokorteksist, mis toimib kõrgema närvitegevuse keskusena. Imetajate aju vahe- ja keskosa on väikesed. Laienevad eesaju poolkerad katavad neid ja purustavad enda alla. Mõnedel imetajatel on sile aju ilma soonte ja keerdudeta, kuid enamikul imetajatel on ajukoores vaod ja keerdud. Vagude ja keerdude ilmumine on tingitud aju kasvust, mille kolju mõõtmed on piiratud. Ajukoore edasine kasv toob kaasa voltimise ilmnemise soonte ja keerdude kujul.
Aju
Kui kõigi selgroogsete seljaaju on enam-vähem võrdselt arenenud, siis aju erineb erinevatel loomadel märkimisväärselt suuruse ja ehituse keerukuse poolest. Evolutsiooni käigus toimuvad eriti dramaatilised muutused eesajus. Madalamatel selgroogsetel on eesaju halvasti arenenud. Kaladel esindavad seda haistmissagarad ja halli aine tuumad aju paksuses. Eesaju intensiivne areng on seotud loomade maale ilmumisega. See eristub vaheseinaks ja kaheks sümmeetriliseks poolkeraks, mida nimetatakse telentsefalon. Hallollus eesaju (koore) pinnal ilmub esmakordselt roomajatel, arenedes edasi lindudel ja eriti imetajatel. Tõeliselt suured eesaju poolkerad muutuvad ainult lindudel ja imetajatel. Viimases katavad need peaaegu kõik muud ajuosad.
Aju asub koljuõõnes. See hõlmab ajutüve ja telentsefaloni (ajukoor).

Ajutüvi koosneb piklikust medullast, sillast, keskajust ja vaheajust.
Medulla on seljaaju otsene jätk ja laienedes läheb tagaajusse. Põhimõtteliselt säilitab see seljaaju kuju ja struktuuri. Medulla oblongata paksuses on halli aine kogunemine - kraniaalnärvide tuumad. Tagatelg sisaldab väikeaju ja sild. Väikeaju paikneb pikliku medulla kohal ja on keerulise ehitusega. Väikeaju poolkerade pinnal moodustab hallaine ajukoore ja väikeaju sees selle tuumad. Nagu seljaaju piklik medulla, täidab see kahte funktsiooni: refleks ja juhtiv. Pikkmedulla refleksid on aga keerulisemad. See väljendub selle tähtsuses südametegevuse, veresoonte seisundi, hingamise ja higistamise reguleerimisel. Kõigi nende funktsioonide keskused asuvad medulla piklikus. Siin on närimise, imemise, neelamise, sülje ja maomahla keskused. Hoolimata oma väikesest suurusest (2,5–3 cm), on piklik medulla kesknärvisüsteemi oluline osa. Selle kahjustus võib hingamise ja südametegevuse seiskumise tõttu põhjustada surma. Medulla oblongata ja silla juhifunktsioon on impulsside edastamine seljaajust ajju ja tagasi.
IN keskaju paiknevad primaarsed (subkortikaalsed) nägemis- ja kuulmiskeskused, mis viivad läbi reflektoorseid orienteerumisreaktsioone valgus- ja helistimulatsioonile. Need reaktsioonid väljenduvad torso, pea ja silmade erinevates liigutustes stiimulite suunas. Keskaju koosneb ajuvarredest ja nelipeaajudest. Keskaju reguleerib ja jaotab skeletilihaste toonust (pinget).
Diencephalon koosneb kahest osakonnast - talamus ja hüpotalamus, millest igaüks koosneb suurest hulgast visuaalse talamuse ja subtalamuse piirkonna tuumadest. Visuaalse talamuse kaudu edastatakse tsentripetaalsed impulsid ajukoorele kõigist keha retseptoritest. Ükski tsentripetaalne impulss, olenemata sellest, kust see pärineb, ei saa visuaalsetest künkadest mööda minna ajukooresse. Seega suhtlevad kõik retseptorid vahekeha kaudu ajukoorega. Subtuberkulaarses piirkonnas on keskused, mis mõjutavad ainevahetust, termoregulatsiooni ja endokriinseid näärmeid.
Väikeaju asub pikliku medulla taga. See koosneb hallist ja valgest ainest. Erinevalt seljaajust ja ajutüvest paikneb aga hallaine – ajukoor – väikeaju pinnal ja valgeaine sees, ajukoore all. Väikeaju koordineerib liigutusi, muudab need selgeks ja sujuvaks, mängib olulist rolli keha tasakaalu hoidmisel ruumis ning mõjutab ka lihastoonust. Väikeaju kahjustumisel tekib inimesel lihastoonuse langus, liigutushäired ja kõnnakumuutused, kõne aeglustub jne. Mõne aja möödudes aga taastub liikumine ja lihaste toonus tänu sellele, et kesknärvisüsteemi puutumatud osad võtavad üle väikeaju funktsioonid.
Suured poolkerad- aju suurim ja arenenum osa. Inimestel moodustavad nad suurema osa ajust ja on kogu pinna ulatuses kaetud ajukoorega. Hallollus katab poolkerad väljastpoolt ja moodustab ajukoore. Inimese ajukoore paksus on 2–4 mm ja see koosneb 6–8 kihist, mis on moodustatud 14–16 miljardist erineva kuju, suuruse ja funktsioonide poolest. Ajukoore all on valge aine. See koosneb närvikiududest, mis ühendavad ajukoort kesknärvisüsteemi alumiste osadega ja poolkerade üksikuid sagaraid omavahel.
