Poznavanje zakona homoloških nizova nasljedne varijabilnosti omogućuje. Zakon N.I. Vavilova (zakon homolognog niza nasljedne varijabilnosti)
Dana 4. lipnja održao je prezentaciju na temu “Zakon homolognih nizova u nasljednoj varijaciji”. Ovo je jedan od onih radova koji se smatraju temeljnim i čine teorijsku osnovu bioloških istraživanja. Bit zakona svodi se na to da vrste i rodove koji su genetski bliski (međusobno povezani jedinstvom podrijetla) karakteriziraju slični nizovi u nasljednoj varijabilnosti. Studentska strast za proučavanjem žitarica, a potom i križarica, mahunarki i bundeva omogućila je Vavilovu i njegovim studentima da pronađu slične mutacije u srodnim vrstama, a potom i rodovima. U tablici razvijenoj kao rezultat pokusa, Vavilov je znakom "+" označio mutacije čija je manifestacija pronađena u tim vrstama, a prazna mjesta pokazuju da bi takve mutacije trebale postojati, ali još nisu otkrivene. Tablica s praznim poljima, koja će se popunjavati daljnjim razvojem znanosti. Jesmo li se već susreli s nečim ovakvim?! Naravno, u kemiji, poznati periodni sustav! Obrazac dvaju zakona potvrdila je znanost. “Prazne” ćelije se popunjavaju i to je osnova za praktičan odabir. Tvrda pšenica poznata je samo u jarom obliku, no po zakonu bi u prirodi trebala postojati i durum pšenica u ozimom obliku. Doista je ubrzo otkriven na granici Irana i Turske. Bundeve i dinje karakteriziraju jednostavni i segmentirani plodovi, ali lubenica ovog oblika nije opisana u vrijeme Vavilova. Ali segmentirane lubenice otkrivene su na jugoistoku europske Rusije. U kulturi prevladava uzgoj repe s tri klice, čiji usjevi zahtijevaju plijevljenje i uklanjanje dva suvišna izdanka. Ali među rođacima cikle u prirodi bilo je i oblika s jednom klicom, pa su znanstvenici uspjeli stvoriti novu sortu repe s jednom klicom. Priroda usjeva žitarica bez tilja je mutacija koja se pokazala korisnom uvođenjem strojne žetve, kada se mehanizmi manje začepljuju. Uzgajivači su, koristeći Vavilovljev zakon, pronašli oblike bez oštrice i stvorili nove sorte žitarica bez oštrice. Činjenice paralelne varijabilnosti u bliskim i udaljenim vrstama bile su poznate čak i Charlesu Darwinu. Na primjer, ista boja krzna glodavaca, albinizam kod predstavnika različitih skupina životinjskog svijeta i ljudi (opisan je slučaj albinizma kod crnaca), nedostatak perja kod ptica, nedostatak ljuski kod riba, slična boja plodova voćnih i bobičastih kultura, varijabilnost korijenskih usjeva, itd. Razlog paralelizma u varijabilnosti leži u činjenici da se homologni karakteri temelje na prisutnosti sličnih gena: što su vrste i rodovi genetski bliži, to je potpunija sličnost u serija varijabilnosti. Odatle i uzrok homolognih mutacija – zajedničko podrijetlo genotipova. U procesu evolucije živa priroda programirana je kao prema jednoj formuli, bez obzira na vrijeme nastanka vrste. N. I. Vavilov zakon homoloških serija u nasljednoj varijabilnosti nije bio samo potvrda Darwinove doktrine o podrijetlu vrsta, već je također proširio ideju nasljedne varijabilnosti. Nikolaj Ivanovič opet može izjaviti: “Hvala Darwinu!”, ali i “Nastavljamo Darwina!” Vratimo se u 1920. Zanimljiva su sjećanja očevidaca. Aleksandra Ivanovna Mordvinkina, koja je bila prisutna na kongresu Saratovskog poljoprivrednog instituta (kasnije kandidat bioloških znanosti), prisjetila se: „Kongres je otvoren u najvećoj dvorani sveučilišta. Nijedan izvještaj kasnije nije na mene ostavio tako snažan dojam kao govor Nikolaja Ivanoviča. Govorio je nadahnuto, svi su slušali bez daha, osjećalo se da se pred nama otvara nešto jako veliko, novo u znanosti. Nakon burnog, dugotrajnog pljeska, profesor Vjačeslav Rafailovič Zelenski je rekao: “Ovo su biolozi koji pozdravljaju svog Mendeljejeva.” Posebno su mi se urezale u sjećanje riječi Nikolaja Maksimoviča Tulajkova: “Što se može dodati ovom izvještaju? Mogu reći jedno: Rusija neće propasti ako ima sinove kao što je Nikolaj Ivanovič. Nikolaj Vladimirovič Timofejev-Resovski, vrsni genetičar koji je Vavilova poznavao ne samo iz njegova rada, nego i osobno, povjerljivo je razgovarao s bliskim poznanicima: “Nikolaj Ivanovič bio je divan čovjek i veliki mučenik, izvrstan oplemenjivač i sakupljač biljaka, putnik, hrabar i sveopći miljenik, ali njegov zakon homologijskog niza - zakon uopće nije homološki, nego analogni niz, da, gospodine! Što je homologija? Ovo je sličnost temeljena na zajedničkom podrijetlu. Što je analogija? Sličnost vanjskih karakteristika, koja je određena sličnim staništem, ali ne i srodstvom. Pa tko je u pravu? Vavilov! Može se samo diviti dubini njegovog biološkog uma! Promjenom samo jednog pojma u nazivu mijenja se bit zakona. Prema zakonu homoloških nizova svi su ljudi jednaki, jer su istog biološkog podrijetla i pripadaju vrsti homo sapiens, tj. svi su jednako pametni, sposobni i talentirani itd., ali imaju vanjske razlike: po visini, omjerima između dijelova tijela itd. Prema zakonu analognog niza, ljudi su slični izgledom, jer imaju slično stanište, ali različito podrijetlo. A to je već prostor za šovinizam, rasizam, nacionalizam, čak i genocid. A Vavillov zakon kaže da su afrički pigmej i američki košarkaš istog genetskog podrijetla i da se jedan ne može stavljati iznad drugoga - to je antiznanstveno! Valjanost univerzalnog biološkog obrasca koji je otkrio Vavilov potvrđena je suvremenim istraživanjima ne samo na biljkama, već i na životinjama. Moderni genetičari vjeruju da zakon otkriva bezgranične izglede za znanstveno znanje, generalizaciju i predviđanje” (profesor M. E. Lobanov). Drugi temeljni rad N. I. Vavilova, "Imunitet biljaka na zarazne bolesti" (1919.), datira iz Saratovskog razdoblja. Na naslovnoj stranici knjige Nikolaj Ivanovič je napisao: “Posvećeno sjećanju na velikog istraživača imuniteta Ilju Iljiča Mečnikova.” Niti jedan veliki znanstvenik sebe ne vidi izdvojenim u znanosti. Tako se Vavilov, zahvaljujući Mečnikovu, zapitao mogu li biljke imati zaštitnu moć ako je imaju životinje? U potrazi za odgovorom na pitanje proveo je istraživanje žitarica originalnom metodom i, sažimajući praksu i teoriju, postavio temelje nove znanosti - fitoimunologije. Rad je imao isključivo praktični značaj - iskoristiti prirodnu imunost biljaka kao najracionalniji i najisplativiji način suzbijanja štetnika. Mladi znanstvenik stvorio je originalnu teoriju o fiziološkoj imunosti biljaka na zarazne bolesti, a temelj njegovog učenja bilo je proučavanje genotipske imunosti. N. I. Vavilov proučavao je reakciju "domaćina" na uvođenje parazita, specifičnost te reakcije i otkrio je li cijela serija imuna ili samo određene vrste ove serije. Nikolaj Ivanovič pridavao je posebnu važnost grupnom imunitetu, vjerujući da je u uzgoju važno razviti sorte koje su otporne ne na jednu rasu, već na cijelu populaciju fizioloških rasa, a takve otporne vrste treba tražiti u domovini biljke. Znanost je kasnije potvrdila da divlje vrste - srodnici kulturnih biljaka - imaju prirodni imunitet i manje su osjetljive na zarazne bolesti. To je uvođenje gena otpornosti u biljke kojima se bave moderni uzgajivači, koristeći teoriju N. I. Vavilova i metode genetskog inženjeringa. Znanstvenik je bio zainteresiran za razvoj pitanja imuniteta tijekom cijele svoje znanstvene karijere: "Doktrina imuniteta biljaka na zarazne bolesti" (1935.), "Zakoni prirodnog imuniteta biljaka na zarazne bolesti (ključevi za pronalaženje imunoloških oblika)" ( objavljen tek 1961.). Akademik Pjotr Mihajlovič Žukovski s pravom je primijetio: „Tijekom Saratovskog razdoblja, iako je bilo kratko (1917.-1921.), izašla je zvijezda N. I. Vavilov - znanstvenik." Vavilov će kasnije napisati: "Emigrirao sam iz Saratova u ožujku 1921. s cijelim laboratorijem od 27 ljudi." Izabran je za šefa Zavoda za primijenjenu botaniku Poljoprivrednog znanstvenog odbora u Petrogradu. Od 1921. do 1929. god - Profesor Odsjeka za genetiku i uzgoj Lenjingradskog poljoprivrednog instituta. Godine 1921. V. I. Lenjin je poslao dva znanstvenika na konferenciju u Ameriku, jedan od njih bio je N. I. Vavilov. Izvješće o genetskom istraživanju učinilo ga je popularnim među znanstvenicima konferencije. U Americi su njegovi nastupi popraćeni ovacijama, sličnim onima koje su uslijedile za Chkalova. "Ako su svi Rusi takvi, onda moramo biti prijatelji s njima", vikali su američki listovi. Za 20-30 god. N. I. Vavilov također se očituje kao glavni organizator znanosti. On je zapravo bio tvorac i stalni ravnatelj Svesaveznog instituta za uzgoj biljaka (VIR). Godine 1929. Svesavezna akademija poljoprivrednih znanosti (VASKhNIL) stvorena je na temelju Svesaveznog instituta za eksperimentalnu agronomiju, koji je prethodno organizirao Vavilov. Izabran je za prvog predsjednika (od 1929. do 1935.). Uz izravno sudjelovanje znanstvenika, organiziran je Institut za genetiku Akademije znanosti SSSR-a. U kratkom vremenu Vavilov talent stvorio je znanstvenu školu genetičara, koja je postala vodeća u svijetu. Sve početne radove u našoj zemlji na području genetike izvodio je on ili pod njegovim vodstvom. U VIR-u je prvi put korištena metoda eksperimentalne poliploidije, a G. D. Karpechenko je započeo rad na njezinoj primjeni u daljinskoj hibridizaciji. Vavilov je inzistirao na početku rada na korištenju fenomena heterozisa i međulinijske hibridizacije. Danas je ovo abeceda selekcije, ali tada je to bio početak. Tijekom 30 godina znanstvenog djelovanja objavljeno je oko 400 radova i članaka! Fenomenalno pamćenje, enciklopedijsko znanje, vlada gotovo dvadeset jezika, svjestan svih novotarija u znanosti. Radilo se 18-20 sati dnevno. Majka ga je prekorila: “Nemaš vremena ni za spavanje...”, prisjeća se Vavilovljev sin.
MUTACIONA VARIJABILNOST
Plan
Razlika između mutacija i modifikacija.
Klasifikacija mutacija.
Zakon N.I.Vavilova
Mutacije. Pojam mutacije. Mutageni čimbenici.
Mutacije – su iznenadne, trajne, prirodne ili umjetne promjene genetskog materijala koje nastaju pod utjecajem mutageni faktori .
Vrste mutagenih faktora:
A) fizički– zračenje, temperatura, elektromagnetsko zračenje.
B) kemijski faktori – tvari koje uzrokuju trovanje tijela: alkohol, nikotin, formaldehid.
U) biološki- virusi, bakterije.
Razlika između mutacija i modifikacija
Klasifikacija mutacija
Postoji nekoliko klasifikacija mutacija.
