البروتينات: تاريخ البحث ، التركيب الكيميائي ، الخصائص ، الوظائف البيولوجية. ما العناصر المكونة للبروتينات وما هي خصائصها؟ ما هو موجود في البروتينات
التركيب الكيميائي للبروتينات.
3.1. السندات الببتيد
البروتينات عبارة عن بوليمرات غير منتظمة مبنية من بقايا حمض أميني بيتا ، ويمكن كتابة الصيغة العامة لها في محلول مائي عند قيم pH قريبة من المحايد على أنها NH 3 + CHRCOO -. ترتبط بقايا الأحماض الأمينية في البروتينات برابطة أميدية بين مجموعات -amino و-carboxyl. رابطة الببتيد بين اثنينيُشار عادةً إلى بقايا الأحماض الأمينية بيتا السندات الببتيد ، والبوليمرات المبنية من بقايا الأحماض الأمينية ألفا المتصلة بواسطة روابط الببتيد تسمى بولي ببتيدات. يمكن أن يكون البروتين كهيكل مهم بيولوجيًا إما عديد ببتيد واحد أو عدة ببتيدات متعددة تشكل معقدًا واحدًا نتيجة للتفاعلات غير التساهمية.
3.2 التركيب الأولي للبروتينات
عند دراسة التركيب الكيميائي للبروتينات ، من الضروري معرفة ، أولاً ، ما هي العناصر الكيميائية التي تتكون منها ، وثانيًا ، بنية مونومراتها. للإجابة على السؤال الأول ، يتم تحديد التركيب الكمي والنوعي للعناصر الكيميائية للبروتين. أظهر التحليل الكيميائي موجود في جميع البروتينات كربون (50-55٪) ، أكسجين (21-23٪) ، نيتروجين (15-17٪) ، هيدروجين (6-7٪) ، كبريت (0.3-2.5٪). تم العثور أيضًا على الفوسفور واليود والحديد والنحاس وبعض العناصر الكبيرة والصغرى الأخرى في تكوين البروتينات الفردية ، بكميات مختلفة ، وغالبًا ما تكون صغيرة جدًا.
يمكن أن يختلف محتوى العناصر الكيميائية الرئيسية في البروتينات ، باستثناء النيتروجين الذي يتميز تركيزه بأكبر ثبات ويبلغ متوسطه 16٪. بالإضافة إلى ذلك ، فإن محتوى النيتروجين في المواد العضوية الأخرى منخفض. وفقًا لهذا ، تم اقتراح تحديد كمية البروتين بواسطة النيتروجين المكون له. مع العلم أن 1 جرام من النيتروجين موجود في 6.25 جرام من البروتين ، يتم ضرب الكمية الموجودة من النيتروجين في معامل 6.25 ويتم الحصول على كمية البروتين.
لتحديد الطبيعة الكيميائية لمونومرات البروتين ، من الضروري حل مشكلتين: فصل البروتين إلى مونومرات ومعرفة تركيبها الكيميائي. يتم تحقيق تفكك البروتين إلى الأجزاء المكونة له عن طريق التحلل المائي - غليان البروتين لفترة طويلة مع الأحماض المعدنية القوية. (التحلل الحمضي)أو أسباب (تحلل قلوي). غالبًا ما يستخدم الغليان عند 110 درجة مئوية مع حمض الهيدروكلوريك لمدة 24 ساعة ، وفي المرحلة التالية ، يتم فصل المواد التي يتكون منها التحلل المائي. لهذا الغرض ، يتم استخدام طرق مختلفة ، في أغلب الأحيان كروماتوغرافيا (لمزيد من التفاصيل ، انظر فصل "طرق البحث ..."). الأحماض الأمينية هي الجزء الرئيسي من التحلل المائي المنفصل.
3.3 أحماض أمينية
حاليًا ، تم العثور على ما يصل إلى 200 من الأحماض الأمينية المختلفة في كائنات مختلفة من الحياة البرية. في جسم الإنسان ، على سبيل المثال ، هناك حوالي 60 نوعًا ، لكن البروتينات تحتوي فقط على 20 حمضًا أمينيًا ، وأحيانًا تسمى الأحماض الأمينية الطبيعية.
الأحماض الأمينية هي أحماض عضوية يتم فيها استبدال ذرة الهيدروجين في ذرة الكربون بمجموعة أمينية - NH 2. لذلك ، بطبيعتها الكيميائية ، هذه أحماض بيتا الأمينية مع الصيغة العامة:
H - C - NH 2
من هذه الصيغة يمكن ملاحظة أن تكوين جميع الأحماض الأمينية يشمل المجموعات العامة التالية: - CH 2 ، - NH 2 ، - COOH. السلاسل الجانبية (الجذور - ص) الأحماض الأمينية تختلف. كما يتضح من الملحق الأول ، فإن الطبيعة الكيميائية للجذور متنوعة: من ذرة الهيدروجين إلى المركبات الحلقية. الجذور هي التي تحدد السمات الهيكلية والوظيفية للأحماض الأمينية.