Ajukoores on soontega eraldatud keerdud, mis suurendavad oluliselt selle pinda. Kolm sügavaimat soont jagavad poolkerad labadeks. Igal poolkeral on neli sagarat: frontaalne, parietaalne, ajaline, kuklaluu. Erinevate retseptorite ergastamine siseneb ajukoore vastavatesse tajupiirkondadesse, nn tsoonid, ja siit kanduvad need edasi konkreetsesse elundisse, ajendades seda tegutsema. Korteksis eristatakse järgmisi tsoone. Kuulmistsoon asub oimusagaras, saab impulsse kuulmisretseptoritelt.

Visuaalne ala asub kuklaluu piirkonnas. Siia saabuvad impulsid silma retseptoritelt.
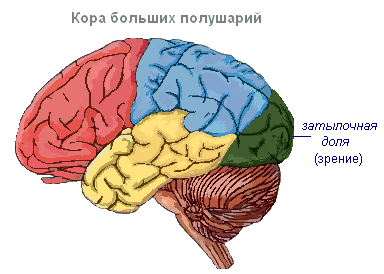
Haistmisvöönd paikneb oimusagara sisepinnal ja on seotud ninaõõnes olevate retseptoritega.
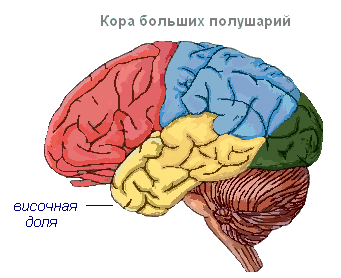
Sensoorne mootor tsoon asub eesmises ja parietaalsagaras. See tsoon sisaldab jalgade, torso, käte, kaela, keele ja huulte peamisi liikumiskeskusi. Siin asub ka kõne keskpunkt.
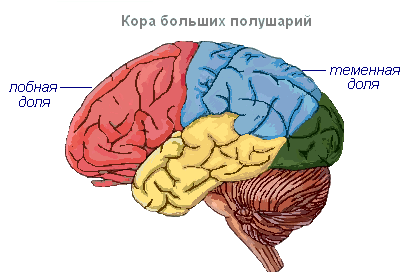
Ajupoolkerad on kesknärvisüsteemi kõrgeim osakond, mis kontrollib imetajate kõigi elundite tööd. Ajupoolkerade tähtsus inimestel seisneb ka selles, et need kujutavad endast vaimse tegevuse materiaalset alust. I. P. Pavlov näitas, et vaimne tegevus põhineb ajukoores toimuvatel füsioloogilistel protsessidel. Mõtlemine on seotud kogu ajukoore tegevusega, mitte ainult selle üksikute piirkondade funktsioonidega.
| Aju osakond | Funktsioonid | |
| Medulla | Dirigent | Ühendus seljaaju ja aju katvate osade vahel. |
| Refleks | Hingamisteede, südame-veresoonkonna, seedesüsteemi aktiivsuse reguleerimine:
|
|
| Pons | Dirigent | Ühendab väikeaju poolkerad omavahel ja ajukoorega. |
| Väikeaju | Koordineerimine | Tahtlike liigutuste koordineerimine ja kehaasendi hoidmine ruumis. Lihaste toonuse ja tasakaalu reguleerimine |
| Keskaju | Dirigent | Ligikaudsed refleksid visuaalsetele ja helistiimulitele ( pöörab pead ja keha). |
| Refleks |
|
|
| Diencephalon | talamus
hüpotalamus
|
|
Ajukoor
Pind ajukoor inimestel on see umbes 1500 cm 2, mis on mitu korda suurem kui kolju sisepind. See suur ajukoore pind tekkis tänu suure hulga soonte ja keerdude kujunemisele, mille tulemusena on suurem osa ajukoorest (umbes 70%) koondunud soontesse. Ajupoolkerade suurimad sooned on keskne, mis kulgeb üle mõlema poolkera ja ajaline, eraldades oimusagara ülejäänud osast. Ajukoorel on vaatamata väikesele paksusele (1,5–3 mm) väga keeruline struktuur. Sellel on kuus peamist kihti, mis erinevad neuronite ja ühenduste struktuuri, kuju ja suuruse poolest. Ajukoores on kõigi sensoorsete (retseptorite) süsteemide keskused, kõigi organite ja kehaosade esindajad. Sellega seoses lähenevad kõigist siseorganitest või kehaosadest pärit tsentripetaalsed närviimpulssid ajukoorele ja see suudab kontrollida nende tööd. Ajukoore kaudu suletakse konditsioneeritud refleksid, mille kaudu keha pidevalt, kogu elu jooksul, kohandub väga täpselt muutuvate eksistentsitingimustega, keskkonnaga.
Närvisüsteemi elusorganismis esindab sidevõrk, mis tagab selle ühenduse välismaailmaga ja oma protsessidega. Selle põhielemendiks on neuron – rakk, mille protsessid (aksonid ja dendriidid) edastavad informatsiooni elektriliselt ja keemiliselt.
Närviregulatsiooni eesmärk
Esimest korda ilmnes närvisüsteem elusorganismides, kuna oli vaja tõhusamat suhtlust keskkonnaga. Lihtsa impulsside edastamise võrgu väljatöötamine aitas mitte ainult väljastpoolt tulevaid signaale tajuda. Tänu sellele sai võimalikuks korraldada oma eluprotsesse edukamaks toimimiseks.
Evolutsiooni käigus muutus närvisüsteemi struktuur keerulisemaks: selle ülesandeks ei saanud mitte ainult välismõjudele adekvaatse vastuse kujundamine, vaid ka oma käitumise korraldamine. I. P. Pavlov nimetas seda toimimismeetodit
Koostoime üherakulise keskkonnaga
Närvisüsteem tekkis esmakordselt rohkem kui ühest rakust koosnevates organismides, kuna see edastab signaale võrgustikku moodustavate neuronite vahel. Kuid juba algloomadel võib täheldada võimet reageerida rakusiseste protsesside poolt põhjustatud välistele stiimulitele.
