Словник ґендерних термінів. Еволюційна теорія статі В
І використовується в різних галузях наукової діяльності: еволюції статевого розмноження, біології рослин і тварин, медицини, соціальної психології, педагогіки та інших.
Поділ на дві статі це спеціалізація зі збереження та зміни інформації у популяції. Одна підлога має бути інформаційно більш тісно пов'язана із середовищем, і бути більш чутливою до її змін. Підвищена смертність чоловічої статі від усіх факторів середовища дозволяє вважати її оперативноюекологічну підсистему популяції. Жіноча стать, як більш стабільна, є консервативноїпідсистемою та зберігає існуючий розподіл генотипів у популяції.
У еволюції статі різних стадіях і рівнях організації виник цілий ряд механізмів, які послідовно забезпечували тісніший зв'язок жіночої статі з генеративним (консервативним) потоком, а чоловічого - з екологічним (оперативним). Так, у чоловічої статі, порівняно з жіночою, вища частота мутацій, менша адитивність успадкування батьківських ознак, вже норма реакції, вища агресивність і допитливість, активніша пошукова, ризикована поведінка та інші якості, що «наближають до середовища». Усі вони, цілеспрямовано виносячи чоловічу стать на периферію розподілу, забезпечують йому переважне отримання екологічної інформації.
Інша група особливостей - величезна надмірність чоловічих гамет, їх малі розміри та висока рухливість, велика активність та мобільність самців, їх схильність до полігамії та інші етолого-психологічні властивості. Тривалі періоди вагітності, годування і турботи про потомство у самок, практично підвищуючи ефективну концентрацію чоловічих особин, перетворюють чоловічу стать на «надлишковий», отже, «дешевий», а жіночий - на дефіцитний і більш цінний.
В результаті консервативно-оперативної спеціалізації статей відбувається їх асинхронна еволюція: нові ознаки з'являються спочатку в оперативній підсистемі (чоловіча стать) і лише потім потрапляють до консервативної (жіноча стать).
Чоловіча стать залишається в небезпечних зонах і піддається дії відбору. Після дії відбору зменшується частка чоловічих особин і звужується їхня генотипна дисперсія. У рушійному середовищі перетворення торкаються дисперсії статей і середні значення ознаки: норма реакції створює тимчасовий, фенотипічний статевий диморфізм, відбір - генотипічний. Чоловіча стать отримує нову екологічну інформацію. Підвищення смертності чоловічої статі збільшує народжуваність чоловічих особин за рахунок негативного зворотного зв'язку.
Статевий процес та статева диференціація діють у протилежних напрямках: перший збільшує різноманітність генотипів, а друга – мінімум вдвічі погіршує його. Тому називати «неоднакову» пару гомологічних хромосом (ХY, ZW) «статевими» лише через те, що вони визначають стать, не зовсім коректно. Набагато більше підстав вважати їх «антистатевими», оскільки саме вони погіршують головне досягнення статі – комбінаторику ознак. Основна роль статевих хромосом - еволюційна,- створення двох зрушених за часом форм (жіночої та чоловічої), для економної еволюції.
Підлогу зиготи визначають при зачатті статеві хромосоми. Далі, до кінця онтогенезу, підлогою управляють статеві гормони. У ссавців базова підлога - гомогаметна (XX) - жіноча; а похідна підлога - гетерогаметна (XY) - чоловіча. Він запускається Y-хромосомою, що перетворює «безстатеві» зачатки гонад ембріона на сім'яники, що продукують андрогени. За відсутності Y-хромосоми, ті ж тканини перетворюються на яєчники, що продукують естрогени. У птахів базова підлога теж гомогаметна (ZZ), але - чоловіча; а похідна жіноча стать має гетерогаметну конституцію (ZW). Він запускається W-хромосомою, що перетворює зачатки на яєчники, що продукують естрогени. За відсутності W-хромосоми, ті ж тканини перетворюються на сім'яники, які продукують андрогени. Тобто, у ссавців андрогени зрушують самців від самок до середовища, а у птахів – естрогени видаляють самок від самців та середовища. В обох випадках, чоловіча стать є «середовищною», а жіноча - «системною». Статеві гормони визначають розвиток не тільки ознак статевої диференціації (статевий диморфізм), але й асиметрії мозку, рук та інших частин тіла (латеральний диморфізм). Естрогени, розширюючи норму реакції, дозволяють жіночим фенотипам залишити зони відбору та зберегтися. Вони діють «відцентрово», видаляючи та ізолюючи систему від середовища. Андрогени - їх хімічні антагоністи, діють, навпаки - «відцентрово», наближаючи систему до середовища, піддаючи його більш інтенсивному дії відбору та прискорюючи еволюцію. Отже, андроген-естрогенне співвідношення регулює інтенсивність інформаційного контакту системи із середовищем.
Еволюційна теорія статі розглядає підвищену смертність чоловічої статі, як вигідну популяції форму інформаційного контакту з середовищем, здійснювану через елімінацію шкідливим чинником середовища частини особин популяції. Наприклад, усі «нові» хвороби, хвороби «століття» чи «цивілізації» (інфаркт, атеросклероз, гіпертонія та ін.), як правило, хвороби чоловічої статі.
У мінливих, екстремальних умовах середовища, підвищується смертність чоловічої статі та падає третинне співвідношення статей популяції. Чим мінливіше середовище, тим менше залишається у популяції чоловічих особин і водночас, тим більше їх потрібно для пристосування. Компенсувати зниження третинного співвідношення статей можна лише підвищивши вторинне. Іншими словами, в екстремальних умовах середовища одночасно підвищуватиметься і смертність і народжуваність чоловічих особин, тобто зростатиме їхня «оборотність».
Негативний зворотний зв'язок реалізується у рослин через кількість пилку, а у тварин через інтенсивність статевої діяльності, старіння, спорідненість і загибель гамет. При цьому мала кількість пилку, інтенсивна статева діяльність самців, свіжа сперма і старі яйцеклітини повинні призводити до збільшення народжуваності чоловічих особин.
Для реалізації популяційного механізму необхідно, щоб ймовірність мати нащадка даної статі відрізнялася у різних особин і визначалася їх генотипом. При цьому повинна існувати зворотна залежність між репродуктивним рангом даної особини та статтю її потомства: чим вищий репродуктивний ранг, тим більше має бути нащадків протилежної статі. У цьому випадку регуляція може здійснюватися на популяційному рівні - більшою або меншою участю в розмноженні особин, що дають у потомстві надлишок самців або самок.
Кожному нащадку батько і мати передають приблизно однакову кількість генетичної інформації, але кількість потомства, якому може передати генетичну інформацію самець, незрівнянно більша за кількість, якій може передати інформацію самка. Кожен самець, в принципі, може передати інформацію всьому потомству популяції, тоді як самки такої можливості позбавлені. Тобто, пропускна здатність – «перетин» – каналу зв'язку самця з потомством значно більша, ніж переріз каналу зв'язку самки.
У строго моногамної популяції число батьків і матерів рівне, тобто самці і самки мають однакове «перетин каналу» зв'язку з потомством. У разі полінії, коли батьків менше, ніж матерів, самці мають більше «перетин» каналу зв'язку. У разі поліандрії – все навпаки.
Широка норма реакції робить жіночу стать більш мінливою і пластичною в онтогенезі. Вона дозволяє особам жіночої статі залишити зони елімінації та дискомфорту, зібратися у зоні комфорту та зменшити фенотипічну дисперсію та смертність.
Вужча норма реакції чоловічої статі не дозволяє йому зменшити фенотипічну дисперсію. Чоловічі особини залишаються в зонах елімінації та дискомфорту та гинуть, або не залишають потомство. Це дозволяє населенню за нову інформацію «платити» насамперед жертвою чоловічих особин.
Висока онтогенетична пластичність жіночої статі забезпечує високу стабільність у філогенезі. Серед поколінь, жіноча стать найповніше зберігає наявний у популяції розподіл генотипів. Генотипічний розподіл чоловічої статі змінюється набагато сильніше. Отже, у філогенетичному плані більш мінлива, пластична чоловіча стать, а в онтогенетичному - навпаки, більш пластична і мінлива жіноча. Такий, на перший погляд парадоксальний, розподіл ролей у філогенезі та онтогенезі, насправді, послідовно та несуперечливо реалізує ідею спеціалізації статей з консервативної та оперативної задач еволюції.
У стабільному середовищі всі перетворення генетичної інформації торкаються дисперсії статей, але не торкаються середніх значень ознак. Тому статевий диморфізм відсутній. Виникає лише різниця у дисперсії, яка при переході до наступного покоління зникає. Однак, необхідно, щоб генотипічний статевий диморфізм за нормою реакції існував заздалегідь (у стабільній фазі), причому генетична інформація про широку норму реакції повинна передаватися тільки по жіночій лінії, а про вузьку - тільки по чоловічій.
У рушійному середовищі фенотипічний розподіл чоловічої статі, до дії відбору, приблизно повторює вихідний генотипний розподіл. Широка норма реакції жіночої статі призводить до зсуву розподілу фенотипів та до появи тимчасового – фенотипного – статевого диморфізму. Жіноча стать залишає зони відбору та дискомфорту, і зберігає спектр минулих генотипів. Виникла різниця між чоловічими та жіночими гаметами частково зберігається і після запліднення, оскільки інформація, що передається через Y-хромосому, ніколи не потрапляє від батька до дочки. На користь того, що частина генетичної інформації залишається в чоловічій підсистемі і не потрапляє в жіночу, свідчить також існування реципрокних ефектів - те, що при гібридизації не байдуже з якоїсь породи батько, а з якої матері.
Отже, різні переріз каналу та норма реакції чоловічої та жіночої статі, у рушійному середовищі, неминуче приводять, вже в одному поколінні, до виникнення генотипного статевого диморфізму. У наступних поколіннях, у рушійному середовищі, він може накопичуватися та зростати.
Якщо перейти до філогенетичного масштабу часу, то у роздільностатевих форм, після зміни стабілізуючого середовища на рушійну, протягом багатьох поколінь ознака змінюється лише у чоловічої статі. У жіночої статі зберігається старе значення ознаки. Траєкторія еволюції ознаки роздвоюється на чоловічу та жіночу гілки, відбувається «розбіжність» ознаки у двох статей – поява та зростання генотипічного статевого диморфізму. Це - дивергентнафаза, в якій швидкість еволюції ознаки більша у чоловічої статі.
Через деякий час, коли вичерпуються можливості норми реакції та інших механізмів захисту жіночої статі, ознака починає змінюватись і в неї. Генотипічний статевий диморфізм, досягнувши свого оптимуму, залишається незмінним. Це - стаціонарнафаза, коли швидкості еволюції ознаки у чоловічої та жіночої статі рівні. Коли у чоловічої статі ознака досягає нового еволюційно-стабільного значення, у жіночої статі він продовжує змінюватися. Це - конвергентнафаза еволюції ознаки, коли швидкість її більша у жіночої статі. Генотипічний статевий диморфізм поступово зменшується і при злитті ознак у двох статей зникає. Отже, фази еволюції ознаки у чоловічої та жіночої статі зрушені у часі: у чоловічої статі вони починаються і закінчуються раніше, ніж у жіночої.
Оскільки еволюція ознаки завжди починається з розширення його генотипної дисперсії і закінчується її звуженням, то дивергентної фазі дисперсія ширше в чоловічої статі, а конвергентної - в жіночого. Значить, за статевим диморфізмом і дисперсією статей можна судити про напрям і фазу еволюції ознаки.
