Kiến thức về quy luật dãy tương đồng của biến dị di truyền cho phép. Định luật N.I. Vavilov (định luật về chuỗi tương đồng về tính biến thiên di truyền)
Vào ngày 4 tháng 6, ông đã có bài thuyết trình về “Quy luật về dãy tương đồng trong biến dị di truyền”. Đây là một trong những công trình được coi là cơ bản và là cơ sở lý luận của nghiên cứu sinh học. Bản chất của quy luật bắt nguồn từ thực tế là các loài và chi có quan hệ gần gũi về mặt di truyền (được kết nối với nhau bởi sự thống nhất về nguồn gốc) được đặc trưng bởi các chuỗi giống nhau về tính biến đổi di truyền. Niềm đam mê nghiên cứu ngũ cốc của sinh viên, sau đó là các loại cây họ cải, cây họ đậu và bí ngô đã cho phép Vavilov và các sinh viên của ông tìm ra những đột biến tương tự ở các loài liên quan và sau đó là các chi. Trong bảng được phát triển từ kết quả của các thí nghiệm, Vavilov đã đánh dấu bằng dấu “+” các đột biến mà biểu hiện của chúng được tìm thấy ở những loài này và các khoảng trống cho thấy rằng những đột biến đó lẽ ra đã tồn tại nhưng vẫn chưa được phát hiện. Một bảng có các ô trống, sẽ được lấp đầy bằng sự phát triển hơn nữa của khoa học. Chúng ta đã từng gặp phải điều gì như thế này trước đây chưa?! Tất nhiên, trong hóa học, bảng tuần hoàn nổi tiếng! Mô hình của hai định luật này đã được khoa học xác nhận. Các ô “trống” được lấp đầy và đây là cơ sở để lựa chọn thực tế. Lúa mì cứng chỉ được biết đến ở dạng mùa xuân, nhưng theo quy luật, lúa mì cứng ở dạng mùa đông cũng phải tồn tại trong tự nhiên. Nó thực sự đã sớm được phát hiện ở biên giới Iran và Thổ Nhĩ Kỳ. Bí ngô và dưa có đặc điểm là quả đơn giản và nhiều múi, nhưng dưa hấu có hình dạng này không được mô tả vào thời Vavilov. Nhưng dưa hấu phân đoạn được phát hiện ở phía đông nam nước Nga thuộc châu Âu. Nền văn hóa chủ yếu là trồng củ cải ba mầm, những loại cây trồng cần làm cỏ và loại bỏ hai chồi thừa. Nhưng trong số họ hàng của củ cải đường trong tự nhiên cũng có những dạng củ cải đơn mầm nên các nhà khoa học đã có thể tạo ra nhiều loại củ cải đường đơn mầm mới. Bản chất không có mái che của cây ngũ cốc là một đột biến đã được chứng minh là có lợi khi áp dụng máy thu hoạch, khi các cơ chế trở nên ít bị tắc nghẽn hơn. Các nhà tạo giống, sử dụng định luật Vavilov, đã tìm ra những dạng không có đầu và tạo ra các giống ngũ cốc không có đầu mới. Sự thật về sự biến đổi song song ở các loài gần và xa đã được Charles Darwin biết đến. Ví dụ, cùng màu lông của loài gặm nhấm, bệnh bạch tạng ở đại diện của các nhóm khác nhau trong thế giới động vật và con người (một trường hợp bệnh bạch tạng ở người da đen đã được mô tả), thiếu bộ lông ở chim, thiếu vảy ở cá, màu sắc tương tự của các loại quả thuộc loại cây ăn quả và quả mọng, tính biến đổi của các loại cây lấy củ, v.v. Lý do tính song song trong tính biến đổi nằm ở chỗ các đặc điểm tương đồng dựa trên sự hiện diện của các gen giống nhau: loài và chi càng gần nhau về mặt di truyền thì sự giống nhau về mặt di truyền càng hoàn thiện hơn. chuỗi biến thiên. Do đó gây ra đột biến tương đồng - nguồn gốc chung của các kiểu gen. Trong quá trình tiến hóa, thiên nhiên sống được lập trình như thể theo một công thức, bất kể thời gian xuất hiện của loài. Định luật về chuỗi tương đồng trong biến dị di truyền của N.I. Vavilov không chỉ là sự xác nhận học thuyết của Darwin về nguồn gốc các loài mà còn mở rộng ý tưởng về biến dị di truyền. Nikolai Ivanovich một lần nữa có thể tuyên bố: “Cảm ơn Darwin!”, nhưng cũng “Tiếp tục Darwin!” Hãy quay trở lại năm 1920. Ký ức của những người chứng kiến thật thú vị. Alexandra Ivanovna Mordvinkina, người có mặt tại đại hội của Viện Nông nghiệp Saratov (sau này là ứng cử viên khoa học sinh học), nhớ lại: “Đại hội khai mạc tại khán phòng lớn nhất của trường đại học. Không một báo cáo nào sau đó gây ấn tượng mạnh với tôi như bài phát biểu của Nikolai Ivanovich. Anh ấy nói đầy hứng khởi, mọi người nín thở lắng nghe, có cảm giác như một điều gì đó rất lớn lao, mới mẻ trong khoa học đang mở ra trước mắt chúng tôi. Khi có những tràng pháo tay như vũ bão và kéo dài, Giáo sư Vyacheslav Rafailovich Zelensky nói: “Đây là những nhà sinh vật học chào đón Mendeleev của họ”. Câu nói của Nikolai Maksimovich Tulaikov đặc biệt in sâu vào trí nhớ của tôi: “Có thể bổ sung thêm điều gì vào báo cáo này? Tôi có thể nói một điều: Nước Nga sẽ không diệt vong nếu có những người con như Nikolai Ivanovich”. Nikolai Vladimirovich Timofeev-Resovsky, một nhà di truyền học xuất sắc, người biết đến Vavilov không chỉ từ công việc của ông mà còn về mặt cá nhân, đã nói chuyện một cách tâm sự với những người quen thân: “Nikolai Ivanovich là một người đàn ông tuyệt vời và một liệt sĩ vĩ đại, một nhà nhân giống và sưu tập thực vật xuất sắc, một nhà du lịch, một người dũng cảm và được mọi người yêu thích, nhưng loạt luật tương đồng của ông - luật hoàn toàn không phải là chuỗi tương đồng, mà là loạt tương tự, vâng, thưa ông! Tương đồng là gì? Đây là sự tương đồng dựa trên nguồn gốc chung. Một sự tương tự là gì? Sự giống nhau về các đặc điểm bên ngoài, được xác định bởi môi trường sống tương tự, nhưng không phải bởi mối quan hệ họ hàng. Vậy ai đúng? Vavilov! Người ta chỉ có thể ngưỡng mộ chiều sâu tâm hồn sinh học của anh ấy! Chỉ thay đổi một thuật ngữ trong tiêu đề sẽ thay đổi bản chất của luật. Theo quy luật chuỗi tương đồng, mọi người đều bình đẳng, vì họ có cùng nguồn gốc sinh học và thuộc loài homo sapiens, tức là mọi người đều thông minh, có năng lực và tài năng như nhau, v.v., nhưng họ có những khác biệt bên ngoài: về chiều cao, tỷ lệ giữa các bộ phận cơ thể, v.v. Theo quy luật chuỗi tương tự, con người có ngoại hình giống nhau, vì họ có môi trường sống giống nhau nhưng nguồn gốc khác nhau. Và đây đã là chỗ cho chủ nghĩa Sô vanh, chủ nghĩa phân biệt chủng tộc, chủ nghĩa dân tộc, thậm chí là diệt chủng. Và định luật Vavilov nói rằng người lùn ở Châu Phi và cầu thủ bóng rổ ở Mỹ có cùng nguồn gốc di truyền, và không thể đặt người này lên trên người kia - điều này phản khoa học! Giá trị của mô hình sinh học phổ quát do Vavilov phát hiện đã được xác nhận bởi nghiên cứu hiện đại không chỉ ở thực vật mà còn ở động vật. Các nhà di truyền học hiện đại tin rằng quy luật này bộc lộ những triển vọng vô tận về kiến thức khoa học, sự khái quát hóa và dự đoán” (Giáo sư M. E. Lobanov). Một tác phẩm cơ bản khác của N. I. Vavilov, “Khả năng miễn dịch của thực vật đối với các bệnh truyền nhiễm” (1919), có từ thời Saratov. Trên trang tựa của cuốn sách, Nikolai Ivanovich viết: “Dành để tưởng nhớ nhà nghiên cứu vĩ đại về khả năng miễn dịch Ilya Ilyich Mechnikov”. Không một nhà khoa học vĩ đại nào tự coi mình là người đứng ngoài khoa học. Vì vậy, nhờ có Mechnikov, Vavilov tự hỏi liệu thực vật có thể có sức mạnh bảo vệ nếu động vật có chúng hay không? Để tìm kiếm câu trả lời cho câu hỏi, ông đã tiến hành nghiên cứu về ngũ cốc bằng phương pháp ban đầu và tóm tắt thực tiễn và lý thuyết, đặt nền móng cho một ngành khoa học mới - miễn dịch thực vật học. Công trình này có ý nghĩa hoàn toàn thực tế - sử dụng khả năng miễn dịch tự nhiên của thực vật như một cách hợp lý và tiết kiệm chi phí nhất để kiểm soát sâu bệnh. Nhà khoa học trẻ đã tạo ra một lý thuyết ban đầu về khả năng miễn dịch sinh lý của thực vật đối với các bệnh truyền nhiễm, và cơ sở giảng dạy của ông là nghiên cứu về khả năng miễn dịch kiểu gen. N.I. Vavilov đã nghiên cứu phản ứng của “vật chủ” trước sự xâm nhập của ký sinh trùng, tính đặc hiệu của phản ứng này và tìm ra liệu toàn bộ loạt bệnh có miễn dịch hay chỉ một số loài nhất định trong loạt này. Nikolai Ivanovich đặc biệt coi trọng khả năng miễn dịch nhóm, tin rằng trong chăn nuôi, điều quan trọng là phải phát triển các giống có khả năng kháng không phải với một chủng tộc mà với toàn bộ quần thể các chủng tộc sinh lý, và những loài kháng bệnh như vậy phải được tìm kiếm ở quê hương của thực vật. Khoa học sau này đã xác nhận các loài hoang dã - họ hàng của cây trồng - có khả năng miễn dịch tự nhiên và ít mắc các bệnh truyền nhiễm hơn. Việc đưa các gen kháng vào thực vật là điều mà các nhà nhân giống hiện đại đang thực hiện, sử dụng lý thuyết của N. I. Vavilov và các phương pháp kỹ thuật di truyền. Nhà khoa học quan tâm đến sự phát triển các vấn đề miễn dịch trong suốt sự nghiệp khoa học của mình: “Học thuyết về khả năng miễn dịch thực vật đối với các bệnh truyền nhiễm” (1935), “Quy luật miễn dịch thực vật tự nhiên đối với các bệnh truyền nhiễm (Chìa khóa để tìm ra các dạng miễn dịch)” ( chỉ được xuất bản vào năm 1961). ). Viện sĩ Pyotr Mikhailovich Zhukovsky đã nhận xét rất đúng: “Trong thời kỳ Saratov, tuy ngắn ngủi (1917-1921), ngôi sao N. đã trỗi dậy. I. Vavilov - nhà khoa học." Vavilov sau này viết: “Tôi di cư từ Saratov vào tháng 3 năm 1921 cùng với toàn bộ phòng thí nghiệm gồm 27 người.” Ông được bầu làm Trưởng phòng Thực vật học Ứng dụng của Ủy ban Khoa học Nông nghiệp ở Petrograd. Từ 1921 đến 1929 - Giáo sư Bộ môn Di truyền và Giống cây trồng, Viện Nông nghiệp Leningrad. Năm 1921, V.I. Lênin cử hai nhà khoa học đi dự hội nghị ở Mỹ, một trong số họ là N.I. Vavilov. Báo cáo về nghiên cứu di truyền đã khiến nó trở nên phổ biến trong giới khoa học tại hội nghị. Ở Mỹ, màn trình diễn của anh ấy đi kèm với sự hoan nghênh nhiệt liệt, tương tự như màn trình diễn sau đó của Chkalov. Các tờ báo Mỹ hét lên: “Nếu tất cả người Nga đều như vậy thì chúng ta cần phải làm bạn với họ”. Trong 20-30 năm nữa. N.I. Vavilov cũng thể hiện mình là nhà tổ chức khoa học lớn. Ông thực sự là người sáng lập và giám đốc thường trực của Viện trồng cây trồng toàn Liên minh (VIR). Năm 1929, Viện Khoa học Nông nghiệp Liên minh (VASKhNIL) được thành lập trên cơ sở Viện Nông học Thực nghiệm Liên minh, trước đây được tổ chức bởi Vavilov. Ông được bầu làm tổng thống đầu tiên (từ 1929 đến 1935). Với sự tham gia trực tiếp của nhà khoa học, Viện Di truyền của Viện Hàn lâm Khoa học Liên Xô đã được thành lập. Trong một thời gian ngắn, tài năng của Vavilov đã tạo ra một trường phái khoa học về di truyền học, trường này trở thành trường hàng đầu thế giới. Tất cả các công việc ban đầu ở nước ta trong lĩnh vực di truyền đều do ông hoặc dưới sự lãnh đạo của ông thực hiện. Tại VIR, phương pháp đa bội thực nghiệm đã được sử dụng lần đầu tiên và G. D. Karpechenko bắt đầu nghiên cứu việc sử dụng phương pháp này trong lai tạo từ xa. Vavilov nhất quyết bắt đầu nghiên cứu việc sử dụng hiện tượng dị hợp và lai giữa các dòng. Ngày nay đây là ABC của sự lựa chọn, nhưng đó là sự khởi đầu. Hơn 30 năm hoạt động khoa học, đã xuất bản khoảng 400 công trình, bài báo! Trí nhớ phi thường, kiến thức bách khoa, thông thạo gần 20 ngôn ngữ, nhận thức được mọi đổi mới trong khoa học. Làm việc 18-20 giờ một ngày. Mẹ anh mắng anh: “Con thậm chí không có thời gian để ngủ…”, con trai Vavilov nhớ lại.
BIẾN ĐỔI
Kế hoạch
Sự khác biệt giữa đột biến và sửa đổi.
Phân loại đột biến.