Mitmerakuliste organismide närvisüsteem erineb kvalitatiivselt algloomade sarnasest moodustisest. Viimased leiavad kogu ühenduste süsteemi ühe raku ainevahetuses. Ripsloomad “õpivad” erinevatest protsessidest, mis toimuvad väliselt või sisemiselt protoplasma koostise muutumise ja mõne muu struktuuri tegevuse tõttu. Mitmerakulistel elusolenditel on süsteem, mis on üles ehitatud funktsionaalsetest üksustest, millest igaühel on oma metaboolsed protsessid.
Seega ilmub esimest korda närvisüsteem inimesel, kellel pole mitte üks, vaid mitu rakku, see tähendab, et prototüübiks on impulsside juhtimine algloomades. Nende elutähtsa aktiivsuse tasemel toodab protoplasma struktuure, mis juhivad impulsse. Samamoodi täidavad keerulisemates elusolendites seda funktsiooni üksikisik
Koelenteraatide närvisüsteemi tunnused
Kolooniates elavad hulkraksed loomad ei jaga omavahel funktsioone ning neil puudub veel närvivõrgustik. See esineb staadiumis, mil mitmerakulise organismi mitmesugused funktsioonid eristuvad.

Esimest korda ilmub närvisüsteem hüdras ja teistes koelenteraatides. See on võrk, mis edastab mittesihitud signaale. Struktuur pole veel moodustunud, see on hajutatud kogu koelenteraadi kehas. Ganglionrakud ja nende Nisslevi aine ei ole täielikult moodustunud. See on närvisüsteemi kõige lihtsam versioon.
Loomade motoorse aktiivsuse tüübi määrab hajus võrgutaoline närvisüsteem. Hydra teeb peristaltilisi liigutusi, kuna sellel pole liikumiseks ja muudeks liigutusteks spetsiaalseid kehaosi. Motoorse aktiivsuse jaoks nõuab see kontraktiilsete elementide pidevat ühendamist ja see eeldab, et suurem osa juhtivatest rakkudest asuks kontraktiilses osas. Millise looma närvisüsteem ilmub esmakordselt hajusa võrgustikuna? Need, kes on inimeste reguleerimissüsteemi rajajad. Selle tõestuseks on asjaolu, et loomade embrüote arengus esineb gastrulatsiooni.
Helmintide närvisüsteemi tunnused
Närviregulatsiooni hilisem paranemine oli seotud kahepoolse sümmeetria kujunemisega radiaalse asemel ja neuronite klastrite moodustumisega erinevates kehaosades.

Nööride kujul ilmneb närvisüsteem esmakordselt 1 Selles etapis esindavad seda paaritud peakiud ja neist välja ulatuvad moodustunud kiud. Võrreldes koelenteraatidega on see süsteem palju keerulisem. Helmintides leidub närvirakkude rühmi sõlmede ja ganglionide kujul. Aju prototüüp on keha esiosas paiknev ganglion, mis täidab regulatoorseid funktsioone. Seda nimetatakse aju ganglioniks. Sellest jooksevad mööda kogu keha kaks närvitüve, mis on ühendatud hüppajatega.

Kõik süsteemi komponendid ei asu väljaspool, vaid on sukeldatud parenhüümi ja on seeläbi kaitstud vigastuste eest. Esimest korda ilmub lameussidel närvisüsteem koos kõige lihtsamate meeleorganitega: puudutus, nägemine ja tasakaalutunne.
Nematoodide närvisüsteemi tunnused
Järgmine arenguetapp on neelu lähedusse rõnga moodustumine ja mitmed sellest ulatuvad pikad kiud. Selliste omadustega närvisüsteem ilmub kõigepealt perifarüngeaalsesse rõngasse, mis on üks ümmargune ganglion ja täidab tajumise põhiorgani funktsioone. Sellega on ühendatud kõhunöör ja seljanärv.
Nematoodide närvitüved paiknevad intraepiteliaalselt, see tähendab hüpodermaalsetes harjades. Tajuorganite rolli täidavad sensillad - setae, papillid, lisaelundid, amfiid ja phasmiidid. Kõik nad on varustatud segase tundlikkusega.

Nematoodide kõige keerulisemad tajuorganid on amfid. Need on paaris, võivad olla erineva kujuga ja asuvad ees. Nende peamine ülesanne on ära tunda kehast kaugel asuvad keemilised mõjurid. Mõnedel ümarussidel on ka retseptorid, mis tajuvad sisemisi ja väliseid mehaanilisi mõjusid. Neid nimetatakse metaneemideks.
Ringleti närvisüsteemi tunnused
Närvisüsteemi ganglionide moodustumine areneb edasi anneliidides. Enamikus neist toimub kõhutüvede ganglionisatsioon nii, et igas ussi segmendis on paar närvisõlme, mis on kiudude kaudu ühendatud naabersegmentidega. neil on medullaarsest ganglionist moodustatud ventraalne närvijuhe ja sellest lähtuv nööripaar. Nad venivad piki kõhu tasapinda. Tajuvad elemendid asuvad ees ja neid esindavad kõige lihtsamad silmad, haistmisrakud, tsiliaarsed lohud ja lokaatorid. Paaritud sõlmedega tekkis närvisüsteem esmalt anneliididel, kuid hiljem areneb see välja lülijalgsetel. Neil on peas olevate ganglionide laienemine ja kehas olevate sõlmede kombinatsioon.