Всі ознаки можна розділити на три групи за ступенем розходження між статями.
До першої групи віднесемо ті ознаки, за якими між чоловічою та жіночою статтю немає жодної різниці. До них відносяться якісні ознаки, які проявляються на рівні виду - загальний для обох статей план та принципова будова тіла, кількість органів та багато інших. Статевий диморфізм за цими ознаками в нормі відсутня. Але він спостерігається у галузі патології. У дівчаток частіше виявляються атавістичні аномалії (повернення або зупинки розвитку), а у хлопчиків – футуристичні (пошук нових шляхів). Наприклад, серед 4000 новонароджених дітей із трьома нирками дівчаток було у 2.5 рази більше, ніж хлопчиків, а серед 2000 дітей з однією ниркою було приблизно у 2 рази більше за хлопчиків. Нагадаємо, що у наших далеких предків у кожному сегменті тіла була пара виділених органів – метанефридіїв. Отже, три нирки у дівчаток – це повернення до предкового типу (атавістичне спрямування), а одна нирка у хлопчиків – футуристична тенденція. Така ж картина спостерігається серед дітей із наднормативним числом ребер, хребців, зубів тощо, тобто органів, які зазнали в процесі еволюції зменшення числа – серед них більше дівчаток. Серед новонароджених з їх нестачею більше хлопчиків. Подібна картина спостерігається і в розподілі вроджених вад серця і магістральних судин.
До другої групи відносяться ознаки, що зустрічаються тільки в однієї статі. Це первинні та вторинні статеві ознаки: статеві органи, молочні залози, борода у людини, грива у лева, а також багато господарських ознак (продукція молока, яєць, ікри тощо). Статевий диморфізм щодо них носить генотипічний характер, оскільки у фенотипі однієї статі ці ознаки відсутні, але спадкова інформація про ці ознаки записана в генотипі обох статей. Тому, якщо вони еволюціонують, то за ними має існувати генотипний статевий диморфізм. Виявляється він у вигляді реципрокних ефектів.
Третя група ознак знаходиться посередині між першою (статевий диморфізм відсутня) та другою групою (статевий диморфізм носить абсолютний характер). До неї відносяться ознаки, які зустрічаються і в чоловічої статі, і в жіночої, але розподілені в популяції з різною частотою та ступенем виразності. Це кількісні ознаки: зростання, вага, розміри та пропорції, багато морфофізіологічних та етолого-психологічних ознак. Статевий диморфізм щодо них проявляється як відношення їх середніх значень. Він справедливий для всієї популяції, але може мати зворотне значення окремої пари особин. Саме цей статевий диморфізм є «компасом» еволюції ознаки.
Статевий диморфізм тісно пов'язаний з еволюцією ознаки: він повинен бути відсутнім або бути мінімальним для стабільних ознак і максимальною, найбільш чітко вираженою, за філогенетично молодою (еволюціонуючою) ознакою. Як і дві інші основні характеристики роздільностатевої популяції - дисперсія та співвідношення статей, статевий диморфізм розглядається не як константа, властива даному виду, як вважалося раніше, а як змінна та регульована величина, тісно пов'язана з умовами середовища та визначальна, у свою чергу, еволюційну пластичність ознаки. Оскільки в мінливому, екстремальному середовищі потрібна більша пластичність, ніж у стабільному (оптимальному), то й статевий диморфізм у стабільному середовищі повинен зменшуватися, а в мінливому - рости.
Статевий диморфізм має бути пов'язаний з репродуктивною структурою популяції: у строгих моногамів він має бути мінімальним, оскільки моногами використовують спеціалізацію статей лише на рівні організму; у полігамних видів, повніше використовують переваги диференціації, він має зростати зі зростанням ступеня полігамії.
За ознаками, властивими лише одній статі (первинні та вторинні статеві ознаки, а також багато господарсько-цінних ознак: продукція яєць, молока, ікри), статевий диморфізм має абсолютний, організмовий, характер. Оскільки у фенотипі однієї статі ці ознаки відсутні, про генотипічний статевий диморфізм за ними можна судити з реципрокних ефектів. Якщо за «старими» (стабільними) ознаками генетичний внесок батька в нащадка в середньому дещо менший за вклад матері (через материнський ефект, зумовлений цитоплазматичною спадковістю, гомогаметною конституцією та утробним розвитком у ссавців), то за «новими» ознаками, згідно з еволюційною теорією статі, має існувати деяке домінування батьківських ознак над материнськими.
Батьківський ефект встановлений по алкоголізму у людини, по інстинкту насиджування, скоростиглості, несучості та живої ваги у курей, по динаміці зростання, числу хребців і довжині тонкого кишечника у свиней, по вудою молока та продукції молочного жиру у великої рогатої худоби. Наявність батьківського ефекту по удою і несучості означає не що інше, як більш високу генотипічну «подійність» у бугаїв і «несучість» у півнів, ніж у корів і курей тих же порід.
Так як жіноча стать спеціалізується більше за генетичним потоком інформації та зв'язків всередині популяції, у них повинні бути краще розвинені мова та вербальні здібності. Етологічні особливості жіночої статі спрямовані на збереження старої, вже освоєної та вдосконалення вже знайдених рішень. Самки більше прагнуть пристосуватися до середовища, вижити та залишити потомство. Тому вони більш податливі, більше схильні до впливу середовища проживання і ефективніше навчаються.
З іншого боку, екологічна спеціалізація чоловічої статі може пояснити їх краще розвинені просторово-зорові здібності, пов'язані з середовищем (захист, полювання, боротьба з ворогами). Поведінкові особливості чоловічої статі спрямовані зміну старого і мають характер пошуку нових рішень. Вони охоче виявляють більш ризиковану, “дослідницьку” поведінку, гірше навчаються і менш конформні.
На основі еволюційного підходу було проведено аналіз психологічних відмінностей між статями у вербальних та фізичних здібностях, імпульсивності та пошуку відчуттів, а також у процесі навчання, психології творчості, відмінності у статусних уподобаннях та прагненні до влади та контролю. . Трофімова запропонувала додавання до теорії Геодакяна у формі поняття "пропалювання непотрібностей" (redundancy prunning). Це поняття визначає тенденцію чоловічої частини статі скорочувати непотрібні ступені свободи порушенням прийнятих правил та умовностей.
За твердженням Геодакяна, уявлення теорії статі про відособленість нової та старої інформації протягом багатьох поколінь дозволяє пояснити ряд незрозумілих явищ в антропології. Так у туркменській популяції методом узагальненого портрета була виявлена чітка різниця по напів - жіночі портрети укладалися в один тип, а чоловічі - у два типи. Аналогічне явище спостерігав Р. М. Юсупов у краніології башкир – жіночі черепи були близькі до угро-фінського типу (у географічному плані це північно-західні сусіди сучасних башкир), а чоловічі – до алтайського, казахського та інших (східні та південно-східні сусіди) ). У удмуртської популяції, дерматогліфіка у жінок відповідала північно-західному типу, а у чоловіків - східно-сибірському. Л. Г. Кавгазова відзначила подібність дерматогліфіки болгар з турками, тоді як болгарки були ближчими до литовців. Жіночі форми фенотипів показують вихідний етнос, тоді як чоловічі форми - кількість джерел та напрямок генних потоків. Факти наведені вище, показують угро-фінське походження удмуртських та башкирських етносів, що різняться за культурою та мовою. Чотирьохмодальний розподіл черепів чоловічої частини популяції, за твердженням В. Геодакяна, пояснюється впливом трьох різних навал з півдня та сходу. Напрямок генних потоків у цих популяціях - з південного сходу північний захід, а населенню болгар - з півдня північ. Він стверджує також, що острівна популяція (японська), у повній відповідності до теорії, є мономодальною для обох статей.
В оптимальних, стабільних умовах середовища, коли немає необхідності у високій еволюційній пластичності, основні характеристики зменшуються і мають мінімальне значення, тобто падає народжуваність (одночасно і смертність) хлопчиків, скорочується їхня різноманітність і різниця між чоловічою та жіночою статтю. Усе це знижує еволюційну пластичність популяції. В екстремальних умовах мінливого середовища, коли для швидкої адаптації потрібна висока еволюційна пластичність, йдуть зворотні процеси: ростуть, одночасно, народжуваність і смертність (тобто «оборотність») чоловічої статі, його різноманітність, чіткіше стає статевий диморфізм. Усе це підвищує еволюційну пластичність населення.
Ознака еволюціонує - якщо у ньому існує статевої диморфізм, і стабільний - коли статевої диморфізм відсутня.
«Якщо за якоюсь ознакою існує генотипний популяційний статевий диморфізм, то ця ознака еволюціонує від жіночої форми до чоловічої».
Правило є частиною "Еволюційної теорії статі". З погляду системного підходу, застосованого В. А. Геодакяном в 1965 до проблеми статі, статевий диморфізм розглядається як наслідок асинхронної еволюції статей. Отже, статевий диморфізм виникає тільки за ознаками, що еволюціонують. Це еволюційна «дистанція» між статями, що виникає з початком еволюції ознаки і зникає з її кінцем. Відповідно, статевий диморфізм може бути наслідком будь-якого виду відбору, а не лише статевого, як вважав Дарвін.
Якщо дисперсія ознаки у чоловічої статі більша, ніж у жіночої - еволюція знаходиться в дивергентної фазиякщо дисперсії статей рівні - фаза еволюції стаціонарнаякщо дисперсія більше у жіночої статі - то фаза конвергентна. Дисперсія - різноманітність ознак у чоловічої та жіночої статі.
За аналогією із співвідношенням статей для різних стадій онтогенезу можна виділити також первинну, вторинну і третинну дисперсію статей. Оскільки дисперсія пов'язана з ознаками, а в зиготі більшість ознак ще в потенції, під первинною дисперсією треба розуміти ті потенції, з яких буде реалізовано ознаку дорослої дефінітивної стадії.
Було виявлено, що жіночі нащадки успадковують батьківські ознаки адитивніше (проміжне, середньоарифметичне успадкування), ніж чоловічі. Відмінності між самцями та самками мишей спостерігалися для відносної ваги надниркових залоз, тимусу, статевих залоз та гіпофіза, а також генів, відповідальних за рухову активність.
Велика фенотипічна дисперсія чоловічої статі є одним з основних положень еволюційної теорії статі. Оскільки фенотипова дисперсія відбиває генотипическую, очікується, що з чоловічих особин вона має бути ширше з допомогою мутацій і неаддитивного успадкування ознак. Ступінь зв'язку генотипу з фенотипом (норма реакції) також визначає величину дисперсії фенотипу.
Можна сказати також, що для початкової, ювенільної стадії онтогенезу характерніша жіноча форма ознаки, а для дефінітивної, зрілої стадії - чоловіча. Інакше кажучи, жіночі форми ознак із віком повинні, зазвичай, слабшати, а чоловічі форми - посилюватися.
На тісніший зв'язок жіночої статі з початковою фазою онтогенезу звернув увагу ще Дарвін. Він писав: «У всьому тваринному царстві, якщо чоловіча і жіноча стать відрізняються один від одного на вигляд, видозмінюється, за рідкісними винятками, самець, а не самка, тому що остання зазвичай залишається схожою з молодими тваринами свого вигляду і з іншими членами всієї групи». Антропологи також відзначали близькість жіночого типу з дитячим (більш грацильні кістки, слабовиражені надбрівні дуги, менше оволосіння тіла та ін.).