جميع الأحماض الأمينية ، باستثناء أبسط الأحماض الأمينية بالنسبة لك الجلايسين (NH 3 + CH 2 COO) لها ذرة مراوان C ويمكن أن توجد على شكل متشابهين (أيزومرات بصرية):
COO-COO-
NH3 + صص NH3 +
إل-ايزوميرد-ايزومير
تشتمل جميع البروتينات المدروسة حاليًا على أحماض أمينية فقط من السلسلة L ، والتي ، إذا أخذنا في الاعتبار الذرة اللولبية من جانب ذرة H ، فإن مجموعات NH 3 + و COO وجذر R تقع في اتجاه عقارب الساعة. إن الحاجة إلى بناء جزيء بوليمر مهم بيولوجيًا من متماثل محدد بدقة أمر واضح - من خليط راسمي من اثنين من المتضادمات ، يمكن الحصول على خليط معقد بشكل لا يمكن تصوره من الأيزومرات ثنائية الاتجاه. لا يزال السؤال عن سبب استناد الحياة على الأرض إلى البروتينات المبنية على وجه التحديد من الأحماض الأمينية L- وليس D-- لغزًا مثيرًا للاهتمام. تجدر الإشارة إلى أن الأحماض الأمينية D منتشرة إلى حد ما بطبيعتها ، علاوة على ذلك ، فهي جزء من oligopeptides ذات أهمية بيولوجية.
تُبنى البروتينات من عشرين نوعًا من الأحماض الأمينية الأساسية ، ومع ذلك ، يتم تكوين باقي الأحماض الأمينية المتنوعة تمامًا من هذه الأحماض الأمينية العشرين الموجودة بالفعل في تكوين جزيء البروتين. من بين هذه التحولات ، ينبغي للمرء أولاً وقبل كل شيء أن يلاحظ التكوين جسور ثاني كبريتيد أثناء أكسدة اثنين من بقايا السيستين في تكوين سلاسل الببتيد المشكلة بالفعل. نتيجة لذلك ، يتم تكوين بقايا حمض ديامينوديكاربوكسيليك من اثنين من بقايا السيستين سيستين (انظر الملحق الأول). في هذه الحالة ، يحدث الارتباط المتقاطع إما داخل سلسلة بولي ببتيد واحدة أو بين سلسلتين مختلفتين. كبروتين صغير يحتوي على سلسلتين متعدد الببتيد متصلتين بجسور ثاني كبريتيد ، بالإضافة إلى روابط متقاطعة داخل إحدى سلاسل البولي ببتيد:
جيفككاSVCSLYQLENYCN
FVNQHLCGSHLVEALYLVCجيرجفيتبكا
مثال مهم لتعديل بقايا الأحماض الأمينية هو تحويل بقايا البرولين إلى بقايا هيدروكسي برولين :
N-CH-CO-N-CH-CO-
CH 2 CH 2 CH 2 CH 2
CH2CHOH
يحدث هذا التحول ، وعلى نطاق واسع ، أثناء تكوين مكون بروتيني مهم للنسيج الضام - الكولاجين .
نوع آخر مهم جدًا من تعديل البروتين هو فسفرة مجموعات الهيدروكسو من بقايا السيرين والثريونين والتيروزين ، على سبيل المثال:
- NH - CH - CO - - NH - CH - CO -
CH 2 OH CH 2 OPO 3 2 -
الأحماض الأمينية في محلول مائي هي في حالة متأينة بسبب تفكك مجموعات الأمينو والكربوكسيل التي تشكل الجذور. بمعنى آخر ، فهي مركبات مذبذبة ويمكن أن توجد إما كأحماض (مانحون بروتون) أو كقاعدة (متقبلات مانحة).
جميع الأحماض الأمينية ، حسب التركيب ، تنقسم إلى عدة مجموعات:
لا دوري. أحماض أمينية أحادية أمينومونوكربوكسيليةتحتوي في تركيبتها على أمين واحد ومجموعة كربوكسيل واحدة ، في محلول مائي تكون محايدة. يحتوي بعضها على ميزات هيكلية مشتركة ، مما يسمح بالنظر فيها معًا:
جلايسين وألانين.الجلايسين (الجليكوكول أو الأحماض الأمينية) غير نشط بصريًا - إنه الحمض الأميني الوحيد الذي لا يحتوي على مادة متناهية الصغر. يشارك الجليسين في تكوين المادة الصفراوية والنووية إلى الهيم ، وهو ضروري لتحييد المنتجات السامة في الكبد. يستخدم الجسم ألانين في عمليات التمثيل الغذائي للكربوهيدرات والطاقة المختلفة. الأيزومير -alanine هو جزء لا يتجزأ من فيتامين بانتوثينيك بالنسبة لك ، أنزيم A (CoA) ، والمواد المستخرجة للعضلات.
سيرين وثريونين.إنهم ينتمون إلى مجموعة أحماض هيدروكسي ، لأن. لديك مجموعة الهيدروكسيل. السرين هو جزء من الإنزيمات المختلفة ، البروتين الرئيسي للحليب - الكازين ، بالإضافة إلى العديد من البروتينات الدهنية. يشارك ثريونين في التخليق الحيوي للبروتين ، كونه حمض أميني أساسي.
السيستين والميثيونين.أحماض أمينية تحتوي على ذرة كبريت. يتم تحديد قيمة السيستين من خلال وجود مجموعة السلفهيدريل (-SH) في تركيبته ، مما يمنحها القدرة على أكسدة الجسم بسهولة وحماية الجسم من المواد ذات القدرة العالية على الأكسدة (في حالة الإصابة الإشعاعية ، الفوسفور تسمم). يتميز الميثيونين بوجود مجموعة ميثيل سهلة الحركة ، والتي تستخدم لتخليق مركبات مهمة في الجسم (الكولين ، الكرياتين ، الثايمين ، الأدرينالين ، إلخ).