Inimese närvisüsteemi hajusvõrgu elemendid
Närvisüsteemi evolutsioonilise arengu tipp on inimese pea- ja seljaaju ilmumine. Kuid isegi selliste keerukate struktuuride olemasolul säilib algne hajus korraldus. See võrk mässib kõiki keharakke: nahka, veresooni jne. Kuid selliste omadustega ilmneb esimest korda närvisüsteem inimesel, kellel polnud isegi võimalust keskkonda erinevalt tajuda.

Tänu nendele “jääk” struktuuriüksustele on inimesel võimalus tunda erinevaid mõjutusi ka mikroskoopilistel aladel. Keha võib reageerida väikseima võõrkeha ilmumisele kaitsereaktsioonide arendamisega. Hajusvõrgu olemasolu inimese närvisüsteemis kinnitavad laboratoorsed uurimismeetodid, mis põhinevad värvaine sisseviimisel.
Närvisüsteemi üldine arengujoon evolutsiooni ajal
Närvisüsteemi evolutsioonilised protsessid toimusid kolmes etapis:
- hajus võrk;
- gangiiliad;
- seljaaju ja aju.

Kesknärvisüsteemi struktuur ja toimimine on varasematest tüüpidest väga erinev. Selle sümpaatiline jaotus sisaldab ganglion- ja retikulaarseid elemente. Närvisüsteem oma fülogeneetilises arengus fragmenteeris ja diferentseerus üha enam. Ganglioniline arengustaadium erines retikulaarsest staadiumist neuronite olemasolul, mis paiknesid endiselt juhtivussüsteemi kohal.
Iga elusorganism on oma olemuselt monoliit, mis koosneb erinevatest organitest ja nende süsteemidest, mis pidevalt ja pidevalt suhtlevad üksteisega ja väliskeskkonnaga. Närvisüsteem tekkis esmakordselt koelenteraatides, see oli hajus võrgustik, mis tagas impulsside elementaarse juhtivuse.
LOENG TEEMAL: INIMESE NÄRVISÜSTEEM
Närvisüsteem on süsteem, mis reguleerib inimese kõigi organite ja süsteemide tegevust. See süsteem määrab: 1) kõigi inimorganite ja süsteemide funktsionaalse ühtsuse; 2) kogu organismi seotus keskkonnaga.
Homöostaasi säilitamise seisukohalt tagab närvisüsteem: sisekeskkonna parameetrite hoidmise etteantud tasemel; käitumuslike reaktsioonide kaasamine; kohanemine uute tingimustega, kui need püsivad pikka aega.
Neuron(närvirakk) - närvisüsteemi peamine struktuurne ja funktsionaalne element; Inimestel on rohkem kui sada miljardit neuronit. Neuron koosneb kehast ja protsessidest, tavaliselt ühest pikast protsessist – aksonist ja mitmest lühikesest hargnenud protsessist – dendriitidest. Mööda dendriite järgnevad impulsid rakukehale, mööda aksonit - raku kehast teistele neuronitele, lihastele või näärmetele. Tänu protsessidele võtavad neuronid omavahel kontakti ja moodustavad närvivõrgustikke ja ringe, mille kaudu ringlevad närviimpulsid.
Neuron on närvisüsteemi funktsionaalne üksus. Neuronid on vastuvõtlikud stimulatsioonile, see tähendab, et nad on võimelised ergastama ja edastama elektrilisi impulsse retseptoritelt efektoritele. Impulsi ülekande suuna alusel eristatakse aferentseid neuroneid (sensoorsed neuronid), efferentseid neuroneid (motoorsed neuronid) ja interneuroneid.
Närvikudet nimetatakse erutuvaks koeks. Vastuseks mõnele löögile tekib ja levib selles ergastusprotsess - rakumembraanide kiire laadimine. Ergastuse (närviimpulsi) tekkimine ja levik on peamine viis, kuidas närvisüsteem täidab oma kontrollifunktsiooni.
Peamised eeldused ergastuse tekkeks rakkudes: elektrilise signaali olemasolu membraanil puhkeolekus - puhkemembraani potentsiaal (RMP);
võime muuta potentsiaali, muutes membraani läbilaskvust teatud ioonide jaoks.
Rakumembraan on poolläbilaskev bioloogiline membraan, sellel on kanalid, mis lasevad läbi kaaliumiioone, kuid puuduvad kanalid rakusiseste anioonide jaoks, mis jäävad kinni membraani sisepinnale, tekitades membraani negatiivse laengu. sees on membraani puhkepotentsiaal, mis on keskmiselt - – 70 millivolti (mV). Rakus on 20-50 korda rohkem kaaliumiioone kui väljas, see säilib kogu elu jooksul membraanpumpade abil (suured valgumolekulid, mis on võimelised kaaliumiioone rakuvälisest keskkonnast sisemusse transportima). MPP väärtus määratakse kaaliumiioonide ülekandega kahes suunas:
1. väljastpoolt rakku pumpade toimel (suure energiakuluga);
2. rakust väljapoole difusiooni teel membraanikanalite kaudu (ilma energiatarbimiseta).
Ergutamise protsessis mängivad peamist rolli naatriumioonid, mida on väljaspool rakku alati 8-10 korda rohkem kui sees. Naatriumikanalid on raku puhkeolekus suletud, nende avamiseks on vaja rakule adekvaatse stiimuliga mõjuda. Kui stimulatsioonilävi saavutatakse, avanevad naatriumikanalid ja naatrium siseneb rakku. Tuhandiksekundi jooksul kaob membraanilaeng esmalt ja muutub seejärel vastupidiseks – see on aktsioonipotentsiaali (AP) esimene faas – depolarisatsioon. Kanalid sulguvad - kõvera tipp, seejärel taastatakse laeng mõlemal pool membraani (kaaliumkanalite tõttu) - repolarisatsiooni staadium. Ergastus peatub ja raku puhkeolekus vahetavad pumbad rakku sisenenud naatriumi kaaliumi vastu, mis rakust lahkus.