Яскравим прикладом може бути зв'язок ступеня розвитку рогів, в різних видів оленів і антилоп, з віком їх появи у самців і самок: чим сильніше виражена рогатість у виду в цілому, тим більш ранньому віці з'являються роги: спершу у самців і пізніше у самок. Онтогенетичне правило статевого диморфізму було підтверджено на 16 антропометричних ознаках: відносна довжина ніг, передпліччя, 4-го і 2-го пальців, головний індекс, коло зубної дуги, епікантус, горбинка спинки носа, оволосіння тіла, обличчя та голови, концентр , частота пульсу , швидкість випорожнення жовчного міхура , асиметрія мозку , час реакції , відчуття гіркого смаку фенілтіо - сечовини та нюх .
«У реципрокних гібридів за ознаками батьків, що дивергують, повинна домінувати батьківська форма (порода), а за конвергувальними - материнська.»
«Аномалії розвитку, що мають „атавістичну“ природу частіше повинні з'являтися у жіночої статі, а мають „футуристичну“ природу (пошук), - у чоловічої». За видовими (і вище рангів спільності) ознаками (багатоклітинність, теплокровність, число органів, план і принципова будова тіла та ін) статевий диморфізм в нормі відсутня. Він спостерігається тільки в області патології і виявляється у різній частоті появи тих чи інших вроджених вад розвитку у чоловічої та жіночої статі. Класифікація (далеких попередників, - серед них більше дівчаток. Серед новонароджених з їх нестачею - навпаки, більше хлопчиків.).
Правило було також перевірено на матеріалі вроджених вад серця та магістральних судин (32 тис. випадків). Було показано, що жіночі аномалії розвитку носять характер збереження ембріональних особливостей будови серця, властивих останнім стадіям внутрішньоутробного розвитку, або ознак, властивих видам, що стоять на нижчих щаблях еволюційних сходів (недалеке минуле) (відкритий овальний отвір у міжпредсердний) . Елементи «чоловічих» вад (стенози, коарктації, транспозиція магістральних судин) мають «футуристичну» природу (пошук).
Мюллера, що встановлює зв'язок між явищами філогенезу та онтогенезу (онтогенез – коротке повторення філогенезу).
Якщо для простоти говорити не про організм у цілому, а лише про одну його ознаку, то явище філогенезу - це динаміка (поява та зміна) ознаки в еволюційному масштабі часу, в історії виду. Явище онтогенезу - це динаміка ознаки історії життя особини. Отже, закон Геккеля-Мюллера пов'язує між собою онтогенетичну та філогенетичну динаміки ознаки.
У 1965 р. В. А. Геодакяном було відкрито закономірність, що пов'язує явище популяційного статевого диморфізму з філогенезом. «Якщо за якоюсь ознакою існує генотипний популяційний статевий диморфізм, цей ознака еволюціонує від жіночої форми до чоловічої».
У 1983 р. їм була теоретично передбачена закономірність, що пов'язує явище статевого диморфізму з онтогенезом . "Якщо за якоюсь ознакою існує популяційний статевий диморфізм, то в онтогенезі ця ознака змінюється, як правило, від жіночої форми до чоловічої".
Введемо поняття двох форм ознаки, пов'язаних з вектором часу, у кожному з трьох явищ (філогенезу, онтогенезу та статевого диморфізму). У філогенезі ознаки розрізнятимемо його «атавістичну» і «футуристичну» форми, в онтогенезі ознаки – його «ювенільну» (молоду) та «дефінітивну» (дорослу) форми, а в популяційному статевому диморфізмі – його «жіночу» та «чоловічу» форми . Тоді узагальнену закономірність, що пов'язує явища філогенезу, онтогенезу та статевого диморфізму, можна сформулювати як «правило відповідності» між атавістичної, ювенільноюі жіночоїформами ознак, з одного боку, і між футуристичної, дефінітивнийі чоловічийформами – з іншого.
«Правило відповідності» можна поширити і інші явища, системно пов'язані з філогенезом і онтогенезом (еволюцією), у яких можна виділити минулу і майбутню форми. Наприклад, явище мутування (філогенетичний процес виникнення генів), явище домінування (онтогенетичний процес прояву генів), явище гетерозису та реципрокних ефектів. На зв'язок між явищами філогенезу, онтогенезу, мутування, домінування та статевого диморфізму вказують такі відомі факти, як: вищий рівень спонтанних мутацій у чоловічої статі; більш адитивне успадкування батьківських ознак нащадками жіночої статі, отже, більш домінантне успадкування нащадками чоловічої статі; дають можливість знаючи одне явище передбачити два інших. Відомо, що у далеких філогенетичних попередників людини очі були розташовані латерально, їх зорові поля не перекривалися і кожне око було пов'язане лише з протилежною півкулею мозку – контралатерально. У процесі еволюції в деяких хребетних, зокрема й у предків людини, у зв'язку з придбанням стереоскопічного зору, очі перейшли вперед. Це призвело до перекриття лівого та правого зорових полів та до появи нових іпсилатеральних зв'язків: ліве око – ліва півкуля, праве око – праве. Таким чином, з'явилася можливість мати в одному місці зорову інформацію від лівого та правого ока - для їх зіставлення та вимірювання глибини. Отже, іпсилатеральні зв'язки філогенетично молодші, ніж контралатеральні. На підставі філогенетичного правила, можна передбачити еволюційно більш просунуті іпсі-зв'язки у чоловічої статі, порівняно з жіночим, - тобто статевий диморфізм за часткою іпсі/контра волокон у зоровому нерві. З онтогенетичного правила можна передбачити зростання частки ипси волокон в онтогенезі. А оскільки візуально-просторові здібності та об'ємна уява тісно пов'язані зі стереоскопією та іпсі зв'язками, то стає зрозумілим, чому у чоловіків вони розвинені краще. Цим пояснюються спостерігаються відмінності чоловіків і жінок у розумінні геометричних завдань, орієнтуванні та визначенні напрямів, читанні креслень та географічних карт (див., напр., // Behavioral Neuroscience).
Застосування тих же правил до нюхового рецептора людини призводить до висновку, що у філогенезі нюх людини, на відміну від зору, погіршується. Оскільки, як було показано, у людей з віком відбувається атрофія нюхових волокон та їх кількість в нюховому нерві неухильно зменшується, то можна передбачити, що їхня кількість у жінок має бути більшою, ніж у чоловіків.
Передбачення було підтверджено аналізом 31814 хворих із вродженими вадами серця та магістральних судин. Наднормативні м'язи в 1,5 рази частіше виявляються у чоловіків, ніж у жінок.
Критика теорії статі загалом у літературі відсутня. Критика окремих аспектів іноді трапляється. Наприклад у книзі Л. А. Гаврилова та Н. С. Гаврилової аналізуються статеві відмінності в тривалості життя. Щодо більшої варіабельності ознак у самців відповідальної їх підвищену смертність, автори відзначають, що «у цій гіпотезі не розкритий конкретний молекулярно-генетичний механізм, що призводить до більшої тривалості життя самок». І там-таки пишуть, що цей недолік, «може бути в принципі усунений у ході подальшого розвитку та конкретизації цієї гіпотези». Вони вважають, що прогноз теорії про переважання чоловіків серед довгожителів не узгоджується з фактами, оскільки, по-перше, «у міру зростання тривалості життя зростають і відмінності за цією ознакою між чоловіками і жінками» і, по-друге, «в останні роки в розвинених країн спостерігається прискорене проти чоловіками зниження смертності жінок старшого віку» . Вони також вважають, що "велика тривалість життя самок зовсім не є загальнобіологічною закономірністю". Слід зазначити, що висновок про більшу тривалість життя самок у більшості вивчених видів був зроблений задовго до появи теорії статі в низці робіт.
Положення теорії про співвідношення статей та "Феномен воєнних років" обговорювалися в роботі В. Іскріна
Оскільки сам Ч. Дарвін вважав, що чоловіча стать змінюється раніше, основне положення концепції В. Геодакяна, що еволюція статей відбувається асинхронно, не суперечить теорії еволюції Дарвіна. Останнім часом у країнах навіть широко вживається новий термін “male-driven evolution”. Теорія В. Геодакяна доповнює та розширює теорію статевого відбору Ч. Дарвіна зазначаючи, що статевий диморфізм може виникнути внаслідок будь-якого (а не лише статевого) відбору. А. С. Кондрашов у класифікації теорій статі помістив її в категорію "Гіпотез негайної переваги" (immediate benefit) оскільки відбір серед "дешевих" самців і чоловічих гамет ефективніший.
Теорія В. Геодакяна аналізує процес статевої диференціації, і тому не суперечить численним теоріям, які намагаються пояснити виникнення та підтримання статевого розмноження, оскільки вони приділяють основну увагу процесу схрещування.
Серед теорій роздільної порожнини, теорія статі є більш загальною, ніж, наприклад, теорія Паркера (1972), яка пояснює статеву диференціацію на рівні гамет і тільки у водних тварин
З чим можуть бути пов'язані індивідуальні відмінності між представниками чоловічої та жіночої статі? Очевидно, що для відповіді на це питання необхідно вийти за межі психології та звернутися до теорій та гіпотез, що існують в етології та біології.
Питання, для чого взагалі існує стать, виникало давно. Найпростіша відповідь – для розмноження – задовільною вважатися не може. У живому світі існує крім раздельнополого ще також безстатеве (вегетативне) і гермафродитне розмноження, причому очевидних переваг перед ними у раздельнополого розмноження немає. Навпаки, комбінаторний потенціал (поєднання генів) у гермафродитів вдвічі більше, а кількість потомства (ефективність розмноження) вище у безстатевих. Однак усі прогресивні форми розмножуються саме статевим шляхом (3, 5).
Для прояснення ролі раздельнополого розмноження 1965 р. вітчизняним біологом В.А.Геодакяном (під очевидним впливом кібернетики та теорії систем) було створено так звана еволюційна теорія статі. У якій автор стверджував, що диференціація статей пов'язана зі спеціалізацією з двох основних аспектів еволюційного процесу збереження та зміни генетичної інформації як вигідної для популяції форми інформаційного контакту з середовищем.Очевидно, що тільки чоловічих (або тільки жіночих) особин недостатньо для забезпечення наступності та розвитку виду. Вони мають співіснувати.
Поклавши в основу своєї теорії принцип сполучених підсистем, Геодакян зазначив, що адаптивні системи, що еволюціонують у рушійному середовищі, значно підвищують свою загальну стійкість за умови диференціації на дві сполучені підсистеми, з консервативною та оперативною спеціалізацією, що належать особам відповідно жіночого та чоловічого. Як це відбувається?