I Klasifikacija mutacija prema značenju: korisne, štetne, neutralne.
Koristan mutacije dovode do povećane otpornosti organizma i materijal su za prirodnu i umjetnu selekciju.
Štetne mutacije smanjuju vitalnost i dovode do razvoja nasljednih bolesti: hemofilije, anemije srpastih stanica.
II Klasifikacija mutacija prema lokalizaciji ili mjestu nastanka: somatske i generativne.
Somatski nastaju u stanicama tijela i zahvaćaju samo dio tijela, dok se razvijaju mozaične jedinke: različite oči, boja kose. Ove se mutacije nasljeđuju samo tijekom vegetativnog razmnožavanja (kod ribiza).
Generativno – javljaju se u spolnim stanicama ili u stanicama iz kojih nastaju gamete. Dijele se na nuklearne i ekstranuklearne (mitohondrijske, plastidne).
III Mutacije po prirodi promjene genotipa: kromosomske, genomske, genske.
Genetski (ili točka) nisu vidljivi pod mikroskopom, povezani su s promjenama u strukturi gena. Ove mutacije nastaju kao rezultat gubitka nukleotida, umetanja ili zamjene jednog nukleotida drugim. Ove mutacije dovode do bolesti gena: daltonizma, fenilketonurije.
Kromosomski (perestrojka) povezana s promjenama u strukturi kromosoma. Što se može dogoditi:
Brisanje: - gubitak dijela kromosoma;
Dupliciranje – udvostručenje dijela kromosoma;
Inverzija – rotacija dijela kromosoma za 180 0;
Translokacija – izmjena dijelova nehomolognih kromosoma i spajanje dva nehomologna kromosoma u jedan.
Uzroci kromosomskih mutacija: pojava dva ili više loma kromosoma nakon čega slijedi njihovo spajanje, ali pogrešnim redoslijedom.
Genomski mutacije dovesti do promjene broja kromosoma. razlikovati heteroploidija I poliploidija.
Heteroploidija povezana s promjenom broja kromosoma, na više kromosoma – 1.2.3. Uzroci: nerazdvajanje kromosoma u mejozi:
- Monosomija – smanjenje broja kromosoma za 1 kromosom. Opća formula kromosomskog seta je 2n-1.
- trisonomija – povećanje broja kromosoma za 1. Opća formula 2n+1 (47 kromosoma Clanfeatherov sindrom; trisonomija 21 para kromosoma - Downov sindrom (znakovi višestrukih urođenih mana koje smanjuju vitalnost organizma i poremećen psihički razvoj).
poliploidija – višestruka promjena broja kromosoma. U poliploidnim organizmima haploidni (n) skup kromosoma u stanicama ponavlja se ne 2 puta, kao u diploidnim, već 4-6 puta, ponekad i mnogo više - do 10-12 puta.
Pojava poliploida povezana je s kršenjem mitoze ili mejoze. Konkretno, nedivergencija homolognih kromosoma u mejozi dovodi do stvaranja gameta s povećanim brojem kromosoma. U diploidnim organizmima ovaj proces može proizvesti diploidne (2n) gamete.
Rasprostranjen u kultiviranim biljkama: heljdi, suncokretu itd., kao iu samoniklim biljkama.
Zakon N.I. Vavilova (zakon homolognog niza nasljedne varijabilnosti).
/ Od davnina su istraživači zapažali postojanje sličnih karakteristika kod različitih vrsta i rodova iste obitelji, na primjer, dinje, slične krastavcima, ili lubenice, slične dinjama. Te su činjenice činile osnovu zakona homoloških nizova u nasljednoj varijabilnosti./
Višestruki alelizam. Paralelna varijabilnost. Gen može postojati u više od dva stanja. Raznolikost alela jednog gena naziva se višestruki alelizam. Različiti aleli određuju različite stupnjeve iste osobine. Što više alela nosi jedinka u populaciji, to je vrsta plastičnija i bolje prilagođena promjenjivim uvjetima okoliša.
U osnovi je višestruki alelizam paralelna varijabilnost - pojava u kojoj se slični znakovi pojavljuju u različitim vrstama i rodovima iste obitelji. Sistematizirao je činjenice paralelne varijabilnosti N.I. Vavilov./
N.I. Vavilov uspoređivao je vrste iz obitelji žitarica. Utvrdio je da ako meka pšenica ima ozime i jare oblike, osnate i bezosne, onda se isti oblici nužno nalaze i u durum pšenici. Štoviše, sastav značajki. Po čemu se oblici razlikuju unutar vrste i roda, često se pokaže da je isto i kod drugih rodova. Na primjer, oblici raži i ječma ponavljaju oblike različitih vrsta pšenice, tvoreći isti paralelni ili homologni niz nasljedne varijabilnosti.
Sistematizacija činjenica omogućila je N.I. Vavilovu da formulira zakon homolognog niza u nasljednoj varijabilnosti (1920): vrste i rodovi koji su genetski bliski karakterizirani su sličnim nizovima nasljedne varijabilnosti s takvom pravilnošću. Da se, poznavajući niz oblika unutar jedne vrste, može predvidjeti prisutnost paralelnih oblika u drugim vrstama i rodovima.
Homologija nasljednih svojstava blisko srodnih vrsta i rodova objašnjava se homologijom njihovih gena, budući da potječu od iste vrste pretka. Osim toga, proces mutacije u genetski bliskim vrstama odvija se na sličan način. Stoga razvijaju slične nizove recesivnih alela i, kao rezultat toga, paralelna svojstva.
Zaključak iz Vavilova zakona: svaka vrsta ima određene granice mutacijske varijabilnosti. Nijedan proces mutacije ne može dovesti do promjena koje nadilaze spektar nasljedne varijabilnosti vrste. Tako kod sisavaca mutacije mogu promijeniti boju dlake iz crne u smeđu, crvenu, bijelu, mogu se pojaviti pruge i pjegavosti, ali je isključena pojava zelene boje.
©2015-2019 stranica
Sva prava pripadaju njihovim autorima. Ova stranica ne polaže pravo na autorstvo, ali omogućuje besplatnu upotrebu.
Datum izrade stranice: 2016-04-12
utvrđuje paralelizam u nasljeđivanju i varijabilnosti organizama. Formulirao N. I. Vavilov 1920. Proučavajući varijabilnost svojstava u vrstama i rodovima žitarica i drugih porodica, N. I. Vavilov je otkrio da: 1. Vrste i rodovi koji su genetski bliski jedni drugima karakteriziraju identični nizovi nasljedne varijabilnosti s takvim pravilnost da se znajući niz oblika za jednu vrstu, može predvidjeti prisutnost identičnih oblika u drugim vrstama i rodovima. Što su rodovi i Linneons bliže genetski smješteni u općem sustavu, to je potpuniji identitet u redovima njihove varijabilnosti. 2. Cijele obitelji biljaka općenito karakterizira određeni ciklus varijabilnosti koji prolazi kroz sve rodove koji čine obitelj.” Iako se zakon u početku odnosio na varijabilnost biljaka, N. I. Vavilov je istaknuo njegovu primjenjivost na životinje. Teorijski osnova homologije fenotipskih nizova. varijabilnost u srodnoj taksonomiji. skupina je ideja o jedinstvu njihova nastanka kroz diverzifikaciju pod utjecajem prirode. izbor. Budući da su zajednički preci trenutno postojećih vrsta imali specifičan, specifičan skup gena, onda njihovi potomci moraju imati, uz manje iznimke, isti skup gena. S obzirom da svaki gen može mutirati u različitim smjerovima (multiple, alelizam) i da je proces mutacije neusmjeren, prirodno je pretpostaviti da će spektar promjena identičnih gena kod jedinki bliskih vrsta biti sličan. Dakle, zakon se temelji na homologiji. redova (3. r.) leži paralelizam genotipskih. varijabilnost kod pojedinaca sa sličnim skupom gena. Biti teoretski. Na temelju komparativne genetike, zakon objašnjava polimorfizam vrsta i time opravdava cjelovitost vrste, unatoč postojanju unutar njezinih granica morfološki jasno različitih oblika. S druge strane, zakon unosi jasnoću u fenotipski fenomen. "homogenost" množina vrste, rub se može povezati s njihovom heterozigotnošću i fenomenom dominacije, koji se otkriva tijekom inbridinga. 3. r., odražavajući opći obrazac procesa mutacije i formiranja organizama, je biol. temelj metoda za namjensko dobivanje potrebnih nasljeđa i promjena. Uzgajivačima ukazuje na smjerove umjetnosti, selekcije ili, kako je napisao N. I. Vavilov, "što tražiti", a metode traženja mogu biti različite: od pronalaženja željenih oblika u prirodi ili njihovog identificiranja kroz inbreeding do dobivanja tih oblika pomoću mutageni. Biokemijski mehanizmi 3. g.r. naširoko se proučavaju na različitim objektima - od promjena u bakterijskom metabolizmu do mikrobioloških procesa. sinteza nasljedstva, ljudske bolesti.
Zakon homologijskog niza
Obrada opsežnog promatračkog i eksperimentalnog materijala, detaljna studija varijabilnosti brojnih Linneovih vrsta (Linneons), ogroman broj novih činjenica dobivenih uglavnom proučavanjem kultiviranih biljaka i njihovih divljih srodnika, omogućili su N.I. Vavilov je okupio sve poznate primjere paralelne varijabilnosti i formulirao opći zakon, koji je nazvao "Zakon homolognih serija u nasljednoj varijabilnosti" (1920.), o čemu je izvijestio na Trećem sveruskom kongresu uzgajivača, održanom u Saratovu. Godine 1921. N.I. Vavilov je poslan u Ameriku na Međunarodni kongres poljoprivrede, gdje je održao prezentaciju o zakonu homoloških serija. Zakon paralelne varijabilnosti bliskih rodova i vrsta, koji je uspostavio N.I. Vavilov i povezan sa zajedničkim podrijetlom, razvijajući evolucijska učenja Charlesa Darwina, cijenjena je od svjetske znanosti. Publika ga je doživjela kao najveći događaj u svjetskoj biološkoj znanosti koji otvara najšire horizonte praksi.
Zakon homolognog niza, prije svega, uspostavlja osnovu za sistematiku ogromne raznolikosti biljnih oblika u kojima je organski svijet tako bogat, omogućava oplemenjivaču da dobije jasnu predodžbu o mjestu svakog, pa čak i najmanju, sustavnu jedinicu u biljnom svijetu i prosuditi moguću raznolikost izvornog materijala za selekciju.
Glavne odredbe zakona homoloških serija su sljedeće.
"1. Genetski bliske vrste i rodovi karakteriziraju slični nizovi nasljedne varijabilnosti s takvom pravilnošću da se, poznavajući nizove oblika unutar jedne vrste, može predvidjeti prisutnost paralelnih oblika u drugim vrstama i rodovima. Što su rodovi i Linneoni genetski locirani u općem sustavu, to je potpunija sličnost u nizu njihove varijabilnosti.
2. Cijele obitelji biljaka općenito karakterizira određeni ciklus varijabilnosti koji prolazi kroz sve rodove i vrste koje čine obitelji.”
Čak i na III Sveruskom izbornom kongresu (Saratov, lipanj 1920.), gdje je N.I. Vavilov prvi izvijestio o svom otkriću, svi su sudionici kongresa priznali da će "poput periodnog sustava (periodnog sustava)" zakon homoloških nizova omogućiti predviđanje postojanja, svojstava i strukture još nepoznatih oblika i vrsta biljaka i životinja, te je visoko cijenio znanstveni i praktični značaj ovog zakona. Suvremeni napredak molekularne stanične biologije omogućuje razumijevanje mehanizma postojanja homološke varijabilnosti u bliskim organizmima – na čemu se točno temelji sličnost budućih oblika i vrsta s već postojećima – te smislenu sintezu novih oblika biljaka koji ne postoje u prirodi. Sada se Vavillovljevom zakonu dodaje novi sadržaj, baš kao što je pojava kvantne teorije dala novi, dublji sadržaj Mendeljejevljevom periodnom sustavu.