Närvikiu suvalises punktis esile kutsutud PD muutub membraani naaberosadele ärritavaks, põhjustades neis AP-d, mis omakorda ergastavad üha rohkem membraani sektsioone, levides seega kogu rakus. Müeliiniga kaetud kiududes tekivad AP-d ainult müeliinivabades piirkondades. Seetõttu suureneb signaali levimise kiirus.


Ergastuse ülekandmine rakust teise toimub keemilise sünapsi kaudu, mida esindab kahe raku kokkupuutepunkt. Sünapsi moodustavad presünaptilised ja postsünaptilised membraanid ning nendevaheline sünaptiline lõhe. AP-st tulenev erutus rakus jõuab presünaptilise membraani piirkonda, kus paiknevad sünaptilised vesiikulid, millest eraldub spetsiaalne aine, saatja. Vahesse sisenev saatja liigub postsünaptilisele membraanile ja seondub sellega. Ioonide jaoks avanevad membraanis poorid, need liiguvad rakku ja toimub ergastusprotsess

Seega muundatakse rakus elektriline signaal keemiliseks ja keemiline signaal jällegi elektriliseks. Signaali ülekanne sünapsis toimub aeglasemalt kui närvirakus ja on ka ühepoolne, kuna saatja vabaneb ainult läbi presünaptilise membraani ja saab seostuda ainult postsünaptilise membraani retseptoritega ja mitte vastupidi.
Vahendajad võivad rakkudes põhjustada mitte ainult erutust, vaid ka inhibeerimist. Sel juhul avanevad membraanil poorid ioonide jaoks, mis tugevdavad membraanil puhkeolekus eksisteerinud negatiivset laengut. Ühel rakul võib olla palju sünaptilisi kontakte. Neuroni ja skeletilihaskiu vahelise vahendaja näide on atsetüülkoliin.
Närvisüsteem jaguneb kesknärvisüsteem ja perifeerne närvisüsteem.
Kesknärvisüsteemis eristatakse aju, kuhu on koondunud peamised närvikeskused ja seljaaju ning siin on madalama taseme keskused ja rajad perifeersetesse organitesse.
Perifeerne sektsioon - närvid, närviganglionid, ganglionid ja põimikud.
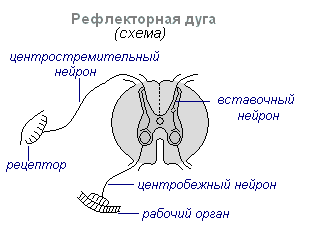
Närvisüsteemi peamine toimemehhanism on refleks. Refleks on keha igasugune reaktsioon välis- või sisekeskkonna muutustele, mis viiakse läbi kesknärvisüsteemi osalusel vastusena retseptorite ärritusele. Refleksi struktuurne alus on refleksi kaar. See sisaldab viit järjestikust linki:
1 - retseptor - signaalseade, mis tajub mõju;
2 - Aferentne neuron – toob signaali retseptorilt närvikeskusesse;
3 - Interneuron – kaare keskosa;
4 - Efferent neuron - signaal tuleb kesknärvisüsteemist täidesaatvasse struktuuri;
5 – efektor – teatud tüüpi tegevust sooritav lihas või nääre

Aju koosneb närvirakkude kehade, närviteede ja veresoonte klastritest. Närvikanalid moodustavad aju valgeaine ja koosnevad närvikiudude kimpudest, mis juhivad impulsse aju halli aine erinevatesse osadesse - tuumadesse või keskustesse või sealt välja. Rajad ühendavad erinevaid tuumasid, aga ka pea- ja seljaaju.
Funktsionaalselt võib aju jagada mitmeks osaks: eesaju (koosneb telentsefalonist ja vaheajust), keskaju, tagaaju (koosneb väikeajust ja sillast) ja piklikaju. Medulla oblongata, silla ja keskaju nimetatakse ühiselt ajutüveks.

Selgroog asub seljaaju kanalis, kaitstes seda usaldusväärselt mehaaniliste kahjustuste eest.
Seljaaju on segmentaalse struktuuriga. Igast segmendist ulatub välja kaks paari eesmisi ja tagumisi juuri, mis vastab ühele selgroolülile. Kokku on 31 paari närve.
Seljajuured moodustavad sensoorsed (aferentsed) neuronid, nende kehad paiknevad ganglionides ja aksonid sisenevad seljaajusse.
Eesmised juured moodustuvad efferentsete (motoorsete) neuronite aksonitest, mille kehad asuvad seljaajus.
Seljaaju jaguneb tinglikult neljaks osaks - emakakaela-, rindkere-, nimme- ja sakraalne. See sulgeb tohutul hulgal reflekskaare, mis tagab paljude kehafunktsioonide reguleerimise.
Hall keskaine on närvirakud, valge on närvikiud.

Närvisüsteem jaguneb somaatiliseks ja autonoomseks.
TO somaatiline närvilisus süsteem (ladina sõnast “soma” - keha) viitab närvisüsteemi osale (nii rakukehad kui ka nende protsessid), mis kontrollib skeletilihaste (keha) ja meeleelundite tegevust. Seda närvisüsteemi osa juhib suuresti meie teadvus. See tähendab, et me suudame oma suva järgi painutada või sirutada kätt, jalga vms.. Samas ei suuda me teadlikult lõpetada näiteks helisignaalide tajumist.
Autonoomne närvilisus süsteem (tõlkes ladina keelest "vegetatiivne" - taim) on osa närvisüsteemist (nii rakukehad kui ka nende protsessid), mis juhib rakkude ainevahetuse, kasvu ja paljunemise protsesse, see tähendab nii loomade kui ka taimede organismide ühiseid funktsioone. . Autonoomne närvisüsteem vastutab näiteks siseorganite ja veresoonte tegevuse eest.