فالين وليوسين وآيزولوسين.وهي عبارة عن أحماض أمينية متفرعة تشارك بنشاط في عملية التمثيل الغذائي ولا يتم تصنيعها في الجسم.
أحماض أمينية أحادية الكربوكسيلتحتوي على مجموعة أمينية واثنين من مجموعات الكربوكسيل وتعطي تفاعلًا حمضيًا في محلول مائي. وتشمل هذه الأسبارتيك والجلوتامين بالنسبة لك ، والأسباراجين والجلوتامين. هم جزء من الوسطاء المثبطين للجهاز العصبي.
الأحماض الأمينية Diaminomonocarboxylicفي محلول مائي لها تفاعل قلوي بسبب وجود مجموعتين أمين. بالنسبة لهم ، يعتبر اللايسين ضروريًا لتخليق الهستونات وأيضًا في عدد من الإنزيمات. يشارك أرجينين في تركيب اليوريا والكرياتين.
دوري. تحتوي هذه الأحماض الأمينية على نواة عطرية أو حلقية غير متجانسة في تكوينها ، وكقاعدة عامة ، لا يتم تصنيعها في جسم الإنسان ويجب تزويدها بالطعام. يشاركون بنشاط في مجموعة متنوعة من عمليات التمثيل الغذائي. لذا
فينيل ألانين هو المصدر الرئيسي لتخليق التيروزين ، وهو مقدمة لعدد من المواد المهمة بيولوجيًا: الهرمونات (هرمون الغدة الدرقية ، الأدرينالين) ، وبعض الأصباغ. التربتوفان ، بالإضافة إلى مشاركته في تخليق البروتين ، هو أحد مكونات فيتامين PP ، السيروتونين ، التربتامين ، وعدد من الأصباغ. الهيستيدين ضروري لتخليق البروتينات ، وهو مقدمة للهيستامين ، الذي يؤثر على ضغط الدم وإفراز العصارة المعدية.
الخصائص
البروتينات هي مركبات جزيئية كبيرة. هذه عبارة عن بوليمرات تتكون من مئات وآلاف من بقايا الأحماض الأمينية - مونومرات.
البروتينات لها وزن جزيئي مرتفع ، بعضها قابل للذوبان في الماء ، وقادر على الانتفاخ ، ويتميز بالنشاط البصري ، والتنقل في مجال كهربائي ، وبعض الخصائص الأخرى.
تشارك البروتينات بنشاط في التفاعلات الكيميائية. ترجع هذه الخاصية إلى حقيقة أن الأحماض الأمينية التي تتكون منها البروتينات تحتوي على مجموعات وظيفية مختلفة يمكن أن تتفاعل مع مواد أخرى. من المهم أن تحدث مثل هذه التفاعلات أيضًا داخل جزيء البروتين ، مما يؤدي إلى تكوين الببتيد وثاني كبريتيد الهيدروجين وأنواع أخرى من الروابط. لجذور الأحماض الأمينية ، وعلى التوالي و الكتلة الجزيئيةتتراوح البروتينات بين 10000 و 1000000. لذلك ، يحتوي الريبونوكلياز (إنزيم يفكك الحمض النووي الريبي) على 124 بقايا من الأحماض الأمينية ويبلغ وزنها الجزيئي 14000 تقريبًا. يحتوي الميوغلوبين (بروتين عضلي) ، الذي يتكون من 153 من بقايا الأحماض الأمينية ، على جزيئات الوزن 17000 ، والهيموغلوبين - 64500 (574 من مخلفات الأحماض الأمينية). الأوزان الجزيئية للبروتينات الأخرى أعلى: -globulin (تشكل أجسامًا مضادة) يتكون من 1250 من الأحماض الأمينية ويبلغ وزنها الجزيئي حوالي 150.000 ، والوزن الجزيئي لإنزيم نازعة هيدروجين الجلوتامات يتجاوز 1000000.
يتم تحديد الوزن الجزيئي من خلال طرق مختلفة: مقياس التناضح ، الترشيح الهلامي ، البصري ، إلخ. ومع ذلك ، فإن الطريقة الأكثر دقة هي طريقة الترسيب التي اقترحها T. Svedberg. يعتمد على حقيقة أنه أثناء التنبيذ الفائق مع تسارع يصل إلى 900000 جم ، يعتمد معدل ترسيب البروتين على وزنه الجزيئي.
أهم خاصية للبروتينات هي قدرتها على إظهار كل من الحمضية والقاعدية ، أي أنها تعمل مذبذبالشوارد. يتم ضمان ذلك من خلال مجموعات منفصلة مختلفة تشكل جذور الأحماض الأمينية. على سبيل المثال ، يتم نقل الخصائص الحمضية إلى البروتين بواسطة مجموعات الكربوكسيل من الأحماض الأمينية الغلوتامية الأسبارتيك ، بينما يتم نقل الخصائص القلوية بواسطة جذور الأرجينين والليسين والهيستيدين. كلما زاد عدد الأحماض الأمينية ثنائية الكربوكسيل التي يحتوي عليها البروتين ، زادت خصائصه الحمضية والعكس صحيح.
