Peptidna veza. Stvaranje peptidne veze Dva ekvivalentna oblika
Elongacija, stvaranje peptidne veze (reakcija transpeptidacije). Translokacija. Translokaza. Raskid. Uloga proteinskih čimbenika u svakoj fazi translacije
Nakon završetka inicijacije, ribosom se nalazi na mRNA na način da se u P-centru nalazi inicijacijski kodon AUG na koji je vezan Met-tRNAshMet, a u A-centru postoji triplet koji kodira uključivanje prva aminokiselina sintetiziranog proteina. Zatim počinje najduža faza sinteze proteina - elongacija, tijekom koje ribosom, koristeći aa-tRNA, sekvencijalno "čita" mRNA u obliku tripleta nukleotida koji slijede inicijacijski kodon u smjeru od 5" do 3" kraju, produžujući polipeptidni lanac dalje zbog sekvencijalnog dodavanja aminokiselina.
Ugradnja svake aminokiseline u protein odvija se u 3 faze, tijekom kojih se: 1) aa-tRNA svake aminokiseline uključene u protein veže na A-centar ribosoma; 2) peptid iz peptidil-tRNA koji se nalazi u P-centru pridružuje se b-NH2-skupini aminoacilnog ostatka aa-tRNA A-centra i stvara novu peptidnu vezu; 3) peptidil-tRNA, produžena za jedan aminokiselinski ostatak, kreće se od A-centra do P-centra kao rezultat translokacije ribosoma.
Vezanje aminoacil-tRNA na A-mjestu. Kodon mRNA smješten u A-centru pored početnog kodona određuje prirodu aa1tRNAaa1 koja će biti uključena u A-centar. aa1tRNAaa1 stupa u interakciju s ribosomom u obliku ternarnog kompleksa koji se sastoji od faktora elongacije EF-1, aa1tRNAaa1 i GTP. Kompleks učinkovito stupa u interakciju s ribosomom samo ako je antikodon aa-tRNAaa1 komplementaran i antiparalelan mRNA kodonu u A-centru. Ugradnja aa-tRNAaa1 u ribosom događa se zahvaljujući energiji hidrolize GTP-a u GDP i anorganski fosfat. Stvaranje peptidne veze događa se neposredno nakon cijepanja EF-1 i GDP kompleksa od ribosoma. Ova faza procesa se zove transpeptidacijske reakcije
Tijekom ove reakcije, metioninski ostatak Met-tRNAIMet veže se na a-amino skupinu prve aminokiseline vezane na tRNAaa1 i smještene u A-centru, nastaje prva peptidna veza.
Translokacija - treća faza elongacije. Faktor elongacije EF-2 veže se za ribosom i, koristeći energiju GTP-a, tjera ribosom duž mRNA jedan kodon do 3" kraja. Kao rezultat, dipeptidil-tRNA, koja ne mijenja svoj položaj u odnosu na mRNA , kreće se od centra A do centra P. TRNAiMet bez metionina napušta ribosom, a sljedeći kodon ulazi u regiju centra A.
Nakon završetka treće faze produljenja, ribosom ima dipeptidil-tRNA u P-centru, a triplet koji kodira uključivanje druge aminokiseline u polipeptidni lanac ulazi u A-centar. Započinje sljedeći ciklus stadija elongacije, tijekom kojeg se gore opisani događaji ponovno odvijaju na ribosomu. Ponavljanje takvih ciklusa prema broju smislenih kodona u mRNA dovršava cijeli stadij elongacije.
Raskid translacija se događa kada jedan od stop kodona: UAG, UAA ili UGA uđe u A-centar ribosoma. Ne postoje odgovarajuće tRNA za stop kodone. Umjesto toga, 2 faktora otpuštanja proteina RF ili terminacijski faktori vezani su za ribosom. Jedan od njih, pomoću centra peptidil transferaze, katalizira hidrolitičko cijepanje sintetiziranog peptida iz tRNA. Drugi, zahvaljujući energiji hidrolize GTP-a, uzrokuje disocijaciju ribosoma na podjedinice
Dakle, predloška priroda procesa prevođenja očituje se u činjenici da je sekvenca ulaska aminoacil-tRNA u ribosom za sintezu proteina strogo određena mRNA, tj. Redoslijed kodona duž lanca mRNA jedinstveno određuje strukturu sintetiziranog proteina. Ribosom skenira mRNA lanac u obliku tripleta i sekvencijalno odabire "potrebne" aa-tRNA iz okoline, oslobađajući deacilirane tRNA tijekom elongacije.