Nauk o središtima nastanka kulturnih biljaka
Već sredinom 20-ih godina prošlog stoljeća počelo je istraživanje geografske distribucije i intraspecifične raznolikosti raznih poljoprivrednih kultura, koje je proveo N.I. Vavilov i pod njegovim vodstvom omogućili su Nikolaju Ivanoviču da formulira ideje o geografskim središtima podrijetla kultiviranih biljaka. Knjiga “Centri podrijetla kultiviranih biljaka” objavljena je 1926. Duboko teoretski utemeljena ideja o središtima podrijetla dala je znanstvenu osnovu za ciljanu potragu za biljkama korisnim za ljude, te je široko korištena u praktične svrhe.
Ne manje važno za svjetsku znanost je učenje N. I. Vavilova o središtima podrijetla kultiviranih biljaka io geografskim obrascima u distribuciji njihovih nasljednih svojstava (prvi put objavljeno 1926. i 1927.). U ovim klasičnim djelima N.I. Vavilov je prvi dao cjelovitu sliku koncentracije golemog bogatstva oblika kultiviranih biljaka u nekoliko primarnih središta njihova nastanka i pristupio pitanju podrijetla kultiviranih biljaka na sasvim nov način. Ako su prije njega botaničari-geografi (Alphonse De-Candolle i drugi) tražili "opću" domovinu pšenice, onda je Vavilov tražio središta podrijetla pojedinih vrsta, skupina vrsta pšenice u raznim regijama zemaljske kugle. Pri tome je posebno važno bilo identificirati područja prirodne rasprostranjenosti (areala) varijeteta određene vrste i odrediti središte najveće raznolikosti njezinih oblika (botaničko-geografska metoda).
Kako bi utvrdio geografsku distribuciju sorti i rasa kultiviranih biljaka i njihovih divljih srodnika, N.I. Vavilov je proučavao središta drevne poljoprivredne kulture, čiji je početak vidio u planinskim predjelima Etiopije, zapadne i središnje Azije, Kine, Indije, u Andama Južne Amerike, a ne u širokim dolinama velikih rijeka - Nila. , Ganges, Tigris i Eufrat, kako su znanstvenici ranije tvrdili. Rezultati naknadnih arheoloških istraživanja potvrđuju ovu hipotezu.
Kako bi pronašao centre raznolikosti i bogatstva biljnih oblika, N.I. Vavilov je organizirao brojne ekspedicije prema određenom planu koji je odgovarao njegovim teorijskim otkrićima (homologne serije i središta podrijetla kultiviranih biljaka), koje su 1922.–1933. posjetio 60 zemalja svijeta, kao i 140 regija naše zemlje. Time je prikupljen vrijedan fond svjetskih biljnih resursa koji broji preko 250.000 primjeraka. Najbogatija prikupljena zbirka pomno je proučavana metodama selekcije, genetike, kemije, morfologije, taksonomije i zemljopisnih kultura. I dalje se čuva u VIR-u i koriste ga naši i strani uzgajivači.
Stvaranje N.I. Vavilov moderna doktrina selekcije
Sustavno proučavanje svjetskih biljnih resursa najvažnijih kulturnih biljaka radikalno je promijenilo shvaćanje sortnog i specijskog sastava čak i tako dobro proučenih usjeva kao što su pšenica, raž, kukuruz, pamuk, grašak, lan i krumpir. Među vrstama i mnogim sortama ovih kultiviranih biljaka donesenih s ekspedicija, pokazalo se da je gotovo polovica novih, još nepoznatih znanosti. Otkriće novih vrsta i sorti krumpira potpuno je promijenilo dotadašnje shvaćanje izvorne sirovine za njegovu selekciju. Na temelju materijala prikupljenog od strane ekspedicija N.I. Vavilov i njegovi suradnici, osnovana je cjelokupna selekcija pamuka i izgrađen je razvoj vlažnih suptropika u SSSR-u.
Na temelju rezultata detaljnog i dugotrajnog proučavanja sortnog bogatstva koje su prikupile ekspedicije izrađene su diferencijalne karte geografske lokalizacije sorti pšenice, zobi, ječma, raži, kukuruza, prosa, lana, graška, leće, graha, sastavljene su grah, slanutak, slanutak, krumpir i druge biljke . Na tim se kartama moglo vidjeti gdje je koncentrirana glavna sortna raznolikost navedenih biljaka, odnosno gdje treba nabaviti izvorni materijal za selekciju pojedine kulture. Čak i za tako drevne biljke kao što su pšenica, ječam, kukuruz i pamuk, koje su se dugo raširile diljem svijeta, bilo je moguće s velikom točnošću utvrditi glavna područja potencijala primarnih vrsta. Osim toga, utvrđeno je da se područja primarne formacije podudaraju za mnoge vrste, pa čak i rodove. Geografska istraživanja dovela su do uspostavljanja čitavih kulturno neovisnih flora specifičnih za pojedine regije.
Proučavanje svjetskih biljnih resursa omogućilo je N.I. Vavilov je u potpunosti ovladao izvornim materijalom za uzgojni rad u našoj zemlji, te je ponovno postavio i riješio problem izvornog materijala za genetička i selekcijska istraživanja. Razvio je znanstvene osnove selekcije: učenje o izvornom materijalu, botaničke i geografske osnove poznavanja biljaka, metode selekcije na gospodarska svojstva hibridizacije, inkubacije i dr., značaj udaljene međuvrsne i međugeneričke hibridizacije. Svi ovi radovi nisu izgubili svoje znanstveno i praktično značenje u današnje vrijeme.
Botaničko i geografsko proučavanje velikog broja kultiviranih biljaka dovelo je do intraspecifične taksonomije kultiviranih biljaka, što je rezultiralo radovima N.I. Vavilov “Linnaeove vrste kao sustav” i “Doktrina podrijetla kultiviranih biljaka nakon Darwina”.
Genotipska varijabilnost. Klasifikacija mutacija.
Modifikacijska varijabilnost, norma reakcije, statistički uzorci.
Uloga genotipa i okolišnih uvjeta u formiranju fenotipa.
Varijabilnost kao svojstvo živih bića. Vrste varijabilnosti.
Citoplazmatsko nasljeđe.
Varijabilnost je svojstvo živih bića, izraženo u sposobnosti stjecanja novih karakteristika ili gubljenja starih. Postoje dvije vrste varijabilnosti: fenotipska (nenasljedna) i genotipska (nasljedna).
Tipovi fenotipske varijabilnosti:
Ontogenetski (dob);
Izmjena.
Tipovi genotipske varijabilnosti:
Kombinativan;
Mutacijski.
Razvoj fenotipa organizma određen je međudjelovanjem njegove nasljedne osnove – genotipa – s uvjetima okoliša (vidi dijagram).
Na ekspresiju gena značajno utječu drugi geni. Mogućnost razvoja neke osobine također ovisi o utjecaju regulacijskih sustava tijela, prije svega; endokrini. Takvi znakovi kod pijetlova kao što su svijetlo perje, veliki češalj, priroda pjevanja i boja glasa posljedica su djelovanja muškog spolnog hormona. Ženski spolni hormoni koji se daju pijetlovima uzrokuju funkcioniranje gena koji određuju sintezu u jetri proteina koji čine žumanjak jajeta. Obično ti geni "rade" samo kod kokoši. Posljedično, unutarnje okruženje organizma također uvelike utječe na ekspresiju gena u obliku svojstva.
Svaki organizam se razvija i živi u određenim vanjskim uvjetima, doživljavajući utjecaj čimbenika okoliša - kolebanja temperature, svjetlosti, vlažnosti, količine i kvalitete hrane, te stupajući u odnose s drugim organizmima. Svi ti čimbenici mogu promijeniti morfološka i fiziološka svojstva organizama, odnosno njihov fenotip. Razni znakovi organizma mijenjaju se u različitim stupnjevima pod utjecajem vanjskog okruženja.
Svi znakovi organizma mogu se podijeliti u dvije skupine - kvalitativne i kvantitativne. Kvalitativne su karakteristike koje se utvrđuju deskriptivno (tipološki). To su boja cvijeća, oblik plodova, boja životinja, boja očiju i spolne razlike. Kvantitativna su svojstva koja se utvrđuju mjerenjem (nejesnost kokoši, proizvodnja mlijeka krava, masa sjemena pšenice). Na mnoge kvalitativne karakteristike okolišni uvjeti utječu manje nego na kvantitativne. Ali kvalitativne karakteristike također se mogu promijeniti. Na primjer, ako se krzno hermelinskog kunića (bijeli kunići s crnim šapama, repom i njuškom) obrije na bilo kojem dijelu tijela, boja novoizraslog krzna ovisit će o temperaturi okoline. Dakle, ako obrijete bijelu dlaku na leđima i držite životinju na temperaturi iznad +2 ° C, tada će na ovom mjestu rasti bijela dlaka. Pri temperaturama zraka nižim od +2 °C umjesto bijele će rasti crna vuna. Dakle, osobine se razvijaju kao rezultat interakcije genotipa i okoline. Isti genotip može dati različite vrijednosti svojstva u različitim uvjetima okoliša.
Granice unutar kojih su moguće promjene svojstava u određenom genotipu nazivaju se reakcijska norma. Neke osobine (na primjer, mliječnost) imaju vrlo široku reakcijsku normu, druge (boja dlake) imaju mnogo užu.
Ne nasljeđuje se osobina kao takva, već sposobnost organizma (njegov genotip), kao rezultat interakcije s razvojnim uvjetima, da proizvede određeni fenotip, drugim riječima, normu reakcije organizma na vanjske uvjete. je naslijeđeno.
Raznolikost fenotipova koja nastaje u organizmima pod utjecajem okolišnih uvjeta naziva se modifikacijska varijabilnost. Klasičan primjer modifikacijske varijabilnosti je promjena oblika maslačka izraslog u ravnici i u planinama iz istog rizoma (Bonnierov pokus, 1895.). Primjer je vrh strijele vodene biljke, koji ima tri oblika lista ovisno o vanjskom okruženju: vrpčasti (potopljeni), bubrežasti (plutajući) i streličasti (izlazeći). Modifikacijska varijabilnost prilično je raširena među živim organizmima. C. Darwin je to nazvao određenom varijabilnošću.
Za karakterizaciju kvantitativnih karakteristika koristi se skup statističkih pokazatelja. Jedan od tih pokazatelja je serija varijacija, koja karakterizira varijabilnost svojstva. Njegova prosječna vrijednost izračunava se pomoću formule:
gdje je X zbroj, v varijanta, p učestalost pojavljivanja varijante, n ukupan broj varijanti u nizu varijacija.
Za karakterizaciju stupnja varijabilnosti svojstva koristi se krivulja varijacije. Da biste to učinili, izgradite grafikon u koordinatnom sustavu, iscrtavajući na x-osi (vodoravno) vrijednost opcije v redoslijedom njihovog povećanja, na ordinatnoj (okomitoj) osi - učestalost pojavljivanja p svake opcije. Kada povežemo sjecišne točke, dobivamo krivulju koja karakterizira varijabilnost svojstva.
Genotipska varijabilnost
Genotipska, odnosno nasljedna varijabilnost obično se dijeli na kombinativnu i mutacijsku. Kombinativna varijabilnost je kombinacija gena u genotipu. Na primjer, roditelji s krvnom grupom I i II, te djeca s II i III:

Dobivanje novih kombinacija gena u genotipu s kombinativnom varijabilnošću postiže se kao rezultat tri procesa:
a) neovisna divergencija kromosoma tijekom mejoze,
b) njihova slučajna kombinacija tijekom oplodnje,
c) rekombinacija gena uslijed crossing overa.
Darwin je ustanovio da su mnoge sorte kultiviranih biljaka i životinjskih pasmina stvorene hibridizacijom već postojećih pasmina. Veliku važnost pridavao je kombinacijskoj varijabilnosti, smatrajući da ona, uz selekciju, igra važnu ulogu u stvaranju novih oblika kako u prirodi tako iu ljudskom gospodarstvu.
Kombinativna varijabilnost je široko rasprostranjena u prirodi. Mikroorganizmi koji se razmnožavaju nespolno razvili su jedinstvene mehanizme (transformacije i transdukcije) koji dovode do pojave kombinacijske varijabilnosti.