Спочатку організм жіночих особин має ширшу норму реакції, ніж чоловічий. Так, якщо чоловік у конфліктній поведінці, наприклад, зазвичай поводиться вибуховим чином, то зробити його терпимим і миролюбним навряд чи вдасться. А жінка може поєднувати у поведінці кілька стратегій, гнучко використовуючи в залежності від ситуації. Завдяки цьому адаптивні здібності жіночих особин набагато вищі, а учня краще. (У дослідженнях з педагогічної психології наголошується, що початковий рівень здібностей, як правило, вищий у хлопчиків, але в процесі навчання вони швидше виходять на плато, тоді як дівчатка, відштовхуючись від нижчих показників, набирають темп і обганяють хлопчиків.) Якщо ми прийдемо в шкільний клас і подивимося на успішність дітей, то виявиться, що дівчатка (як і хлопчики) однаково розподіляються на відмінниць, двієчниць та посередніх учениць. Однак якщо ми поставимо питання інакше: хто найзапекліший двієчник і хуліган, хто найталановитіший учень? - виявиться, що ці групи заповнені, як правило, хлопчиками. Тобто чоловіча підвиборка має більш спеціалізовану поведінку, що в цілому заважає адаптації на рівні індивіда. Усі крайнощі яскравіше представлені у чоловіків, але жінки більш навчальні.
Припустимо, що середовище існування виду практично не змінюється (таке середовище називають стабілізуючим). У цьому середовищі природний відбір веде до простого збільшення чисельності особин, без зміни їхнього генотипу. Для цієї мети немає необхідності присутності великої кількості чоловічих особин у популяції, головне, щоб було чимало жіночих особин. І справді, у стабільних умовах хлопчиків народжується трохи менше (існує навіть прикмета, що багато хлопчиків народжується до війни).
Але якщо середовище різко змінює свої умови (стає рушійним), то завдання відбору у пристосуванні дещо змінюються; він призводить не лише до збільшення кількості особин, а й до зміни генотипу. В умовах катастроф (екологічних, соціальних, історичних) елімінація та усунення від розмноження в основному зачіпають чоловічу стать, а модифікація – жіноча. Завдяки диференціації статей з'явилися дві основні зміни порівняно з безстатевим розмноженням – це ширший переріз інформаційного каналу взаємодії у чоловічої особини та ширша норма реакції у жіночої особини. Таким чином, чоловіча особина може запліднити більшу кількість самок, а жіноча забезпечити спектр фенотипів з одного генотипу.
Після зникнення катастрофічного чинника та закінчення дії відбору частка чоловічих особин зменшується, і їхня генотипна дисперсія звужується (ті, хто не вижив, не залишають генетичних слідів). Отже, жінки забезпечують постійну філогенетичну пам'ять виду, а чоловіки – тимчасову, онтогенетичну (3).
Для ілюстрації цієї думки Геодакян наводить такий поетичний приклад. Коли настало загальне похолодання на планеті, то у жінок, як високоадаптованих істот, збільшився жировий прошарок. А чоловіки через слабку пристосованість виявилися до цього нездатними і здебільшого просто вимерли. Зате решта винайшов вогонь, щоб зігрівати всю громаду, і з цього моменту став закріплюватися саме його генотип. Отже, чоловіки здійснюють пошук, а жінки – удосконалення. Такий механізм еволюційного біологічного (та психологічного) прогресу.
Очевидно, що, маючи вузьку норму реакції, чоловіки більш біологічно (і психологічно) вразливі. Тому і тривалість життя у них нижча. Новонароджені хлопчики частіше гинуть, аніж дівчатка. Проте більшість довгожителів таки чоловіки.
Звичайно, розвиваються та змінюються не всі анатомо-фізіологічні та поведінкові ознаки, а лише деякі. Наявність відмінностей ознак у чоловічих та жіночих особин називається статевим диморфізмом, тобто. існуванням двох форм (а психології вже почали використовувати і вираз статевої дипсихізм). У сучасних людей, наприклад, існує статевий диморфізм за ознаками зростання, ваги, оволосіння, але немає диморфізму за ознакою кількості пальців чи вух, за кольором очей.
У стабілізуючому середовищі статевий диморфізм відсутня (немає необхідності пристосовуватися, і чоловічі та жіночі особини мають одне і те ж еволюційно вигідне значення ознаки). А в рушійному середовищі вже в одному поколінні з'являється генотипний статевий диморфізм, що зростає у наступних поколіннях. По варіативності ознаки можна будувати висновки про фазі еволюційного процесу за ознакою. Так, якщо в чоловічій підвиборці дисперсія вища, ніж у жіночій, це свідчить про початок еволюційного процесу, а фазу відбору називають дивергентною. Потім настає паралельна фаза, коли дисперсії в обох групах приблизно рівні. І, нарешті, конвергентна фаза, де варіативність в жінок зростає проти чоловіками, свідчить у тому, що еволюційний процес близький до завершення.
Геодакяном сформульовано філогенетичне правило статевого диморфізму: якщо за якоюсь ознакою існує популяційний статевий диморфізм, то ця ознака еволюціонує від жіночої форми до чоловічої. Тобто населення маскулінізується, а значення ознаки, що існують у чоловічій підвиборці, є еволюційно вигідними. Це стосується всіх видів, що мають роздільностатеве розмноження. Так, наприклад, якщо у ссавців самка за розміром менше, ніж самець, це означає, що по ходу еволюційного процесу самки збільшуватимуться у розмірі, тому що це вигідно для виду. А у комах (наприклад, у павуків) самки, навпаки, значно більше ніж самці; це говорить про те, що легкій істоті у його середовищі вижити легше. Отже, і самки ставатимуть менше.
У селекції цей факт також використовується: оскільки селекційні ознаки більше просунуті у батьків, то відбір виробника є ключовою проблемою для виведення нових порід, навіть якщо це стосується прихованих ознак, наприклад подійності.
Існує також онтогенетичне правило статевого диморфізму: якщо за якоюсь ознакою існує популяційний статевий диморфізм, то в онтогенезі ця ознака змінюється, як правило, від жіночої форми до чоловічої. Правило батьківського ефекту в селекції полягає в тому, що за ознаками батьків, що дивергують, (що є предметом уваги) повинна домінувати батьківська форма (порода), а за конвергуючою (несуттєвою для виведення породи) - жіноча.
Цікаво, що в онтогенезі жіночі форми ознаки виявляються раніше, а чоловічі пізніше. Так, маленькі діти обох статей більше схожі на дівчаток, а у людей похилого віку, знову ж таки, незалежно від статі, починають проявлятися чоловічі риси (грубий голос, ріст волосся на обличчі тощо). За характерологічними ознаками маленької дівчинки можна з більшою достовірністю передбачити структуру особистості та поведінку дорослої жінки, ніж у хлопчиків. Тому можна говорити не тільки про диморфізм, а й про дихрономорфізм (тобто тимчасове розбіжність прояви жіночих і чоловічих ознак) (3, 6).
Примітно, що вроджені аномалії, що мають «атавістичну» природу, найчастіше виявляються у жінок, а «футуристичну» у чоловіків. Так, серед новонароджених дівчаток частіше трапляються хвостиками. Однак найдовший хвостик, що становить 13 см, все ж таки належав хлопчику. Спостерігається статевий диморфізм і в області появи хвороб (усі нові захворювання, такі, як рак, СНІД, спочатку з'являлися у чоловіків), і в будові мозку (у чоловіків виразніше виражена асиметрія півкуль і оперативні системи - кора і ліва півкуля, а у жінок - Консервативні системи - підкорка і права півкуля, що обумовлює переважання у чоловіків аналітичного мислення, а у жінок - інтуїтивного, образного і чуттєвого пізнання). Завдяки меншій асиметрії жінки також більш навчаються. Крім того, і в культурно-історичному процесі спостерігається флагманська роль чоловіків: кожна нова професія була спочатку тільки чоловічою і лише потім ставала жіночою, а основні наукові відкриття та культурні революції також відбувалися чоловіками.
"Основне питання - навіщо стать?"
Белл (1982)
Теорію В. Геодакяна можна звести до однієї фрази:
Чоловіки-піддослідні кролики Природи.
Підлога - це не стільки спосіб розмноження, як прийнято вважати,
скільки метод асинхронної еволюції.
В. Геодакян (1991)
Для розуміння явища статі недостатньо знань його репродуктивної та рекомбінативної ролі. Потрібно знати його еволюційну роль. Раздельнополость включає два фундаментальні явища: схрещування(об'єднання генетичної інформації батьків ) та диференціацію(поділ на дві статі). Наявність схрещування відрізняє статеві форми розмноження від безстатевих, наявність диференціації-роздільні форми від гермафродитних. Класична генетика розглядає, однак, лише результати схрещування особин, тому деякі явища, пов'язані з самою диференціацією, не можуть бути пояснені.

Нова теорія розглядає диференціацію статей як вигідну для популяції форму інформаційного контакту з середовищем, як еволюційну спеціалізацію з двох основних альтернативних аспектів еволюції: збереження(консервативний) та зміни(Оперативний).
Які якості «наближають» чоловічу стать до середовища та забезпечують йому отримання екологічної інформації? У чоловічої статі в порівнянні з жіночою вище частота мутацій, менша адитивність успадкування батьківських ознак, вже норма реакції, вища агресивність і допитливість, активніша пошукова, ризикована поведінка. Інша група особливостей — величезна надмірність чоловічих гамет, їхні малі розміри та висока рухливість, велика активність та мобільність самців, та їх схильність до полігамії. Тривалі періоди вагітності, годування і турботи про потомство у самок, практично підвищуючи ефективну концентрацію чоловічих особин, перетворюють чоловічу стать на «надлишковий», отже, «дешевий», а жіночий — на дефіцитний і більш цінний.
Це призводить до того, що відбір діє в основному за рахунок усунення чоловічих особин, але великі потенційні можливості дозволяють їм запліднити усі жіночі (у панміктній чи полігамній популяції). В результаті невелика кількість чоловічих особин передає потомству стільки ж інформації, скільки і велика кількість жіночих. Можна сказати, що канал зв'язку з потомством у чоловічої статі ширший, ніж у жіночої. Спадкова інформація, отримана потомством від матерів, краще відображає розподіл генотипів у популяції і в минулих поколіннях. Інформація, отримана від батьків, більш селективна, вона відображає генотипи найбільш пристосовані до умов середовища.
Широка норма реакції жіночих особин забезпечує їм більш високу онтогенетичну пластичність (адаптивність), дозволяє залишити зони елімінації та дискомфорту та групуватися навколо норми популяції, тобто у стабільному середовищі зменшити свою фенотипічну дисперсію. Вузька норма реакції чоловічої статі зберігає їх широку фенотипічну дисперсію і робить їх більш схильними до відбору. Це означає, що чоловіча стать першим піддається еволюційним змінам.
У безстатевих та гермафродитних популяціях інформація від середовища потрапляє до всіх осіб:
У разі статевої диференціації послідовність появи контролюючої інформації від середовища така:
середа → чоловіча стать → жіноча стать
Отже, чоловіча стать можна розглядати як еволюційний "авангард" популяції, а статевий диморфізм за ознакою як еволюційну "дистанцію" між статями і як "компас", що показує напрямок еволюції цієї ознаки (“Філогенетичне правило статевого диморфізму”).Тому ознаки найчастіше й найбільш виражені у самок повинні мати “атавістичну” природу, тоді як ознаки більш виражені у самців повинні мати “футуристичну” природу (пошук). Максимальний статевий диморфізм має спостерігатися за еволюційно молодими (новими) ознаками.
У реципрокних гібридів дивергуючих форм за еволюціонуючими (новими) ознаками має спостерігатися реципрокний "батьківський ефект"(Домінування батьківської породи, лінії). За ознаками батьків, що дивергують, повинна домінувати батьківська форма, за конвергувальними—материнська. Зокрема, теорія успішно передбачає існування батьківського ефекту за всіма господарсько-цінними ознаками у сільськогосподарських тварин та рослин.