Định luật N.I.Vavilov
Đột biến. Khái niệm đột biến. Yếu tố gây đột biến.
Đột biến – là những thay đổi đột ngột, lâu dài, tự nhiên hoặc nhân tạo trong vật liệu di truyền xảy ra dưới tác động của yếu tố gây đột biến .
Các loại yếu tố gây đột biến:
MỘT) thuộc vật chất- Bức xạ, nhiệt độ, bức xạ điện từ.
B) các yếu tố hoá học các chất gây ngộ độc cơ thể: rượu, nicotin, formaldehyde.
TRONG) sinh học- Virus, vi khuẩn.
Sự khác biệt giữa đột biến và sửa đổi
Phân loại đột biến
Có một số cách phân loại đột biến.
I Phân loại đột biến theo ý nghĩa: có lợi, có hại, trung tính.
Hữu íchđột biến dẫn đến tăng sức đề kháng của sinh vật và là nguyên liệu cho chọn lọc tự nhiên và nhân tạo.
Đột biến có hại giảm sức sống và dẫn đến phát triển các bệnh di truyền: bệnh máu khó đông, bệnh thiếu máu hồng cầu hình liềm.
II Phân loại đột biến theo vị trí hoặc nơi xảy ra: soma và thế hệ.
Dạng cơ thể phát sinh trong các tế bào của cơ thể và chỉ ảnh hưởng đến một phần cơ thể, trong khi các cá thể khảm phát triển: mắt khác nhau, màu tóc. Những đột biến này chỉ được di truyền trong quá trình nhân giống sinh dưỡng (ở cây nho).
sáng tạo – xảy ra trong tế bào mầm hoặc trong tế bào từ đó giao tử được hình thành. Chúng được chia thành hạt nhân và ngoại hạt nhân (ty thể, plastid).
III Đột biến do tính chất thay đổi kiểu gen: nhiễm sắc thể, bộ gen, gen.
di truyền (hoặc điểm) không thể nhìn thấy dưới kính hiển vi, có liên quan đến sự thay đổi cấu trúc gen. Những đột biến này xảy ra do mất nucleotide, chèn hoặc thay thế một nucleotide này bằng một nucleotide khác. Những đột biến này dẫn đến các bệnh về gen: mù màu, phenylketon niệu.
Nhiễm sắc thể (perestroika) liên quan đến sự thay đổi cấu trúc nhiễm sắc thể. Điều gì có thể xảy ra:
Xóa: - mất một phần nhiễm sắc thể;
Sao chép – nhân đôi đoạn nhiễm sắc thể;
Đảo ngược – quay một phần của nhiễm sắc thể 180 0;
Chuyển vị – trao đổi các đoạn nhiễm sắc thể không tương đồng và sáp nhập hai nhiễm sắc thể không tương đồng thành một.
Nguyên nhân gây đột biến nhiễm sắc thể: sự xuất hiện của hai hoặc nhiều sự đứt gãy nhiễm sắc thể sau đó là sự kết hợp của chúng nhưng theo thứ tự sai.
Bộ gen đột biến dẫn tới sự thay đổi số lượng nhiễm sắc thể. Phân biệt dị bội Và đa bội.
Dị thể liên quan đến sự thay đổi số lượng nhiễm sắc thể trên một số nhiễm sắc thể – 1.2.3. nguyên nhân: sự không phân ly của nhiễm sắc thể trong giảm phân:
- Độc thân – số lượng nhiễm sắc thể giảm đi 1 nhiễm sắc thể. Công thức chung của bộ nhiễm sắc thể là 2n-1.
- Tam giác học – tăng số lượng nhiễm sắc thể lên 1. Công thức chung 2n+1 (hội chứng Clanfeather 47 nhiễm sắc thể; tam phân 21 cặp nhiễm sắc thể - Hội chứng Down (dấu hiệu của nhiều khuyết tật bẩm sinh làm giảm sức sống của cơ thể và kém phát triển trí tuệ).
Đa bội – nhiều thay đổi về số lượng nhiễm sắc thể. Ở các sinh vật đa bội, bộ nhiễm sắc thể đơn bội (n) trong tế bào được lặp lại không phải 2 lần như ở các sinh vật lưỡng bội mà là 4-6 lần, đôi khi nhiều hơn - lên đến 10-12 lần.
Sự xuất hiện của đa bội có liên quan đến sự vi phạm nguyên phân hoặc giảm phân. Đặc biệt, sự không phân kỳ của các nhiễm sắc thể tương đồng trong giảm phân dẫn đến sự hình thành giao tử với số lượng nhiễm sắc thể tăng lên. Ở sinh vật lưỡng bội, quá trình này có thể tạo ra giao tử lưỡng bội (2n).
Được tìm thấy rộng rãi trong các loại cây trồng: kiều mạch, hướng dương, v.v., cũng như trong thực vật hoang dã.
Định luật N.I. Vavilov (định luật về chuỗi tương đồng về tính biến thiên di truyền).
/ Từ xa xưa, các nhà nghiên cứu đã quan sát thấy sự tồn tại của những đặc điểm giống nhau ở các loài và chi khác nhau trong cùng một họ, chẳng hạn như dưa, giống dưa chuột, hoặc dưa hấu, giống dưa. Những sự thật này đã hình thành nên cơ sở của quy luật về chuỗi tương đồng trong tính biến thiên di truyền./
Đa allelism. Biến thiên song song. Một gen có thể tồn tại ở nhiều hơn hai trạng thái. Tính đa dạng về số alen của một gen được gọi là đa allelism. Các alen khác nhau xác định mức độ khác nhau của cùng một tính trạng. Càng mang nhiều alen trong quần thể thì loài càng có tính dẻo thì càng thích nghi tốt hơn với các điều kiện môi trường thay đổi.
Cơ sở của đa allelism biến thiên song song - hiện tượng trong đó các đặc điểm giống nhau xuất hiện ở các loài và chi khác nhau trong cùng một họ. Hệ thống hóa các sự kiện về biến thiên song song N.I. Vavilov./
N.I. Vavilov so sánh các loài thuộc họ Ngũ cốc. Ông phát hiện ra rằng nếu lúa mì mềm có dạng mùa đông và mùa xuân, có lông và không có lông, thì những dạng tương tự nhất thiết phải được tìm thấy ở lúa mì cứng. Hơn nữa, thành phần của các tính năng. Về những dạng khác nhau trong một loài và chi, nó thường giống nhau ở các chi khác. Ví dụ, các dạng lúa mạch đen và lúa mạch lặp lại các dạng của các loại lúa mì khác nhau, tạo thành cùng một chuỗi biến đổi di truyền song song hoặc tương đồng.
Hệ thống hóa các sự kiện cho phép N.I. Vavilov hình thành định luật về chuỗi tương đồng trong tính biến thiên di truyền (1920): các loài và chi gần gũi về mặt di truyền được đặc trưng bởi chuỗi biến đổi di truyền tương tự nhau với tính đều đặn như vậy. Rằng, khi biết một số dạng trong một loài, người ta có thể dự đoán sự hiện diện của các dạng song song ở các loài và chi khác.
Sự tương đồng về đặc điểm di truyền của các loài và chi có quan hệ gần gũi được giải thích bằng sự tương đồng về gen của chúng, vì chúng có nguồn gốc từ cùng một loài tổ tiên. Ngoài ra, quá trình đột biến ở các loài gần gũi về mặt di truyền cũng diễn ra tương tự. Do đó, chúng phát triển một loạt các alen lặn tương tự nhau và kết quả là các tính trạng song song.
Kết luận từ định luật Vavilov: mỗi loài có ranh giới nhất định về sự biến đổi đột biến. Không có quá trình đột biến nào có thể dẫn đến những thay đổi vượt ra ngoài phạm vi biến đổi di truyền của loài. Do đó, ở động vật có vú, đột biến có thể làm thay đổi màu lông từ đen sang nâu, đỏ, trắng, sọc và đốm, nhưng loại trừ sự xuất hiện của màu xanh lục.
©2015-2019 trang web
Tất cả các quyền thuộc về tác giả của họ. Trang web này không yêu cầu quyền tác giả nhưng cung cấp quyền sử dụng miễn phí.
Ngày tạo trang: 2016-04-12
thiết lập sự song song trong sự di truyền và biến đổi của sinh vật. Được công thức hóa bởi N. I. Vavilov vào năm 1920. Trong khi nghiên cứu tính biến đổi của các đặc tính trong loài và chi của ngũ cốc và các họ khác, N. I. Vavilov đã phát hiện ra rằng: 1. Các loài và chi gần nhau về mặt di truyền được đặc trưng bởi chuỗi biến dị di truyền giống hệt nhau với Tính quy luật là khi biết nhiều dạng của một loài, người ta có thể dự đoán sự hiện diện của các dạng giống hệt nhau ở các loài và chi khác. Các chi và Linneons càng có vị trí di truyền gần nhau trong hệ thống chung thì sự đồng nhất về cấp độ biến đổi của chúng càng hoàn thiện. 2. Toàn bộ họ thực vật thường có đặc điểm là một chu kỳ biến đổi nhất định trải qua tất cả các chi tạo nên họ.” Mặc dù luật ban đầu liên quan đến tính biến đổi ở thực vật, nhưng N.I. Vavilov đã chỉ ra khả năng áp dụng nó đối với động vật. lý thuyết cơ sở tương đồng của dãy kiểu hình. sự biến đổi trong phân loại liên quan. các nhóm là ý tưởng về sự thống nhất về nguồn gốc của họ thông qua sự đa dạng hóa dưới tác động của thiên nhiên. lựa chọn. Vì tổ tiên chung của các loài hiện có đều có một đặc tính cụ thể, cụ thể một bộ gen thì con cháu của chúng phải có cùng một bộ gen, trừ những ngoại lệ nhỏ. Xét rằng mỗi gen có thể đột biến theo các hướng khác nhau (đa bội, allelism) và quá trình đột biến là không định hướng, điều tự nhiên là giả định rằng phổ biến đổi ở các gen giống hệt nhau ở các cá thể của các loài gần gũi sẽ tương tự nhau. Vì vậy, pháp luật dựa trên sự tương đồng. hàng (3. r.) nằm ở sự song song của kiểu gen. sự đa dạng ở những cá thể có bộ gen giống nhau. Mang tính lý thuyết. Trên cơ sở di truyền học so sánh, định luật giải thích tính đa hình của loài và do đó biện minh cho tính toàn vẹn của loài, bất chấp sự tồn tại trong ranh giới của các dạng hình thái khác biệt rõ ràng. Mặt khác, quy luật mang lại sự rõ ràng cho hiện tượng kiểu hình. "đồng nhất" số nhiều loài, rìa có thể liên quan đến tính dị hợp tử của chúng và hiện tượng thống trị, được bộc lộ trong quá trình cận huyết. 3. r., phản ánh mô hình chung của quá trình đột biến và hình thành sinh vật, là biol. cơ sở của các phương pháp nhằm mục đích đạt được những kế thừa và thay đổi cần thiết. Ông chỉ ra cho các nhà tạo giống các hướng đi của nghệ thuật, chọn lọc, hoặc, như N.I. Vavilov đã viết, “cần tìm kiếm gì” và các phương pháp tìm kiếm có thể khác nhau: từ việc tìm kiếm các dạng mong muốn trong tự nhiên hoặc xác định chúng thông qua cận huyết cho đến thu được các dạng này bằng cách sử dụng chất gây đột biến. Sinh hóa cơ chế 3. g.r. được nghiên cứu rộng rãi trên nhiều đối tượng khác nhau - từ những thay đổi trong quá trình trao đổi chất của vi khuẩn đến các quá trình vi sinh vật. tổng hợp di truyền, bệnh tật của con người.
Luật chuỗi tương đồng
Việc xử lý tài liệu quan sát và thực nghiệm sâu rộng, nghiên cứu chi tiết về tính biến đổi của nhiều loài Linnaean (Linneons), một số lượng lớn thông tin mới thu được chủ yếu từ nghiên cứu cây trồng và họ hàng hoang dã của chúng, đã cho phép N.I. Vavilov đã tập hợp tất cả các ví dụ đã biết về sự biến đổi song song và xây dựng một quy luật chung mà ông gọi là “Quy luật về chuỗi tương đồng trong biến thể di truyền” (1920), mà ông đã báo cáo tại Đại hội các nhà tạo giống toàn Nga lần thứ ba, được tổ chức tại Saratov. Năm 1921 N.I. Vavilov được cử đến Mỹ để tham dự Đại hội Nông nghiệp Quốc tế, nơi ông trình bày về quy luật của chuỗi tương đồng. Quy luật biến đổi song song của các chi và loài gần gũi, được thiết lập bởi N.I. Vavilov và gắn liền với nguồn gốc chung, phát triển giáo lý tiến hóa của Charles Darwin, được khoa học thế giới đánh giá cao. Nó được khán giả coi là sự kiện lớn nhất trong khoa học sinh học thế giới, mở ra những chân trời rộng lớn nhất cho thực hành.
Quy luật chuỗi tương đồng trước hết thiết lập cơ sở cho tính hệ thống của sự đa dạng to lớn của các dạng thực vật trong đó thế giới hữu cơ rất phong phú, cho phép nhà tạo giống có được ý tưởng rõ ràng về vị trí của mỗi loài, thậm chí cả đơn vị nhỏ nhất, có hệ thống trong thế giới thực vật và đánh giá tính đa dạng có thể có của nguồn nguyên liệu để lựa chọn.
Các quy định chính của định luật về chuỗi tương đồng như sau.
"1. Các loài và chi gần nhau về mặt di truyền được đặc trưng bởi chuỗi biến đổi di truyền giống nhau với mức độ đều đặn đến mức khi biết chuỗi các dạng trong một loài, người ta có thể dự đoán sự hiện diện của các dạng song song ở các loài và chi khác. Các chi và Linneons càng có vị trí di truyền gần nhau trong hệ thống chung thì sự giống nhau về chuỗi biến đổi của chúng càng hoàn thiện.
2. Toàn bộ họ thực vật thường được đặc trưng bởi một chu kỳ biến đổi nhất định xuyên qua tất cả các chi và loài tạo nên các họ đó.”