Kombinacijske varijacije mogu čak igrati ulogu u specijaciji. Opisane su vrste cvjetnica i riba koje kombiniraju karakteristike dviju blisko povezanih postojećih vrsta. Međutim, nastanak vrsta samo kao rezultat hibridizacije rijedak je fenomen.
Fenomen heterozisa susjedan je kombinacijskoj varijabilnosti.
Mutacijska varijabilnost nije uzrokovana rekombinacijom gena, već kršenjem njihove strukture. Charles Darwin (1859) je govorio o mogućnosti mutacija, nazivajući ih nesigurnom varijabilnošću ili pojedinačnim promjenama. Primijetio je iznenadnost njihove pojave.
Izraz "mutacija" predložio je 1889. G. De Friesom kako bi definirao promjene koje je uočio u šumi jasike. Primijetio je da ova biljka često doživljava promjene. Dakle, jedan mutant ima lišće i plodove s crvenim venama; drugi ima osjetno šire listove od izvornog oblika, samo ženske cvjetove,
a ne biseksualac; treći mutant je patuljaste veličine; četvrti je visok i ima velike cvjetove, plodove i sjemenke.
De Vries je u svom djelu "Teorija mutacije" (1901. - 1903.) formulirao glavne odredbe teorije mutacija:
Mutacija nastaje iznenada.
Novi oblici su prilično stabilni.
Mutacije su kvalitativne promjene.
Mutacije mogu biti korisne ili štetne.
Iste se mutacije mogu ponavljati.
Klasifikacija mutacija (vidi tablicu)
Primarnu ulogu imaju generativne mutacije koje se javljaju u spolnim stanicama. Ako je mutacija dominantna, tada se nova osobina ili svojstvo pojavljuje čak i kod heterozigotne jedinke. Ako je mutacija recesivna, tada se može pojaviti tek nakon nekoliko generacija kada postane homozigotna. Primjer generativne dominantne mutacije kod ljudi je pojava mjehurića na koži stopala, katarakte očiju i brahifalanksije (kratki prsti s insuficijencijom falangi). Primjer spontane recesivne generativne mutacije kod ljudi je hemofilija u pojedinim obiteljima.
Somatske mutacije su mutacije koje se javljaju u somatskim stanicama; zadržavaju se u potomstvu samo vegetativnim razmnožavanjem (pojava grane s bijelim bobicama na grmu crnog ribiza, bijeli pramen kose i različite boje očiju kod ljudi). Nasljeđe somatskih mutacija trenutno postaje važno za proučavanje uzroka raka kod ljudi. Pretpostavlja se da se kod malignih tumora transformacija normalne stanice u kancerogenu događa prema vrsti somatskih mutacija.
Genske ili točkaste mutacije najčešća su klasa mutacijskih promjena. To su citološki nevidljive promjene na kromosomima. Genske mutacije mogu biti dominantne ili recesivne. Primjer genske mutacije kod ljudi je vitamin D - rahitis otporan, metabolički poremećaj aminokiseline fenilalanin itd.
Molekularni mehanizmi genskih mutacija očituju se u promjenama redoslijeda parova nukleotida u molekuli nukleinske kiseline. Bit unutarnjih promjena može se svesti na četiri vrste preustroja nukleotida: a) zamjena para baza u molekuli DNA; b) delecija (gubitak) jednog para ili skupine baza u molekuli DNA; "
c) umetanje jednog para ili skupine parova baza u molekulu DNA;
d) preuređivanje položaja nukleotida unutar gena.
Promjene u molekularnoj strukturi gena dovode do novih oblika kopiranja s njega genetske informacije potrebne za odvijanje kemijskih procesa u stanici, što u konačnici dovodi do pojave novih svojstava u stanici i organizmu u cjelini. Za evoluciju su najvažnije točkaste mutacije. Prema njihovom učinku na prirodu kodiranih polipeptida, točkaste mutacije mogu se predstaviti u obliku tri klase:
1) mutacije koje nastaju zamjenom nukleotida unutar kodona, što uzrokuje zamjenu jedne netočne aminokiseline na određenom mjestu u polipeptidnom lancu. Fiziološka uloga proteina se mijenja, što stvara polje za prirodnu selekciju. Ovo je glavna klasa točkastih, intragenih mutacija koje se javljaju prirodnom mutagenezom pod utjecajem zračenja i kemijskih mutagena;
2) nonsens mutacije, tj. pojava terminalnih kodona unutar gena zbog promjena pojedinih baza unutar kodona. Kao rezultat toga, proces prevođenja završava na mjestu gdje se pojavljuje završni kodon. Ispostavilo se da je gen sposoban kodirati samo fragmente polipeptida do točke gdje se pojavljuje terminalni kodon;
3) očitavanje mutacija pomaka okvira događa se kada se unutar gena pojave insercije i delecije, te se cijeli semantički sadržaj gena promijeni. To je uzrokovano novom kombinacijom nukleotida u tripletima. Kao rezultat toga, cijeli polipeptidni lanac nakon točkaste mutacije dobiva druge netočne aminokiseline.
Kromosomske preraspodjele obično se klasificiraju kao mutacije, budući da je njihova prisutnost u stanicama povezana s promjenama u svojstvima tih stanica ili organizama koji nastaju iz tih stanica. Tamo su:
1) nedostatak dijela kromosoma (nedostatak i delecije). Delecija je gubitak (nedostatak) srednjeg dijela kromosoma zbog njegovog prekida na dvije točke. Ako je distalni, terminalni fragment otkinut, nedostatak se naziva nedostatak. Nedostatak je rijedak, jer nakon destrukcije distalne regije kromosom nije sposoban za daljnje postojanje. Nedostaci obično smanjuju sposobnost preživljavanja i plodnost jedinke;
2) udvostručenje ili umnažanje određenih dijelova kromosoma (duplikacija). Primjer umnožavanja je jačanje svojstva Vag (oči u obliku trake) kod Drosophile s povećanjem broja gena koji ga kontroliraju. Fenomen dupliciranja relativno je čest u prirodi i pripisuje mu se određena evolucijska uloga;
3) promjena linearnog rasporeda gena u kromosomu zbog inverzije pojedinih dijelova kromosoma za 180° (inverzija). Zanimljiv primjer inverzija su razlike u setovima kromosoma u obitelji mačaka. Svi njegovi predstavnici imaju 36 kromosoma, ali kariotipovi različitih vrsta razlikuju se u prisutnosti inverzija u različitim kromosomima. Inverzije dovode do promjena niza morfoloških i fizioloških karakteristika organizma i mogu biti čimbenik biološke izolacije populacije;
4) umetanje - pomicanje fragmenata kromosoma duž njegove duljine, zamjenjujući lokalizaciju gena.
Interkromosomski preraspodjeli povezani su s izmjenom regija između nehomolognih kromosoma. Takve se preraspodjele nazivaju translokacije. Teške translokacije mogu dovesti do oštrog smanjenja održivosti stanice i organizma u cjelini.
Mutacije koje utječu na genom stanice nazivaju se genomske. Do promjene broja kromosoma u genomu može doći zbog povećanja ili smanjenja broja haploidnih skupova ili pojedinačnih kromosoma. Organizmi u kojima su se umnožili cijeli haploidni skupovi nazivaju se poliploidima. Organizmi u kojima broj kromosoma nije višekratnik haploidnog broja nazivaju se aneuploidima ili heteroploidima.
Poliploidija je genomska mutacija koja se sastoji od povećanja broja kromosoma koji je višestruki od haploidnog broja. Stanice s različitim brojem haploidnih skupova kromosoma nazivaju se: 3G-triploid; 4d-tetraploid itd. Poliploidija dovodi do promjena u svojstvima organizma: stanice su velike i imaju povećanu plodnost. Poliploidija se pak dijeli na autopoliploidiju (povećanje broja kromosoma zbog umnažanja genoma jedne vrste) i alopoliploidiju (povećanje broja kromosoma zbog spajanja genoma različitih vrsta). Poliploidija je poznata kod životinja (trepetljikaši, valjkasti crvi, vodeni rakovi, svilene bube, vodozemci). Aneuploidija ili heteroploidija je promjena u broju kromosoma koja nije višekratnik haploidnog skupa kromosoma (primjerice 2n+\,2n- 1, 2n - 2, 2n + 2). Kod ljudi je to sindrom trisomije na X kromosomu ili na 21. kromosomu (Downov sindrom), monosomija na X kromosomu itd. Fenomen aneuploidije pokazuje da kršenje broja kromosoma dovodi do promjene strukture i održivost organizma.
Promjena u plazmogenima (bilo koja jedinica citoplazmatskog nasljeđa koja odgovara kromosomskom nasljeđu), koja dovodi do promjene karakteristika i svojstava organizma, naziva se citoplazmatske mutacije. Ove su mutacije stabilne i prenose se s koljena na koljeno (gubitak citokrom oksidaze u mitohondrijima kvasca).
Prema adaptivnoj vrijednosti mutacije se dijele na korisne, štetne (letalne i poluletalne) i neutralne. Ova podjela je uvjetna. Primjeri letalnih i polu-letalnih mutacija kod ljudi: epiloja (sindrom karakteriziran rastom kože, mentalnom retardacijom) i epilepsija, kao i prisutnost tumora srca, bubrega, amaurotična idiocija (taloženje masnih tvari u središnjem živčanom sustavu). sustava, praćeno degeneracijom moždane tvari, sljepoćom).
Mutacije koje se javljaju prirodno bez posebnog izlaganja neobičnim uzročnicima nazivaju se spontane. Obrasci spontane mutacije svode se na sljedeće točke:
1. Razina spontanih mutacija u gotovo svim živim organizmima je niska, ali učestalost mutacija varira među različitim vrstama životinja i biljaka.
2. Različiti geni u istom genotipu mutiraju različitim frekvencijama.
3. Slični geni u različitim genotipovima mutiraju različitim brzinama. Na primjer, u različitim laboratorijskim sojevima Drosophila, učestalost mutacija oka i krila nije ista; Mutacija albinizma je češća kod glodavaca nego kod drugih sisavaca.
4. Vrste i rodovi koji su genetski bliski karakteriziraju slični nizovi nasljedne varijabilnosti s takvom pravilnošću da se poznavajući nizove oblika unutar jedne vrste može predvidjeti prisutnost paralelnih oblika u drugim vrstama i rodovima. Što su rodovi i vrste bliže smješteni u općoj shemi, to je potpunija sličnost u nizu njihove varijabilnosti. “Cijele obitelji biljaka općenito karakterizira određeni ciklus varijacija koji prolazi kroz sve rodove i vrste koje čine obitelj.” Posljednju pravilnost otkrio je N. I. Vavilov 1920. On je istaknuo da homološke serije često nadilaze rodove, pa čak i obitelji. Kratki prsti uočeni su kod predstavnika mnogih redova sisavaca: goveda, ovaca, pasa, ljudi. Albinizam se uočava kod svih klasa kralješnjaka. Zakon homoloških serija izravno je povezan s proučavanjem nasljednih bolesti čovjeka. Mnoge mutacije pronađene kod životinja mogu poslužiti kao modeli za nasljedne ljudske bolesti. Dakle, psi imaju spolno povezanu hemofiliju. Albinizam je zabilježen kod mnogih vrsta glodavaca, mačaka, pasa i ptica. Miševi, goveda i konji mogu poslužiti kao modeli za proučavanje mišićne distrofije; epilepsija - zečevi, štakori, miševi; nasljedna gluhoća postoji kod zamoraca, miševa i pasa. Miševi boluju od nasljednih metaboličkih bolesti (pretilost, dijabetes).
Inducirani mutacijski proces shvaća se kao pojava nasljednih promjena pod utjecajem posebnog utjecaja čimbenika okoliša. Svi čimbenici mutageneze mogu se podijeliti u tri vrste: fizikalne, kemijske i biološke. Najučinkovitiji fizički mutagen je ionizirajuće zračenje. Prolazeći kroz stanice, X-zrake, gama-zrake, alfa-čestice i druga ionizirajuća radijacija na svom putu izbacuju elektrone iz vanjske ljuske atoma ili molekula, pretvarajući ih u pozitivno nabijene čestice. Ionizirajuće zračenje može imati izravan učinak na DNA i neizravan učinak preko ioniziranih molekula i atoma drugih tvari. Doza zračenja mjeri se u rentgenima ili radovima - vrijednostima koje su blizu apsolutne vrijednosti. Učestalost mutacija jako ovisi o dozi zračenja i njoj je izravno proporcionalna.