Новий погляд на еволюційну роль статі дозволяє краще зрозуміти багато явищ, пов'язаних із статтю: статевий диморфізм (ПД), співвідношення статей (СП), роль статевих хромосом (ПХ) та статевих гормонів (ПГ), психологічні відмінності між чоловіками та жінками та ін.
Замість існуючого раніше погляду на основні характеристики роздільностатевої популяції як на константи, специфічні для виду, пропонується новий: співвідношення статей, дисперсія і статевий диморфізм - змінні, регульовані величини, тісно пов'язані з умовами середовища. У стабільних умовах (оптимальне середовище) вони мають падати, а в мінливих умовах (екстремальне середовище) — рости. У першому випадку еволюційна пластичність виду зменшується, тоді як у другому—збільшується.
Статевий диморфізм це “дистанція” між статями у процесі еволюції будь-якої ознаки. Це генетична інформація, яка завдяки спеціалізації статей на популяційному рівні вже потрапила до чоловічої підсистеми, але ще не досягла жіночої.
Інформаційний контакт організму з середовищем визначається та регулюється співвідношенням у ньому чоловічих (андрогенів) та жіночих (естрогенів) гормонів. Андрогени "наближають" (в інформаційному сенсі) організм до середовища, а естрогени, навпаки, "видаляють" його від середовища.
Розкриті філогенетичні та онтогенетичні закономірності диференціації статей сформульовані у вигляді правил.
Досі вважалося, що розподіл на дві статі необхідний для самовідтворення, що підлога є способом розмноження. А виявляється, що підлога це швидше спосіб еволюції .
Теорія дозволяє з єдиних позицій відповісти на багато питань, на які не може відповісти теорія статевого відбору Дарвіна і передбачити нові явища.
МедицинаЗ'являється можливість пояснити статевий диморфізм щодо багатьох хворобам. Співвідношення статей захворюваності залежить від фази взаємодії популяції зі шкідливим чинником середовища.
Детальніше...
Детальніше про теорію статі
Перша публікація:Хлопчик або дівчинка. Співвідношення статей-величина, регульована природою? (В. А. Геодакян). Наука та Життя, 1965 № 1, с.55-58.
Популярний виклад: Еволюційна теорія статі . (А. Гордон). Програма "00:30" НТВ, 06.03.2002
Більш повний науковий виклад:
Найсвіжіша публікація: Чоловік і жінка. Еволюційно-біологічне призначення . Геодакян Ст А. Між. Конф.: Жінка та свобода. Шляхи вибору у світі традицій та змін.Москва, 1-4 червня 1994, с. 8–17.
Copyright © 2005 -2012 S. Geodakyan. Всі права захищені.
Чому існують чоловіки та жінки? Чому б нам не бути, як амебам, безстатевими? Чи, як земляним черв'якам, гермафродитами? Чому статей існує два, не три? Вченим важко відповісти на це питання. Важко усі - крім одного. Доктор біологічних наук, провідний науковий співробітник Інституту загальних проблем екології та еволюції РАН Віген Артаваздович Геодакян вже сорок років розробляє теорію, покликану пояснити в чому сенс поділу живих істот на дві статі. І всі, кому доводилося познайомитися з цією теорією, розкривали рота від подиву.
Піддослідні кролики
Всю теорію Геодакяна можна висловити лише у фразі - чоловіки є піддослідними кроликами природи. Як відомо, життя на землі розвивається від амеби до динозаврів, і від динозаврів - до людини, і відбувається це тому, що живі істоти набувають у процесі еволюції нових ознак і властивостей. Але зміни в живих організмах можуть бути не лише корисними, а й шкідливими. Інша необережна мутація може привести до вимирання біологічного виду. Ось для цього більшість біологічних істот поділяться на чоловіків і жінок, і весь ризик, пов'язаний із зміною, беруть на себе чоловіки, яких можна сміливо назвати першопрохідниками еволюції. Адже якщо мутація виявиться шкідливою, і більшість чоловіків загине - на долі виду це не позначиться, не численні чоловіки, що залишилися в живих, зможу, використовуючи численних жінок, легко відновити свою чисельність. Таким чином, чоловіки якийсь час пробують нові мутації на собі, і якщо виживають - передають нову ознаку жінкам.
Відповідно до поки що не відкинутої теорією Дарвіна, еволюція живих істот відбувається у результаті природного відбору. Припустимо, на землі настало глобальне похолодання, і всім тваринам добре б обзавестися густим хутром. Відбір діє грубо - всі, у кого хутра не вистачає, гинуть, залишаються найшерстистіші, які і дають початок новому поколінню морозостійких тварин. Природа надходить як селекціонер: вибирає овець із найгустішою вовною і робить їх засновником нових, густошерстих порід. Лихо в тому, що чим «ретельніше» відбувається селекція, тим менше особин залишається в живих. Якщо після перших морозів овець залишиться замало, то, нехай навіть будуть дуже вовняні, але вони вже просто не зможуть відновити чисельність свого вигляду. Але якщо в живих залишиться занадто багато, то нові ознаки - («густіша вовна») не виявляться досить чітко. Тобто будь-який біологічний вид перебуває між двома небезпеками - потрібно зберігати свою чисельність і забезпечувати високу швидкість еволюції.
І ось тут на допомогу вівцям, як і всім іншим живим істотам, приходить поділ на дві статі. Жінки відповідають за підтримку чисельності свого виду. Чоловіки постійно гинуть, але передають потомству нові ознаки, які вийшли в результаті найжорсткішої селекції.

Чоловічі та жіночі таланти
Зрозуміло виникає питання - а як жінкам вдається виживати в умовах, що змінюються, скажімо серед тих же холодів? Але в кожної статі свій талант. Чоловіки мають талант змінюватися генетично, набувати нових мутацій, які передають своїм дітям. Жінки набагато легше самі пристосовуються до нових умов, та їх гени у своїй залишаються незмінними. Жінки легше виживають і в холоді, і в спеку, але саме тому на них не діє природний відбір, і їм нема чого передати потомству. Як каже Віген Артаваздович, якщо чоловіка з дитинства погано годувати – він помре, якщо жінку погано годувати, вона виросте менше на зріст. Коли настають холоди, вівця, на відміну барана, легко сама обростає вовною, але це ознака не передається потомству. Саме тому для підтримки молочної породи худоби молочні бики - як це не смішно звучить - відіграють набагато більшу роль, ніж молочні корови.
Поклавши в основу своєї теорії принцип сполучених підсистем, Геодакян зазначив, що адаптивні системи, що еволюціонують у рушійному середовищі, значно підвищують свою загальну стійкість за умови диференціації на дві сполучені підсистеми, з консервативною та оперативною спеціалізацією, що належать особам відповідно жіночого та чоловічого. Як це відбувається?
Спочатку організм жіночих особин має ширшу норму реакції, ніж чоловічий. Так, якщо чоловік у конфліктній поведінці, наприклад, зазвичай поводиться вибуховим чином, то зробити його терпимим і миролюбним навряд чи вдасться. А жінка може поєднувати у поведінці кілька стратегій, гнучко використовуючи в залежності від ситуації. Завдяки цьому адаптивні здібності жіночих особин набагато вищі, а учня краще. (У дослідженнях з педагогічної психології наголошується, що початковий рівень здібностей, як правило, вищий у хлопчиків, але в процесі навчання вони швидше виходять на плато, тоді як дівчатка, відштовхуючись від нижчих показників, набирають темп і обганяють хлопчиків.) Якщо ми прийдемо в шкільний клас і подивимося на успішність дітей, то виявиться, що дівчатка (як і хлопчики) однаково розподіляються на відмінниць, двієчниць та середніх учениць. Однак якщо ми поставимо питання інакше: хто найзапекліший двієчник і хуліган, хто найталановитіший учень? - то виявиться, що це групи заповнені, зазвичай, хлопчиками. Тобто чоловіча підвиборка має більш спеціалізовану поведінку, що в цілому заважає адаптації на рівні індивіда. Усі крайнощі яскравіше представлені у чоловіків, але жінки більш навчальні.

Наступне питання, яке потрібно вирішити: чому чоловіки довгий час, іноді протягом багатьох поколінь тримають нові ознаки «в собі», і лише потім передають їх жінкам? Цю проблему Геодакян вирішує на генетичному рівні. Як відомо з генетики, у клітинах чоловіків існують так звані «ігрок-хромосоми» – власне, саме наявність цих хромосом відрізняє чоловіків від жінок. За теорією Геодакяна, нові гени з'являються одному кінці игрек-хромосомы, а перейти до жінок можуть лише з іншого кінця. З кожним наступним поколінням ген переміщається від початку хромосоми до кінця. Але поки він не досягне протилежного кінця, нова ознака передається тільки від батька до сина, але ніколи – до дочки. І доки так відбувається, нова ознака проходить перевірку часом.
Нещодавно розшифрування людського геному принесло ще одне підтвердження теорії Геодакяна. Виявилося, що більшість мутацій у людських генах справді виникають «в ігрек-хромосомах». Тобто чоловічі хромосоми – це справді експериментальна лабораторія, де виробляються нові гени.

Чоловіки як компас еволюції
З теорії Геодакяна випливає, що жінки йдуть в еволюції із деяким відставанням від чоловіків. Чудовий доказ цього знайдено на території Ізраїлю. У печерах гори Кармель виявлено скелети стародавніх людей, причому всі жінки – типові неандертальці, без підборіддя та з низьким лобом, а всі чоловіки – кроманьйонці, тобто сучасні люди. Вчені вважають, що плем'я кроманьйонців захопило жінок-неандертальців. Але Геодакян посміхається на це: а куди вони поділи тих чоловіків, і тим більше – своїх жінок?
На думку Геодакяна, це типовий приклад того, як чоловіки вже отримали нові ознаки - підбродок та високий лоб, а жінки за ними ще не підтягнулися.
Вивчаючи, чим чоловіки відрізняються від жінок (крім власне статевих ознак), можна зрозуміти, в який бік йде еволюція. Як відомо, у вищих тварин самці завжди більші за самок. Це означає, що ці істоти з еволюцією поступово збільшували свої розміри. Справді - предки слона були не більше свині, а предки коня - трохи більші за кішку. Але за теорією Геодакяна першими свої розміри збільшували самці, а самки за ними підтягувалися з деяким відставанням. Саме тому жеребці завжди більші за кобил - а кобила трохи ближче до свого дрібного, кішкоподібного предка.
У комах все навпаки - жінки набагато більші за чоловіків. Про що це каже? Про те, що еволюція йшла шляхом зменшення розмірів цих істот. У далекі часи на землі жили бабки з метровим розмахом крил, але їх «чоловіки» поступово дрібніли, а за чоловіками – але з відставанням, неохоче розлучаючись із розкішними розмірами – встигали жінки.

Але еволюція продовжується і сьогодні. Наприклад, дані атропометричних вимірів говорять, що як правило у чоловіків безіменний палець довший за вказівний, а у жінок навпаки, зазвичай довший за вказівний. По теорії Геодакяна це означає, що еволюція йде за «чоловічим» варіантом, і що через століття у всіх людей без винятку довшими пальцями буде безіменний.
Однак, – питаю я Вігена Артаваздовича, – цивілізація дала всім їжу та житло, сьогодні люди не гинуть від холоду, і, принаймні, не відрощують собі шерсть для боротьби з погодою. Чи не зупинилася еволюція людини?