Ngay cả tại Đại hội tuyển chọn toàn Nga lần thứ III (Saratov, tháng 6 năm 1920), nơi N.I. Vavilov lần đầu tiên báo cáo về khám phá của mình, tất cả những người tham gia đại hội đều công nhận rằng “giống như bảng tuần hoàn (hệ thống tuần hoàn)”, định luật về chuỗi đồng đẳng sẽ giúp dự đoán sự tồn tại, tính chất và cấu trúc của các dạng và loài thực vật vẫn chưa được biết đến và động vật và đánh giá cao ý nghĩa khoa học và thực tiễn của bộ luật này. Những tiến bộ hiện đại trong sinh học tế bào phân tử giúp người ta có thể hiểu được cơ chế tồn tại của sự biến đổi tương đồng ở các sinh vật gần gũi - dựa trên sự giống nhau chính xác của các dạng và loài trong tương lai với các dạng hiện có - và tổng hợp một cách có ý nghĩa các dạng thực vật mới không tồn tại trong tự nhiên. Giờ đây nội dung mới đang được bổ sung vào định luật Vavilov, giống như sự xuất hiện của lý thuyết lượng tử đã mang lại nội dung mới, sâu sắc hơn cho hệ thống tuần hoàn của Mendeleev.
Học thuyết về trung tâm nguồn gốc cây trồng
Vào giữa những năm 20, nghiên cứu về sự phân bố địa lý và tính đa dạng trong loài của các loại cây nông nghiệp khác nhau, được thực hiện bởi N.I. Vavilov và dưới sự lãnh đạo của ông, đã cho phép Nikolai Ivanovich hình thành các ý tưởng về trung tâm địa lý xuất xứ của cây trồng. Cuốn sách “Trung tâm xuất xứ của cây trồng” được xuất bản năm 1926. Ý tưởng dựa trên lý thuyết sâu sắc về trung tâm xuất xứ đã cung cấp cơ sở khoa học cho việc tìm kiếm có mục tiêu các loại thực vật có ích cho con người và được sử dụng rộng rãi cho các mục đích thực tế.
Tầm quan trọng không kém đối với khoa học thế giới là lời giảng dạy của N.I. Vavilov về trung tâm nguồn gốc của cây trồng và về các mô hình địa lý trong sự phân bố các đặc điểm di truyền của chúng (xuất bản lần đầu vào năm 1926 và 1927). Trong những tác phẩm kinh điển này N.I. Vavilov là người đầu tiên đưa ra một bức tranh mạch lạc về sự tập trung của vô số dạng cây trồng ở một vài trung tâm chính về nguồn gốc của chúng và tiếp cận câu hỏi về nguồn gốc của cây trồng theo một cách hoàn toàn mới. Nếu trước ông, các nhà thực vật học-địa lý (Alphonse De-Candolle và những người khác) đang tìm kiếm quê hương “chung” của lúa mì, thì Vavilov đang tìm kiếm trung tâm nguồn gốc của từng loài, nhóm loài lúa mì ở nhiều khu vực khác nhau trên thế giới. Trong trường hợp này, điều đặc biệt quan trọng là xác định các khu vực phân bố tự nhiên (các khu vực) giống của một loài nhất định và xác định trung tâm của sự đa dạng lớn nhất về các dạng của nó (phương pháp địa lý thực vật).
Để thiết lập sự phân bố địa lý của các giống và chủng loại cây trồng và họ hàng hoang dã của chúng, N.I. Vavilov đã nghiên cứu các trung tâm văn hóa nông nghiệp cổ đại, khởi đầu là các vùng núi ở Ethiopia, Tây và Trung Á, Trung Quốc, Ấn Độ, trên dãy Andes của Nam Mỹ, chứ không phải ở các thung lũng rộng lớn của các con sông lớn - sông Nile , Ganges, Tigris và Euphrates, như các nhà khoa học đã tuyên bố trước đó. Kết quả nghiên cứu khảo cổ học tiếp theo đã xác nhận giả thuyết này.
Để tìm ra trung tâm đa dạng và phong phú của các dạng thực vật, N.I. Vavilov đã tổ chức nhiều cuộc thám hiểm theo một kế hoạch cụ thể tương ứng với những khám phá lý thuyết của ông (chuỗi tương đồng và trung tâm nguồn gốc của cây trồng), vào năm 1922–1933. đã đến thăm 60 quốc gia trên thế giới, cũng như 140 vùng của nước ta. Kết quả là một nguồn tài nguyên thực vật quý giá của thế giới đã được thu thập, lên tới hơn 250.000 mẫu vật. Bộ sưu tập phong phú nhất được thu thập đã được nghiên cứu cẩn thận bằng các phương pháp chọn lọc, di truyền, hóa học, hình thái, phân loại và cây trồng địa lý. Nó vẫn được lưu trữ trong VIR và được sử dụng bởi các nhà chăn nuôi của chúng tôi và nước ngoài.
Sự sáng tạo của N.I. Học thuyết lựa chọn hiện đại của Vavilov
Nghiên cứu có hệ thống về tài nguyên thực vật trên thế giới đối với các loại cây trồng quan trọng nhất đã thay đổi hoàn toàn sự hiểu biết về thành phần giống và loài của ngay cả những loại cây trồng được nghiên cứu kỹ lưỡng như lúa mì, lúa mạch đen, ngô, bông, đậu Hà Lan, hạt lanh và khoai tây. Trong số các loài và nhiều giống cây trồng này được mang về từ các cuộc thám hiểm, gần một nửa là loài mới, chưa được khoa học biết đến. Việc phát hiện ra các loài và giống khoai tây mới đã thay đổi hoàn toàn cách hiểu trước đây về nguồn nguyên liệu để lựa chọn. Dựa trên tài liệu được thu thập bởi các cuộc thám hiểm của N.I. Vavilov và các cộng tác viên của ông đã thành lập toàn bộ quá trình tuyển chọn bông và xây dựng sự phát triển của các vùng cận nhiệt đới ẩm ở Liên Xô.
Dựa trên kết quả nghiên cứu chi tiết và lâu dài về sự phong phú về chủng loại được thu thập bởi các cuộc thám hiểm, các bản đồ khác biệt về vị trí địa lý của các giống lúa mì, yến mạch, lúa mạch, lúa mạch đen, ngô, kê, lanh, đậu Hà Lan, đậu lăng, đậu, đậu, đậu xanh, đậu xanh, khoai tây và các loại cây khác đã được tổng hợp. Trên những bản đồ này, người ta có thể thấy nơi tập trung sự đa dạng về giống chính của các loài thực vật được đặt tên, tức là nơi cần lấy nguyên liệu nguồn để chọn lọc một loại cây trồng nhất định. Ngay cả đối với những loài thực vật cổ xưa như lúa mì, lúa mạch, ngô và bông, vốn đã lan rộng khắp toàn cầu từ lâu, vẫn có thể xác định với độ chính xác cao về các lĩnh vực chính có tiềm năng loài chính. Ngoài ra, người ta còn xác định rằng các khu vực hình thành sơ cấp trùng khớp với nhiều loài và thậm chí cả chi. Nghiên cứu địa lý đã dẫn đến việc hình thành toàn bộ hệ thực vật độc lập về mặt văn hóa cụ thể cho từng khu vực.
Việc nghiên cứu tài nguyên thực vật trên thế giới đã cho phép N.I. Vavilov đã làm chủ hoàn toàn nguồn nguyên liệu cho công tác nhân giống ở nước ta, đồng thời ông đã đặt lại và giải quyết vấn đề nguồn nguyên liệu cho nghiên cứu di truyền và chọn lọc. Ông đã phát triển các cơ sở khoa học về chọn lọc: học thuyết về nguồn nguyên liệu, cơ sở thực vật và địa lý của kiến thức về thực vật, các phương pháp chọn lọc các đặc điểm kinh tế liên quan đến lai tạo, ươm tạo, v.v., tầm quan trọng của lai xa giữa các loài và giữa các giống. Tất cả những công trình này không hề mất đi ý nghĩa khoa học và thực tiễn ở thời điểm hiện tại.
Nghiên cứu thực vật và địa lý của một số lượng lớn cây trồng đã dẫn tới sự phân loại cùng loài của cây trồng, dẫn đến các công trình của N.I. Vavilov “Các loài Linnaean như một hệ thống” và “Học thuyết về nguồn gốc của cây trồng sau Darwin.”
Sự biến đổi kiểu gen. Phân loại đột biến.
Sửa đổi độ biến thiên, định mức phản ứng, mô hình thống kê.
Vai trò của kiểu gen và điều kiện môi trường trong việc hình thành kiểu hình.
Tính biến đổi như một đặc tính của sinh vật. Các loại biến thiên.
Di truyền tế bào chất.
Tính biến đổi là một đặc tính của sinh vật, thể hiện ở khả năng có được những đặc tính mới hoặc mất đi những đặc tính cũ. Có hai loại biến dị: kiểu hình (không di truyền) và kiểu gen (di truyền).
Các dạng biến đổi kiểu hình:
Bản thể (tuổi);
Sửa đổi.
Các dạng biến đổi kiểu gen:
Kết hợp;
Đột biến.
Sự phát triển kiểu hình của sinh vật được xác định bởi sự tương tác của cơ sở di truyền - kiểu gen - với các điều kiện môi trường (xem sơ đồ).
Sự biểu hiện của một gen bị ảnh hưởng đáng kể bởi các gen khác. Khả năng phát triển một tính trạng trước hết còn phụ thuộc vào ảnh hưởng của hệ thống điều hòa của cơ thể; Nội tiết. Những dấu hiệu như vậy ở gà trống như bộ lông sáng, mồng lớn, tính chất hót và âm sắc của giọng nói là do tác động của hormone sinh dục nam. Hormon giới tính cái được tiêm cho gà trống gây ra hoạt động của các gen quyết định sự tổng hợp trong gan các protein tạo nên lòng đỏ trứng. Thông thường, những gen này chỉ “hoạt động” ở gà. Do đó, môi trường bên trong sinh vật cũng ảnh hưởng rất lớn đến sự biểu hiện của gen dưới dạng một tính trạng.
Mỗi sinh vật phát triển và sống trong những điều kiện bên ngoài nhất định, chịu tác động của các yếu tố môi trường - sự biến động về nhiệt độ, ánh sáng, độ ẩm, số lượng và chất lượng thức ăn và có mối quan hệ với các sinh vật khác. Tất cả những yếu tố này có thể làm thay đổi các đặc tính hình thái và sinh lý của sinh vật, tức là kiểu hình của chúng. Các dấu hiệu khác nhau của cơ thể thay đổi ở những mức độ khác nhau dưới tác động của môi trường bên ngoài.
Tất cả các dấu hiệu của sinh vật có thể được chia thành hai nhóm - định tính và định lượng. Định tính là những đặc điểm được thiết lập mang tính mô tả (loại hình). Đó là màu sắc của hoa, hình dạng của trái cây, màu sắc của động vật, màu mắt và sự khác biệt về giới tính. Định lượng là những tính trạng được xác định bằng phép đo (sản lượng trứng gà, sản lượng sữa bò, trọng lượng hạt lúa mì). Nhiều đặc tính định tính ít bị ảnh hưởng bởi điều kiện môi trường hơn các đặc tính định lượng. Nhưng đặc điểm chất lượng cũng có thể thay đổi. Ví dụ, nếu lông của thỏ ermine (thỏ trắng có bàn chân, đuôi và mõm đen) bị cạo ở bất kỳ bộ phận nào trên cơ thể thì màu sắc của lông mới mọc sẽ phụ thuộc vào nhiệt độ môi trường. Vì vậy, nếu bạn cạo phần lông trắng ở lưng và giữ con vật ở nhiệt độ trên +2 ° C thì lông trắng sẽ mọc ở nơi này. Ở nhiệt độ không khí dưới +2 °C, len đen sẽ phát triển thay vì trắng. Vì vậy, các tính trạng phát triển là kết quả của sự tương tác giữa kiểu gen và môi trường. Cùng một kiểu gen có thể cho các giá trị khác nhau của một tính trạng trong các điều kiện môi trường khác nhau.
Các giới hạn trong đó những thay đổi về đặc tính của một kiểu gen nhất định có thể xảy ra được gọi là chuẩn phản ứng. Một số tính trạng (ví dụ như độ sữa) có chuẩn phản ứng rất rộng, những tính trạng khác (màu lông) có chuẩn phản ứng hẹp hơn nhiều.
Di truyền không phải là một đặc điểm mà là khả năng của một sinh vật (kiểu gen của nó), do sự tương tác với các điều kiện phát triển, tạo ra một kiểu hình nhất định, nói cách khác, là chuẩn mực phản ứng của sinh vật với các điều kiện bên ngoài. được kế thừa.
Sự đa dạng về kiểu hình phát sinh ở sinh vật dưới tác động của điều kiện môi trường được gọi là tính biến đổi biến đổi. Một ví dụ kinh điển về khả năng biến đổi biến đổi là sự thay đổi hình dạng của một cây bồ công anh trồng trên đồng bằng và trên núi từ cùng một thân rễ (thí nghiệm của Bonnier, 1895). Một ví dụ là đầu mũi tên của thực vật thủy sinh, chúng tạo ra ba hình dạng lá tùy thuộc vào môi trường bên ngoài: hình dải băng (ngập nước), hình thận (nổi) và hình mũi tên (nổi lên). Sự biến đổi sửa đổi khá phổ biến trong các sinh vật sống. C. Darwin gọi đó là sự biến đổi nhất định.
Để mô tả các đặc điểm định lượng, một bộ chỉ số thống kê được sử dụng. Một trong những chỉ số này là chuỗi biến thể, đặc trưng cho tính biến đổi của một tính trạng. Giá trị trung bình của nó được tính bằng công thức:
trong đó X là tổng, v là biến thể, p là tần suất xuất hiện của biến thể, n là tổng số biến thể trong chuỗi biến thể.
Để mô tả mức độ biến đổi của một tính trạng, người ta sử dụng đường cong biến đổi. Để thực hiện điều này, hãy xây dựng biểu đồ trong hệ tọa độ, vẽ trên trục x (ngang) giá trị của tùy chọn v theo thứ tự tăng dần của chúng, trên trục tọa độ (dọc) - tần suất xuất hiện p của mỗi tùy chọn. Khi kết nối các điểm giao nhau, chúng ta thu được một đường cong mô tả tính biến đổi của tính trạng.