Autonoomset närvisüsteemi teadvus praktiliselt ei kontrolli, see tähendab, et me ei suuda oma suva järgi leevendada sapipõie spasme, peatada rakkude jagunemist, peatada sooletegevuse, laiendada ega ahendada veresooni.
Nagu teada, ilmub närvisüsteem esmakordselt madalamate hulkrakuliste selgrootute puhul. Närvisüsteemi tekkimine on suur verstapost loomamaailma evolutsioonis ja selle poolest erinevad isegi primitiivsed mitmerakulised selgrootud kvalitatiivselt algloomadest. Siin on oluline punkt ergastuse juhtivuse järsk kiirenemine närvikoes: uprotoplasmas ei ületa ergastuse juhtivuse kiirus 1-2 mikronit sekundis, kuid isegi kõige primitiivsemas närvisüsteemis, mis koosneb närvirakkudest, on see 0,5 meetrit sekundis!
Närvisüsteem eksisteerib madalamates hulkraksetes organismides väga erineval kujul: võrkjas (näiteks hüdras), rõngakujuline (meduus), radiaalne (täht) ja kahepoolne. Kahepoolset vormi esindavad madalamad (soolestiku) lameussid ja primitiivsed molluskid (kitoon) ainult kehapinna lähedal asuva võrgustikuga, kuid mitmed pikisuunalised nöörid eristuvad võimsama arengu poolest. Närvisüsteem vajub järk-järgult arenedes lihaskoe alla ja pikisuunalised nöörid muutuvad tugevamaks, eriti keha ventraalsel küljel. Samal ajal muutub üha olulisemaks keha eesmine ots, ilmub pea (tsefaliseerimise protsess) ja koos sellega aju - närvielementide kogunemine ja tihendamine eesmises otsas. Lõpuks omandab kesknärvisüsteem kõrgematel ussidel juba täielikult "närviredeli" tüüpilise struktuuri, milles aju asub seedetrakti kohal ja on ühendatud kahe sümmeetrilise kommissuuriga ("periofarüngeaalne ring") subfarüngeaalsete ganglionidega. paiknevad kõhupoolsel küljel ja seejärel paaris kõhu närvitüvedega. Olulised elemendid on siin ganglionid, mistõttu räägitakse ka ganglionärvisüsteemist ehk “ganglionaarsest trepist”. Mõnel selle loomarühma esindajal (näiteks kaanidel) koonduvad närvitüved nii tihedalt kokku, et tekib “närvikett”.
Võimsad juhtivad kiud väljuvad ganglionidest, mis moodustavad närvitüvesid. Hiiglaslikes kiududes viiakse närviimpulsid palju kiiremini läbi nende suure läbimõõdu ja väikese arvu sünaptiliste ühenduste (mõnede närvirakkude aksonite ning teiste rakkude dendriitide ja rakukehade kokkupuutekohad) tõttu. Mis puutub peaganglionidesse, st. aju, siis on nad rohkem arenenud aktiivsematel loomadel, kellel on ka kõige arenenumad retseptorisüsteemid.
Närvisüsteemi tekke ja evolutsiooni määrab vajadus koordineerida paljurakulise organismi erineva kvaliteediga funktsionaalüksusi, ühtlustada selle eri osades väliskeskkonnaga suheldes toimuvaid protsesse ning tagada kompleksse organismi aktiivsus nagu ühtne terviklik süsteem. Ainult koordineeriv ja organiseeriv keskus, näiteks kesknärvisüsteem, suudab mitmerakulises organisatsioonis pakkuda paindlikkust ja varieeruvust organismi reaktsioonis.
Suur tähtsus oli selles osas ka tsefalisaapia protsessil, s.o. organismi peaotsa eraldamine ja sellega seotud aju välimus. Ainult aju juuresolekul on võimalik perifeeriast tulevate signaalide tõeliselt tsentraliseeritud "kodeerimine" ja kaasasündinud käitumise terviklike "programmide" moodustamine, rääkimata looma kogu välistegevuse kõrgest koordineerimisest.
Loomulikult ei sõltu vaimse arengu tase ainult närvisüsteemi struktuurist. Näiteks anneliididega tihedalt seotud rotiferidel on sarnaselt neile kahepoolne närvisüsteem ja aju, samuti spetsiaalsed sensoorsed ja motoorsed närvid. Kuid erinedes ripslastest oma suuruse, välimuse ja elustiili poolest vähe, on rotiferid oma käitumiselt väga sarnased viimastele ega ilmuta kõrgemaid vaimseid võimeid kui ripslased. See näitab taas, et vaimse aktiivsuse arengu juhtivaks teguriks ei ole mitte üldine struktuur, vaid looma spetsiifilised elutingimused, tema suhete ja keskkonnaga suhtlemise iseloom. Samas näitab see näide veel kord, kui hoolikalt tuleb läheneda “kõrgemate” ja “madalamate” iseloomude hindamisele, kui võrrelda erineval filogeneetilisel positsioonil olevaid organisme, eriti kui võrrelda algloomi ja mitmerakulisi selgrootuid.
3.1. Närvisüsteemi päritolu ja funktsioonid.
Kõigi loomade närvisüsteem on ektodermaalset päritolu. See täidab järgmisi funktsioone:
Organismi suhtlemine keskkonnaga (ärrituse tajumine, edasikandumine ja reageerimine ärritusele);
Kõigi elundite ja organsüsteemide ühendamine ühtseks tervikuks;
Närvisüsteem on kõrgema närviaktiivsuse kujunemise aluseks.