Чому? – щиро дивується Геодакян. - Просто раніше чоловік гинув від зустрічі з тигром, а сьогодні його опрацьовують на науковій раді чи парткомі – та інфаркт.
Ось так. Відповідно до теорії Геодакяна парткоми виконували роль інструменту природного добору.
Чоловіки на право, жінки на ліво
Але Геодакян іде ще далі. Він вважає, що поділ живих істот на дві статі - лише частковий приклад поділу всього, що є в живій природі на ліву і праву половини. Усі нововведення з'являються праворуч. У риб серце знаходиться строго посередині, але коли риби вийшли на сушу і в них замість зябер з'явилося перше легке, то серце відповзло ліворуч, щоб поступитися місцем. Умовно кажучи, права половина в будь-якому живому тілі – «чоловіча», а ліва – «жіноча». Нові еволюційні ознаки йдуть шляхом з права на ліво, і від чоловіків - до жінок. Будь-яка новація в людському організмі з'являється з правого боку у чоловіків, а закінчує свій шлях на лівому боці у жінок. Ось, наприклад, очі в процесі еволюції поступово переповзали з боків (як у коней) на обличчя. Але якщо точно поміряти, то можна побачити, що у чоловіків у середньому очі трохи ближчі до носа, ніж у жінок. Жінки часто мають таку вражаючу східних поетів «волокістю» - широко розставленими очима, але з погляду еволюції це реліктова ознака. При цьому праве око завжди трохи ближче до носа, ніж ліве, так само як і праве вухо зазвичай трохи вище, ніж ліве.
Коли людина починає штучно виводити собак із довгими вухами, то у самців вуха відростають швидше, ніж у самок, а праві вуха у них обох - трохи швидше, ніж ліві. Найдовше вухо - у самця з права, найкоротше - у панночки зліва. Більше того, Віген Артаваздович навіть береться вивести математичний закон, у відповідність до якого сума довжин чоловічого правого та жіночого лівого вуха має дорівнювати сумі довжин чоловічого лівого та жіночого правого.
Зуби у людей поступово дрібніють. Тому, якщо поставити поруч приблизно однакових за зростом і комплекцією чоловіка і жінку, то найдрібніші зуби будуть у чоловіка праворуч, а найбільші – у жінок зліва.
Втім, жінкам не варто ображатися, що вони відстають. Адже зміни бувають не лише корисними. І. як каже Віген Геодакян, хоча серед чоловіків більше генієм, ніж серед жінок, але серед них також набагато більше повних недоумків. Краще варто згадати кумедну пісеньку, яку у кінофільмі «Труба» співає Олег Янковський голосом Анатолія Солов'яненка:
Чоловіки, я хочу нагадати вам,
Що пробним екземпляром був Адам!
На ньому працював Бог у міру сил,
На ньому спочатку досвід нагромадив.
Список літератури:
Уся світова література.

За мотивами статті Фрумкіна et al.
© В.А. Геодакян
ЕВОЛЮЦІЙНА ТЕОРІЯ ПІДЛОГИ В.А. Геодакян
Віген Артаваздович Геодакян, доктор біологічних наук, старший науковий співробітник Інституту еволюційної морфології та екології тварин ім. О.М. Северцова АН СРСР. Біолог-теоретик. Наукові інтереси - пов'язані зі статтю проблеми еволюції, генетики, екології, асиметрії мозку та психології, а також питання інформації та організації систем.На жаль, з технічних причин не наведені малюнки. V.V.
ЖОДНИЙ природний феномен не викликав такого інтересу і не містив так багато загадок, як стать. Проблемою статі займалися найбільші біологи: Ч. Дарвін, А. Воллес, А. Вейсман, Р. Гольдшмідт, Р. Фішер, Г. Меллер. А загадки залишалися і сучасні авторитети продовжували говорити про кризу еволюційної біології. "Підлога - головний виклик сучасної теорії еволюції... королева проблем еволюційної біології",- вважає Г. Белл - "Інтуїції Дарвіна та Менделя, які висвітлили так багато загадок, не змогли впоратися з центральною загадкою статевого розмноження". Для чого існують дві статі? Що дає?
Головні переваги статевого розмноження прийнято пов'язувати із забезпеченням генетичного розмаїття, придушенням шкідливих мутацій, перешкодою для спорідненого схрещування - інбридингу. Однак все це результат запліднення, яке є і у гермафродітів, а не диференціації (поділу) на дві статі. До того ж комбінаторний потенціал гермафродитного розмноження вдвічі вищий, ніж раздельнополого, а кількісна ефективність безстатевих методів вдвічі вища, ніж статевих. Виходить, роздільностатевий спосіб - найгірший? Чому ж тоді всі еволюційно прогресивні форми тварин (ссавці, птахи, комахи) та рослин (дводомні) роздільностатеві?
Автор цих рядків ще на початку 60-х років висловив думку, що диференціація статей - це економна форма інформаційного контакту з середовищем, спеціалізація за двома головними "аспектами еволюції - консервативним і оперативним". теорію, яка пояснює з єдиних позицій безліч різноманітних фактів і передбачає нові, сутність теорії і буде представлена у статті.
ДВА ПІДЛОГИ - ДВА ПОТОКИ ІНФОРМАЦІЇ
У принципі для системи можливі два вирішення цього конфлікту: бути на певній оптимальній "відстанні" від середовища або розділитися на дві сполучені підсистеми - консервативну та оперативну, першу "прибрати подалі" від середовища, щоб зберегти наявну інформацію, а другу "наблизити" до середовища для отримання нової. Друге рішення підвищує загальну стійкість системи, тому часто зустрічається серед еволюціональних, адаптивних, стежать систем (незалежно від їх конкретної природи) – біологічних, соціальних, технічних і т. д. Саме в цьому еволюційна логіка диференціації статей. Безстатеві форми "дотримуються" першого рішення, роздільностатеві - другого.
Якщо виділити два потоки інформації: генеративний (передача генетичної інформації від покоління до покоління, з минулого в майбутнє) та екологічний (інформація від середовища, із сьогодення в майбутнє), то легко переконатися, що дві статі по-різному беруть участь у них. У еволюції статі різних стадіях і рівнях організації виник цілий ряд механізмів, які послідовно забезпечували тісніший зв'язок жіночої статі з генеративним (консервативним) потоком, а чоловічого - з екологічним (оперативним). Так, у чоловічої статі порівняно з жіночою вище частота мутацій, менша адитивність успадкування батьківських ознак, вже норма реакції, вища агресивність і допитливість, активніша пошукова, ризикована поведінка та інші якості, що "наближають до середовища". Усі вони, цілеспрямовано виносячи чоловічу стать на периферію розподілу, забезпечують йому переважне отримання екологічної інформації. Інша група особливостей - величезна надмірність чоловічих гамет, їх малі розміри та висока рухливість, велика активність та мобільність самців, їх схильність до полігамії та інші етого-психологічні властивості. Тривалі періоди вагітності, годування та турботи про потомство у самок, практично підвищуючи ефективну концентрацію чоловічих особин, перетворюють чоловічу стать на "надлишковий", отже, "дешевий", а жіночий - на дефіцитний і більш цінний.
Це призводить до того, що відбір діє в основному за рахунок усунення чоловічих особин, "надмірність" і "дешевизна" дозволяють йому працювати з більшими коефіцієнтами. У результаті популяції зменшується кількість чоловічих особин, але великі потенційні можливості дозволяють їм запліднити всі жіночі. Невелика кількість чоловічих особин передає потомству стільки ж інформації, як і велика кількість жіночих, іншими словами, канал зв'язку з потомством у чоловічої статі ширше, ніж у жіночої. Значить, генетична інформація, передана по жіночій лінії, більш репрезентативна, а по чоловічій селективна, тобто в жіночій лінії повніше зберігається минуле розмаїття генотипів, у чоловічій - сильніше змінюється середній генотип.
Перейдемо до популяції - елементарної одиниці, що еволюціонує.
Будь-яка роздільностатева популяція характеризується трьома основними параметрами: співвідношенням статей (відношенням числа чоловічих особин до числа жіночих), дисперсією статей (відношенням значень дисперсії ознаки, або його різноманітності, у чоловічих і жіночих особин), статевим диморфізмом (відношенням середніх підлог). Приписуючи жіночому підлозі консервативну місію, а чоловічому - оперативну, теорія пов'язує ці параметри популяції з умовами середовища проживання і еволюційної пластичністю образу.
У стабільному (оптимальному) середовищі, коли немає необхідності нічого змінювати, сильні консервативні тенденції та еволюційна пластичність мінімальна. У рушійному (екстремальному) середовищі, коли потрібно підвищити пластичність, посилюються оперативні тенденції. В одних видів, скажімо нижчих ракоподібних, ці переходи здійснюються перемиканням з одного типу розмноження на інший (наприклад, в оптимальних умовах – партеногенетичний, в екстремальних – роздільностатевий). У більшості ж роздільностатевих видів ця регуляція плавна: в оптимальних умовах основні характеристики знижуються (знижується народжуваність чоловічих особин, звужується їх дисперсія, зменшується статевий диморфізм), а в екстремальних - ростуть (це екологічне правило диференціації статей).
Оскільки екологічний стрес призводить до різкого зростання, ці параметри популяції можуть бути індикатором стану екологічної ніші. У цьому зв'язку. показово, що народжуваність хлопчиків у Каракалпакії за останнє десятиліття підвищилася на 5%. Згідно з екологічним правилом, основні параметри повинні зростати за будь-яких природних чи соціальних катаклізм (сильні землетруси, війни, голод, переселення тощо). Тепер про елементарний крок еволюції.
ПЕРЕТВОРЕННЯ ГЕНЕТИЧНОЇ ІНФОРМАЦІЇ В ОДНОМУ ПОКОЛІННІ
Генотип - це програма, яка в різних середовищах може реалізуватися в один із цілого спектру фенотипів (ознак). Отже, в генотипі записано певне значення ознаки, а діапазон можливих значень. В онтогенезі реалізується один, найкращий для конкретного середовища фенотип. Отже, генотип задає діапазон реалізацій, середовище "вибирає" точку всередині цього діапазону, ширина якого є норма реакції, що характеризує ступінь участі середовища у визначенні ознаки
За одними ознаками, наприклад групі крові чи кольору очей, норма реакції вузька, тому середовище мало впливає ними;, за іншими - психологічним, інтелектуальним здібностям - дуже широка, тому багато хто пов'язують лише з впливом середовища, т. е. вихованням; треті ознаки, скажімо зростання, маса, займають проміжне становище.
З урахуванням двох відмінностей статей - за нормою реакції (яка ширша у жіночих особин) і перерізу каналу зв'язку (ширше у чоловічих особин) - розглянемо перетворення генетичної інформації в одному поколінні, тобто від зигот до зигот, ставши білізуючою і рушійною середовищі . Припустимо, що вихідний розподіл генотипів у популяції однаково для чоловічих і жіночих зигот, тобто статевий диморфізм за ознакою відсутня. Щоб з розподілу генотипів зигот отримати розподіл фенотипів (організмів до і після відбору), з нього, у свою чергу, розподіл генотипів яйцеклітин і сперміїв, і, нарешті, розподіл зигот наступного покоління, достатньо простежити перетворення двох крайніх генотипів зигот на крайні фенотипи, гамети і знову в зиготи. Інші генотипи проміжні і залишаться такими у всіх розподілах. Більш широка норма реакції жіночої статі дозволяє йому за рахунок модифікаційної пластичності залишити зони відбору, зберегти та передати потомству весь спектр вихідних генотипів.