Inducirane mutacije? uzrokovane zračenjem prvi su eksperimentalno dobili sovjetski znanstvenici G. A. Nadson i G. S. Filippov, koji su 1925. promatrali proces mutacije kod kvasaca nakon izlaganja ionizirajućem zračenju. Godine 1927. američki genetičar G. Meller pokazao je da X-zrake mogu izazvati mnoge mutacije kod drozofila, a kasnije je mutageno djelovanje X-zraka potvrđeno na mnogim predmetima.
U fizičke mutagene spada i ultraljubičasto zračenje. Međutim, njegov mutageni učinak znatno je manji od djelovanja ionizirajućeg zračenja. Još slabije djeluje povišena temperatura, koja za toplokrvne životinje i čovjeka nema gotovo nikakvog značajnijeg značaja zbog postojanosti njihove tjelesne temperature."
Druga skupina čimbenika su kemijski mutageni. Kemijski mutageni uzrokuju uglavnom točkaste, odnosno genske mutacije, za razliku od fizičkih mutagena, koji znatno povećavaju vjerojatnost kromosomskih mutacija. Prioritet za otkrivanje kemijskih mutagena pripada sovjetskim istraživačima. Godine 1933 V. V. Sakharov dobio je mutacije djelovanjem joda, 1934. M. E. Lobashev - pomoću amonijaka. Godine 1946. sovjetski genetičar I. A. Rappoport otkrio je snažno mutageno djelovanje formalina i etilenimina, a engleski istraživač S. Auerbach snažno mutageno djelovanje iperita.
Biološki mutageni uključuju viruse i toksine niza organizama, osobito plijesni. Godine 1958. sovjetski genetičar S.I. Alikhanyan pokazao je da virusi uzrokuju mutacije u aktinomicetama. Nadalje, u domaćim i inozemnim laboratorijima utvrđeno je da su brojne dobro proučene mutacije kod životinja, biljaka i ljudi rezultat djelovanja virusa.
Ne mora se svako oštećenje DNK koje se dogodi nužno pretvoriti u mutaciju; ispravak se često događa uz pomoć posebnih enzima. Taj se proces naziva reparacija.
Trenutačno su poznata tri mehanizma popravka: fotoreaktivacija, tamni popravak i postreplikacijski popravak. Fotoreaktivacija uključuje eliminaciju dimera timina, koji su posebno česti u DNA pod utjecajem UV zraka, pomoću vidljive svjetlosti. Ovo je enzimski proces. Enzim, koristeći svjetlosnu energiju, cijepa dimere i tako obnavlja UV oštećenja u DNK faga i bakterija. Tamni popravak ne zahtijeva svjetlo. Sposoban je ispraviti širok raspon oštećenja DNK. Popravak tamne boje odvija se u nekoliko faza uz sudjelovanje nekoliko enzima:
Molekule prvog enzima (endonukleaza) neprestano ispituju molekulu DNA, prepoznajući oštećenje, enzim reže lanac DNA u blizini;
Drugi enzim (također endonukleaza ili egzonukleaza) čini drugi rez u ovoj niti, izrezujući oštećeno područje;
Treći enzim (egzonukleaza) značajno proširuje nastali jaz, odsijecajući desetke ili stotine nukleotida;
Četvrti enzim (polimeraza) popravlja prazninu prema redoslijedu nukleotida u drugom (neoštećenom) lancu DNA.
Prije nego što dođe do replikacije oštećenih molekula uočava se popravak u svjetlu i tami. Ako dođe do replikacije oštećenih molekula, tada se molekule kćeri mogu podvrgnuti postreplikacijskom popravku. Njegov mehanizam još nije jasan. Fenomen popravka DNK raširen je od bakterija do ljudi i važan je za održavanje stabilnosti genetskih informacija koje se prenose s generacije na generaciju.
Citoplazmatsko nasljeđe
Nasljedstvo, u kojem su materijalna osnova nasljeđivanja elementi citoplazme, naziva se nekromosomskim ili citoplazmatskim. Budući da je kod životinja i biljaka jajna stanica, a ne spermij, bogata citoplazmom, citoplazmatsko nasljeđivanje, za razliku od kromosomskog, mora se dogoditi po majčinoj liniji. Utemeljitelji citoplazmatskog nasljeđivanja bili su njemački genetičari K. Correns i Z. B a u r. Nasljedni faktori lokalizirani u citoplazmi i njenim organelama označavaju se pojmom plazmatip.
Ekstranuklearno nasljeđe se dijeli na:
I. Zapravo nekromosomsko ili citoplazmatsko nasljeđe:
Plastidno nasljeđe;
Nasljeđivanje preko mitohondrija;
Citoplazmatski muški sterilitet.
II. Predeterminacija citoplazme.
III. Nasljeđivanje infekcijom ili inkluzijama (pseudocitoplazmatsko nasljeđe).
1. Fenotip ovisi o:
a) od genotipa;
b) iz okoline;
c) ne ovisi ni o čemu;
d) iz genotipa i iz okoline.
2. Imaju usku reakcijsku normu... znakove.
kvaliteta;
b) kvantitativni.
3. Znakovi varijabilnosti izraženi su u obliku serije varijacija i krivulje varijacija:
a) mutacijski;
b) izmjena;
c) kombinatorni?
4. Do mutacije dolazi:
a) prilikom križanja;
b) tijekom prijelaza;
c) iznenada u DNK ili kromosomima.
5. Mutacije:
a) uvijek recesivan;
b) uvijek dominantan;
c) može biti dominantan ili recesivan.
6. Mutacije se manifestiraju fenotipski:
a) u svakom slučaju;
b) u homozigotnom organizmu;
c) u heterozigotnom organizmu.
7. Prema adaptivnoj vrijednosti mutacije mogu biti:
a) somatski;
b) polusmrtonosni, smrtonosni;
c) gen, odnosno točka.
8. Besmislene mutacije su:
a) mutacije koje nastaju zamjenom nukleotida unutar kodona;
b) pojava terminalnih kodona unutar gena;
c) čitanje mutacija pomaka okvira.
9. Maligni tumori mogu biti uzrokovani:
a) virusi;
b) kemikalije;
c) ionizirajuće zračenje;
d) virusi, kemikalije i ionizirajuće zračenje. _
10. Citoplazmatsko nasljeđivanje vrši:
a) po majčinoj liniji;
b) očinska linija.
Književnost
1. R.G. Zayats, I.V. Rachkovskaya i dr. Biologija za pristupnike. Minsk, Unipress, 2009., str. 578-597 (prikaz, ostalo).
2. L.N. Peseckaja. Biologija. Minsk, “Aversev”, 2007., str. 23-35.
3. N.D. Lisov, N.A. Lemeza i dr. Biologija. Minsk, “Aversev”, 2009., str. 33-37.
4. E.I. Shepelevich, V.M. Gluško, T.V. Maksimova. Biologija za školarce i kandidate. Minsk, “UniversalPress”, 2007., str. 37-50.
PREDAVANJE 16. Selekcija biljaka, životinja i mikroorganizama
Predmet i ciljevi selekcije.
Vavilov N.I. o podrijetlu kultiviranih biljaka.
Osnovne selekcijske metode. Heterozis, njegova upotreba u uzgoju.
Metode selekcije i genetski rad I. V. Michurina.
Dostignuća u uzgoju biljaka i životinja. Sorte kultiviranih biljaka koje su stvorili bjeloruski uzgajivači.
Glavni smjerovi su biotehnologija (mikrobiološka industrija, genetski i stanični inženjering).
Selekcija (od latinskog selectio - selekcija) je znanost o metodama stvaranja novih i poboljšanja postojećih sorti kultiviranih biljaka, pasmina domaćih životinja i sojeva mikroorganizama koje koristi čovjek.
Oplemenjivanje nove sorte ili pasmine je složen višefazni proces, koji se svodi na pažljiv odabir roditeljskih parova, njihovo križanje, metodičnu selekciju u hibridnom potomstvu, zatim križanje odabranih oblika i ponovno selekciju. U tom smislu, moderna znanost o uzgoju uključuje sljedeće glavne dijelove:
1. Proučavanje izvornog materijala - sortne, pasminske i specijske raznolikosti biljaka, životinja i mikroorganizama.
2. Razvoj metoda i sustava hibridizacije na temelju genetskih obrazaca nasljeđivanja svojstava i njihove varijabilnosti.
3. Razvoj metoda umjetne selekcije.
Pasmina, sorta ili soj je populacija organizama koju je umjetno stvorio čovjek i koja ima određene nasljedne karakteristike. Sve jedinke unutar pasmine, sorte ili soja imaju slična, nasljedno utvrđena svojstva, kao i istu vrstu reakcije na čimbenike okoliša. Na primjer, Leghorn pilići imaju malu težinu, ali visoku proizvodnju jaja.
Ciljevi suvremenog oplemenjivanja su povećanje produktivnosti biljnih sorti, pasmina domaćih životinja i sojeva mikroorganizama. U vezi s industrijalizacijom i mehanizacijom poljoprivrede, selekcija je usmjerena na stvaranje sorti žitarica, sorti vinove loze, rajčice, čajeva i pamuka, pogodnih za strojnu berbu, s niskim stabljikama koje ne polegu; uzgoj sorti povrća za uzgoj u staklenicima i hidroponima; stvaranje pasmina životinja za držanje u velikim stočarskim kompleksima.
Poboljšanje biljnih sorti, pasmina domaćih životinja i sojeva mikroorganizama nemoguće je bez proučavanja njihova podrijetla i evolucije. U tom smislu, rad N.I.Vavilova na proučavanju centara podrijetla kultiviranih biljaka je od posebne važnosti. Vavilov je organizirao niz ekspedicija, gdje je prikupio najvredniji materijal o rasprostranjenosti kultiviranih biljaka na kugli zemaljskoj. Vavilov je utvrdio da je rodno mjesto kukuruza Meksiko i Srednja Amerika, a rodno mjesto krumpira Južna Amerika. U Afganistanu je pronašao mnogo sorti meke pšenice, au Etiopiji - durum pšenice. Otkrio je i opisao 8 središta nastanka kultiviranih biljaka:
Indijski (južnoazijski tropski) - središte podrijetla sorti riže, šećerne trske, agruma;
Srednja Azija - meka pšenica, mahunarke i drugi usjevi;
kineski (ili istočnoazijski) - proso, heljda, soja, žitarice;
Zapadna Azija - pšenica i raž, kao i voćarstvo;
Mediteran - masline, djetelina, leća, kupus, krmno bilje;
Abyssinian - sirak, pšenica, ječam;
južnomeksički - pamuk, kukuruz, kakao, bundeva, grah;
Južna Amerika - središte krumpira, ljekovito bilje (grm koke, cinhona).
Poznavanje prirodnih uvjeta u postojbini pojedine biljke omogućuje isključivanje nekih besmislenih smjerova u selekciji i omogućuje provođenje selekcije na znanstvenoj osnovi, uzimajući u obzir svojstva kako samih biljaka tako i utjecaj okoliša. čimbenici na njihov nastanak. Vrijednost Vavilovljeva rada leži u činjenici da je postavio temelje za prikupljanje i pohranjivanje biljnih genofonda i stvorio prvu zbirku gena u svijetu u Svesaveznom institutu za biljke u Lenjingradu. Ova zbirka se svake godine nadopunjuje novim uzorcima koji dolaze sa svih kontinenata, a čak je i tijekom lenjingradske blokade potpuno očuvana. Trenutno pruža izvorni materijal za uzgajivače širom svijeta.