3.2. Närvisüsteemi areng selgrootute loomade seas.
Närvisüsteem ilmus esmakordselt koelenterates ja oli difuusne või retikulaarne tüüp närvisüsteem, st. Närvisüsteem on närvirakkude võrgustik, mis on jaotatud üle kogu keha ja on omavahel ühendatud õhukeste protsessidega. Tüüpilise ehitusega on tal hüdras, kuid juba meduusidel ja polüüpidel tekivad teatud kohtades (suu lähedal, vihmavarju äärtes) närvirakkude kobarad, need närvirakkude kobarad on meeleelundite eelkäijad.
Edasi kulgeb närvisüsteemi areng närvirakkude koondumise teed teatud kehakohtadesse, s.o. mööda närvisõlmede (ganglionide) moodustumise teed. Need sõlmed tekivad peamiselt seal, kus asuvad rakud, mis tajuvad keskkonna ärritust. Seega tekib radiaalse sümmeetriaga närvisüsteemi radiaalne tüüp ja kahepoolse sümmeetriaga närviganglionide kontsentratsioon keha eesmises otsas. Pea sõlmedest ulatuvad mööda keha ulatuvad paarisnärvitüved. Seda tüüpi närvisüsteemi nimetatakse ganglionvarreks.
Seda tüüpi närvisüsteemil on lameussidele tüüpiline struktuur, st. keha eesmises otsas on paaritud ganglionid, millest edasi ulatuvad närvikiud ja meeleelundid ning mööda keha kulgevad närvitüved.
Ümarussidel ühinevad peaganglionid perifarüngeaalseks närvirõngaks, millest piki keha ulatuvad ka närvitüved.
Anneliidides moodustub närviahel, s.o. Igas segmendis moodustuvad sõltumatud paaris närvisõlmed. Kõik need on ühendatud nii piki- kui ka põiksuunaliste kiududega. Selle tulemusena omandab närvisüsteem redelilaadse struktuuri. Sageli lähenevad mõlemad ahelad üksteisele lähemale, ühendudes mööda keha keskosa paarituks kõhu närviahelaks.
Lülijalgsetel on sama tüüpi närvisüsteem, kuid närviganglionide arv väheneb ja nende suurus suureneb, eriti peas või tsefalotoraksis, s.o. tsefaliseerimise protsess on käimas.
Molluskitel esindavad närvisüsteemi erinevates kehaosades paiknevad sõlmed, mis on omavahel ühendatud sõlmedest välja ulatuvate nööride ja närvidega. Seedejalgsetel on pedaali-, aju- ja pleura-vistseraalsed sõlmed; kahepoolmelistel – pedaal ja pleura-vistseraalne; peajalgsetel - pleura-vistseraalsed ja ajunärvi ganglionid. Peajalgsete neelu ümber täheldatakse närvikoe kogunemist.
3.3. Närvisüsteemi evolutsioon akordides.
Akordide närvisüsteemi esindab neuraaltoru, mis eristub pea- ja seljaajuks.
Alumiste akordide korral on neuraaltoru õõnestoru (neurocoel) välimus, mille närvid ulatuvad torust välja. Lantsetis moodustub peaosas väike laienemine - aju rudiment. Seda laienemist nimetatakse vatsakesteks.
Kõrgemates akordides moodustuvad neuraaltoru eesmises otsas kolm turset: eesmised, keskmised ja tagumised vesiikulid. Esimesest ajupõieklist moodustuvad seejärel eesaju ja vahepeaaju, keskmisest ajupõiekesest - mesencephalon, tagumisest - väikeaju ja medulla oblongata, mis läheb seljaajusse.
Kõigis selgroogsete loomade klassides koosneb aju 5 sektsioonist (eesmine, keskmine, keskmine, tagumine ja medulla), kuid nende arenguaste ei ole erinevate klasside loomadel ühesugune.
Seega paiknevad tsüklostoomides kõik ajuosad üksteise järel horisontaaltasandil. Medulla oblongata läheb nutria keskkanaliga otse seljaajusse.
Kaladel on aju tsüklostoomidega võrreldes rohkem diferentseeritud. Eesaju maht on suurenenud, eriti kopsukaladel, kuid eesaju pole veel poolkeradeks jagatud ja toimib funktsionaalselt kõrgeima haistmiskeskusena. Eesaju katus on õhuke, koosneb ainult epiteelirakkudest ja ei sisalda närvikudet. Diencephalonis, millega on ühendatud käbikeha ja hüpofüüsi, asub hüpotalamus, mis on endokriinsüsteemi keskpunkt. Kaladel on kõige arenenum keskaju. Optilised labad on selles hästi väljendunud. Keskaju piirkonnas on kõikidele kõrgematele selgroogsetele iseloomulik painutus. Lisaks on keskaju analüüsikeskus. Tagaaju osaks olev väikeaju on kalade liikumise keerukuse tõttu hästi arenenud. See esindab liikumise koordinatsioonikeskust, selle suurus varieerub sõltuvalt erinevate kalaliikide liikumisaktiivsusest. Medulla oblongata tagab side aju kõrgemate osade ja seljaaju vahel ning sisaldab hingamis- ja vereringekeskusi.
Kala ajust väljub 10 paari kraniaalnärve.
Seda tüüpi aju, mille kõrgeim integratsioonikeskus on keskaju, nimetatakse ihtüopsiidiks.
Kahepaiksete närvisüsteem on oma struktuurilt lähedane kopsukalade närvisüsteemile, kuid seda iseloomustab märkimisväärne areng ja paaristatud piklike poolkerade täielik eraldumine, samuti väikeaju nõrk areng, mis on tingitud kahepaiksete vähesest liikuvusest. ja nende liigutuste monotoonsus. Kuid kahepaiksed arendasid eesaju jaoks katuse, mida nimetatakse esmaseks medullaarseks võlviks - arhipalliumiks. Kraniaalnärve on nagu kaladelgi kümme. Ja ajutüüp on sama, st. ihtüopsid.