Вузька норма реакції чоловічої статі змушує його залишитися в зонах елімінації і зазнати інтенсивного відбору. Тому чоловіча стать передає наступному поколінню лише вузьку частину вихідного спектру генотипів, що максимально відповідає умовам середовища в даний момент. У стабілізуючому середовищі це середня частина спектру, у рушійному - край розподілу. Отже, генетична інформація, передана жіночою статтю нащадку, більш репрезентативна, а передана чоловічою - більш селективна. Інтенсивний відбір зменшує кількість чоловічих особин, але оскільки на освіту зигот потрібно рівну кількість чоловічих і жіночих гамет, чоловічим особинам доводиться запліднювати не одну жіночу. Широкий переріз каналу чоловічої статі це дозволяє. Отже, у кожному поколінні популяції яйцеклітини широкого розмаїття, що несуть інформацію про минулому багатстві генотипів, зливаються зі сперміями вузького розмаїття, генотипи яких містять інформацію лише про найбільш відповідних поточних умов середовища. Таким чином, наступне покоління отримує інформацію про минуле по материнській лінії, про сьогодення – за батьківською.
У стабілізуючому середовищі середні генотипи чоловічих і жіночих гамет однакові, відрізняються лише їх дисперсії, тому генотипний розподіл зигот наступного покоління збігається з вихідним. Єдиний результат диференціації статей при цьому зводиться до розплати популяції за екологічну інформацію "дешевшою" чоловічою статтю. Інша картина в рушійному середовищі, де зміни зачіпають як дисперсії, а й середні значення генотипів. Виникає генотипічний статевий диморфізм гамет, що є нічим іншим, як запис (фіксацію) екологічної інформації у розподілі чоловічих гамет. Яка його подальша доля?
Якщо батьківська генетична інформація передається синам та дочкам стохастично, при заплідненні вона повністю змішається та статевий диморфізм зникне. Але якщо існують будь-які механізми, що перешкоджають повному змішанню, деяка частка цієї інформації потрапить від батьків лише до синів і, отже, частина статевого диморфізму збережеться у зиготів. А такі механізми є. Наприклад, тільки синам потрапляє інформація з генів У-хромосоми; по-різному проявляються гени у нащадків, залежно від цього, успадковані вони від батька чи матері. Без подібних бар'єрів важко пояснити також домінування батьківського генотипу у нащадків від реципрокних схрещувань, відоме у тваринництві, скажімо, висока удійність корів, що передається через бика. Все це дозволяє вважати, що достатньо лише відмінностей статей за нормою реакції та перерізу каналу зв'язку, щоб у рушійному середовищі вже в одному поколінні виник генотипічний статевий диморфізм, який при зміні поколінь накопичуватиметься і зростатиме.
ДИМОРФІЗМ І ДИХРОНІЗМ У ФІЛОГЕНЕЗІ
Отже, коли для цієї ознаки стабілізуюче середовище стає рушійним, починається еволюція ознаки у чоловічої. статі, а в жіночої він зберігається, т. е. настає дивергенція ознаки, з мономорфного він перетворюється на диморфний.
З кількох можливих еволюційних сценаріїв дозволяють вибрати єдиний два очевидні факти: еволюціонують обидві статі; існують ознаки як моно-, і диморфні. Це можливо лише в тому випадку, якщо фази еволюції ознаки у статей зрушені за часом: у чоловічої зміни ознака починається і закінчується раніше, ніж у жіночої. При цьому, згідно з екологічним правилом, мінімальна в стабілізуючому середовищі дисперсія ознаки розширюється з початком еволюції та звужується після його завершення.
Траєкторія еволюції ознаки роздвоюється на чоловічу та жіночу гілки, з'являється і зростає статевий диморфізм. Це дивергентна фаза, в якій швидкість еволюції та дисперсія ознаки болю чоловічої статі. Через багато поколінь і в жіночої статі починає розширюватися дисперсія та змінюватись ознака. Статевий диморфізм, досягнувши оптимуму, залишається незмінним. Це паралельна фаза: швидкості еволюції ознаки та її дисперсії в обох статей постійні і рівні. Коли у чоловічої статі ознака досягає нового, стабільного значення, дисперсія звужується і еволюція припиняється, але ще продовжується у жіночої статі. Це конвергентна фаза, в якій швидкість еволюції та дисперсія більша у жіночої статі. Статевий диморфізм поступово зменшується і коли ознака у статей стає однаковим, зникає, а дисперсії вирівнюються і стають мінімальними. Цим завершується диморфна стадія еволюції ознаки, за якою знову слідує мономорфна, або стадія стабільності.
Таким чином, вся філогенетична траєкторія еволюції ознаки складається з мономорфних і диморфних стадій, що чергуються, наявність же самого диморфізму теорія розглядає як критерій еволюції ознаки.
Отже, статевий диморфізм за будь-якою ознакою тісно пов'язаний з його еволюцією: з'являється з її початком, зберігається поки вона йде, і зникає, як тільки еволюція закінчується. Отже, статевий диморфізм - наслідок як статевого відбору, як вважав Дарвін, але будь-якого: природного, статевого, штучного. Це неодмінна стадія, модус еволюції будь-якої ознаки у роздільностатевих форм, пов'язана з утворенням "дистанції" між статями по морфологічній та хронологічній осях. Статевий диморфізм і статевий дихронізм - це два виміри загального явища - дихрономорфізм.
Сказане можна сформулювати у вигляді філогенетичних правил статевого диморфізму та дисперсії статей: якщо за якоюсь ознакою існує популяційний статевий диморфізм, то ознака еволюціонує від жіночої форми до чоловічої; якщо дисперсія ознаки більша у чоловічої статі - фаза дивергентна, дисперсії рівні - паралельна, дисперсія більше у жіночої статі - фаза конвергентна. За першим правилом можна визначити напрямок еволюції ознаки, за другим - її фазу, або пройдений шлях. Скориставшись правилом статевого диморфізму, можна зробити цілий ряд передбачень, що легко перевіряються. Так, виходячи з того, що еволюція більшості видів хребетних супроводжувалася збільшенням розмірів, можна встановити напрямок статевого диморфізму - у великих форм самці, як правило, більші за самок. І навпаки, оскільки багато комах і павукоподібних в ході еволюції дрібнішали, у дрібних форм самці повинні бути менше самок.
Правило легко перевірити на сільськогосподарських тварин та рослинах, штучну еволюцію (селекцію) яких спрямовувала людина. Селекційні – господарсько цінні – ознаки мають бути більш просунуті у самців. Таких прикладів багато: у м'ясних порід тварин – свиней, овець, корів, птахів – самці швидше ростуть, набирають вагу і дають кращу якість м'ясо; жеребці перевершують кобил спортивними та робочими якостями; барани тонкорунних порід дають у 1,5-2 рази більше вовни, ніж вівці; у самців хутрових звірів хутро краще, ніж у самок; самці шовкопряда дають на 20% більше шовку і т.д.
Перейдемо тепер від масштабу філогенезу часу до онтогенетичного.
ДИМОРФІЗМ І ДИХРОНІЗМ В ОНТОГЕНЕЗІ
Якщо кожну з фаз філогенетичного сценарію спроектувати на онтогенез (за законом рекапітуляції, онтогенез - коротке повторення філогенезу), можна отримати відповідні їм шість (три фази в еволюційній стадії і три в стабільній; доеволюційна, післяеволюційна та міжеволюційна) розвитку. Дихронізм проявиться в онтогенезі як вікове запізнення у розвитку ознаки у жіночої статі, тобто домінування жіночої форми диморфної ознаки на початку онтогенезу та чоловічої наприкінці. Це онтогенетичне правило статевого диморфізму: якщо за якоюсь ознакою існує статевий популяційний диморфізм, в онтогенезі ця ознака змінюється, як правило, від жіночої форми до чоловічої. Інакше висловлюючись, ознаки материнської породи з віком мають слабшати, а батьківській - посилюватися. Перевірка цього правила за двома десятками антропометричних ознак повністю підтверджує передбачення теорії. Яскравий приклад - розвиток рогів у різних видів оленів та антилоп: чим сильніша "рогатість" виду, тим раніше в онтогенезі з'являються роги спочатку у самців, а потім у самок. Та ж закономірність - вікове запізнювання розвитку у жіночої статі за функціональною асиметрією мозку - виявлено С. Вітельзон. Вона дослідила здібності 200 праворуких дітей впізнавати предмети на дотик лівою та правою рукою та з'ясувала, що хлопчики вже у 6 років мають правопівкульну спеціалізацію, а дівчатка до 13 років – "симетричні".
Описані закономірності відносяться до диморфних ознак, що еволюціонують. Але є і мономорфні, стабільні, за якими в нормі статевий диморфізм відсутній. Це фундаментальні ознаки видового і вищих рангів спільності, такі як багатоклітинність, теплокровність, загальний для обох статей план будови тіла, число органів і т. д. За теорією, якщо їх дисперсія більше у чоловічої статі, то фаза доеволюційна, якщо у жіночої - післяеволюційна В останній фазі теорія передбачає існування "реліктів" статевого диморфізму і дисперсії статей у патології. диморфізму: вроджені аномалії, що мають атавістичну природу, частіше повинні з'являтися у жіночої статі, а мають футуристичну природу (пошук) - у чоловічої, наприклад, серед новонароджених дітей із наднормативним числом нирок, ребер, хребців, зубів і т.д. , що зазнали редукції числа в ході еволюції, має бути більше дівчаток, а з їх нестачею - хлопчиків, медична статистика це підтверджує: серед 2 тис. дітей, що народилися з однією ниркою, приблизно в 2,5 рази більше хлопчиків, а серед 4 тис. дітей із трьома нирками майже вдвічі більше дівчаток. Такий розподіл - невипадково, він відбиває еволюцію видільної системи. Отже, три нирки у дівчаток – це повернення до предкового типу розвитку, атавістичне спрямування; одна нирка у хлопчиків – футуристичне, продовження редукційної тенденції. Аналогічна і статистика за аномальним числом ребер. З вивихом стегна, уродженою пороком, з якою діти краще бігають і лазять по деревах, ніж здорові, дівчаток народжується у п'ять-шість разів більше, ніж хлопчиків.
Подібна картина й у розподілі вроджених вад серця та магістральних судин. З 32 тис. вивірених діагнозів у всіх "жіночих" пороках переважали елементи, властиві серцю ембріона або філогенетичних попередників людини: відкритий овальний отвір у міжпередсердній перегородці, незаростаюча ботала протока (судина, що з'єднує у плоду легеневу артерію). вади найчастіше були новими (пошук): ні в філогенезі, ні в ембріонів не мали аналогій - різного роду стенози (звуження) та транспозиції магістральних судин.