Osnovne selekcijske metode
| Metode | Uzgoj životinja | Uzgoj biljaka |
| Odabir roditeljskih parova | Po gospodarski vrijednim svojstvima i po eksterijeru (skup fenotipskih svojstava) | Prema mjestu podrijetla (geografski udaljeni) ili genetski udaljeni (nesrodni) |
| Hibridizacija: a) nesrodna (outbreeding) | Križanje udaljenih pasmina s kontrastnim karakteristikama radi dobivanja heterozigotnih populacija i manifestacije heterozisa. Rezultat je neplodno potomstvo. | Intraspecifično, interspecifično, međugeneričko križanje koje dovodi do heterozisa da bi se dobile heterotične populacije, kao i visoka produktivnost |
| b) u bliskom srodstvu (inbreeding) | Križanje između bliskih srodnika radi dobivanja homozigotnih (čistih) linija s poželjnim osobinama | Samooprašivanje u biljkama koje se međusobno oprašuju umjetnim utjecajem radi dobivanja homozigotnih (čistih) linija |
| Izbor a) masa | Nije primjenjivo | Pogodan za unakrsno oprašivanje biljaka |
| b) individualni | Stroga individualna selekcija koristi se za ekonomski vrijedne osobine, izdržljivost i eksterijer | Primijenjeno na samooplodne biljke, čiste linije su izolirane - potomci jedne samooplodne jedinke |
| Metoda ispitivanja očeva po potomstvu | Koriste se metodom umjetnog osjemenjivanja od najboljih mužjaka, čije kvalitete provjeravaju brojni potomci. | Nije primjenjivo |
| Eksperimentalna proizvodnja poliploida u | Nije primjenjivo | Koristi se u genetici i uzgoju za dobivanje produktivnijih, produktivnijih oblika |
Heterozis, njegova upotreba u uzgoju
Veća održivost i produktivnost F hibrida u odnosu na križane oblike izražava značenje fenomena heterozisa. "
Još sredinom 18.st. I. Kelreuter, akademik Ruske akademije, poznati botaničar, skrenuo je pozornost na činjenicu da su u nekim slučajevima, pri križanju biljaka, hibridi prve generacije mnogo snažniji od svojih roditeljskih oblika. Novo razdoblje u proučavanju fenomena heterozisa počinje 20-ih godina. prošlog stoljeća iz radova američkih genetičara Schella, Easta, Hell, Johnsona. Kao rezultat njihovog rada samooprašivanjem u kukuruzu dobivene su hibridne linije karakterizirane jakom depresijom. Ali kada je Shell međusobno ukrstio depresivne linije, neočekivano je dobio vrlo moćne F hibride. Ovim radovima započela je široka primjena heterozisa u procesu oplemenjivanja."
Genetičari su predložili nekoliko hipoteza za objašnjenje heterozisa. Hipotezu dominacije razvio je američki genetičar Johnson. Temelji se na prepoznavanju blagotvorno djelujućih dominantnih gena u homozigotnom ili heterozigotnom stanju:

Ako križani oblici imaju samo dva dominantna gena, tada ih hibrid ima četiri. To određuje heterozis hibrida, odnosno njegove prednosti u odnosu na izvorne oblike.
Hipotezu o naddominaciji predložili su američki genetičari Schell i East. Temelji se na spoznaji da biti heterozigot za jedan ili više gena daje prednost u odnosu na homozigote za jedan ili više gena.
Sovjetski genetičar V. A. Strunnikov predložio je hipotezu o kompenzacijskom kompleksu gena.
Značenje fenomena heterozisa je veća održivost i produktivnost F hibrida u odnosu na križane oblike.
Zbog učinka heterozisa dobiva se povećanje prinosa do 25-50 %.Smanjenje heterozisa u drugoj i sljedećim generacijama povezano je s cijepanjem hibridnog potomstva. U razvoju biljaka i životinja mogu se uočiti učinci koji u manifestaciji nalikuju heterozisu, ali nisu uzrokovani genetskim razlozima (ne kao rezultat križanja), već utjecajem određenih vanjskih utjecaja. To je takozvani fiziološki heterozis. Na primjer, za biljke su utvrđeni slučajevi uklanjanja štetnih posljedica samooprašivanja oštrom promjenom uvjeta rasta. Priroda takvih učinaka još nije jasna. Postoje tri vrste heterozisa:
Reproduktivni (hibridni potomci su superiorniji od roditeljskih oblika u plodnosti);
Somatski (kod hibridnog potomstva povećava se vegetativna masa kod biljaka i konstitucionalna snaga kod životinja);
Adaptivni (hibridi su bolje prilagođeni uvjetima okoline od roditeljskih oblika).
Metode selekcije i genetski rad I. V. Michurina
| Metode | Suština metode | Primjeri | |
| Biološki udaljena hibridizacija: a) interspecifična | Križanje predstavnika različitih vrsta za dobivanje sorti sa željenim svojstvima | Vladimirska trešnja x Winkler bijela trešnja - Ljepota sjeverne trešnje (dobar okus, zimska otpornost) | |
| b) međugenerički | Križanje predstavnika različitih rodova za dobivanje novih biljaka | Trešnja x ptičja trešnja = cerapadus | |
| Zemljopisno udaljena hibridizacija | Križanje predstavnika geografski udaljenih regija kako bi se hibridu usadile potrebne kvalitete (ukus, stabilnost) | Divlja Ussuri kruška x Bere royal (Francuska) = Bere zimska Michurina | |
| Metode | Suština metode | Primjeri | |
| Izbor | Višestruka, tvrda: po veličini, obliku, otpornosti na zimu, imunološkim svojstvima, kakvoći, okusu plodova i njihovoj očuvanosti | Na sjeveru su promovirane mnoge sorte stabala jabuka s dobrim okusom i visokim prinosom | |
| Mentorska metoda | Njegovanje poželjnih osobina u hibridnoj sadnici (povećanje dominacije), za što sadnicu na biljku cijepi učitelj od kojeg se te osobine žele postići. | Bellefleur kineski (hibridna podloga) x kineski (izdanak) = Bellefleur kineski (dugoročna sorta kasnog sazrijevanja) | |
| Medijatorska metoda | U udaljenoj hibridizaciji, korištenje divlje vrste kao posrednika za prevladavanje neukrštanja | Divlji mongolski badem x divlja Davidova breskva = srednji badem Kultivirana breskva x srednji badem = hibridna breskva (napredni sjever) | |
| Izloženost uvjetima okoline | Pri uzgoju mladih hibrida pazilo se na način skladištenja sjemena, prirodu i stupanj ishrane; izloženost niskim temperaturama, tlo siromašno hranjivim tvarima, česte transplantacije | ||
| Miješanje peludi | Za prevladavanje interspecifične nekompatibilnosti (nekompatibilnosti) | Pelud majčine biljke miješao se s peludom očeve, vlastiti pelud iritirao je stigmu, a ona je opažala strani pelud | |
Uspjesi sovjetske selekcije
Dostignuća uzgoja biljaka u SSSR-u povezana su s uporabom genetskih metoda u kombinaciji s metodama uzgoja. Tako su N.V. Tsin (1898-1980) i njegovi kolege, kao rezultat daljinske hibridizacije i selekcije, razvili visoko prinosne (do 70 c/ha) hibride pšenice i pšenične trave koji su bili otporni na polijeganje. Koristeći metodu složene hibridizacije zemljopisno udaljenih oblika praćene pažljivom individualnom selekcijom kroz više generacija, P. P. Lukjanenko je stvorio niz izvanrednih sorti ozime pšenice. Sorta pšenice Bezostaya-1 postala je široko rasprostranjena. Ova sorta ima visok prinos (65-70 ili više c/ha) u širokom rasponu ekoloških uvjeta: u sjevernom Kavkazu, južnoj Ukrajini, Moldaviji, Zakavkazju, u nekim područjima srednje Azije i Kazahstana, kao iu Mađarskoj. , Bugarska, Rumunjska, Jugoslavija.
P. P. Lukyanenko također je stvorio sorte pšenice (Aurora, Kavkaz), čiji prinos doseže gotovo 100 c/ha. V. N. Remeslov postigao je veliki uspjeh u selekciji ozime pšenice prilagođene širokom rasponu klimatskih i geografskih uvjeta u Mironovskom institutu za oplemenjivanje pšenice. Sorta Mironovskaya-808 koju je uzgojio bila je zonirana na gotovo cijelom teritoriju Ukrajinske SSR, u Moldavskoj i Bjeloruskoj saveznoj republici iu gotovo 50 regija RSFSR-a. Ovu sortu karakterizira visoka produktivnost (55-60 c/ha), zimska otpornost i dobra reakcija na gnojiva. Godine 1974. dobivena je sorta Ilyichevka (100 c/ha).
U selekciji proljetne pšenice najveće uspjehe postigli su u Istraživačkom institutu za poljoprivredu Jugoistoka (Saratov) izvrsni uzgajivači A. P. Shekhurdin i V. N. Mamontova.
Raznolikost pšenice Saratovskaya-29, koju su uzgajali ovi uzgajivači, uz visoku produktivnost, karakteriziraju izuzetne kvalitete pečenja.
Značajne uspjehe u oplemenjivanju suncokreta postigao je V. S. Pustovoyt, koji je započeo oplemenjivački rad sa sortama koje sadrže oko 30% ulja u sjemenu. Kao rezultat korištenja kontinuirane obiteljske grupne selekcije, uspio je dobiti sorte s udjelom ulja od 50 posto ili više. V. S. Pustovoit razvio je sorte suncokreta "Mayak" i "Peredovik". Zahvaljujući zoniranju ovih sorti, moguće je dobiti dodatne tisuće tona suncokretovog ulja uz isti trošak.
M. I. Xajinov 30-ih godina. prošlog stoljeća otkrili fenomen citoplazmatske muške sterilnosti. Na temelju hibridizacije i selekcije biljaka na citoplazmatski faktor razvijene su visokoprinosne sorte kukuruza.
Lutkov i Zosimovich povećali su sadržaj šećera i prinos šećerne repe zbog poliploidije. Veliki uspjeh postigli su i stočari. Križanjem je M. F. Ivanov stvorio visokoproduktivne pasmine (stepska bijela ukrajinska pasmina svinja, askanska finoruna pasmina ovaca). Na temelju hibridizacije divlje argali ovce s merino ovcom, zatim odabirom životinja željenog tipa i korištenjem inbreedinga u Kazahstanu, N. S. Baturin i njegovi kolege razvili su argali-merino pasminu, koja kombinira visokokvalitetnu vunu i visoku prilagodljivost argala. na uvjete visokoplaninskih pašnjaka . Metode hibridizacije i selekcije bile su osnova za stvaranje nove pasminske skupine svinja "Kazahstanski hibrid" (Institut za eksperimentalnu biologiju Kazahstanske SSR). Kemerovske svinje i divlje svinje korištene su kao početni oblici u procesu uzgoja ove pasminske skupine.
Dostignuća bjeloruskih uzgajivača
Znanstvenici Bjeloruskog istraživačkog instituta za krumpir i uzgoj voća i povrća (Samokhvalovichi, regija Minsk) od 1925. do 1995. godine. Uzgajano je oko 50 sorti krumpira, više od 70 povrća, 124 voća i 23 sorte bobičastog voća. Pod vodstvom i izravnim sudjelovanjem akademika P.I. Alsmika uzgojene su sorte krumpira kao što su Temp, Loshitsky, Ravaristy, Ogonyok, Sadko, Novinka, Verba, Ivushka, Lasunak, Zorka itd. 12 sorti krumpira s potencijalnim prinosom od 500-700 c/ha, otporan na bolesti i štetnike, visokih okusnih svojstava, pogodan za preradu u prehrambene poluproizvode. A.G. Voluznev razvio je 23 sorte bobičastog voća. Najčešće sorte crnog ribiza su Bjeloruski slatki, Cantata, Minai Shmyrev, Pamyat Vavilov. Princeza,
Katjuša, Partizanka; crveni ribiz - Voljeni; ogrozd - Proljeće, Ščedri; -jagode - Minskaja, Čajka.