Sự biến đổi kiểu gen
Kiểu gen hoặc biến đổi di truyền thường được chia thành tổ hợp và đột biến. Biến dị tổ hợp là sự kết hợp của các gen trong một kiểu gen. Ví dụ: cha mẹ có nhóm máu I và II và con có nhóm máu II và III:

Việc thu được các tổ hợp gen mới trong một kiểu gen có tính biến dị tổ hợp đạt được nhờ ba quá trình:
a) sự phân kỳ độc lập của các nhiễm sắc thể trong quá trình giảm phân,
b) sự kết hợp ngẫu nhiên của chúng trong quá trình thụ tinh,
c) Tái tổ hợp gen do trao đổi chéo.
Darwin chứng minh rằng nhiều giống cây trồng và vật nuôi đã được tạo ra thông qua việc lai tạo các giống có sẵn. Ông rất coi trọng tính biến đổi tổ hợp, tin rằng cùng với sự chọn lọc, nó đóng một vai trò quan trọng trong việc tạo ra các dạng mới cả trong tự nhiên và nền kinh tế con người.
Sự biến đổi kết hợp là phổ biến trong tự nhiên. Các vi sinh vật sinh sản vô tính đã phát triển các cơ chế độc đáo (biến đổi và tải nạp) dẫn đến sự xuất hiện của biến dị tổ hợp.
Biến thể kết hợp thậm chí có thể đóng một vai trò trong sự hình thành loài. Các loài thực vật có hoa và cá được mô tả là sự kết hợp các đặc điểm của hai loài hiện có có liên quan chặt chẽ với nhau. Tuy nhiên, sự xuất hiện của các loài chỉ do lai tạo là một hiện tượng hiếm gặp.
Hiện tượng dị hợp liền kề với biến thiên tổ hợp.
Sự biến đổi đột biến không phải do sự tái tổ hợp của các gen mà do sự vi phạm cấu trúc của chúng. Charles Darwin (1859) đã nói về khả năng đột biến, gọi chúng là những biến đổi không chắc chắn hoặc những thay đổi đơn lẻ. Anh nhận thấy sự xuất hiện đột ngột của họ.
Thuật ngữ “đột biến” được G. De Friesom đề xuất vào năm 1889 để xác định những thay đổi mà ông quan sát được ở rừng dương. Ông nhận thấy loài cây này thường xuyên trải qua những thay đổi. Như vậy, một đột biến có lá và quả có gân đỏ; loại còn lại có lá rộng hơn đáng kể so với dạng ban đầu, chỉ có hoa cái,
và không lưỡng tính; dị nhân thứ ba có kích thước lùn; cây thứ tư cao, có hoa, quả và hạt to.
De Vries, trong tác phẩm “Lý thuyết đột biến” (1901 - 1903), đã đưa ra những quy định chính của lý thuyết đột biến:
Đột biến xảy ra đột ngột.
Các hình thức mới khá ổn định.
Đột biến là những thay đổi về chất.
Đột biến có thể có lợi hoặc có hại.
Những đột biến tương tự có thể xảy ra nhiều lần.
Phân loại đột biến (xem bảng)
Vai trò chính thuộc về các đột biến thế hệ xảy ra trong tế bào mầm. Nếu đột biến chiếm ưu thế thì một đặc điểm hoặc đặc tính mới sẽ xuất hiện ngay cả ở một cá thể dị hợp tử. Nếu đột biến là lặn thì nó chỉ có thể xuất hiện sau vài thế hệ khi trở thành đồng hợp tử. Một ví dụ về đột biến gen trội ở người là biểu hiện phồng rộp da ở bàn chân, đục thủy tinh thể ở mắt và chứng đau ngón tay ngắn (ngón tay ngắn bị thiếu các đốt ngón tay). Một ví dụ về đột biến gen lặn tự phát ở người là bệnh máu khó đông ở từng gia đình.
Đột biến soma là đột biến xảy ra ở tế bào soma; chỉ được bảo tồn ở con cái thông qua quá trình nhân giống sinh dưỡng (sự xuất hiện của một cành có quả mọng màu trắng trên bụi nho đen, một sợi tóc trắng và các màu mắt khác nhau ở người). Sự kế thừa các đột biến soma hiện đang trở nên quan trọng trong việc nghiên cứu nguyên nhân gây ung thư ở người. Người ta cho rằng trong các khối u ác tính, sự biến đổi của một tế bào bình thường thành tế bào ung thư xảy ra tùy theo loại đột biến soma.
Đột biến gen hay đột biến điểm là loại đột biến phổ biến nhất. Đây là những thay đổi vô hình về mặt tế bào học trong nhiễm sắc thể. Đột biến gen có thể là gen trội hoặc gen lặn. Một ví dụ về đột biến gen ở người là bệnh còi xương kháng vitamin D, bệnh rối loạn chuyển hóa axit amin phenylalanine, v.v..
Cơ chế phân tử của đột biến gen được thể hiện ở sự thay đổi thứ tự các cặp nucleotide trong phân tử axit nucleic. Bản chất của những thay đổi bên trong có thể được quy về bốn loại sắp xếp lại nucleotide: a) thay thế một cặp bazơ trong phân tử DNA; b) mất (mất) một cặp hoặc một nhóm bazơ trong phân tử ADN; "
c) chèn một cặp hoặc một nhóm cặp bazơ vào phân tử ADN;
d) sắp xếp lại vị trí các nucleotit trong gen.
Những thay đổi trong cấu trúc phân tử của gen dẫn đến các hình thức sao chép mới từ gen đó thông tin di truyền cần thiết cho sự xuất hiện của các quá trình hóa học trong tế bào, cuối cùng dẫn đến sự xuất hiện các đặc tính mới trong toàn bộ tế bào và sinh vật. Đột biến điểm là quan trọng nhất đối với sự tiến hóa. Tùy theo ảnh hưởng của chúng đến bản chất của các polypeptide được mã hóa, các đột biến điểm có thể được chia thành ba loại:
1) đột biến xảy ra khi một nucleotide được thay thế trong codon, gây ra sự thay thế một axit amin không chính xác ở một vị trí nhất định trong chuỗi polypeptide. Vai trò sinh lý của protein thay đổi, tạo ra môi trường cho chọn lọc tự nhiên. Đây là loại đột biến điểm, đột biến gen chính xuất hiện thông qua quá trình gây đột biến tự nhiên dưới tác động của bức xạ và các chất gây đột biến hóa học;
2) các đột biến vô nghĩa, tức là sự xuất hiện của các codon cuối cùng trong gen do sự thay đổi các bazơ riêng lẻ trong các codon đó. Kết quả là quá trình dịch mã kết thúc tại vị trí xuất hiện codon cuối cùng. Hóa ra gen này chỉ có khả năng mã hóa các đoạn polypeptide cho đến khi xuất hiện codon cuối cùng;
3) các đột biến đọc dịch khung xảy ra khi các phần chèn và xóa xuất hiện trong một gen và toàn bộ nội dung ngữ nghĩa của gen đó thay đổi. Điều này được gây ra bởi sự kết hợp mới của các nucleotide trong bộ ba. Kết quả là toàn bộ chuỗi polypeptide sau đột biến điểm thu được các axit amin không chính xác khác.
Sự sắp xếp lại nhiễm sắc thể thường được phân loại là đột biến, vì sự hiện diện của chúng trong tế bào có liên quan đến sự thay đổi tính chất của các tế bào này hoặc các sinh vật phát sinh từ các tế bào này. Có:
1) thiếu một phần nhiễm sắc thể (thiếu và mất đoạn). Xóa là sự mất (thiếu) phần giữa của nhiễm sắc thể do nó bị đứt ở hai điểm. Nếu đoạn xa, đoạn cuối bị đứt ra thì gọi là thiếu hụt. Sự thiếu hụt này rất hiếm xảy ra vì sau khi vùng ngoại vi bị phá hủy, nhiễm sắc thể không thể tồn tại được nữa. Sự thiếu hụt thường làm giảm khả năng tồn tại và khả năng sinh sản của một cá nhân;
2) nhân đôi hoặc nhân đôi một số phần nhất định của nhiễm sắc thể (nhân đôi). Một ví dụ về sự nhân đôi là việc củng cố đặc điểm Vag (mắt hình dải) ở Drosophila với sự gia tăng số lượng gen kiểm soát nó. Hiện tượng nhân đôi tương đối phổ biến trong tự nhiên và nó có một vai trò tiến hóa nhất định;
3) sự thay đổi trong cách sắp xếp tuyến tính của các gen trong nhiễm sắc thể do sự đảo ngược 180° của các phần riêng lẻ của nhiễm sắc thể (đảo ngược). Một ví dụ thú vị về sự đảo ngược là sự khác biệt về bộ nhiễm sắc thể ở họ mèo. Tất cả các đại diện của nó đều có 36 nhiễm sắc thể, nhưng kiểu nhân của các loài khác nhau khác nhau ở chỗ có sự đảo ngược ở các nhiễm sắc thể khác nhau. Sự đảo ngược dẫn đến những thay đổi về một số đặc điểm hình thái và sinh lý của sinh vật và có thể là một yếu tố dẫn đến sự cô lập sinh học của quần thể;
4) chèn - sự di chuyển của các đoạn nhiễm sắc thể dọc theo chiều dài của nó, thay thế sự định vị của gen.
Sự sắp xếp lại giữa các nhiễm sắc thể có liên quan đến sự trao đổi vùng giữa các nhiễm sắc thể không tương đồng. Sự sắp xếp lại như vậy được gọi là chuyển vị. Sự chuyển vị nghiêm trọng có thể dẫn đến giảm mạnh khả năng tồn tại của tế bào và toàn bộ sinh vật.
Đột biến ảnh hưởng đến bộ gen của tế bào được gọi là bộ gen. Sự thay đổi số lượng nhiễm sắc thể trong bộ gen có thể xảy ra do sự tăng hoặc giảm số lượng bộ đơn bội hoặc số lượng nhiễm sắc thể riêng lẻ. Các sinh vật trong đó toàn bộ bộ đơn bội đã nhân lên được gọi là đa bội. Các sinh vật trong đó số lượng nhiễm sắc thể không phải là bội số của số đơn bội được gọi là thể lệch bội hoặc dị bội.
Thể đa bội là một đột biến gen bao gồm sự gia tăng số lượng nhiễm sắc thể là bội số của số lượng đơn bội. Các tế bào có số lượng bộ nhiễm sắc thể đơn bội khác nhau được gọi là: tam bội 3G; 4d-tứ bội, v.v. Thể đa bội dẫn đến những thay đổi về đặc điểm của sinh vật: tế bào lớn và khả năng sinh sản tăng lên. Ngược lại, đa bội được chia thành autopolyploidy (sự gia tăng số lượng nhiễm sắc thể do sự nhân lên của bộ gen của một loài) và allopolyploidy (sự gia tăng số lượng nhiễm sắc thể do sự hợp nhất của bộ gen của các loài khác nhau). Thể đa bội được biết đến ở động vật (ciliates, giun tròn, động vật giáp xác thủy sinh, tằm, lưỡng cư). Aneuploidy, hay dị thể, là sự thay đổi về số lượng nhiễm sắc thể không phải là bội số của bộ nhiễm sắc thể đơn bội (ví dụ: 2n+\,2n- 1, 2n - 2, 2n + 2). Ở người, đây là hội chứng trisomy trên nhiễm sắc thể X hoặc trên nhiễm sắc thể 21 (hội chứng Down), monosomy trên nhiễm sắc thể X, v.v. Hiện tượng lệch bội cho thấy sự vi phạm số lượng nhiễm sắc thể dẫn đến thay đổi cấu trúc và khả năng sống sót của cơ thể.
Sự thay đổi về plasmogens (bất kỳ đơn vị di truyền tế bào chất nào tương ứng với di truyền nhiễm sắc thể), dẫn đến sự thay đổi về đặc điểm và tính chất của sinh vật, được gọi là đột biến tế bào chất. Những đột biến này ổn định và được truyền từ thế hệ này sang thế hệ khác (mất cytochrome oxidase trong ty thể của nấm men).
Theo giá trị thích ứng, đột biến có thể được chia thành đột biến có lợi, có hại (gây chết người và bán gây chết người) và trung tính. Sự phân chia này có điều kiện. Ví dụ về các đột biến gây chết người và bán gây chết người ở người: epiloia (một hội chứng đặc trưng bởi sự phát triển của da, chậm phát triển trí tuệ) và động kinh, cũng như sự hiện diện của các khối u ở tim, thận, chứng ngu ngốc (lắng đọng chất béo trong hệ thần kinh trung ương). hệ thống, kèm theo thoái hóa chất não, mù lòa).
Đột biến xảy ra tự nhiên mà không có sự tiếp xúc đặc biệt với các tác nhân bất thường được gọi là tự phát. Các mô hình đột biến tự phát có những điểm sau:
1. Mức độ đột biến tự phát ở hầu hết các sinh vật sống là thấp, nhưng tần suất đột biến khác nhau giữa các loài động vật và thực vật khác nhau.
2. Các gen khác nhau trong cùng một kiểu gen đột biến ở tần số khác nhau.
3. Các gen giống nhau ở các kiểu gen khác nhau đột biến với tốc độ khác nhau. Ví dụ, ở các chủng Drosophila trong phòng thí nghiệm khác nhau, tần suất đột biến ở mắt và cánh là không giống nhau; Đột biến bệnh bạch tạng phổ biến hơn ở loài gặm nhấm so với các động vật có vú khác.
4. Các loài và chi gần nhau về mặt di truyền được đặc trưng bởi chuỗi biến dị di truyền tương tự nhau với mức độ đều đặn đến mức khi biết chuỗi các dạng trong một loài, người ta có thể dự đoán sự hiện diện của các dạng song song ở các loài và chi khác. Các chi và loài càng nằm gần nhau trong sơ đồ chung thì sự tương đồng về chuỗi biến đổi của chúng càng hoàn thiện. “Toàn bộ họ thực vật thường được đặc trưng bởi một chu kỳ biến đổi nhất định trải qua tất cả các chi và loài tạo nên họ.” Sự đều đặn cuối cùng được N.I. Vavilov phát hiện vào năm 1920. Ông chỉ ra rằng chuỗi tương đồng thường vượt ra ngoài phạm vi chi và thậm chí cả họ. Ngón tay ngắn đã được quan sát thấy ở đại diện của nhiều loài động vật có vú: gia súc, cừu, chó, con người. Bệnh bạch tạng được quan sát thấy ở tất cả các lớp động vật có xương sống. Quy luật chuỗi tương đồng liên quan trực tiếp đến việc nghiên cứu các bệnh di truyền ở người. Nhiều đột biến được tìm thấy ở động vật có thể đóng vai trò là mô hình cho các bệnh di truyền ở người. Như vậy, chó mắc bệnh máu khó đông liên quan đến giới tính. Bệnh bạch tạng đã được ghi nhận ở nhiều loài gặm nhấm, mèo, chó và chim. Chuột, gia súc và ngựa có thể dùng làm mô hình để nghiên cứu chứng loạn dưỡng cơ; bệnh động kinh - thỏ, chuột cống, chuột nhắt; bệnh điếc di truyền tồn tại ở chuột lang, chuột và chó. Chuột mắc các bệnh chuyển hóa di truyền (béo phì, tiểu đường).