Mala i velika podjedinica ribosoma obavljaju različite funkcije tijekom translacije: mala podjedinica veže mRNA i dekodira informacije pomoću tRNA i mehanizma translokacije, a velika podjedinica je odgovorna za stvaranje peptidnih veza.
Tijekom emitiranja Informacije zabilježene na mRNA kao niz nukleotidnih baza pretvaraju se u niz aminokiselina. Taj se proces odvija na ribosomima, a za njegovu uspješnu provedbu potrebna je još jedna vrsta RNA - kratka prijenosna RNA (tRNA). Svaka molekula tRNA ima specifičnu prostornu konfiguraciju, pomalo podsjećajući na list djeteline.
U središtu molekule (na vrhu srednjeg lista djeteline) nalazi se triplet - antikodon sposoban za uparivanje sa svojim komplementarnim tripletom (kodonom) mRNA. Triplet na kraju tRNA može formirati kovalentnu vezu sa specifičnom aminokiselinom. U stanici postoje tRNA s različitim antikodonima, odnosno sposobnim vezati se na svaku od aminokiselina potrebnih za sintezu proteina.
Sam ribosom je složen biokemijski sustav dizajniran za sintezu proteina u skladu s uputama zapisanim u strukturi mRNA. Prvo se ribosom veže na mRNA, a zatim se tRNA koja nosi aminokiselinu, čiji je antikodon komplementaran s prvim kodonom mRNA, dodaje kompleksu mRNA-ribosom. Zatim se uz prvu tRNA dodaje druga s antikodonom komplementarnim drugom kodonu mRNA, itd. Poseban enzim povezuje dvije aminokiseline koje isporučuju te dvije tRNA, a koje i dalje ostaju vezane za kompleks. Nakon toga, prva tRNA napušta ribosom kako bi pričvrstila novu molekulu svoje odgovarajuće aminokiseline. U međuvremenu, ribosom se kreće duž mRNA, a druga tRNA s aminokiselinom vezanom za nju zauzima mjesto prve. Sve se to ponavlja mnogo puta dok ribosom ne dođe do zaustavnog kodona na mRNA, koji završava bilo koji strukturni gen. Došavši do nje, ribosom i novosintetizirani protein odvajaju se od mRNA i prelaze u citoplazmu stanice.
Na jednu molekulu mRNA obično se vežu mnogi ribosomi koji, krećući se duž nje, prevode nove proteinske molekule kodon po kodon. Ova struktura se zove polisoma. Ribosomi rade vrlo učinkovito: u 1 sekundi ljudsko tijelo sintetizira 5 · 10 14 molekula hemoglobina – proteina s jedinstvenim nizom od 574 aminokiseline.
Proces biosinteze proteina jedna je od energetski najintenzivnijih reakcija u plastičnom metabolizmu stanice. Formiranje jedne peptidne veze u sintetiziranom proteinu zahtijeva četiri ATP molekule – dvije kada se aminokiselina doda tRNA i dvije izravno na ribosomu.
Aminokiseline se mogu međusobno povezivati putem veza koje se nazivaju peptidne veze, tvoreći tako polimernu molekulu. Ako broj aminokiselina ne prelazi 10, tada se zove novi spoj peptid; ako od 10 do 40 aminokiselina – polipeptid, ako više od 40 aminokiselina – protein.
Peptidna veza je veza između α-karboksilne skupine jedne aminokiseline i α-amino skupine druge aminokiseline.
Stvaranje peptidne veze
Ako je potrebno imenovati peptid, sufiks "-il" dodaje se svim nazivima aminokiselina; samo posljednja aminokiselina zadržava svoje ime nepromijenjeno. Na primjer, Alan mulj-ser mulj-triptof hr ili γ-glutamin mulj-cistein mulj-sjaj I n (nazvano drugačije glutation).
Svojstva peptidne veze uključuju:
1. Koplanarnost
Svi atomi uključeni u peptidnu skupinu nalaze se u istoj ravnini, s atomima "H" i "O" koji se nalaze na suprotnim stranama peptidne veze.