Uzgajane su 124 sorte voćaka, uključujući 24 sorte jabuka (Antey, Belorusskoe malina, Bananovoe, Minskoe itd.), 8 sorti krušaka (Beloruska, Maslyanistaya, Loshitskaya itd.), 15 sorti trešanja (Zolotaya Loshitskaya , Krasavitsa) i mnogi drugi. Utemeljitelji bjeloruske selekcije voćaka su E. P. Syubarova i A. E. Syubarov. Uzgajivači jabuka nastavili su ono što su započeli - G. K. K ovalenko i E. V. Semashko, za kruške - N. I. M ihnevič, za trešnje - R. M. Sulimova, za šljivu - V. A. Matv e e v. Utemeljitelji selekcije povrtnih kultura su: G. I. Artemenko i A. M. Polyanskaya (rajčica), E. I. Chulkova (kupus), V. F. Devyatova (luk, češnjak).Oni su postavili i razvili temelje znanstvene selekcije povrće u Bjelorusiji.
Sorte povrtnih kultura bjeloruske selekcije su zonirane: rajčice otvorenog tla - Peramoga, Odlično, Dokhodny, Ruzha, Neman; rajčice za filmske staklenike - Vezha; krastavci - Dolzhik, Verasen, Zarnitsa; kupus - Rusinovka, Yubileynaya; luk - Amber, Vetraz; češnjak - Polet itd.
Osim toga, bjeloruski uzgajivači uzgojili su i zonirali mnoge sorte žitarica i mahunarki, industrijskih i krmnih biljaka. Na Bjeloruskom istraživačkom institutu za poljoprivredu (Žodino), N.D. Mukhiny je uzgojio tetraploidnu sortu ozime raži Belta. Autor je i koautor sorti ozime raži Beloruskaja 23, Družba, jare pšenice Minskaja, heljde Iskra i Jubilejnaja 2. Sorta ozime pšenice Berezina (74 c/ha) odlikuje se visokim svojstvima za mljevenje brašna i pečenjem. . Sorte za upotrebu u stočnoj hrani uključuju ozimu pšenicu Nadzeya (79 c/ha). Dobro su se dokazale sorte proljetnog ječma Zazersky 85 i Zhodinsky 5. Raznolikost žute lupine Narochansky odlikuje se visokom kvalitetom. Prinos zrna 27, zelena masa 536 c/ha. Sadržaj proteina u zrnu je 45,8%. Najpoznatije sorte šećerne repe su Belorusskaya single-55, polihibrid Belorussky-31 (uzgojen na Institutu za genetiku i citologiju Akademije znanosti Bjelorusije u suradnji s Ganusovskom eksperimentalnom oplemenjivačkom stanicom križanjem tetraploidnih i diploidnih oblika šećerna repa). Prosječni prinos korijenskih usjeva je 410-625 c/ha, sadržaj šećera je 15,3-19,5%, skupljanje šećera je 56,3-99,1 c/ha.
Glavna područja biotehnologije (mikrobiološki, genetski i stanični inženjering)
Biotehnologija je korištenje živih organizama i bioloških procesa u proizvodnji, tj. proizvodnja tvari potrebnih čovjeku korištenjem dostignuća mikrobiologije, biokemije i tehnologije.
Mikrobiološka industrija, koja se pojavila 60-ih godina prošlog stoljeća. XX. stoljeća rješava niz problema:
1) osigurava stoci potpunu bjelančevinu krmiva;
2) biotehnologijom se dobivaju i koriste enzimi (proteaza, amilaza, pektinaza);
3) korištenjem biotehnologije dobivaju se mikrobiološki proizvodi - aminokiseline, antibiotici (penicilini, cefalosporini, tetraciklini, eritromicini, streptomicini);
4) uz pomoć mikroorganizama dobivaju se dodatni izvori energije u obliku bioplina, etanola, vodika zbog korištenja industrijskog i poljoprivrednog otpada od strane mikroba.
Stanično inženjerstvo - metode uzgoja stanica u posebnim hranjivim podlogama.
Kultura tkiva je kultura stanica uzgojena u sterilnim uvjetima u posebnim podlogama. Stanične kulture (ili kulture tkiva) mogu poslužiti za proizvodnju vrijednih tvari. Na primjer, kultura stanica biljke ginseng proizvodi ljekovite tvari, kao i cijela biljka.
Stanične kulture također se koriste za hibridizaciju stanica. Posebnim tehnikama moguće je spojiti stanice različitog podrijetla iz organizama kod kojih je konvencionalna spolna hibridizacija nemoguća. Ovo otvara temeljno novi način stvaranja hibrida koji se temelji na kombiniranju somatskih, a ne zametnih stanica u jedan sustav. Već su dobivene hibridne stanice i organizmi krumpira i rajčice, jabuke i trešnje.
Kod životinja, proizvodnja hibridnih stanica također otvara nove mogućnosti, uglavnom za medicinu. Na primjer, u kulturi su dobiveni hibridi između stanica raka (koje imaju sposobnost neograničenog rasta) i nekih krvnih stanica - limfocita. Hibridomi su hibridi stanica raka i limfocita. Limfociti proizvode tvari koje osiguravaju imunitet (Imunitet) na zarazne bolesti, uključujući virusne bolesti. Korištenjem takvih hibridnih stanica moguće je dobiti vrijedne ljekovite tvari koje povećavaju otpornost organizma na infekcije.
Genetski inženjering je skup metoda koje omogućuju prijenos genetskih informacija s jednog organizma na drugi u epruveti. Prijenos gena omogućuje prevladavanje međuvrsnih barijera i prijenos pojedinačnih nasljednih karakteristika jednog organizma na drugi. Cilj genetskog inženjeringa je "dobiti stanice (bakterijske) sposobne preraditi određene "ljudske" proteine u industrijskim razmjerima. Tako se od 1980. ljudski hormon rasta - somatotropin - dobiva iz E. coli. Somatotropin je jedini lijek za djeca koja pate od patuljastog rasta zbog nedostatka ovog hormona.Prije razvoja genetskog inženjeringa izoliran je iz hipofize leševa.
Od 1982. inzulin za liječenje dijabetesa proizvodi se u industrijskim razmjerima iz E. coli koja sadrži gen za ljudski inzulin. Prije toga ovaj lijek nije bio dostupan svim pacijentima.
Najčešća metoda genetskog inženjeringa je metoda dobivanja rekombinantnih, tj. plazmida koji sadrže strani gen. Plazmidi su kružne dvolančane molekule DNK. Svaka bakterija, osim svoje osnovne DNA, sadrži nekoliko različitih plazmida, koje izmjenjuje s drugim bakterijama. To su plazmidi koji nose gene otpornosti na lijekove u bakterijama. Genetski inženjeri koriste plazmide za uvođenje gena viših organizama u bakterijske stanice.
Alat genetskog inženjeringa su enzimi otkriveni 1974. godine - restrikcijske endonukleaze, odnosno restrikcijski enzimi (doslovno restrikcija). Restrikcijski enzimi prepoznaju mjesta (mjesta prepoznavanja) i uvode simetrične prekide koji se nalaze koso jedni od drugih u lance DNK. Kao rezultat toga, na krajevima svakog fragmenta ograničene DNK formiraju se kratki jednolančani "repovi" koji se nazivaju "ljepljivi" krajevi.
Metode genetskog inženjeringa. Da bi se dobio rekombinantni plazmid, strani gen (na primjer, ljudski gen) uvodi se u razdvojeni plazmid. Enzim ligaza povezuje oba dijela DNK zajedno kako bi se formirao rekombinantni plazmid, koji se uvodi u E. coli. Svi potomci ove bakterije nazivaju se klon.
Cijeli proces dobivanja takvih bakterija, nazvan kloniranje, sastoji se od uzastopnih faza:
1. Restrikcija - rezanje ljudske DNK restrikcijskim enzimom na fragmente s “ljepljivim” krajevima.
2. Ligacija - uključivanje fragmenata ljudske DNA u plazmide zbog "šivanja ljepljivih krajeva" enzimom ligazom.
3. Transformacija - uvođenje rekombinantnih plazmida u bakterijske stanice. Međutim, plazmidi prodiru samo u dio tretiranih bakterija. Transformirane bakterije, zajedno s plazmidom, stječu otpornost na određeni antibiotik. To im omogućuje odvajanje od netransformiranih koji umiru na mediju koji sadrži antibiotik. Bakterije se siju na hranjivu podlogu i svaka od transformiranih bakterija se razmnožava i stvara koloniju od više tisuća potomaka - klon.
4. Probir - selekcija među klonovima transformiranih bakterija onih koje sadrže plazmide sa željenim ljudskim genom. Bakterijske kolonije prekrivene su posebnim filtrom, nakon uklanjanja na njemu ostaje otisak kolonija. Zatim se provodi molekularna hibridizacija. Filtri su uronjeni u otopinu koja sadrži radioaktivno obilježenu sondu. Proba je polinukleotid komplementaran dijelu željenog gena. Hibridizira se samo s onim rekombinantnim plazmidima koji sadrže željeni gen. Nakon hibridizacije, rendgenski fotografski film se stavlja na filtar u mraku i razvija nakon nekoliko sati. Položaj osvijetljenih područja omogućuje pronalaženje plazmida sa željenim genom.
1. Interspecifični hibridi:
a) su neplodni;
b) odlikuju se povećanom plodnošću;
c) uvijek žensko;
d) uvijek muško.
2. Metoda koja se koristi u selekciji i nije popraćena promjenom genetskih svojstava organizama je:
a) poliploidija; b) umjetna mutageneza;
c) hibridizacija; d) kloniranje.
3. Domovina kukuruza i suncokreta je središte podrijetla kultiviranih biljaka (prema N. I. Vavilovu):
a) južnoazijski;
b) srednjoamerički;
c) abesinska;
d) Sredozemlje.
4. Sama poliploidija ili euploidija je:
a) preuređenje kromosoma;
b) promjena broja kromosoma, višekratnik haploidnog broja;
c) promjena slijeda nukleotida.
5. Po prvi put je bilo moguće razviti načine za prevladavanje neplodnosti interspecifičnih hibrida:
a) K. A. Timirjazev; b) M. F. Ivanov; c) G. D. Karpečenko; d) N. S. Butarin.
6. Koji se oblik umjetne selekcije koristi u uzgoju životinja:
a) masivan;
b) individualni?
7. U XX. stoljeću. Metodama odabira dodano je sljedeće:
a) poliploidija;
b) umjetna mutageneza;
c) hibridizacija stanica;
d) poliploidija, umjetna mutageneza i hibridizacija stanica.
8. Homogena skupina biljaka gospodarski vrijednih svojstava koju je stvorio čovjek naziva se:
a) klon;
b) pasmina;
c) raznolikost.
9. Biotehnologija je:
a) korištenje proizvoda biološkog podrijetla (treseta, ugljena, nafte) za pogon strojeva i mehanizama;
b) korištenje tehnologije u stočarstvu i biljnoj proizvodnji;
c) korištenje živih organizama i bioloških procesa u proizvodnji;
d) korištenje živih organizama kao modela pri stvaranju različitih struktura i mehanizama.
Književnost
1. R.G. Zayats, I.V. Rachkovskaya i dr. Biologija za pristupnike. Minsk, Unipress, 2009., str. 674-686 (prikaz, ostalo).
2. L.N. Peseckaja. Biologija. Minsk, “Aversev”, 2007., str. 72-85.
3. E.I. Shepelevich, V.M. Gluško, T.V. Maksimova. Biologija za školarce i kandidate. Minsk, “UniversalPress”, 2007., str.95-104.
PREDAVANJE 17. Postanak i razvoj života na Zemlji.
Evolucijska doktrina Charlesa Darwina
Prva evolucijska teorija J. B. Lamarcka.
Preduvjeti za nastanak darvinizma.
Bit evolucijske doktrine Charlesa Darwina i njezin utjecaj na biološke znanosti.
Sintetička teorija evolucije života.
Zasluga za stvaranje prve holističke teorije o evoluciji organskog svijeta pripada J. B. Lamarcku (1744.-1829.). Glavne odredbe ove teorije iznio je u svom djelu "Filozofija zoologije" (1809). Lamarck je postulirao sljedeća načela:
Organizmi su promjenjivi;
Vrste (i druge taksonomske kategorije) su privremene i postupno se transformiraju u nove vrste;
Opći trend povijesnih promjena u organizmima je postupno usavršavanje njihove organizacije (gradacija), čiji je pokretač izvorna (od tvorca pokrenuta) želja za napretkom;
Organizmi imaju urođenu sposobnost brzog reagiranja na promjene u vanjskim uvjetima;
Promjene u organizmima stečene tijekom života kao odgovor na promjenjive uvjete nasljeđuju se.