تحتوي هذه المجموعات أيضًا على شحنات كهربائية تشكل الشحنة الكلية لجزيء البروتين. في البروتينات التي تسود فيها الأحماض الأمينية الأسبارتيك والجلوتامين ، تكون شحنة البروتين سالبة ؛ ففائض الأحماض الأمينية الأساسية يعطي شحنة موجبة لجزيء البروتين. نتيجة لذلك ، في مجال كهربائي ، تتحرك البروتينات نحو القطب السالب أو الأنود ، اعتمادًا على حجم شحنتها الإجمالية. لذلك ، في البيئة القلوية (الرقم الهيدروجيني 7-14) ، يتبرع البروتين بالبروتون ويصبح سالبًا ، بينما في البيئة الحمضية (الرقم الهيدروجيني 1-7) ، يتم قمع تفكك المجموعات الحمضية ويصبح البروتين كاتيونًا.
وبالتالي ، فإن العامل الذي يحدد سلوك البروتين ككاتيون أو أنيون هو تفاعل الوسط ، والذي يتم تحديده بواسطة تركيز أيونات الهيدروجين ويتم التعبير عنه بقيمة الأس الهيدروجيني. ومع ذلك ، عند بعض قيم الأس الهيدروجيني ، يتساوى عدد الشحنات الموجبة والسالبة ويصبح الجزيء محايدًا كهربائيًا ، أي لن تتحرك في مجال كهربائي. تُعرَّف قيمة الأس الهيدروجيني للوسط بأنها نقطة تساوي الكهرباء للبروتينات. في هذه الحالة ، يكون البروتين في أقل حالة استقرارًا ، ومع تغيرات طفيفة في درجة الحموضة إلى الجانب الحمضي أو القلوي ، فإنه يترسب بسهولة. بالنسبة لمعظم البروتينات الطبيعية ، تكون النقطة الكهربية في بيئة حمضية قليلاً (الرقم الهيدروجيني 4.8 - 5.4) ، مما يشير إلى غلبة الأحماض الأمينية ثنائية الكربوكسيل في تركيبها.
تكمن الخاصية المتذبذبة في خواص التخزين المؤقت للبروتينات ومشاركتها في تنظيم درجة الحموضة في الدم. قيمة الرقم الهيدروجيني لدم الإنسان ثابتة وتتراوح بين 7.36 - 7.4 ، على الرغم من المواد المختلفة ذات الطبيعة الحمضية أو الأساسية ، والتي يتم تزويدها بانتظام بالطعام أو تتشكل في عمليات التمثيل الغذائي - لذلك ، هناك آليات خاصة لتنظيم القاعدة الحمضية توازن البيئة الداخلية للجسم. وتشمل هذه الأنظمة التي تم النظر فيها في الفصل. نظام عازلة الهيموجلوبين "التصنيف" (الصفحة 28). يشير التغيير في درجة الحموضة في الدم بأكثر من 0.07 إلى تطور عملية مرضية. يسمى التحول في درجة الحموضة إلى الجانب الحمضي بالحماض ، ويسمى الجانب القلوي القلاء.
من الأهمية بمكان بالنسبة للجسم قدرة البروتينات على امتصاص بعض المواد والأيونات على سطحها (الهرمونات والفيتامينات والحديد والنحاس) ، والتي تكون إما ضعيفة الذوبان في الماء أو سامة (البيليروبين والأحماض الدهنية الحرة). تنقلها البروتينات عبر الدم إلى أماكن أخرى من التحولات أو التحييد.
المحاليل المائية للبروتينات لها خصائصها الخاصة. أولاً ، البروتينات لها انجذاب كبير للماء ، أي أنهم محبة للماء.هذا يعني أن جزيئات البروتين ، مثل الجسيمات المشحونة ، تجذب ثنائيات أقطاب الماء ، والتي توجد حول جزيء البروتين وتشكل قشرة مائية أو قشرة هيدراتية. تحمي هذه القشرة جزيئات البروتين من الالتصاق ببعضها البعض والترسيب. يعتمد حجم قشرة الماء على بنية البروتين. على سبيل المثال ، ترتبط الألبومات بسهولة بجزيئات الماء ولها قشرة مائية كبيرة نسبيًا ، في حين أن الجلوبيولين والفيبرينوجين يعلقان الماء بشكل أسوأ ، وتكون قشرة الماء أصغر. وبالتالي ، فإن استقرار المحلول المائي للبروتين يتحدد بعاملين: وجود شحنة على جزيء البروتين وقشرة الماء المحيطة به. عندما تتم إزالة هذه العوامل ، يترسب البروتين. يمكن أن تكون هذه العملية قابلة للعكس ولا رجعة فيها.