Перелічені правила охоплюють диморфні ознаки, властиві обох статей. А як із ознаками, властивими тільки одній підлозі, такими як несучість, удійність? Фенотиповий статевий диморфізм за такими ознаками має абсолютний, організмовий характер, але спадкова інформація про них записана в генотипі обох статей. Тому, якщо вони еволюціонують, за ними має існувати генотипічний статевий диморфізм, який можна виявити у реципрокних гібридів. За такими ознаками (у числі інших еволюціонують) теорія передбачає напрямок реципрокних ефектів. У реципрокних гібридів за ознаками батьків, що диергують, повинна домінувати батьківська форма (порода), а по конвергуючих - материнська. Це еволюційне правило реципрокних ефектів. Воно дає дивовижну можливість виявити велику генотипічну просунутість чоловічої статі навіть за суто жіночими ознаками. Таке, здавалося б, па^ радоксальне передбачення теорії повністю підтверджується: в одній і тій же породі бики генотипно "подійніше" корів, а півні більш "яйценоски", ніж кури, тобто ці ознаки передаються переважно самцями.
Проблеми еволюції переважно ставляться до " чорним ящикам " без входу - у яких прямий експеримент неможливий. Необхідну інформацію еволюційне вчення черпало із трьох джерел: палеонтології, порівняльної анатомії та ембріології. Кожна з них має суттєві обмеження, оскільки охоплює лише частину ознак. Сформульовані правила дають новий метод для еволюційних досліджень абсолютно за всіма ознаками роздільностатевих форм. Тому особливу цінність метод представляє вивчення еволюції людини, таких її ознак, як темперамент, інтелект, функціональна асиметрія мозку, вербальні, просторово-зорові, творчі здібності, гумор та інші психологічні властивості, яких традиційні методи не застосовні.
ФУНКЦІОНАЛЬНА АСИМЕТРІЯ МОЗКУ І ПСИХОЛОГІЧНІ ОСОБЛИВОСТІ
Довгий час її вважали привілеєм людини, пов'язуючи з промовою, праворукістю, самосвідомістю, вважали, що асиметрія вторинна – наслідок цих унікальних особливостей людини. Наразі встановлено, що асиметрія широко поширена у плацентарних тварин, більшість дослідників визнають також і різницю її виразності у чоловіків та жінок. Дж. Леві вважає, наприклад, що жіночий мозок подібний до мозку чоловіка-шульги, тобто менш асиметричний, ніж у чоловіка-правші.
З позицій теорії статі більш асиметричний мозок у чоловіків (і самців деяких хребетних) означає, що еволюція йде від симетрії до асиметрії. Статевий диморфізм по асиметрії мозку дає надію зрозуміти та пояснити відмінності у здібностях та схильностях чоловіків та жінок.
Відомо, що у наших далеких філогенетичних предків були бічні очі (у людських ембріонів ранніх стадій розвитку вони розташовуються так само), зорові поля не перекривалися, кожне око було пов'язане лише з протилежною півкулею (контралатеральні зв'язки). В процесі еволюції очі перемістилися на лицьову сторону, зорові поля перекрилися, але щоб виникла стереоскопічна картина, зорова інформація від обох очей мала зосередитися в одній області мозку.
Зір став стереоскопічним тільки після того, як виникли додаткові – іпсилатеральні – волокна, які поєднали ліве око з лівою півкулею, праве – з правим. Значить, іпсилатеральні зв'язки еволюційно молодші за контралатеральні, а тому у чоловіків вони повинні бути більш просунуті, тобто іпсилатеральних волокон у зоровому нерві більше.
Оскільки об'ємна уява та просторово-зорові здібності пов'язані зі стереоскопією (і кількістю іпсиволокон), у чоловіків вони мають бути розвинені краще, ніж у жінок. І справді, психологам добре відомо, що у розумінні геометричних завдань чоловіки набагато перевершують жінок, як й у читанні географічних карт, орієнтуванні біля і т.д.
Як виник психологічний статевий диморфізм, з погляду теорії статі? Немає принципової різниці в еволюції морфофізіологічних та психологічних чи поведінкових ознак. Широка норма реакції жіночої статі забезпечує йому вищу, ніж у чоловічої статі, пластичність (адаптивність) в онтогенезі. Це стосується і психологічних ознак. Відбір у зонах дискомфорту у чоловічої та жіночої статі йде в різних напрямках: завдяки широкій нормі реакції жіноча стать може "вибратися" з цих зон за рахунок виховності, навченості, конформності, тобто загалом - адаптивності. Для чоловічої статі такий шлях закрито через вузьку норму реакції; тільки винахідливість, кмітливість, винахідливість можуть забезпечити виживання в дискомфортних умовах. Іншими словами, жінки пристосовуються до ситуації, чоловіки виходять із неї, знайшовши нове рішення, дискомфорт стимулює пошук.
Тому чоловіки охочіше беруться за нові, що вимагають пошуку, неординарні завдання (часто виконуючи їх учорне), а жінки краще доводять вирішення знайомих завдань до досконалості. Чи не тому вони процвітають у тих видах діяльності, в яких можна обійтися добре відшліфованими навичками, наприклад у роботі на конвеєрі?
Якщо оволодіння мовою, листом, будь-яким ремеслом розглянути в еволюційному аспекті, можна виділити фазу пошуку (знаходження нових рішень), освоєння та фазу закріплення, вдосконалення. Чоловіча перевага у першій фазі та жіноча у другій виявлено у спеціальних дослідженнях.
Новаторство у будь-якій справі – місія чоловічої статі. Чоловіки першими освоювали всі професії, види спорту, навіть в'язання, в якому зараз монополія жінок незаперечна, винайшли чоловіки (Італія, XIII ст.). Роль авангарду належить чоловікам і у схильності до деяких хвороб і соціальних пороків. Саме чоловіча стать частіше піддається "новим" хворобам, або, як їх називають, хворобам століття,; цивілізації, урбанізації - атеросклерозу, раку, шизофренії, СНІДу, а також соціальним вадам - алкоголізму, куріння, наркоманії, азартних ігор, злочинності тощо.
За теорією, повинні існувати і два протилежні типи психічних хвороб, пов'язаних з авангардною роллю чоловічої статі та ар'єргардної - жіночої.
Патологія, що супроводжується недостатньою асиметрією мозку, малими розмірами мозолистого тіла та великими передньою комісурою, повинна у два – чотири рази частіше зустрічатися у жінок, аномалії з протилежними характеристиками – у чоловіків. Чому?
Якщо за кількісною ознакою немає відмінностей між статями, то розподіл його значень у популяції часто описується кривою гауса. Дві крайні області такого розподілу і є зони патології - "плюс" і "мінус" відхилення, від норми, в кожну з яких чоловічі та жіночі особини потрапляють з рівною ймовірністю. Але якщо статевий диморфізм існує, то у кожної статі ознака розподіляється по -своєму, утворюються дві розсунуті на величину статевого диморфізму криві.Оскільки вони залишаються всередині загальнопопуляційного розподілу, то одна зона патології виявиться збагаченою чоловічими особинами, інша - жіночими.До речі, так само пояснюється характерна для населення майже всіх країн світу "статева спеціалізація" багатьох інших хвороб.
Наведені приклади показують, як "працює" теорія статі лише в деяких проблемах людини, насправді вона охоплює набагато більший масив явищ, у тому числі й соціальний аспект.
Оскільки диморфний стан ознаки свідчить про те, що він на "еволюційному марші", максимальними повинні бути відмінності за останніми еволюційними придбаннями людини-абстрактного мислення, творчих здібностей, просторової уяви, гумору, саме вони повинні переважати у чоловіків. І справді, видатні вчені, композитори, художники, письменники, режисери – переважно чоловіки, а серед виконавців багато жінок.
Проблема статі зачіпає дуже важливі області інтересів людини: демографію та медицину, психологію та педагогіку, дослідження алкоголізму, наркоманії та злочинності, через генетику вона пов'язана з економікою. Правильна соціальна концепція статі необхідна вирішення проблем народжуваності і смертності, сім'ї та виховання, професійної орієнтації. Така концепція повинна будуватися на природній біологічній основі, бо без розуміння біологічних, еволюційних ролей чоловічої та жіночої статі не можна правильно визначити їхні соціальні ролі.
Тут наведено лише деякі загальнобіологічні висновки теорії статі, з єдиних позицій пояснено різноманітні, раніше незрозумілі явища та факти, згадані прогностичні можливості. Отже, підсумуємо. Еволюційна теорія статі дозволяє:
- 1) передбачити поведінку основних характеристик роздільностатевої популяції в стабільному (оптимальному) та рушійному (екстремальному) середовищах;
- 2) диференціювати еволюціонуючі та стабільні ознаки;
- 3) визначити напрямок еволюції будь-якої ознаки;
- 4) встановити фазу (пройдений шлях) еволюції ознаки;
- 5) визначити середню швидкість еволюції ознаки: V = диморфізм / дихронізм
- 6) передбачити шість різних варіантів онтогенетичної динаміки статевого диморфізму, що відповідають кожній фазі філогенезу;
- 7) передбачити напрямок домінування ознаки батьківської або материнської породи у реципрокних гібридів;
- 8) передбачити та розкрити "релікти" дисперсії статей та статевого диморфізму в галузі вроджених патологій;
- 9) встановити зв'язок між віковою та статевою епідеміологією.
Отже, спеціалізація жіночої статі щодо збереження генетичної інформації, а чоловічої - щодо її зміни досягається гетерохронною еволюцією статей. Отже, стать - це не так спосіб розмноження, як прийнято вважати, скільки метод асинхронної еволюції.
Оскільки представлена тут робота - плід теоретичних роздумів та узагальнень, не можна не сказати кілька слів про роль теоретичних досліджень у біології. Природознавство, за словами відомого фізика, лауреата Нобелівської премії Р. Міллікена, рухається на двох ногах – теорії та експерименті. Але такі справи - у фізиці, в біології ж панує культ фактів, вона досі живе спостереженнями та експериментом, теоретичної біології як такої, аналога теоретичної фізики не існує. Звичайно, це пов'язано зі складністю живих систем, звідси і скепсис біологів, які звикли йти традиційним шляхом - від фактів та експериментів до узагальнюючих висновків та теорії. Але чи може наука про живе, як і раніше, залишатися суто емпіричною в "століття біології", який, за визнанням багатьох сучасників, йде на зміну "століття фізики"? Здається, що й біології настав час стати на обидві ноги.
Література
Bell G., The Masterprice of Nature. The Evolution and Genetics of Sexuality, London, 1982.
. Геодакян В. А. // Пробл. передачі інформ. 1965. Т. 1. № 1. С. 105-112.
. Детальніше див; Геодакян В. А. Еволюційна логіка диференціації статей // Природа. 1983. № 1. С. 70-80.
. Геодакян У. А. // Докл. АН СРСР. 1983. Т. 269. № 12. С. 477-482.
. Vitelson S.F..// Science. 1976. V. 193. М 4251. Р. 425-427.
. Геодакян Ст А., Шерман А. Л.// Журн. заг. біології. 1971. Т. 32. № 4. С. 417-424.
. Геодакян В. А.// Системні дослідження: методологічні проблеми. Щорічник. 1986. М., 1987. С. 355-376.
. Геодакян В. А. Теорія диференціації статей у проблемах людини // Людина у системі наук. М., 1989. С. 171-189.
 Завантажити роботу на тему: Науково-дослідна робота студентів та шляхи її вдосконалення
Завантажити роботу на тему: Науково-дослідна робота студентів та шляхи її вдосконалення У чому полягає суперечливість суспільного процесу
У чому полягає суперечливість суспільного процесу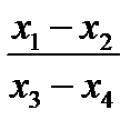 Вимірювальні шкали Абсолютна шкала вимірювань
Вимірювальні шкали Абсолютна шкала вимірювань