2. Trans položaj supstituenata
Radikali aminokiselina u odnosu na peptidnu osovinu C-N- spojevi su na “različitim” stranama, u poprečnom položaju.
![]()
3. Dva ekvivalentna oblika
Peptidna veza se nalazi u keto obliku i enolnom obliku.

4. Sposobnost stvaranja vodikovih veza.
Atomi kisika i vodika uključeni u peptidnu skupinu imaju sposobnost stvaranja vodikovih veza s atomima kisika i vodika drugih peptidnih skupina.

5. Peptidna veza ima djelomično karakter dvostruke veze.
Duljina peptidne veze je kraća od jednostruke veze, krute je strukture i rotacija oko nje je otežana. No budući da u proteinu osim peptidne veze postoje i druge veze, lanac aminokiselina može se okretati oko glavne osi, što proteinima daje različite konformacije (prostorni raspored atoma).
Ribosomi- intracelularne organele promjera 20-22 nm koje provode biosintezu proteina. Nalaze se u stanicama svih živih organizama. Oblik ribosoma je blizak sferičnom. Prokariotske stanice (bakterije, modrozelene alge), kao i kloroplaste i mitohondrije eukariota karakteriziraju 70 S ribosomi; 80 S ribosomi se nalaze u citoplazmi svih eukariota. S je pokazatelj brzine taloženja (sedimentacije), što je veći broj S, veća je brzina taloženja. Položaj ribosoma u citoplazmi može biti slobodan, ali najčešće su povezani s EPS-om, tvoreći polisome (asocijacije ribosoma).Bozomi u citoplazmi mogu biti slobodni, ali najčešće su povezani s EPS-om, tvoreći polisome (jedinice ribosoma pomoću glasničke RNA).
Sastav i struktura ribosoma. Ribosomi se sastoje od dvije podjedinice: velike i male. Velika podjedinica svakog ribosoma pričvršćena je na membranu najgrubljeg ER-a, a mala podjedinica strši u citoplazmatski matriks. Mali kombinuje 1 molekulu rRNA i 33 molekule različitih proteina, veliki - tri molekule rRNA i oko 40 proteina. rRNA (ribosomska) funkcionira kao okvir za proteine (imaju strukturnu i enzimatsku ulogu), a također osigurava vezanje ribosoma na specifičnu nukleotidnu sekvencu mRNA (informacijska RNA K). Obrazovanje

Ribosomi u stanicama nastavljaju samosastavljanjem iz prethodno sintetizirane RNA i proteina. Prekursori ribosomske RNA sintetiziraju se u jezgrici na nukleolarnoj DNA.
Funkcije ribosoma:
. specifično vezanje i zadržavanje komponenti sustava za sintezu proteina (glasnička RNA; transportna RNA, (GTP) i faktori translacije proteina);
. katalitičke funkcije (stvaranje peptidnih veza, hidroliza gvanozin trifosfata);
. funkcije mehaničkog kretanja supstrata (glasničke i transportne RNA), odnosno translokacije.
Emitiranje- proces stvaranja polipeptidnog lanca na matrici i RNA. Sinteza proteinskih molekula odvija se na ribosomima koji se nalaze slobodno u citoplazmi ili na grubom ER.
Faze prevođenja (Sl. 13):

Riža. 13. Shema emitiranja
Uzastopne faze sinteze polipeptida:
. mala ribosomska podjedinica veže se na met-tRNA, zatim na mRNA;
. ribosom se miješa duž RNA, što je popraćeno višestrukim ponavljanjem ciklusa dodavanja sljedeće aminokiseline u rastući polipeptidni lanac;
. Ribosom doseže jedan od zaustavnih kodona mRNA, a polipeptidni lanac se oslobađa i odvaja od ribosoma.
Aktivacija aminokiselina. Svaka od 20 aminokiselina proteina povezana je kovalentnim vezama na određenu tRNA koristeći energiju ATP-a. Reakciju katalizira specijalizirani enzim koji zahtijeva prisutnost iona magnezija – aminoacil-tRNA sintetaza.