Quá trình đột biến cảm ứng được hiểu là sự xuất hiện những biến đổi di truyền dưới sự tác động đặc biệt của các yếu tố môi trường. Tất cả các yếu tố gây đột biến có thể được chia thành ba loại: vật lý, hóa học và sinh học. Tác nhân gây đột biến vật lý hiệu quả nhất là bức xạ ion hóa. Đi qua tế bào, tia X, tia gamma, hạt alpha và các bức xạ ion hóa khác dọc theo đường đi của chúng sẽ đánh bật các electron khỏi lớp vỏ ngoài của nguyên tử hoặc phân tử, biến chúng thành các hạt tích điện dương. Bức xạ ion hóa có thể vừa tác động trực tiếp lên DNA vừa tác động gián tiếp thông qua các phân tử ion hóa và nguyên tử của các chất khác. Liều bức xạ được đo bằng roentgen hoặc rads - những giá trị gần với giá trị tuyệt đối. Tần số đột biến phụ thuộc rất nhiều vào liều bức xạ và tỷ lệ thuận với nó.
Đột biến gây ra? gây ra bởi chiếu xạ lần đầu tiên được thực hiện bởi các nhà khoa học Liên Xô G. A. Nadson và G. S. Filippov, người vào năm 1925 đã quan sát quá trình đột biến ở nấm men sau khi tiếp xúc với bức xạ ion hóa. Năm 1927, nhà di truyền học người Mỹ G. Meller đã chỉ ra rằng tia X có thể gây ra nhiều đột biến ở Drosophila, và sau đó tác động gây đột biến của tia X đã được xác nhận trên nhiều vật thể.
Bức xạ tia cực tím cũng thuộc về chất gây đột biến vật lý. Tuy nhiên, tác dụng gây đột biến của nó ít hơn đáng kể so với bức xạ ion hóa. Nhiệt độ tăng thậm chí còn có tác động yếu hơn, điều này đối với động vật máu nóng và con người hầu như không có ý nghĩa đáng kể do nhiệt độ cơ thể của chúng không đổi."
Nhóm yếu tố thứ hai là các tác nhân gây đột biến hóa học. Các chất gây đột biến hóa học chủ yếu gây ra đột biến điểm hoặc đột biến gen, trái ngược với các chất gây đột biến vật lý, làm tăng đáng kể khả năng xảy ra đột biến nhiễm sắc thể. Ưu tiên phát hiện ra chất gây đột biến hóa học thuộc về các nhà nghiên cứu Liên Xô. Năm 1933 V.V. Sakharov nhận được đột biến nhờ tác dụng của iốt, vào năm 1934 M.E. Lobashev - sử dụng amoni. Năm 1946, nhà di truyền học Liên Xô I. A. Rappoport đã phát hiện ra tác động gây đột biến mạnh mẽ của formalin và ethyleneimine, còn nhà nghiên cứu người Anh S. Auerbach đã phát hiện ra tác động gây đột biến mạnh mẽ của khí mù tạt.
Tác nhân gây đột biến sinh học bao gồm virus và độc tố của một số sinh vật, đặc biệt là nấm mốc. Năm 1958, nhà di truyền học Liên Xô S.I. Alikhanyan đã chỉ ra rằng virus gây đột biến ở xạ khuẩn. Hơn nữa, trong các phòng thí nghiệm trong và ngoài nước, người ta đã xác định rằng một số đột biến được nghiên cứu kỹ lưỡng ở động vật, thực vật và con người là kết quả của hoạt động của virus.
Không phải mọi tổn thương DNA xảy ra nhất thiết đều biến thành đột biến, sự điều chỉnh thường xảy ra với sự trợ giúp của các enzyme đặc biệt. Quá trình này được gọi là sửa chữa.
Hiện tại, ba cơ chế sửa chữa đã được biết đến: kích hoạt quang học, sửa chữa tối và sửa chữa sau sao chép. Quá trình kích hoạt quang học liên quan đến việc loại bỏ các chất làm mờ thymine, đặc biệt phổ biến trong DNA dưới tác động của tia UV, sử dụng ánh sáng khả kiến. Đây là một quá trình enzyme. Enzym sử dụng năng lượng ánh sáng sẽ phân tách các dimer và do đó phục hồi tổn thương do tia cực tím trong DNA của thể thực khuẩn và vi khuẩn. Sửa chữa tối không cần ánh sáng. Nó có khả năng sửa chữa nhiều loại tổn thương DNA. Sửa chữa tối xảy ra trong một số giai đoạn với sự tham gia của một số enzyme:
Các phân tử của enzyme đầu tiên (endonuclease) liên tục kiểm tra phân tử DNA, nhận ra sự hư hỏng, enzyme sẽ cắt chuỗi DNA gần nó;
Một enzyme khác (cũng là endnuclease hoặc exonuclease) thực hiện vết cắt thứ hai trên sợi này, cắt bỏ vùng bị tổn thương;
Enzim thứ ba (exonuclease) mở rộng đáng kể khoảng cách thu được, cắt bỏ hàng chục hoặc hàng trăm nucleotide;
Enzym thứ tư (polymerase) sửa chữa khoảng trống theo thứ tự các nucleotide trong chuỗi DNA thứ hai (không bị hư hại).
Quá trình sửa chữa sáng và tối được quan sát trước khi xảy ra quá trình sao chép của các phân tử bị hư hỏng. Nếu sự sao chép của các phân tử bị hư hỏng xảy ra thì các phân tử con có thể được sửa chữa sau quá trình sao chép. Cơ chế của nó vẫn chưa rõ ràng. Hiện tượng sửa chữa DNA phổ biến từ vi khuẩn sang người và rất quan trọng để duy trì sự ổn định của thông tin di truyền được truyền từ thế hệ này sang thế hệ khác.
Di truyền tế bào chất
Di truyền, trong đó cơ sở vật chất của sự di truyền là các yếu tố của tế bào chất, được gọi là không nhiễm sắc thể hoặc tế bào chất. Vì ở động vật và thực vật, tế bào trứng chứ không phải tinh trùng rất giàu tế bào chất, nên sự di truyền tế bào chất, không giống như di truyền nhiễm sắc thể, phải xảy ra thông qua dòng mẹ. Người sáng lập ra di truyền tế bào chất là nhà di truyền học người Đức K. Correns và Z. B aur. Các yếu tố di truyền khu trú trong tế bào chất và các bào quan của nó được gọi là plasmatype.
Di truyền ngoài nhân được chia thành:
I. Thực chất là di truyền không nhiễm sắc thể hoặc tế bào chất:
Kế thừa plastid;
Di truyền qua ty thể;
Vô sinh nam tế bào chất.
II. Xác định trước tế bào chất.
III. Di truyền thông qua nhiễm trùng hoặc thể vùi (di truyền giả tế bào chất).
1. Kiểu hình phụ thuộc vào:
a) từ kiểu gen;
b) từ môi trường;
c) không phụ thuộc vào bất cứ điều gì;
d) Do kiểu gen và môi trường.
2. Họ có chuẩn mực phản ứng hẹp... dấu hiệu.
Chất lượng;
b) định lượng.
3. Dấu hiệu của sự biến thiên được thể hiện dưới dạng chuỗi biến thiên và đường cong biến thiên:
a) đột biến;
b) sửa đổi;
c) tổ hợp?
4. Xảy ra đột biến:
a) khi băng qua đường;
b) khi vượt qua;
c) đột ngột trong DNA hoặc nhiễm sắc thể.
5. Đột biến:
a) luôn lặn;
b) luôn chiếm ưu thế;
c) Tính trạng trội hoặc lặn.
6. Đột biến biểu hiện về kiểu hình:
a) trong mọi trường hợp;
b) ở sinh vật đồng hợp tử;
c) Ở cơ thể dị hợp tử.
7. Theo giá trị thích nghi của đột biến, chúng có thể là:
a) cơ thể;
b) bán gây chết người, gây chết người;
c) gen hoặc điểm.
8. Đột biến vô nghĩa là:
a) đột biến xảy ra khi một nucleotide được thay thế trong codon;
b) sự xuất hiện của các codon cuối cùng trong gen;
c) đọc các đột biến dịch khung.
9. Khối u ác tính có thể do:
a) virus;
b) hóa chất;
c) bức xạ ion hóa;
d) virus, hóa chất và bức xạ ion hóa. _
10. Sự di truyền qua tế bào chất được thực hiện bằng:
a) dòng mẹ;
b) dòng họ nội.
Văn học
1. R.G. Zayats, I.V. Rachkovskaya và cộng sự Sinh học dành cho người nộp đơn. Minsk, Unipress, 2009, tr. 578-597.
2. L.N. Pesetskaya. Sinh vật học. Minsk, “Aversev”, 2007, trang 23-35.
3. ND Lisov, N.A. Lemeza và cộng sự Sinh học. Minsk, “Aversev”, 2009, trang 33-37.
4. E.I. Shepelevich, V.M. Glushko, T.V. Maksimova. Sinh học cho học sinh và người nộp đơn. Minsk, “UniversalPress”, 2007, trang 37-50.
BÀI 16. Tuyển chọn thực vật, động vật và vi sinh vật
Đối tượng và mục đích tuyển chọn.
Vavilov N.I. về nguồn gốc của cây trồng.
Các phương pháp tuyển chọn cơ bản Heterosis, ứng dụng của nó trong chăn nuôi.
Phương pháp chọn lọc và di truyền I. V. Michurina.
Thành tựu trong chăn nuôi cây trồng, vật nuôi. Các giống cây trồng được tạo ra bởi các nhà lai tạo Bêlarut.
Các hướng chính là công nghệ sinh học (công nghiệp vi sinh, kỹ thuật di truyền và tế bào).
Chọn lọc (từ tiếng Latin selectio - chọn lọc) là khoa học về các phương pháp tạo ra mới và cải tiến các giống cây trồng, giống vật nuôi và các chủng vi sinh vật mới được con người sử dụng hiện có.
Nhân giống một giống hoặc giống mới là một quá trình phức tạp gồm nhiều giai đoạn, bao gồm việc lựa chọn cẩn thận các cặp bố mẹ, việc lai tạo của chúng, chọn lọc có phương pháp ở con lai, sau đó là lai các dạng đã chọn và một lần nữa để chọn lọc. Về vấn đề này, khoa học chăn nuôi hiện đại bao gồm các phần chính sau:
1. Nghiên cứu nguồn nguyên liệu - đa dạng giống, giống, loài thực vật, động vật và vi sinh vật.
2. Phát triển các phương pháp và hệ thống lai dựa trên mô hình di truyền các tính trạng và tính biến dị của chúng.
3. Phát triển phương pháp chọn lọc nhân tạo.
Giống, giống hoặc chủng là một quần thể sinh vật được con người tạo ra một cách nhân tạo và có những đặc điểm di truyền nhất định. Tất cả các cá thể trong một giống, giống hoặc chủng đều có những đặc tính tương tự, cố định về mặt di truyền, cũng như cùng một kiểu phản ứng với các yếu tố môi trường. Ví dụ, gà Leghorn có trọng lượng nhỏ nhưng sản lượng trứng cao.
Mục tiêu của chăn nuôi hiện đại là tăng năng suất các giống cây trồng, giống vật nuôi và các chủng vi sinh vật. Gắn với quá trình công nghiệp hóa, cơ giới hóa nông nghiệp, việc chọn lọc nhằm tạo ra các giống ngũ cốc, giống nho, giống nho, cà chua, giống chè, bông thích hợp cho việc thu hoạch bằng máy; nhân giống các loại rau trồng trong nhà kính, thủy canh; tạo ra các giống vật nuôi để nuôi trong các tổ hợp chăn nuôi lớn.
Việc cải tiến giống cây trồng, giống vật nuôi, các chủng vi sinh vật là không thể nếu không nghiên cứu nguồn gốc và quá trình tiến hóa của chúng. Về vấn đề này, công trình của N.I.Vavilov về nghiên cứu trung tâm nguồn gốc cây trồng có tầm quan trọng đặc biệt. Vavilov đã tổ chức một số cuộc thám hiểm, nơi ông thu thập những tài liệu có giá trị nhất về sự phân bố của các loại cây trồng trên toàn cầu. Vavilov xác định rằng nơi sinh ra ngô là Mexico và Trung Mỹ, còn nơi sinh ra khoai tây là Nam Mỹ. Ở Afghanistan, ông đã tìm thấy nhiều loại lúa mì mềm và ở Ethiopia - lúa mì cứng. Ông đã phát hiện và mô tả 8 trung tâm nguồn gốc cây trồng:
Ấn Độ (nhiệt đới Nam Á) - trung tâm xuất xứ các giống lúa, mía, trái cây có múi;
Trung Á - lúa mì mềm, các loại đậu và các loại cây trồng khác;
Trung Quốc (hoặc Đông Á) - kê, kiều mạch, đậu nành, ngũ cốc;
Tây Á - lúa mì và lúa mạch đen, cũng như trồng cây ăn quả;
Địa Trung Hải - ô liu, cỏ ba lá, đậu lăng, bắp cải, cây thức ăn gia súc;
Abyssinian - lúa miến, lúa mì, lúa mạch;
Nam Mexico - bông, ngô, ca cao, bí ngô, đậu;
Nam Mỹ - trung tâm của khoai tây, cây thuốc (cây coca, cây cinchona).
Kiến thức về các điều kiện tự nhiên ở quê hương của một loại cây cụ thể giúp loại trừ một số hướng vô nghĩa trong quá trình chọn lọc và giúp thực hiện chọn lọc trên cơ sở khoa học, có tính đến các đặc tính của cả bản thân cây đó và ảnh hưởng của môi trường. các yếu tố hình thành chúng. Giá trị công việc của Vavilov nằm ở chỗ ông đã đặt nền móng cho việc thu thập và lưu trữ các nguồn gen thực vật và tạo ra bộ sưu tập gen đầu tiên trên thế giới tại Viện Thực vật All-Union ở Leningrad. Bộ sưu tập này được bổ sung hàng năm với các mẫu mới đến từ tất cả các châu lục, và ngay cả trong cuộc phong tỏa Leningrad, nó vẫn được bảo tồn hoàn toàn. Hiện tại, nó cung cấp nguồn nguyên liệu cho các nhà chăn nuôi trên toàn thế giới.