ترسيب عكسي للبروتينات(التمليح) ينطوي على ترسيب البروتين تحت تأثير بعض المواد ، وبعد إزالته يعود إلى حالته الأصلية (الأصلية). تستخدم أملاح الفلزات الأرضية القلوية والقلوية لتمليح البروتينات (غالبًا ما يستخدم الصوديوم وكبريتات الأمونيوم في الممارسة العملية). تزيل هذه الأملاح قشرة الماء (تسبب الجفاف) وتزيل الشحنة. هناك علاقة مباشرة بين حجم القشرة المائية لجزيئات البروتين وتركيز الأملاح: كلما كانت قشرة الماء أصغر ، قل الأملاح المطلوبة. وهكذا ، فإن الجلوبيولين ، الذي يحتوي على جزيئات كبيرة وثقيلة وقشرة مائية صغيرة ، يترسب عندما يكون المحلول مشبعًا بشكل غير كامل بالأملاح ، والألبومين ، كجزيئات أصغر محاطة بقشرة مائية كبيرة ، تترسب عندما يكون المحلول مشبعًا تمامًا.
جزيء البروتين الأصلي
جزيء بروتين مشوه. تشير الشرطات إلى الروابط الموجودة في جزيء البروتين الأصلي التي تنكسر أثناء التمسخ.


يرتبط الترسيب غير القابل للانعكاس بالتغيرات العميقة داخل الجزيئية في بنية البروتين ، مما يؤدي إلى فقدان خصائصها الأصلية (قابلية الذوبان ، والنشاط البيولوجي ، وما إلى ذلك). يسمى هذا البروتين بالتشويه ، والعملية تمسخ. يحدث تمسخ البروتين في المعدة ، حيث توجد بيئة حمضية بقوة (درجة الحموضة 0.5 - 1.5) ، وهذا يساهم في تكسير البروتينات بواسطة الإنزيمات المحللة للبروتين. يعتبر تمسخ البروتين هو الأساس لعلاج التسمم بالمعادن الثقيلة ، عندما يتم إعطاء المريض حليبًا أو بيضًا نيئًا حسب نظام التشغيل ("عن طريق الفم") بحيث تفسد المعادن بروتينات الحليب أو البيض.
تم امتصاصها على سطحها ولم تؤثر على بروتينات الغشاء المخاطي للمعدة والأمعاء ، ولم يتم امتصاصها في الدم.
يقع حجم جزيئات البروتين في حدود 1 ميكرومتر إلى 1 نانومتر ، وبالتالي فهي كذلك غروانيالجزيئات التي تشكل المحاليل الغروانية في الماء. تتميز هذه المحاليل بلزوجة عالية ، والقدرة على تشتيت أشعة الضوء المرئية ، ولا تمر عبر أغشية شبه منفذة.
تعتمد لزوجة المحلول على الوزن الجزيئي وتركيز المذاب. كلما زاد الوزن الجزيئي ، زاد لزوجة المحلول. تشكل البروتينات كمركبات جزيئية كبيرة محاليل لزجة. على سبيل المثال ، محلول بياض البيض في الماء.
ماء

لا تمر الجسيمات الغروانية عبر الأغشية شبه المنفذة (السيلوفان ، الغشاء الغرواني) ، لأن مسامها أصغر من الجسيمات الغروية. جميع الأغشية البيولوجية غير منفذة للبروتينات. تُستخدم خاصية محاليل البروتين هذه على نطاق واسع في الطب والكيمياء لتنقية مستحضرات البروتين من الشوائب. تسمى عملية الفصل هذه بغسيل الكلى. تكمن ظاهرة غسيل الكلى في عمل جهاز "الكلى الاصطناعية" ، والذي يستخدم على نطاق واسع في الطب لعلاج الفشل الكلوي الحاد.
غسيل الكلى (الدوائر البيضاء الكبيرة عبارة عن جزيئات بروتينية ، والدوائر السوداء عبارة عن جزيئات كلوريد الصوديوم)
معادن الحليب
يحتوي رماد الحليب على معادن مثل الكالسيوم والفوسفور والمغنيسيوم والبوتاسيوم والصوديوم والكلور والكبريت والسيليكون. يتم تحديد كمية العناصر الفردية في الحليب بشكل أساسي من خلال العوامل الوراثية. التغذية والعوامل البيئية الأخرى لها تأثير طفيف فقط على صيانتها. تظل كمية المعادن في الحليب ثابتة حتى في حالة وجود القليل من العناصر الفردية في النظام الغذائي. مع عدم كفاية تناول المعادن مع الطعام ، يتم تعبئة احتياطيات الجسم وبالتالي يتم الحفاظ على تركيزها في الحليب عند مستوى معين. مع وجود نقص كبير في عنصر واحد أو أكثر ، يظل المحتوى المعدني لكل وحدة حجم من الحليب ثابتًا إلى حد ما. ومع ذلك ، فإن إنتاجية الحليب ، ومن ثم الكمية الإجمالية للمعادن في الحليب ، تنخفض.
|
المعادن |
يحتوي ، ز |
المعادن |
|
الكمية الإجمالية للعناصر النزرة في الحليب أقل من 0.15٪ ، ويعتمد محتوى العناصر النزرة في الحليب بشكل وثيق على وجودها في العلف.
الخصائص الهيكلية والميكانيكية للزيت.
وفقًا لـ Rebinder ، هناك نوعان رئيسيان من الهياكل.
النوع الأول هو بنية التخثر- هذه هي الشبكات المكانية التي تنشأ عن الالتصاق العشوائي لأصغر جسيمات الطور المشتت أو الجزيئات الدقيقة من خلال طبقات رقيقة من وسط معين.
النوع الثاني هو هيكل التبلور-التكثيف، والتي تتشكل نتيجة للنمو البيني المباشر للبلورات مع تكوين مادة صلبة متعددة البلورات.