Inicijacija proteinskog lanca. U maloj podjedinici ribosoma nalazi se funkcionalni centar s dva odjeljka – peptidil (P-odsjek) i aminoacil (A-odsjek). U prvom položaju nalazi se tRNA koja nosi određenu aminokiselinu, u drugom je tRNA koja je napunjena lancem aminokiselina. 5" kraj mRNA, koji sadrži informacije o ovom proteinu, veže se na P-mjesto s malom česticom ribosoma i s početnom aminokiselinom (formilmetionin u prokariota; metionin u eukariota) vezanom za odgovarajuću tRNA. tRNA je komplementarna tripletu sadržanom u mRNA, signalizirajući početak proteinskog lanca.
Elongacija je ciklički ponavljajući događaj u kojem dolazi do produljenja peptida. Polipeptidni lanac produžuje se sekvencijalnim dodavanjem aminokiselina, od kojih se svaka isporučuje u ribosom i umeće u određeni položaj pomoću odgovarajuće tRNA. Peptidna veza nastaje između aminokiseline iz peptidnog lanca i aminokiseline povezane s tRNA. Ribosom se kreće duž mRNA i tRNA s lancem aminokiselina ulazi u A-mjesto. Ovaj slijed događaja se ponavlja sve dok ribosomi ne dođu do terminator kodona za koji ne postoji odgovarajuća tRNA.
Raskid. Nakon završetka lančane sinteze, koju signalizira tzv. stop kodon mRNA (UAA, UAG, UGA). U tom slučaju, posljednjoj aminokiselini u peptidnom lancu dodaje se voda i njezin se karboksilni kraj odvaja od tRNA, a ribosom se raspada na dvije subčestice.
Sinteza peptida ne događa se jednim ribosomom, već nekoliko tisuća, koji tvore kompleks - polisom.
Preklapanje i obrada. Da bi poprimio normalan oblik, protein se mora saviti u određenu prostornu konfiguraciju. Prije ili nakon savijanja, polipeptid se može podvrgnuti obradi koju izvode enzimi i sastoji se od uklanjanja viška aminokiselina, dodavanja fosfata, metila i drugih skupina itd.
Pod, ispod prijevod u biologiji odnosi se na sintezu polipeptida iz aminokiselina koji se javlja u citoplazmi na ribosomima uz sudjelovanje 1) mRNA kao matrica, 2) tRNA kao nosač aminokiselina, kao i 3) broj proteinski faktori, obavljajući katalitičku funkciju u različitim fazama procesa. Translacija se događa u stanicama svih živih organizama; to je temeljni proces žive prirode.
S informacijske točke gledišta, prevođenje se može definirati kao mehanizam za prevođenje sekvence tripleta mRNA u sekvencu aminokiselina proteina.
Funkcija ribosoma je da drže mRNA, tRNA i proteinske faktore na mjestu dok ne dođe do određene kemijske reakcije. Najčešće je to stvaranje peptidne veze između susjednih aminokiselina.
Translacija i biosinteza proteinaA obično znače istu stvar. Međutim, kada govorimo o biosintezi proteina, ona često uključuje posttranslacijske modifikacije polipeptida (njihovo stjecanje sekundarne, tercijarne i kvaternarne strukture), a ponekad može uključivati i proces transkripcije. S ove točke gledišta, prevođenje se smatra važnim korakom u biosintezi proteina.
Proces prevođenja u eukariota i prokariota ima niz razlika, uglavnom povezanih s raznolikošću i aktivnošću proteinskih čimbenika.
Može postojati nekoliko ribosoma na jednom lancu mRNA, formirajući polisoma. U ovom slučaju sintetizira se nekoliko identičnih polipeptida odjednom (ali svaki je u svojoj fazi sinteze).
Sinteza jednog proteina obično traje nekoliko sekundi.
Aminokiseline iz kojih se sintetizira polipeptid nužno prolaze kroz fazu aktivacije. Sam proces prevođenja uključuje tri faze: inicijaciju, produljenje i završetak.
Proces prevođenja ima svojstvo specifičnosti. Prvo, specifični kodoni mRNA imaju vlastite tRNA. Drugo, aminokiseline su vezane samo za "njihove" tRNA.
Aktivacija aminokiselina
Aktivacija aminokiselina je nužna jer se samo u tom stanju one mogu spojiti s tRNA i kasnije međusobno formirati peptidne veze.
Citoplazma stanica uvijek sadrži slobodne (nekombinirane s drugim tvarima) aminokiseline. Specifični enzimi, u prisutnosti ATP-a, pretvaraju aminokiselinu u aminoaciladenilat, koji se već može vezati za tRNA.