Các phương pháp tuyển chọn cơ bản
| phương pháp | Chăn nuôi | Nhân giống cây trồng |
| Lựa chọn cặp bố mẹ | Bởi những đặc điểm có giá trị kinh tế và bởi bên ngoài (một tập hợp các đặc điểm kiểu hình) | Theo nơi xuất xứ (xa về mặt địa lý) hoặc xa về mặt di truyền (không liên quan) |
| Lai tạo: a) không liên quan (giao phối cận huyết) | Lai các giống xa có đặc điểm tương phản để thu được quần thể dị hợp tử và biểu hiện dị hợp tử. Kết quả là con cái bị vô sinh. | Sự lai chéo giữa các loài, giữa các loài, giữa các loài dẫn đến dị hợp để thu được các quần thể dị chủng, cũng như năng suất cao |
| b) quan hệ gần gũi (cận huyết) | Lai giữa các họ hàng gần để tạo ra các dòng đồng hợp tử (thuần chủng) có các tính trạng mong muốn | Tự thụ phấn ở cây thụ phấn chéo nhờ tác động nhân tạo để thu được dòng đồng hợp tử (thuần chủng) |
| Lựa chọn a) khối lượng | Không áp dụng | Thích hợp cho cây thụ phấn chéo |
| b) cá nhân | Lựa chọn cá thể cứng nhắc được áp dụng cho các đặc điểm, độ bền và ngoại hình có giá trị kinh tế. | Dùng cho cây tự thụ phấn, các dòng thuần chủng được phân lập - con của một cá thể tự thụ phấn |
| Phương pháp kiểm tra đực giống qua con cái | Họ sử dụng phương pháp thụ tinh nhân tạo từ những con đực đực tốt nhất, chất lượng của phương pháp này đã được kiểm tra bởi rất nhiều con cái. | Không áp dụng |
| Thí nghiệm sản xuất các thể đa bội ở | Không áp dụng | Được sử dụng trong di truyền và nhân giống để thu được các hình thức năng suất cao hơn |
Heterosis, ứng dụng của nó trong chăn nuôi
Khả năng sống sót và năng suất cao hơn của các giống lai F so với các dạng lai thể hiện ý nghĩa của hiện tượng dị hợp. "
Trở lại giữa thế kỷ 18. I. Kelreuter, viện sĩ của Học viện Nga, một nhà thực vật học nổi tiếng, đã thu hút sự chú ý đến thực tế là trong một số trường hợp, khi lai thực vật, các cây lai thế hệ đầu tiên mạnh hơn nhiều so với dạng bố mẹ của chúng. Một thời kỳ mới trong nghiên cứu về hiện tượng dị hợp bắt đầu vào những năm 20. thế kỷ trước từ tác phẩm của các nhà di truyền học người Mỹ Schell, East, Hell, Johnson. Kết quả nghiên cứu của họ là các dòng lai có đặc điểm trầm cảm nặng đã thu được ở ngô thông qua quá trình tự thụ phấn. Nhưng khi Shell vượt qua những ranh giới chán nản với nhau, anh ta bất ngờ thu được những giống F lai rất mạnh. Với những công trình này, việc sử dụng rộng rãi phương pháp lai trong quá trình nhân giống đã bắt đầu."
Các nhà di truyền học đã đề xuất một số giả thuyết để giải thích dị hợp tử. Giả thuyết thống trị được phát triển bởi nhà di truyền học người Mỹ Johnson. Nó dựa trên sự công nhận các gen trội hoạt động có lợi ở trạng thái đồng hợp tử hoặc dị hợp tử:

Nếu kiểu lai chỉ có hai gen trội thì kiểu lai có bốn gen trội. Điều này quyết định tính ưu việt của giống lai, tức là những ưu điểm của nó so với các dạng ban đầu.
Giả thuyết về sự vượt trội được đề xuất bởi các nhà di truyền học người Mỹ Schell và East. Nó dựa trên sự thừa nhận rằng dị hợp tử về một hoặc nhiều gen mang lại lợi thế hơn so với đồng hợp tử về một hoặc nhiều gen.
Nhà di truyền học Liên Xô V. A. Strunnikov đã đề xuất giả thuyết về một phức hợp gen bù.
Ý nghĩa của hiện tượng dị hợp là khả năng sống sót và năng suất của các giống lai F cao hơn so với các dạng lai.
Do ảnh hưởng của dị hợp nên năng suất tăng lên tới 25-50%, giảm dị hợp ở thế hệ thứ hai và các thế hệ tiếp theo có liên quan đến việc phân chia các thế hệ con lai. Trong quá trình phát triển của thực vật và động vật, có thể quan sát thấy các hiệu ứng giống với biểu hiện dị hợp, nhưng không phải do lý do di truyền (không phải do lai tạo), mà do ảnh hưởng của một số ảnh hưởng bên ngoài. Đây được gọi là dị hợp sinh lý. Ví dụ, đối với thực vật, các trường hợp đã được thiết lập để loại bỏ hậu quả có hại của việc tự thụ phấn khi có sự thay đổi mạnh mẽ về điều kiện sinh trưởng. Bản chất của những hiệu ứng như vậy vẫn chưa rõ ràng. Có ba loại dị hợp:
Sinh sản (con lai vượt trội so với dạng bố mẹ về khả năng sinh sản);
Somatic (ở thế hệ con lai, khối lượng sinh dưỡng ở thực vật và sức mạnh hiến pháp ở động vật tăng lên);
Thích nghi (con lai thích nghi tốt hơn với điều kiện môi trường so với dạng bố mẹ).
Phương pháp chọn lọc và di truyền I. V. Michurina
| phương pháp | Bản chất của phương pháp | Ví dụ | |
| Lai xa về mặt sinh học: a) khác loài | Lai các đại diện của các loài khác nhau để thu được giống có đặc tính mong muốn | Anh đào Vladimir x anh đào trắng Winkler - Vẻ đẹp của anh đào miền Bắc (vị ngon, độ cứng mùa đông) | |
| b) liên thế hệ | Lai các đại diện của các chi khác nhau để thu được cây mới | Anh đào x anh đào chim = cerapadus | |
| Sự lai tạo xa về mặt địa lý | Vượt qua các đại diện của các vùng xa xôi về mặt địa lý để truyền cho con lai những phẩm chất cần thiết (hương vị, tính ổn định) | Lê Ussuri hoang dã x Bere royal (Pháp) = Bere mùa đông Michurina | |
| phương pháp | Bản chất của phương pháp | Ví dụ | |
| Lựa chọn | Nhiều, cứng: về kích thước, hình dạng, độ cứng trong mùa đông, đặc tính miễn dịch, chất lượng, mùi vị của trái cây và chất lượng bảo quản của chúng | Nhiều giống táo ngon, năng suất cao được quảng bá ra phía Bắc | |
| Phương pháp cố vấn | Nuôi dưỡng những phẩm chất mong muốn ở cây con lai (tăng ưu thế), cây con sẽ được giáo viên ghép vào cây mà họ muốn có được những phẩm chất này | Bellefleur Chinese (gốc ghép lai) x Chinese (cành ghép) = Bellefleur Chinese (giống dài ngày, chín muộn) | |
| Phương thức hòa giải | Trong lai xa, sử dụng loài hoang dại làm vật trung gian để khắc phục tình trạng không thể lai xa | Hạnh nhân Mông Cổ hoang x đào David hoang = hạnh nhân trung gian Đào trồng x hạnh nhân trung gian = đào lai (tiên tiến phía bắc) | |
| Tiếp xúc với điều kiện môi trường | Khi nuôi con lai non cần chú ý đến phương pháp bảo quản hạt giống, tính chất và mức độ dinh dưỡng; tiếp xúc với nhiệt độ thấp, đất nghèo dinh dưỡng, cấy ghép thường xuyên | ||
| Trộn phấn hoa | Để khắc phục sự không tương thích giữa các quốc gia (không tương thích) | Phấn hoa của cây mẹ trộn lẫn với phấn hoa của cây bố, phấn hoa của cây mẹ kích thích đầu nhụy và nó nhận biết được phấn hoa lạ. | |
Thành công của tuyển chọn Liên Xô
Những thành tựu của nhân giống cây trồng ở Liên Xô gắn liền với việc sử dụng các phương pháp di truyền kết hợp với phương pháp nhân giống. Do đó, N.V. Tsin (1898-1980) và các đồng nghiệp của ông, nhờ lai tạo và chọn lọc xa, đã phát triển các giống lai lúa mì-cỏ lúa mì có năng suất cao (lên tới 70 c/ha) có khả năng chống đổ ngã. Sử dụng phương pháp lai phức tạp của các dạng ở xa nhau về mặt địa lý, sau đó là chọn lọc từng cá thể cẩn thận qua nhiều thế hệ, P. P. Lukyanenko đã tạo ra một số giống lúa mì mùa đông đáng chú ý. Giống lúa mì Bezostaya-1 đã trở nên phổ biến. Giống này có năng suất cao (65-70 c/ha trở lên) trong nhiều điều kiện môi trường: ở Bắc Caucasus, miền nam Ukraine, Moldova, Transcaucasia, ở một số khu vực ở Trung Á và Kazakhstan, cũng như ở Hungary. , Bulgaria, Romania, Nam Tư.
P. P. Lukyanenko cũng đã tạo ra các giống lúa mì (Aurora, Caucasus), năng suất đạt gần 100 c/ha. V. N. Remeslov đã đạt được thành công lớn trong việc lựa chọn lúa mì mùa đông thích nghi với nhiều điều kiện khí hậu và địa lý tại Viện giống lúa mì Mironovsky. Giống Mironovskaya-808 mà ông nhân giống đã được phân vùng trên hầu hết toàn bộ lãnh thổ của SSR Ucraina, tại Cộng hòa Liên minh Moldavian và Belarus và ở gần 50 khu vực của RSFSR. Giống này được đặc trưng bởi năng suất cao (55-60 c/ha), độ cứng mùa đông và khả năng đáp ứng tốt với phân bón. Năm 1974, người ta thu được giống Ilyichevka (100 c/ha).
Trong việc tuyển chọn lúa mì vụ xuân, những thành công lớn nhất đã đạt được tại Viện Nghiên cứu Nông nghiệp Đông Nam Bộ (Saratov) bởi các nhà lai tạo xuất sắc A.P. Shekhurdin và V.N. Mamontova.
Giống lúa mì Saratovskaya-29 do các nhà lai tạo này nhân giống, có năng suất cao, được đặc trưng bởi chất lượng nướng đặc biệt.
V.S. Pustovoyt đã đạt được thành công đáng kể trong việc nhân giống hoa hướng dương, ông bắt đầu công việc nhân giống các giống có chứa khoảng 30% dầu trong hạt. Nhờ sử dụng phương pháp chọn lọc nhóm gia đình liên tục, ông đã có thể thu được các giống có hàm lượng dầu từ 50% trở lên. V. S. Pustovoit đã phát triển các giống hoa hướng dương Mayak và Peredovik. Nhờ phân vùng các giống này, có thể thu được thêm hàng nghìn tấn dầu hướng dương với cùng chi phí.
M.I. Xajanov ở những năm 30. thế kỷ trước đã phát hiện ra hiện tượng vô sinh nam ở tế bào chất. Dựa trên việc lai tạo và chọn lọc cây trồng có yếu tố tế bào chất, các giống ngô năng suất cao đã được phát triển.
Lutkov và Zosimovich đã làm tăng hàm lượng đường và năng suất củ cải đường nhờ tính đa bội. Người chăn nuôi cũng đạt được thành công lớn. Bằng cách lai giống, M.F. Ivanov đã tạo ra các giống có năng suất cao (giống lợn Ukraina trắng thảo nguyên, giống cừu lông mịn Askanian). Dựa trên việc lai giữa cừu argali hoang dã với cừu merino, sau đó là chọn lọc động vật thuộc loại mong muốn và sử dụng cận huyết ở Kazakhstan, N.S. Baturin và các đồng nghiệp đã phát triển giống argali-merino, kết hợp len chất lượng cao và khả năng thích ứng cao của argali. với điều kiện của đồng cỏ trên núi cao. Các phương pháp lai và chọn lọc là cơ sở cho việc tạo ra nhóm giống lợn mới “Kazakh hybrid” (Viện Sinh học Thực nghiệm của Cộng hòa Xã hội chủ nghĩa Xô viết Kazakhstan). Lợn Kemerovo và lợn rừng được sử dụng làm hình thức ban đầu trong quá trình nhân giống nhóm giống này.
Thành tựu của các nhà chăn nuôi Belarus
Các nhà khoa học của Viện nghiên cứu trồng khoai tây và rau quả Belarus (Samokhvalovichi, vùng Minsk) từ năm 1925 đến năm 1995. Khoảng 50 giống khoai tây, hơn 70 loại rau, 124 loại trái cây và 23 loại cây mọng đã được nhân giống. Dưới sự lãnh đạo và tham gia trực tiếp của Viện sĩ P.I. Alsmik, những giống khoai tây như Temp, Loshitsky, Ravaristy, Ogonyok, Sadko, Novinka, Verba, Ivushka, Lasunak, Zorka, v.v. Có 12 giống khoai tây được phân vùng ở nước cộng hòa với một tiềm năng năng suất 500 - 700 c/ha, kháng sâu bệnh, có vị ngon, thích hợp chế biến thành bán thành phẩm. A.G. Voluznev đã phát triển 23 giống cây mọng. Các loại nho đen phổ biến nhất là ngọt Belarus, Cantata, Minai Shmyrev, Pamyat Vavilov. Công chúa,
Katyusha, Đảng phái; nho đỏ - Người yêu dấu; lý gai - Mùa xuân, Shchedry; -dâu tây - Minskaya, Chaika.