Seega on kõigil anamnial (tsüklostoomidel, kaladel ja kahepaiksetel) ihtüopsiidi tüüpi aju.
Kõrgemate selgroogsete hulka kuuluvate roomajate aju ehituses, s.o. amnioni jaoks on progressiivse organisatsiooni tunnused selgelt väljendatud. Eesaju poolkeradel on märkimisväärne ülekaal teiste ajuosade ees. Nende aluses on suured närvirakkude akumulatsioonid - juttkeha. Vana ajukoore saared, arhikorteks, ilmuvad kummagi poolkera lateraalsele ja mediaalsele küljele. Keskaju suurus väheneb ja see kaotab oma tähtsuse juhtiva keskusena. Analüüsikeskuseks saab eesaju põhja, s.o. triibulised kehad. Seda tüüpi aju nimetatakse sauropsiidiks või striataaliks. Väikeaju on roomajate liikumise mitmekesisuse tõttu suurenenud. Medulla oblongata moodustab järsu painde, mis on iseloomulik kõigile amnionitele. Ajust lahkub 12 paari kraniaalnärve.
Sama tüüpi aju on omane lindudele, kuid teatud tunnustega. Eesaju poolkerad on suhteliselt suured. Lindude haistmissagarad on halvasti arenenud, mis viitab lõhna rollile lindude elus. Seevastu keskaju esindavad suured optilised sagarad. Väikeaju on hästi arenenud, ajust väljub 12 paari närve.
Imetajate aju saavutab maksimaalse arengu. Poolkerad on nii suured, et katavad keskaju ja väikeaju. Ajukoor on eriti arenenud, selle pindala on suurenenud keerdude ja soonte tõttu. Ajukoorel on väga keeruline struktuur ja seda nimetatakse uueks ajukooreks – neokorteksiks. Ilmub sekundaarne medullaarne võlv, neopallium. Poolkerade ees paiknevad suured haistmissagarad. Vahepea, nagu ka teised klassid, hõlmab käbinääret, hüpofüüsi ja hüpotalamust. Keskaju on suhteliselt väike, see koosneb neljast tuberklist - neljakesi. Eesmine ajukoor on ühendatud visuaalse analüsaatoriga, tagumine kuulmisanalüsaatoriga. Koos eesajuga areneb suurel määral ka väikeaju. Ajust lahkub 12 paari kraniaalnärve. Analüüsikeskus on ajukoor. Seda tüüpi aju nimetatakse rinnanäärmeks.
3.4. Närvisüsteemi anomaaliad ja väärarengud inimestel.
1. Atsefaalia- aju, võlvi, kolju ja näo skeleti puudumine; seda häiret seostatakse eesmise neuraaltoru alaarenguga ning see on kombineeritud seljaaju, luude ja siseorganite defektidega.
2. Anentsefaalia- ajupoolkerade ja kolju katuse puudumine koos ajutüve alaarenguga ja on kombineeritud muude arengudefektidega. Selle patoloogia põhjuseks on neuraaltoru pea mittesulgumine (düsrafism). Sel juhul kolju katuse luud ei arene ning koljupõhja luudel ilmnevad mitmesugused kõrvalekalded. Anentsefaalia ei sobi kokku eluga, keskmine esinemissagedus on 1/1500 ja esineb sagedamini naissoost loodetel.
3. Atelencefaalia- neuraaltoru eesmise osa arengu seiskumine (heterokroonia) kolme vesiikuli staadiumis. Selle tulemusena ei moodustu ajupoolkerad ja subkortikaalsed tuumad.
4. Prosentsefaalia– telentsefalon on jagatud pikisuunalise soonega, kuid sügavuti jäävad mõlemad poolkerad omavahel seotuks.
5. Holoprosentsefaalia- telentsefalon ei ole poolkeradeks jagatud ja sellel on ühe õõnsusega (vatsakesega) poolkera välimus.
6. Alobar prosentsefaalia- telentsefaloni jagunemine toimub ainult tagumises osas ja otsmikusagarad jäävad jagamata.
7. Aplaasia või kollakeha hüpoplaasia– aju kompleksse komissiooni täielik või osaline puudumine, s.t. corpus callosum.
8. Hüdroentsefaalia- ajupoolkerade atroofia kombinatsioonis vesipeaga.
9. Agiriya- ajupoolkerade soonte ja keerdude (sile aju) täielik puudumine.
10. Mikrogüüria- vagude arvu ja mahu vähendamine.
11. Kaasasündinud hüdrotsefaalia- aju vatsakeste süsteemi osa ja selle väljundite obstruktsioon, see on põhjustatud närvisüsteemi arengu esmasest häirest.
12. Spina bifida- seljaaju neuraaltoru sulgumise ja eraldumise defekt naha ektodermist. Mõnikord kaasneb selle anomaaliaga diplomüelia, mille korral seljaaju jaguneb teatud pikkuses kaheks osaks, millest igaühel on oma keskne süvend.
13. Inientsefaalia- haruldane, eluga kokkusobimatu anomaalia esineb sagedamini naissoost loodetel. See on pea ja aju tagaosa jäme anomaalia. Pea pööratakse nii, et nägu oleks suunatud ülespoole. Dorsaalselt jätkub peanahk lumbodorsaalse või ristluu piirkonna nahka.
 Graham Greene'i lühike elulugu
Graham Greene'i lühike elulugu Scipio Aafrika vanem Rooma kindral Scipio
Scipio Aafrika vanem Rooma kindral Scipio Näiteid rahvakeelest vene keeles
Näiteid rahvakeelest vene keeles