Postoji klasa enzima - aminoacil-tRNA sintetazas, – koji aktiviraju aminokiseline koristeći energiju ATP-a. Svaku aminokiselinu aktivira vlastiti enzim, nakon čega se veže samo za svoju tRNA. Nastaje kompleks aminokiselina s tRNA - aminoacil-tRNA (aa-tRNA).
Pokretanje emitiranja
Inicijacija prevođenja uključuje sljedeće sekvencijalne faze uz sudjelovanje čimbenika inicijacije:
Pričvršćivanje 5" kraja mRNA na malu podjedinicu ribosoma. U ovom slučaju, početni kodon (AUG) nalazi se na nedovršenom (zbog nepostojanja velike podjedinice) P-mjestu ribosoma.
Kompleks aa-tRNA s odgovarajućim antikodonom vezan je za početni kodon mRNA. Kod eukariota kodon AUG kodira aminokiselinu metionin; kod prokariota kodon formil-metionin. Ove početne aminokiseline kasnije se izrezuju iz gotovog polipeptida.
Ribosomske podjedinice su ujedinjene, zbog čega su njihova P- i A-mjesta dovršena.
Dakle, u fazi inicijacije, ribosom prepoznaje početni kodon i priprema se za početak sinteze.
Rezultirajuća veza između ribosoma i mRNA je reverzibilna; nakon sinteze polipeptida, mRNA se može odvojiti od ribosoma. Nakon toga, mRNA se ponovno koristi ili uništava posebnim enzimima.
Početni kodon AUG razlikuje se od drugih sličnih kodona u sredini mRNA po tome što mu prethodi kapica i određeni nizovi nukleotida. Zahvaljujući njima, AUG je prepoznat kao starter. (Ovo se uglavnom odnosi na eukariote.)
Emitiranje istezanja
U ovoj fazi dolazi do izravne sinteze polipeptidnog lanca. Proces elongacije sastoji se od mnogo ciklusa. Jedan ciklus produljenja je dodavanje jedne aminokiseline rastućem polipeptidnom lancu.
Već u fazi inicijacije, P-mjesto ribosoma zauzima prva tRNA koja nosi aminokiselinu metionin. U prvom ciklusu elongacije, drugi aa-tRNA kompleks ulazi u A mjesto ribosoma. To će biti tRNA čiji je antikodon komplementaran sljedećem (nakon početnog AUG) kodonu.
Mjesta A(aminoacil) i P(peptidil) postavljaju komplekse aa-tRNA tako da dolazi do kemijske reakcije između aminokiselina i stvaranja peptidne veze.
Nakon toga, prva (smještena na P-mjestu) tRNA se oslobađa svoje aminokiseline. Kao rezultat toga, čini se da je potonja povezana samo s drugom aminokiselinom peptidnom vezom. Druga aminokiselina je vezana za drugu tRNA koja se nalazi na A mjestu.
Ribosom se pomiče duž lanca mRNA jedan triplet. U tom slučaju prva tRNA završava na E-mjestu (izlazu) ribosoma, nakon čega ga napušta. Druga tRNA, vezana za dvije aminokiseline, ide na P mjesto. Mjesto A je oslobođeno za ulazak trećeg aa-tRNA kompleksa.
Sljedeći ciklusi elongacije odvijaju se slično prvom. Kada se A-mjesto oslobodi, aa-tRNA može ući u njega, čiji je antikodon komplementaran mRNA kodonu koji se trenutno nalazi na A-mjestu.
Prekid emitiranja
Terminacija je završetak sinteze polipeptidnog lanca i njegovo odvajanje. Terminacija se događa kada ribosom naiđe na jedan od terminacijskih kodona (UAA, UAG, UGA), za koje ne postoje tRNA. Ove dijelove mRNA prepoznaju posebni proteini - čimbenici prekida.
 Bojni brod “Victory” – Legendarni jedrenjaci
Bojni brod “Victory” – Legendarni jedrenjaci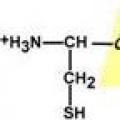 Stvaranje peptidne veze Dva ekvivalentna oblika
Stvaranje peptidne veze Dva ekvivalentna oblika Kasatkin Mihail Aleksandrovič (1902–1974) M Kasatkin
Kasatkin Mihail Aleksandrovič (1902–1974) M Kasatkin