124 giống cây ăn quả đã được nhân giống, trong đó có 24 giống cây táo (Antey, mâm xôi Belorusskoe, Bananovoe, Minskoe, v.v.), 8 giống lê (Beloruska, Maslyanistaya, Loshitskaya, v.v., 15 giống anh đào (Zolotaya Loshitskaya, v.v.). , Krasavitsa) và nhiều người khác. Những người sáng lập việc chọn lọc cây ăn quả của Belarus là E. P. Syubarova và A. E. Syubarov. Các nhà nhân giống táo đã tiếp tục những gì họ đã bắt đầu - G. K. K ovalenko và E. V. Semashko, cho quả lê - N. I. M ikhnevich, cho quả anh đào - R. M. Sulimova, cho mận - V. A. Matv e e v. Những người sáng lập việc lựa chọn cây rau là: G. I. Artemenko và A. M. Polyanskaya (cà chua), E. I. Chulkova (bắp cải), V. F. Devyatova (hành, tỏi). Họ đã đặt nền móng và phát triển nền tảng cho việc lựa chọn khoa học các loại cây trồng cây rau ở Belarus.
Các giống cây rau chọn lọc của Belarus đã được khoanh vùng: cà chua mặt đất mở - Peramoga, Excellent, Dokhodny, Ruzha, Neman; cà chua cho nhà kính phim - Vezha; dưa chuột - Dolzhik, Verasen, Zarnitsa; bắp cải - Rusinovka, Yubileynaya; hành tây - Hổ phách, Vetraz; tỏi - Polet, v.v.
Ngoài ra, các nhà lai tạo Bêlarut đã nhân giống và khoanh vùng nhiều loại ngũ cốc và cây họ đậu, cây công nghiệp và cây thức ăn gia súc. Tại Viện Nghiên cứu Nông nghiệp Belarus (Zhodino), N.D. Mukhiny đã nhân giống giống lúa mạch đen mùa đông Belta tứ bội. Ông là tác giả và đồng tác giả của các giống lúa mạch đen mùa đông Belorusskaya 23, Druzhba, lúa mì mùa xuân Minskaya, kiều mạch Iskra và Yubileynaya 2. Giống lúa mì mùa đông Berezina (74 c/ha) nổi bật bởi chất lượng xay và nướng bột cao . Các loại ngũ cốc sử dụng làm thức ăn gia súc bao gồm lúa mì mùa đông Nadzeya (79 c/ha). Các giống lúa mạch xuân Zazersky 85 và Zhodinsky 5 đã được chứng minh là tốt, giống lupin vàng Narochansky nổi bật bởi chất lượng cao. Năng suất hạt 27, khối lượng xanh 536 c/ha. Hàm lượng protein trong hạt là 45,8%. Các giống củ cải đường nổi tiếng nhất là Belorusskaya hạt đơn-55, đa bội Belorussky-31 (được nhân giống tại Viện Di truyền và Tế bào học của Viện Hàn lâm Khoa học Belarus phối hợp với Trạm nhân giống thực nghiệm Ganusov bằng cách lai các dạng tứ bội và lưỡng bội của củ cải đường). Năng suất bình quân các loại cây lấy củ là 410-625 c/ha, hàm lượng đường 15,3-19,5%, thu đường 56,3-99,1 c/ha.
Các lĩnh vực chính của công nghệ sinh học (vi sinh, di truyền và kỹ thuật tế bào)
Công nghệ sinh học là việc sử dụng các sinh vật sống và các quá trình sinh học trong sản xuất, tức là sản xuất các chất cần thiết cho con người sử dụng các thành tựu của vi sinh, hóa sinh và công nghệ.
Ngành công nghiệp vi sinh xuất hiện vào những năm 60. Thế kỷ XX giải quyết một số vấn đề:
1) cung cấp cho vật nuôi nguồn protein thức ăn hoàn chỉnh;
2) sử dụng công nghệ sinh học, thu được và sử dụng các enzyme (protease, amylase, pectinase);
3) sử dụng công nghệ sinh học, thu được các sản phẩm vi sinh - axit amin, kháng sinh (penicillin, cephalosporin, tetracyclines, erythromycins, streptomycin);
4) với sự trợ giúp của vi sinh vật, các nguồn năng lượng bổ sung thu được dưới dạng khí sinh học, ethanol, hydro do vi khuẩn sử dụng chất thải công nghiệp và nông nghiệp.
Kỹ thuật tế bào - phương pháp phát triển tế bào trong môi trường dinh dưỡng đặc biệt.
Nuôi cấy mô là nuôi cấy tế bào được nuôi cấy trong điều kiện vô trùng trong môi trường đặc biệt. Nuôi cấy tế bào (hoặc nuôi cấy mô) có thể phục vụ để tạo ra các chất có giá trị. Ví dụ, nuôi cấy tế bào của cây nhân sâm sẽ tạo ra dược chất giống như toàn bộ cây.
Nuôi cấy tế bào cũng được sử dụng để lai tế bào. Sử dụng một số kỹ thuật đặc biệt, có thể kết hợp các tế bào có nguồn gốc khác nhau từ các sinh vật mà việc lai tạo giới tính thông thường là không thể. Điều này mở ra một phương pháp mới về cơ bản để tạo ra các giống lai dựa trên việc kết hợp các tế bào soma chứ không phải tế bào mầm vào một hệ thống duy nhất. Người ta đã thu được các tế bào và sinh vật lai của khoai tây, cà chua, cây táo và anh đào.
Ở động vật, việc sản xuất tế bào lai cũng mở ra triển vọng mới, chủ yếu cho y học. Ví dụ, các giống lai đã thu được trong quá trình nuôi cấy giữa các tế bào ung thư (có khả năng phát triển không giới hạn) và một số tế bào máu - tế bào lympho. Tế bào lai là sự kết hợp giữa tế bào ung thư và tế bào lympho. Tế bào lympho sản xuất các chất cung cấp khả năng miễn dịch (Miễn dịch) đối với các bệnh truyền nhiễm, bao gồm cả các bệnh do virus. Sử dụng các tế bào lai như vậy, có thể thu được các dược chất quý giúp tăng khả năng chống nhiễm trùng của cơ thể.
Kỹ thuật di truyền là một tập hợp các phương pháp giúp chuyển thông tin di truyền từ sinh vật này sang sinh vật khác trong ống nghiệm. Chuyển gen giúp vượt qua các rào cản giữa các loài và chuyển các đặc điểm di truyền cá thể của sinh vật này sang sinh vật khác. Mục tiêu của kỹ thuật di truyền là "thu được các tế bào (vi khuẩn) có khả năng xử lý một số protein "của con người" ở quy mô công nghiệp. Do đó, kể từ năm 1980, hormone tăng trưởng của con người - somatotropin - được lấy từ E. coli. Somatotropin là phương pháp điều trị duy nhất cho trẻ em mắc bệnh lùn do thiếu hormone này. Trước khi kỹ thuật di truyền phát triển, nó được phân lập từ tuyến yên của xác chết.
Từ năm 1982, insulin để điều trị bệnh tiểu đường đã được sản xuất ở quy mô công nghiệp từ vi khuẩn E. coli có chứa gen insulin của người. Trước đó, loại thuốc này không được cung cấp cho tất cả bệnh nhân.
Phương pháp phổ biến nhất của kỹ thuật di truyền là phương pháp thu được tái tổ hợp, tức là chứa gen ngoại lai, plasmid. Plasmid là các phân tử DNA sợi đôi hình tròn. Mỗi vi khuẩn, ngoài DNA cơ bản của nó, còn chứa một số plasmid khác nhau mà nó trao đổi với các vi khuẩn khác. Plasmid mang gen kháng thuốc ở vi khuẩn. Plasmid được các kỹ sư di truyền sử dụng để đưa gen từ sinh vật bậc cao vào tế bào vi khuẩn.
Công cụ của kỹ thuật di truyền là các enzyme được phát hiện vào năm 1974 - endonuclease hạn chế, hay enzyme hạn chế (nghĩa đen là hạn chế). Các enzyme hạn chế nhận biết các vị trí (vị trí nhận biết) và tạo ra các điểm đứt đối xứng nằm xiên với nhau trong các chuỗi DNA. Kết quả là, các “đuôi” chuỗi đơn ngắn gọi là đầu “dính” được hình thành ở đầu mỗi đoạn DNA bị hạn chế.
Các phương pháp kỹ thuật di truyền. Để thu được plasmid tái tổ hợp, một gen ngoại lai (ví dụ: gen người) được đưa vào plasmid đã phân tách. Enzim ligase nối cả hai đoạn DNA lại với nhau để tạo thành một plasmid tái tổ hợp, được đưa vào E. coli. Tất cả hậu duệ của vi khuẩn này được gọi là bản sao.
Toàn bộ quá trình thu được những vi khuẩn như vậy, được gọi là nhân bản, bao gồm các giai đoạn kế tiếp:
1. Hạn chế - cắt DNA của con người bằng enzyme giới hạn thành các đoạn có đầu “dính”.
2. Thắt - đưa các đoạn DNA của con người vào các plasmid do "khâu các đầu dính" bằng enzyme ligase.
3. Biến nạp - đưa plasmid tái tổ hợp vào tế bào vi khuẩn. Tuy nhiên, các plasmid chỉ xâm nhập vào một phần vi khuẩn được xử lý. Vi khuẩn biến nạp cùng với plasmid có khả năng đề kháng với một loại kháng sinh cụ thể. Điều này cho phép chúng được tách ra khỏi những cá thể không biến đổi gen chết trên môi trường chứa kháng sinh. Vi khuẩn được gieo trên môi trường dinh dưỡng và mỗi vi khuẩn biến đổi sẽ nhân lên và tạo thành một đàn gồm hàng nghìn con cháu - một bản sao.
4. Sàng lọc - lựa chọn giữa các dòng vi khuẩn biến nạp có chứa plasmid với gen người mong muốn. Các khuẩn lạc vi khuẩn được bao phủ bởi một bộ lọc đặc biệt, khi nó được loại bỏ, dấu vết của các khuẩn lạc vẫn còn trên đó. Quá trình lai phân tử sau đó được thực hiện. Các bộ lọc được ngâm trong dung dịch có chứa đầu dò được đánh dấu phóng xạ. Đầu dò là một polynucleotide bổ sung cho một phần của gen mong muốn. Nó chỉ lai với những plasmid tái tổ hợp có chứa gen mong muốn. Sau khi lai, phim chụp ảnh tia X được đặt trên bộ lọc trong bóng tối và phát triển sau vài giờ. Vị trí của các vùng được chiếu sáng cho phép bạn tìm thấy các plasmid mang gen mong muốn.
1. Các giống lai khác loài:
a) Vô sinh;
b) được đặc trưng bởi khả năng sinh sản tăng lên;
c) luôn là nữ;
d) luôn là nam giới.
2. Phương pháp chọn lọc không làm thay đổi đặc tính di truyền của sinh vật là:
a) đa bội; b) gây đột biến nhân tạo;
c) lai tạo; d) nhân bản.
3. Quê hương của ngô và hướng dương là trung tâm xuất xứ của cây trồng (theo N. I. Vavilov):
a) Nam Á;
b) Trung Mỹ;
c) Abyssinian;
d) Địa Trung Hải.
4. Bản thân thể đa bội hay thể bội bội là:
a) sắp xếp lại nhiễm sắc thể;
b) thay đổi số lượng nhiễm sắc thể, bội số của số đơn bội;
c) thay đổi trình tự nucleotide.
5. Lần đầu tiên, người ta có thể phát triển các phương pháp khắc phục tình trạng vô sinh của các giống lai khác loài:
a) K. A. Timiryazev; b) M. F. Ivanov; c) G. D. Karpechenko; d) N. S. Butarin.
6. Các hình thức chọn lọc nhân tạo được sử dụng trong chăn nuôi:
khổng lồ;
b) cá nhân?
7. Vào thế kỷ XX. Sau đây đã được thêm vào các phương pháp lựa chọn:
a) đa bội;
b) gây đột biến nhân tạo;
c) lai tế bào;
d) đa bội, đột biến nhân tạo và lai tế bào.
8. Nhóm thực vật đồng nhất có những đặc điểm có giá trị kinh tế do con người tạo ra gọi là:
a) nhân bản;
b) giống;
c) sự đa dạng.
9. Công nghệ sinh học là:
a) việc sử dụng các sản phẩm có nguồn gốc sinh học (than bùn, than đá, dầu) để cung cấp năng lượng cho máy móc và cơ chế;
b) việc sử dụng công nghệ trong chăn nuôi và trồng trọt;
c) việc sử dụng các sinh vật sống và các quá trình sinh học trong sản xuất;
d) việc sử dụng các sinh vật sống làm mô hình khi tạo ra các cấu trúc và cơ chế khác nhau.
Văn học
1. R.G. Zayats, I.V. Rachkovskaya và cộng sự Sinh học dành cho người nộp đơn. Minsk, Unipress, 2009, tr. 674-686.
2. L.N. Pesetskaya. Sinh vật học. Minsk, “Aversev”, 2007, trang 72-85.
3. E.I. Shepelevich, V.M. Glushko, T.V. Maksimova. Sinh học cho học sinh và người nộp đơn. Minsk, “UniversalPress”, 2007, tr.95-104.
BÀI 17. Nguồn gốc và sự phát triển của sự sống trên Trái đất.
Học thuyết tiến hóa của Charles Darwin
Thuyết tiến hóa đầu tiên của J. B. Lamarck.
Điều kiện tiên quyết cho sự xuất hiện của học thuyết Darwin.
Bản chất của học thuyết tiến hóa của Charles Darwin và ảnh hưởng của nó đối với khoa học sinh học.
Lý thuyết tổng hợp về sự tiến hóa của sự sống.
Công lao tạo ra lý thuyết tổng thể đầu tiên về sự tiến hóa của thế giới hữu cơ thuộc về J. B. Lamarck (1744-1829). Những quy định chính của lý thuyết này đã được ông nêu ra trong tác phẩm “Triết học động vật học” (1809). Lamarck đưa ra các nguyên tắc sau:
Các sinh vật có thể thay đổi;
Các loài (và các hạng phân loại khác) là tạm thời và dần dần được chuyển đổi thành loài mới;
Xu hướng chung của những thay đổi lịch sử ở sinh vật là sự hoàn thiện dần dần tổ chức của chúng (sự phân cấp), động lực của nó là mong muốn tiến bộ ban đầu (do người sáng tạo khởi xướng);
Các sinh vật vốn có khả năng ứng phó nhanh chóng với những thay đổi của điều kiện bên ngoài;
Những thay đổi ở sinh vật thu được trong suốt cuộc đời để đáp ứng với các điều kiện thay đổi đều được di truyền.