تنتمي القواعد الدهنية للسمن النباتي إلى نوع التخثر من الهياكل. يتم تحديد خصائص الاتساق والبلاستيك للقواعد الدهنية للمارجرين بشكل أساسي من خلال نسبة المراحل الصلبة والسائلة في دهون معينة صالحة للأكل. هذه النسبة من الأطوار الصلبة والسائلة نموذجية لبعض ظروف التبلور المحددة (درجة الحرارة ، الوقت ، الخلط). في هذه الحالة ، يكون لتكوين الوسيط المستمر والمرحلة المشتتة وطبيعة موقع المرحلة المشتتة في الوسط السائل المستمر أهمية كبيرة.
بالنسبة لبعض أنواع الدهون الصالحة للأكل عند درجة حرارة معينة وظروف التبلور ، قد تتجاوز كمية المرحلة الصلبة المشتتة نسبة الطور الأمثل ، ومن ثم تتشكل مثل هذه الأغشية الرقيقة من وسط سائل مستمر على سطح البلورات بحيث لا تستطيع ذلك. تتداخل مع الالتحام الفوضوي الهائل للبلورات مع بعضها البعض. في هذه الحالة ، سيكون لدينا دائمًا أكبر صلابة للقاعدة الدهنية ، والاتساق المتفتت وأسوأ خصائص البلاستيك.
إذا كانت أفلام الوسط السائل المستمر في درجة حرارة الغرفة هي السماكة المثلى ، أي بحيث لا تخلق ظروفًا لانصهار البلورات أثناء التخزين ، في ظل تأثيرات ميكانيكية أو حرارية على النظام ، ففي هذه الحالة المثالية سنحصل دائمًا على هياكل تخثر معززة تحدد أفضل الخصائص البلاستيكية للقواعد الدهنية.
من أجل الحصول على هياكل التخثر المقواة بأفضل خصائص البلاستيك ، غالبًا ما يتم إدخال نوعين من شحم الخنزير بنقطة انصهار تبلغ 32 درجة مئوية و 42 درجة مئوية في تركيبة قاعدة الدهون في الخارج. في هذه الحالة ، يتم إدخال كمية كبيرة إلى حد ما من الزيوت النباتية السائلة. هذا ، من ناحية ، يخلق أفضل نسبة من المراحل الصلبة والسائلة في قاعدة الدهون ، مما يوفر اتساقًا مشابهًا للزبدة ، ومن ناحية أخرى ، يخلق ظروفًا لاتساق قوام المارجرين في نطاق درجة حرارة واسع إلى حد ما. إلى جانب ذلك ، فإن إدخال الشحم عالي الذوبان في قاعدة الدهون يتعارض مع متطلبات علماء الفسيولوجيا لتكوين الدهون الغذائية.
بادئ ذي بدء ، تجدر الإشارة إلى أن وجود مثبتات مستحلب عالية الفعالية فقط هو الذي جعل من الممكن إنشاء تقنية حديثة في إنتاج المارجرين وضمان إنتاج منتج دهني عالي الجودة صالح للأكل. توفر المضافات النشطة على السطح مستحلبًا مشتتًا بدقة في رابطة قوية بين جزيئات الطور المشتت والوسط المستمر (الدهون الصلبة في درجة حرارة الغرفة). القضية الرئيسية في إنتاج المارجرين هي تأثير المضافات السطحية النشطة على الخواص التركيبية والميكانيكية للمارجرين ، وخاصة على القدرة على الذوبان.
تعمل طبقة الامتصاص الخاصة بالمستحلب على زيادة ثبات المستحلب ، خاصة في الحالات التي يتم فيها تكوين هذه الطبقة ، مما يؤدي إلى تكوين طبقة من هلام السطح مع زيادة اللزوجة والقوة بشكل كبير.
هذه الخصائص لها أهمية خاصة لإنتاج المارجرين ، حيث أن المنتج النهائي عبارة عن مستحلب من أصغر جزيئات الطور السائل ، موزعة بشكل موحد في وسط مستمر من المرحلة الصلبة عند درجة حرارة الغرفة.
ترتبط مشكلة قوة المستحلب ارتباطًا وثيقًا بمسألة نوع المستحلبات المتكونة مع مستحلب معين. هناك إمكانية لتشكيل نوعين. يتم تفسير قيمة نسبة أحجام الطور لنوع معين من المستحلب المتكون من خلال حقيقة أن اندماج وفصل مستحلب من نوع معين يحدث بشكل أكثر كثافة ، كلما كان حجم وسط التشتت أصغر وكلما كانت المرحلة المشتتة أكبر . إذا كان المستحلب يوفر مستحلبًا ثابتًا من نوع واحد فقط ، فإن نسبة الحجم تتوقف عن أن تكون حاسمة في تحديد نوع المستحلب. لا يعتمد الانعكاس على نسبة أحجام الطور فحسب ، بل يعتمد أيضًا على التركيز والطبيعة الكيميائية للمستحلب.
يجب أن تحتوي المستحلبات على الخصائص التالية:
تقليل التوتر السطحي
- سريع بما يكفي ليتم امتصاصه على واجهة الطور ، مما يمنع اندماج القطرات ؛
- لها بنية جزيئية محددة مع مجموعات قطبية وغير قطبية ؛
- يؤثر على لزوجة المستحلب.