Ocjena Lamarckove teorije. Lamarckova izvanredna zasluga leži u stvaranju prve evolucijske doktrine. Odbacio je ideju postojanosti vrsta, suprotstavljajući je ideji promjenjivosti vrsta. Njegovo učenje tvrdilo je postojanje evolucije kao povijesnog razvoja od jednostavnog prema složenom. Prvi put se postavilo pitanje čimbenika evolucije. Lamarck je potpuno ispravno vjerovao da uvjeti okoliša imaju važan utjecaj na tijek evolucijskog procesa. Bio je jedan od prvih koji je ispravno procijenio važnost vremena u procesu evolucije i primijetio izvanredno trajanje razvoja života na Zemlji: Međutim, Lamarck je napravio ozbiljne pogreške, prije svega, u razumijevanju čimbenika evolucijskog procesa. , deducirajući ih iz navodno? inherentna želja za savršenstvom u svim živim bićima. Također je krivo shvatio razloge za nastanak fitnessa, izravno ih povezujući s utjecajem okolišnih uvjeta.
Lamarckova evolucijska doktrina nije bila dovoljno uvjerljiva i nije dobila široko priznanje među njegovim suvremenicima.
Darwin je 1859. u knjizi “Podrijetlo vrsta prirodnim odabirom ili očuvanje omiljenih pasmina u borbi za život” iznio osnovna načela te teorije. Svih 1250 primjeraka knjige prodano je prvog dana, a navodno je po utjecaju na ljudsko razmišljanje odmah iza Biblije. Zatim je razvio teoriju u sljedećim djelima, “Promjene na životinjama i biljkama pod utjecajem pripitomljavanja” (1868.) i “Porijeklo čovjeka i spolni odabir” (1871.). Do sličnih je zaključaka neovisno o Darwinu došao engleski zoolog A. Wallace (1858). Naziv "darvinizam" predložio je T. Geksl (1860).
Osnovna načela evolucijskog učenja Charlesa Darwina:
1. Svaka je vrsta sposobna za neograničeno razmnožavanje.
2. Ograničeni vitalni resursi sprječavaju neograničenu reprodukciju. Većina jedinki umire u borbi za opstanak i ne ostavlja potomke.
3. Smrt ili uspjeh u borbi za opstanak su selektivni. Charles Darwin je selektivno preživljavanje i razmnožavanje najprilagođenijih organizama nazvao prirodnom selekcijom.
4. Pod utjecajem prirodne selekcije koja se odvija u različitim uvjetima, skupine jedinki iste vrste akumuliraju različite adaptivne karakteristike iz generacije u generaciju. Skupine jedinki postižu tako značajne razlike da se pretvaraju u nove vrste (načelo divergencije karaktera).
Na temelju Darwinova učenja utvrđeno je da su pokretačke snage evolucije organskog svijeta borba za opstanak i prirodna selekcija temeljena na nasljednoj varijabilnosti, a pokretačke snage evolucije pasmina i sorti nasljedna varijabilnost i umjetna selekcija.
Pod naslijeđem je Darwin razumio sposobnost organizama da u potomstvu sačuvaju svoje vrste, sortne i individualne osobine, a pod varijabilnošću - sposobnost organizama da pod utjecajem okolišnih uvjeta stječu nova svojstva. Razlikovao je određenu, neodređenu i relativnu varijabilnost.
Određena (ili grupna) varijabilnost je manifestacija sličnih karakteristika kod svih jedinki pod utjecajem istih uvjeta okoline. Sada je utvrđeno da ta varijabilnost ne utječe na genotip organizama i naziva se modifikacija ili fenotapija.
Neodređena (ili individualna) varijabilnost je pojava individualnih razlika kod jedinki iste vrste. Individualne razlike se nasljeđuju. Ovo je genetska ili nasljedna varijacija.
Nadalje, Darwin je identificirao korelativnu varijabilnost, kada promjena u jednom organu ili osobini povlači za sobom promjene u drugim organima ili osobinama. Na primjer, dugonoge životinje imaju duge vratove. Kod sorti stolne repe dosljedno se mijenja boja korijena, peteljki i lisnih žila.
Borba za opstanak po Darwinu predstavlja složene i raznolike odnose organizama između njih samih i nežive prirode. Postoje različiti oblici borbe za egzistenciju: unutarvrsna, međuvrsna i borba protiv nepovoljnih uvjeta.
Intraspecifična (konkurencija). Rezultat je očuvanje populacije i vrste zbog smrti slabijih. Pobjeda održivije populacije nad manje održivom populacijom koja zauzima istu ekološku nišu. Primjeri: natjecanje između predatora iste populacije za plijen; intraspecifični kanibalizam - uništavanje mladih životinja kada je veličina populacije prekomjerna; borba za prevlast u čoporu; jednodobna borova šuma.
Dakle, sve vrste borbe za opstanak u konačnici dovode do opstanka onih organizama koji se pokažu najprilagođenijim određenim uvjetima, odnosno prirodnoj selekciji.
Prirodna selekcija je proces koji se neprestano odvija u prirodi, u kojem najsposobnije jedinke svake vrste prežive i ostavljaju potomstvo, a manje sposobne umiru. Neophodan uvjet za prirodnu selekciju je nasljedna varijabilnost, a neposredni rezultat je stvaranje prilagodbi organizama na specifične uvjete postojanja. Klasičan primjer prirodne selekcije je promjena boje brezinog moljca. Postoji pokretačka, stabilizirajuća i disruptivna (kidajuća) prirodna selekcija.
Pogonska ili usmjerena selekcija je selekcija koja favorizira samo jedan smjer varijacije (slika 4). Opisao ga je Darwin. Na primjer, trenutna pojava skupina štakora i insekata koji su otporni na pesticide; sojevi mikroorganizama otporni na antibiotike.
Stabilizirajuća selekcija je selekcija čiji je cilj očuvanje prosječnog, prethodno utvrđenog svojstva u populacijama i djelovanje protiv manifestacija fenotipske varijabilnosti (slika 5). Opisao I.I.Shmalhausen 1946. Na primjer, veličina i oblik cvjetova u biljkama koje se oprašuju kukcima su stabilniji nego u biljkama koje se oprašuju vjetrom (struktura cvjetova biljaka koje se oprašuju kukcima odgovara strukturi kukaca oprašivača). Potomstvo ostavljaju samo one biljke čija se struktura cvijeta ne mijenja. Ptice imaju prosječnu duljinu krila.

Riža. 4. Pogonski oblik prirodne selekcije: A - D - sukcesivne promjene brzine reakcije pod pritiskom pokretačke sile prirodne selekcije.
Disruptivna ili diskontinuirana selekcija je selekcija koja favorizira dva ili više smjerova varijabilnosti u organizmima. Na primjer, kukci na oceanskim otocima (slika 6). Disruptivna selekcija usmjerena je protiv održavanja prosječne vrijednosti svojstva. Ovaj oblik selekcije opisao je K. Mazer (1973).

Riža. 5. Stabilizirajući oblik prirodne selekcije

Slika: 6. Disruptivni oblik prirodne selekcije
Svi oblici prirodne selekcije čine jedinstven mehanizam koji održava ravnotežu populacije s okolišem. Selekcija počinje unutar populacije.
Osnovna evolutivna jedinica je populacija, jer samo ona predstavlja ekološku, morfofiziološku i genetsku cjelinu. Zbirka gena u populaciji naziva se genski fond. U velikim populacijama, gdje nema mutacija, selekcije i miješanja s drugim populacijama, uočava se konstantnost frekvencija alela, homo- i heterozigota (Hardy-Weinbergov zakon).
Svi čimbenici koji uzrokuju odstupanje od Hardy-Weinbergovog zakona su elementarni evolucijski čimbenici. To uključuje: mutacije, prirodnu selekciju, populacijske valove i izolaciju. Mutacije se neprestano javljaju u populacijama pod utjecajem mutagenih čimbenika okoliša i uzrokuju promjene u njezinom genskom fondu. Populacijski valovi su pravilne fluktuacije u broju populacije povezane s periodičkim promjenama intenziteta čimbenika okoliša. Populacijski valovi značajno pospješuju prirodnu selekciju i mijenjaju učestalost geni u populacijama.Izolacija dovodi do divergentnosti svojstava unutar jedne vrste i sprječava međusobno križanje jedinki iz različitih populacija i vrsta.Razlikuju se geografska, ekološka i biološka izolacija.
Sintetička teorija evolucije
Prvi koji je objedinio podatke genetike i darvinizma bio je ruski zoolog i komparativni anatom N. K. Koltsov (1872.-1940.). Njegov učenik i kolega S. S. Chetverikov (1880.-1959.) prvi je dao genetsku osnovu za Darwinova učenja o evoluciji. U poznatom djelu S. S. Chetverikova “O nekim aspektima evolucijskog procesa s gledišta moderne genetike” (1926.) pokazano je da u prirodnim uvjetima u prirodi unutar svake vrste postoji ogroman broj nasljednih promjena koje ne manifestiraju se fenotipski zbog recesivnosti. Vrsta je zasićena mutacijama, koje predstavljaju neiscrpan materijal za evoluciju.
Počeo u 20-ima. prošlog stoljeća, ujedinjenje darvinizma i genetike pridonijelo je širenju i produbljivanju sinteze darvinizma s drugim znanostima; 30-40-ih godina Smatra se razdobljem nastanka sintetičke teorije evolucije (STE).
Važnu ulogu u formiranju STE ima rad F. G. Dobzhanskog “Genetika i podrijetlo vrsta” (1937.), koji je sažeo sintezu genetike s darvinizmom. Značajan doprinos stvaranju STE dao je sovjetski znanstvenik I. I. Shmalgauzen (1887.-1963.). Istraživao je odnos između ontogeneze i filogeneze, proučavao glavne smjerove evolucijskog procesa i identificirao dva oblika prirodne selekcije. Njegova djela su “Putovi i uzorci evolucijskog procesa” (1939.), “Čimbenici evolucije” (1946.).
(1940.), objavljivanjem knjiga E. Mayra “Sustavnost i podrijetlo vrsta” (1944.) i J. Huxleya “Evolucija: Moderna sinteza” (1942.). Izraz "sintetička teorija evolucije" svoje ime duguje naslovu knjige J. Huxleya "Evolucija: Moderna sinteza".
Osnovni postulati STE
1. Materijal za evoluciju su u pravilu vrlo male, ali diskretne promjene nasljeđa – mutacije.
2. Proces mutacije, valovi brojeva - dobavljači materijala za selekciju - slučajni su i neusmjereni po prirodi.
3. Jedini vodeći čimbenik evolucije je prirodna selekcija, temeljena na očuvanju i gomilanju nasumičnih i malih mutacija.
4. Najmanja evolucijska jedinica je populacija, a ne jedinka, kako se pretpostavljalo na temelju ideja o mogućnosti “nasljeđivanja stečenih karakteristika”. Otuda posebna pažnja proučavanju populacije kao elementarne strukture jedinice vrste.
5. Evolucija je divergentne prirode, to jest, jedan takson može postati predak nekoliko svojti kćeri, ali svaka vrsta ima jednu vrstu predaka, jednu populaciju predaka.
6. Evolucija je postupna i dugotrajna. Specijacija kao faza evolucijskog procesa je sekvencijalna zamjena jedne privremene populacije nizom sljedećih privremenih populacija.
 Zbirka ruskih čudaka i modifikatora tijela
Zbirka ruskih čudaka i modifikatora tijela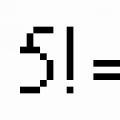 Teorija vjerojatnosti i matematička statistika
Teorija vjerojatnosti i matematička statistika Osnovni pojmovi teorije vjerojatnosti i matematičke statistike
Osnovni pojmovi teorije vjerojatnosti i matematičke statistike