Đánh giá lý thuyết của Lamarck. Công lao nổi bật của Lamarck nằm ở việc tạo ra học thuyết tiến hóa đầu tiên. Ông bác bỏ ý tưởng về tính bất biến của loài, đối lập nó với ý tưởng về khả năng thay đổi của loài. Lời dạy của ông khẳng định sự tồn tại của tiến hóa như một quá trình phát triển lịch sử từ đơn giản đến phức tạp. Lần đầu tiên, câu hỏi về các yếu tố tiến hóa được đặt ra. Lamarck tin tưởng hoàn toàn chính xác rằng điều kiện môi trường có ảnh hưởng quan trọng đến quá trình tiến hóa. Ông là một trong những người đầu tiên đánh giá chính xác tầm quan trọng của thời gian trong quá trình tiến hóa và ghi nhận khoảng thời gian phát triển phi thường của sự sống trên Trái đất: Tuy nhiên, Lamarck đã mắc sai lầm nghiêm trọng, trước hết là khi hiểu các yếu tố của quá trình tiến hóa. , suy ra chúng từ giả định? mong muốn vốn có về sự hoàn hảo trong mọi sinh vật. Ông cũng hiểu sai lý do xuất hiện của thể lực, liên kết trực tiếp chúng với ảnh hưởng của điều kiện môi trường.
Học thuyết tiến hóa của Lamarck chưa đủ thuyết phục và không nhận được sự công nhận rộng rãi của những người cùng thời với ông.
Darwin đã phác thảo những nguyên tắc cơ bản của lý thuyết này vào năm 1859 trong cuốn sách “Nguồn gốc các loài thông qua chọn lọc tự nhiên, hay việc bảo tồn các giống được ưa chuộng trong cuộc đấu tranh giành sự sống”. Tất cả 1.250 bản của cuốn sách đã được bán trong ngày đầu tiên và người ta nói rằng tác động của nó đối với suy nghĩ của con người chỉ đứng sau Kinh thánh. Sau đó, ông phát triển lý thuyết này trong các tác phẩm tiếp theo, “Những thay đổi ở động vật và thực vật dưới ảnh hưởng của việc thuần hóa” (1868) và “Nguồn gốc của con người và lựa chọn giới tính” (1871). Nhà động vật học người Anh A. Wallace (1858) đã đi đến kết luận tương tự một cách độc lập với Darwin. Cái tên “Chủ nghĩa Darwin” do T. Geksl (1860) đề xuất.
Các nguyên tắc cơ bản trong học thuyết tiến hóa của Charles Darwin:
1. Mỗi loài đều có khả năng sinh sản không giới hạn.
2. Nguồn lực quan trọng hạn chế ngăn cản sự sinh sản không giới hạn. Hầu hết các cá thể chết trong cuộc đấu tranh sinh tồn và không để lại con cháu.
3. Cái chết hay thành công trong cuộc đấu tranh sinh tồn đều có tính chọn lọc. Charles Darwin gọi sự sinh tồn và sinh sản có chọn lọc của những sinh vật thích nghi nhất là chọn lọc tự nhiên.
4. Dưới tác động của chọn lọc tự nhiên diễn ra trong những điều kiện khác nhau, các nhóm cá thể cùng loài tích lũy những đặc tính thích nghi khác nhau từ thế hệ này sang thế hệ khác. Các nhóm cá thể có được sự khác biệt đáng kể đến mức chúng biến thành loài mới (nguyên tắc phân kỳ các đặc tính).
Dựa trên lời dạy của Darwin, người ta xác định rằng động lực tiến hóa của thế giới hữu cơ là cuộc đấu tranh sinh tồn và chọn lọc tự nhiên dựa trên sự biến đổi di truyền, và động lực tiến hóa của các giống và giống là biến đổi di truyền và chọn lọc nhân tạo.
Bằng di truyền, Darwin hiểu được khả năng của sinh vật trong việc bảo tồn loài, đặc điểm giống và cá thể ở con cái của chúng, và bằng tính biến đổi - khả năng của sinh vật có được những đặc điểm mới dưới tác động của điều kiện môi trường. Ông phân biệt giữa sự biến đổi xác định, không xác định và tương đối.
Sự biến đổi nhất định (hoặc nhóm) là biểu hiện của những đặc điểm tương tự ở tất cả các cá thể dưới ảnh hưởng của cùng điều kiện môi trường. Hiện nay người ta đã xác định rằng sự biến đổi này không ảnh hưởng đến kiểu gen của sinh vật và được gọi là biến đổi hoặc kiểu hình.
Tính biến thiên không xác định (hoặc cá thể) là sự xuất hiện của những khác biệt cá thể ở các cá thể cùng loài. Sự khác biệt cá nhân được kế thừa. Đây là biến thể di truyền hoặc di truyền.
Ngoài ra, Darwin còn xác định được sự biến đổi tương quan, khi một sự thay đổi ở một cơ quan hoặc đặc điểm kéo theo những thay đổi ở các cơ quan hoặc đặc điểm khác. Ví dụ, động vật chân dài có cổ dài. Ở các giống củ cải đường, màu sắc của rễ, cuống lá và gân lá thay đổi liên tục.
Cuộc đấu tranh sinh tồn theo Darwin thể hiện mối quan hệ phức tạp và đa dạng của các sinh vật giữa chúng với thiên nhiên vô tri. Có nhiều hình thức đấu tranh sinh tồn khác nhau: giữa các loài, giữa các loài và đấu tranh chống lại những điều kiện không thuận lợi.
Nội bộ (cạnh tranh). Kết quả là sự bảo tồn quần thể và loài do cái chết của kẻ yếu. Chiến thắng của một quần thể có khả năng tồn tại cao hơn trước một quần thể kém khả thi hơn trong cùng một khu vực sinh thái. Ví dụ: sự cạnh tranh giữa những kẻ săn mồi trong cùng một quần thể để giành con mồi; ăn thịt đồng loại - tiêu diệt động vật trẻ khi quy mô dân số quá lớn; đấu tranh giành quyền thống trị trong đàn; rừng thông đồng tuổi.
Vì vậy, tất cả các hình thức đấu tranh sinh tồn cuối cùng đều dẫn đến sự sống sót của những sinh vật thích nghi nhất với các điều kiện cụ thể, tức là với chọn lọc tự nhiên.
Chọn lọc tự nhiên là một quá trình diễn ra liên tục trong tự nhiên, trong đó những cá thể phù hợp nhất của mỗi loài sẽ sống sót và để lại con cái, những cá thể kém phù hợp hơn sẽ chết. Một điều kiện cần thiết của chọn lọc tự nhiên là tính biến đổi di truyền và kết quả trước mắt là sự hình thành những thích nghi của sinh vật với những điều kiện tồn tại cụ thể. Một ví dụ kinh điển về chọn lọc tự nhiên là sự thay đổi màu sắc của bướm bạch dương. Có sự chọn lọc tự nhiên thúc đẩy, ổn định và phá vỡ (xé bỏ).
Lựa chọn dẫn động hay chọn hướng là lựa chọn chỉ thiên về một hướng biến đổi (Hình 4). Được mô tả bởi Darwin. Ví dụ, sự xuất hiện hiện nay của các nhóm chuột và côn trùng có khả năng kháng thuốc trừ sâu; các chủng vi sinh vật kháng thuốc kháng sinh.
Ổn định chọn lọc là chọn lọc nhằm mục đích bảo tồn một đặc điểm trung bình đã được thiết lập trước đó trong quần thể và hành động chống lại các biểu hiện của sự biến đổi kiểu hình (Hình 5). Được I.I. Shmalhausen mô tả năm 1946. Ví dụ, kích thước và hình dạng hoa ở cây thụ phấn nhờ côn trùng ổn định hơn ở cây thụ phấn nhờ gió (cấu trúc hoa của cây thụ phấn nhờ côn trùng tương ứng với cấu trúc của hoa thụ phấn nhờ côn trùng). Chỉ những cây có cấu trúc hoa không thay đổi mới để lại cây con. Chim duy trì chiều dài cánh trung bình.

Cơm. 4. Hình thức thúc đẩy chọn lọc tự nhiên: A - D - những thay đổi liên tiếp về tốc độ phản ứng dưới áp lực của động lực chọn lọc tự nhiên
Chọn lọc gián đoạn hoặc gián đoạn là chọn lọc thiên về hai hoặc nhiều hướng biến đổi ở sinh vật. Ví dụ: côn trùng trên các đảo đại dương (Hình 6). Lựa chọn đột phá nhằm mục đích chống lại việc duy trì giá trị trung bình của một tính trạng. Hình thức chọn lọc này được mô tả bởi K. Mazer (1973).

Cơm. 5. Ổn định hình thức chọn lọc tự nhiên

Hình: 6. Hình thức đột phá của chọn lọc tự nhiên
Tất cả các hình thức chọn lọc tự nhiên tạo thành một cơ chế duy nhất duy trì sự cân bằng của quần thể với môi trường. Sự lựa chọn bắt đầu trong một quần thể.
Đơn vị tiến hóa cơ bản là quần thể, vì chỉ có nó mới đại diện cho sự thống nhất về sinh thái, hình thái sinh lý và di truyền. Tập hợp các gen trong quần thể được gọi là vốn gen. Trong các quần thể lớn, nơi không có đột biến, chọn lọc và trộn lẫn với các quần thể khác, người ta quan sát thấy sự không đổi của tần số alen, đồng hợp tử và dị hợp tử (định luật Hardy-Weinberg).
Tất cả các yếu tố gây ra sự sai lệch so với định luật Hardy-Weinberg đều là những yếu tố tiến hóa cơ bản. Chúng bao gồm: đột biến, chọn lọc tự nhiên, làn sóng dân số và sự cô lập. Đột biến liên tục xảy ra trong quần thể dưới tác động của các yếu tố môi trường gây đột biến và gây ra những thay đổi về vốn gen của quần thể. gen trong quần thể. Sự cách ly dẫn đến sự khác biệt về các đặc điểm trong cùng một loài và ngăn cản các cá thể thuộc các quần thể, loài khác nhau giao phối với nhau. Sự cách ly về địa lý, sinh thái và sinh học được phân biệt.
Thuyết tiến hóa tổng hợp
Người đầu tiên kết hợp dữ liệu di truyền học và học thuyết Darwin là nhà động vật học và nhà giải phẫu học so sánh N.K. Koltsov (1872-1940). Học trò và đồng nghiệp của ông, S.S. Chettorikov (1880-1959) là người đầu tiên cung cấp cơ sở di truyền cho những lời dạy về tiến hóa của Darwin. Trong tác phẩm nổi tiếng của S. S. Chettorikov “Về một số khía cạnh của quá trình tiến hóa theo quan điểm của di truyền học hiện đại” (1926), người ta đã chỉ ra rằng trong các điều kiện tự nhiên trong tự nhiên ở mỗi loài, có một số lượng lớn các thay đổi di truyền không xảy ra. biểu hiện ra kiểu hình do tính trạng lặn. Loài này chứa đầy các đột biến, tạo thành vật liệu vô tận cho quá trình tiến hóa.
Bắt đầu vào những năm 20. thế kỷ trước, sự thống nhất giữa học thuyết Darwin và di truyền học đã góp phần mở rộng và đào sâu sự tổng hợp của học thuyết Darwin với các ngành khoa học khác; 30-40 tuổi Đây được coi là thời kỳ hình thành thuyết tiến hóa tổng hợp (STE).
Một vai trò quan trọng trong việc hình thành STE thuộc về công trình “Di truyền học và nguồn gốc các loài” của F. G. Dobzhansky (1937), trong đó tóm tắt sự tổng hợp di truyền học với học thuyết Darwin. Nhà khoa học Liên Xô I. I. Shmalgauzen (1887-1963) đã đóng góp đáng kể vào việc tạo ra STE. Ông đã nghiên cứu mối quan hệ giữa sự phát sinh bản thể và phát sinh loài, nghiên cứu các hướng chính của quá trình tiến hóa và xác định hai hình thức chọn lọc tự nhiên. Các tác phẩm của ông là “Con đường và mô hình của quá trình tiến hóa” (1939), “Các yếu tố tiến hóa” (1946).
(1940), với việc xuất bản các cuốn sách của E. Mayr “Hệ thống và nguồn gốc các loài” (1944) và J. Huxley “Tiến hóa: Tổng hợp hiện đại” (1942). Thuật ngữ “thuyết tiến hóa tổng hợp” được đặt tên theo tựa đề cuốn sách “Tiến hóa: Sự tổng hợp hiện đại” của J. Huxley.
Các tiên đề cơ bản của STE
1. Nguyên liệu của sự tiến hóa, theo quy luật, là những thay đổi rất nhỏ nhưng rời rạc trong di truyền - đột biến.
2. Quá trình đột biến, các làn sóng số - nguồn cung cấp nguyên liệu cho quá trình chọn lọc - mang tính chất ngẫu nhiên và không định hướng.
3. Yếu tố định hướng duy nhất của quá trình tiến hóa là chọn lọc tự nhiên, dựa trên sự bảo tồn và tích lũy các đột biến nhỏ và ngẫu nhiên.
4. Đơn vị tiến hóa nhỏ nhất là một quần thể chứ không phải một cá thể, như đã được giả định dựa trên ý tưởng về khả năng “thừa kế các đặc điểm có được”. Do đó, đặc biệt chú ý đến việc nghiên cứu quần thể như là cấu trúc cơ bản của một đơn vị loài.
5. Sự tiến hóa có tính chất khác nhau, nghĩa là một taxon có thể trở thành tổ tiên của nhiều taxon con, nhưng mỗi loài chỉ có một loài tổ tiên duy nhất, một quần thể tổ tiên duy nhất.
6. Sự tiến hóa là dần dần và lâu dài. Sự hình thành loài là một giai đoạn của quá trình tiến hóa là sự thay thế tuần tự một quần thể tạm thời bằng một loạt quần thể tạm thời tiếp theo.
 Bộ sưu tập những kẻ lập dị và người sửa đổi cơ thể người Nga
Bộ sưu tập những kẻ lập dị và người sửa đổi cơ thể người Nga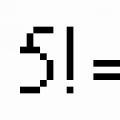 Lý thuyết xác suất và thống kê toán học
Lý thuyết xác suất và thống kê toán học Các khái niệm cơ bản của lý thuyết xác suất và thống kê toán học
Các khái niệm cơ bản của lý thuyết xác suất và thống kê toán học