فعالية المستحلب هي خاصية محددة تعتمد على طبيعتها ، ونوع المواد القابلة للاستحلاب ، ودرجة الحرارة ، ودرجة الحموضة للوسط ، والتركيز ، ووقت الاستحلاب ، وما إلى ذلك.
تحدد فعالية الإجراء وطبيعة المستحلب نوع المستحلب.
المستحلبات المحبة للماء ، قابلة للذوبان في الماء بشكل أفضل من الهيدروكربونات ، تساهم في تكوين مستحلبات الزيت والماء ، وهي كارهة للماء ، قابلة للذوبان بشكل أفضل في الهيدروكربونات ، مستحلبات زيت الماء. تتميز نسبة أحجام الأجزاء القطبية وغير القطبية لجزيئات المستحلب بمؤشر خاص - التوازن المحبة للماء - المحبة للدهون. إذا كان HLB للمستحلب هو 3-6 ، يتم تكوين مستحلب زيت الماء ، بقيمة HLB من 8-13 ، يتم تكوين مستحلب الزيت والماء في الغالب.
المارجرين هو مستحلب فائق التبريد بالماء في الزيت. في هذه الحالة ، لا يتم استبعاد إمكانية تكوين مستحلب من النوع المختلط مع غلبة مستحلب الزيت والماء.
الوظائف الرئيسية للمستحلبات:
تكوين مستحلب مستقر شديد التشتت ؛
- التثبيت ومنع فصل الرطوبة والدهون في المنتج النهائي ؛
- ضمان الاستقرار أثناء التخزين ؛
- توفير القدرة على مقاومة الطرطشة أثناء القلي ؛
- ضمان اللدونة ؛
- ضمان إنشاء شكل مستقر للشبكة البلورية في عملية تكوين الهيكل ؛
- ضمان الخصائص الوظيفية المحددة للمنتج النهائي ، حسب مساحة استخدام السمن.
في أوكرانيا ، لسنوات عديدة ، تم استخدام المستحلبات المنتجة في روسيا ومن إنتاجنا الخاص ، والتي يتم إنتاجها في الإنتاج شبه الصناعي. وتشمل هذه المستحلبات:
T-1 هو أحد منتجات تحلل الدهون في لحم البقر أو شحم الخنزير.
- Т-2 - منتج بلمرة الجلسرين ، أسترة بحمض دهني ؛
- T-F - خليط من مستحلب T-1 وتركيز فوسفاتيد الطعام بنسبة 2: 1 ؛
- PMD - أحادي جليسريد الطعام ؛
- CE - مستحلب مشترك - خليط من PMD وتركيز الفوسفاتيد بنسبة 3: 1.
مجموعة واسعة من مستحلبات نبات نيجني نوفغورود - أنواع مختلفة من أحادي الجليسريد المقطر. في الوقت الحاضر ، تم إتقان إنتاج سلسلة من المستحلبات الجديدة القائمة على الليسيثين في نيجني نوفغورود. هذه هي الليسيثين القياسي ، الليسيثين المجزأ - الفوسفاديتيالكولين والفوسفاديليسيرين ، وكذلك الليسيثين المتحلل بالماء.
في السنوات الأخيرة ، تم استخدام المستحلبات ذات التعديلات المختلفة لسلسلة Dimodan و Palsgaard (في بعض شركات Quest) بشكل أساسي في أوكرانيا.
في فترات مختلفة ، تحولت ميزة الطلب على هذين النوعين من المستحلبات من أحدهما إلى الآخر. يمكننا القول أن هناك منافسة الجودة - السعر.
اعتمادًا على محتوى الدهون في المارجرين ونطاقه ، يتم استخدام Dimodan PVP (Dimodan HP) و Dimodan OT (Dimodan S-T PEL / B) ومستحلبات Dimodan SR. بالنسبة للسمن النباتي الذي يحتوي على نسبة دهون أقل من 40٪ ، والتي يطلبها السكان حاليًا ، بالإضافة إلى ذلك (باستثناء Dimodan OT ، أو Dimodan СР. ، أو Dimodan LS) ، يتم استخدام استرات البولي جلسرين وحمض الريسينوليك - Grinsted PGPR90.
في إنتاج السمن قليل الدسم ، خاصة تلك التي تحتوي على نسبة دهون 25٪ أو أقل ، يتم استخدام أنظمة التثبيت - الغروانيات المائية (الجينات ، البكتين ، إلخ).
وتجدر الإشارة إلى أن الشركات المصنعة تقدم توصيات بشأن استخدام أنواع مختلفة من المستحلبات وأنظمة التثبيت ، اعتمادًا على الغرض من المارجرين. يتيح لك الامتثال لهذه التوصيات الحصول على منتجات عالية الجودة.
بروتينات العضلات
تحتوي لحوم الدواجن على حوالي 20-23٪ بروتينات. يمكن تقسيم بروتينات العضلات إلى ثلاث مجموعات حسب قابليتها للذوبان: البروتينات الليفية العضلية ، والبروتينات الساركوبلازمية ، والبروتينات اللحمية.
عضلي ليفي، أو قابل للذوبان في الملح السناجبغير قابل للذوبان في الماء ، ولكن معظمه يذوب في محاليل كلوريد الصوديوم بتركيز يزيد عن 1٪. تتكون هذه المجموعة من حوالي 20 بروتينًا فرديًا تشكل اللييفات العضلية للعضلة المقلصة. يمكن تقسيم بروتينات اللييفات العضلية إلى ثلاث مجموعات حسب وظيفتها: الانقباض ، المسؤول عن تقلصات العضلات ، والتنظيم ، والمشاركة في التحكم في عملية الانكماش ، والهيكل الخلوي ، وربط اللييفات العضلية والمساعدة في الحفاظ على سلامتها الهيكلية.
بروتينات مقلصةللميوسين والأكتين تأثير كبير على وظيفة بروتين العضلات. نظرًا لوجود الأكتين والميوسين كمركب أكتيوموسين في العضلات المتيبسة ، فإن وظيفة الميوسين تتغير في كل من منتجات الدواجن المستحلب والمقولبة. تعتمد خصائص المنتجات أيضًا على النسبة الإجمالية للأكتين والميوسين ونسبة الميوسين والأكتين في الحالة الحرة. تؤثر البروتينات الساركوبلازمية والبروتينات اللحمية بدورها على الخصائص الوظيفية لبروتينات اللييفات العضلية.
بروتينات الساركوبلازمقابل للذوبان في الماء أو في محاليل ذات قوة أيونية منخفضة (
بروتينات ستروما، غالبًا ما يشار إليها باسم بروتينات النسيج الضام ، تعمل بمثابة سقالات تدعم بنية العضلات. الكولاجين هو البروتين الرئيسي في السدى. يشكل الإيلاستين والشبكية جزءًا صغيرًا من السدى. كل هذه البروتينات غير قابلة للذوبان في الماء والمحاليل المالحة. تميل حنان اللحم إلى الانخفاض مع تقدم العمر بسبب الارتباط المتبادل والتغيرات الأخرى في الكولاجين.
الدم وكسوره
يستخدم الدم الكامل كمواد خام رئيسية لإنتاج النقانق والقريدس والأطعمة المعلبة وغيرها من المنتجات الغذائية ، بالإضافة إلى مادة مضافة تعطي اللون التقليدي للمنتجات عند استخدام مستحضرات البروتين (0.6-1.0٪) ؛ لنفس الغرض ، يتم استخدام مستحضر الهيموجلوبين أو خليط من العناصر المكونة بعد الترطيب في الماء (1: 1).
بالمقارنة مع الأنواع الأخرى من المواد الخام المحتوية على البروتين ، لا يتم استخدام الدم الكامل على نطاق واسع بسبب وجود لون وطعم معينين يغيران الخصائص الحسية للمنتجات النهائية. في الوقت الحالي ، يجري البحث عن تنقية الدم ، ومع ذلك ، ولعدد من الأسباب ، لم تجد الطرق المقترحة تطبيقًا عمليًا في الصناعة. تعتمد الخصائص الوظيفية والتكنولوجية للدم وجزيئاته (البلازما ، المصل) بشكل أساسي على تكوين البروتين. يحتوي الدم الكامل على حوالي 150 بروتينًا لها خصائص فيزيائية كيميائية مختلفة ، وغالبًا ما تتكون بروتينات العناصر والألبومين والجلوبيولين والفيبرينوجين. في هذا الصدد ، على أساس الدم الكامل ، يُنصح بإعداد مستحلبات مخصصة لإدخالها في تركيبات منتجات اللحوم وزيادة استقرار أنظمة اللحوم ، والقيمة الغذائية والمحصول ، وتحسين المؤشرات الحسية والخصائص الهيكلية والميكانيكية.
كإعداد للبروتين ، يُنصح باستخدام عزل الصويا أو كازينات الصوديوم.
يمكن أن يصل مستوى إدخال المستحلبات المحضرة على أساس الدم الكامل في أنظمة اللحوم إلى 30-40٪ بوزن المادة الخام الرئيسية.
تحتوي بروتينات بلازما الدم على مركب FCS فريد من نوعه. تتفاعل البومينات بسهولة مع البروتينات الأخرى ، ويمكن أن تترافق مع الدهون والكربوهيدرات ، ولها قدرة عالية على ربط الماء ورغوة.
الجلوبيولين هي مستحلبات جيدة.
الفيبرينوجين - له قدرة واضحة على تكوين الهلام ، ويتحول إلى ليفبرين تحت تأثير عدد من العوامل (تحول الأس الهيدروجيني إلى نقطة متساوية ، وإدخال أيونات Ca ++ في البلازما) وتشكيل إطار مكاني.
المخاليط يمكن استخدام خصائص الفيبرينوجين هذه في إنتاج بروتين متعدد المكونات يحتوي على ، بما في ذلك مواد تشبه مادة الهلام ، في عملية تكوين البنية الثانوية لمستحلبات اللحوم في إنتاج النقانق المسلوقة.
تتميز جميع بروتينات البلازما بقابلية ذوبان جيدة ، ونتيجة لذلك - قدرة عالية على الارتباط بالماء والاستحلاب ، فهي قادرة على تكوين مواد هلامية عند تسخينها. إن إدخال ملح الطعام له تأثير سلبي على ثبات المستحلبات المعتمدة على بلازما الدم عند درجة الحموضة 7.0. أهم خصائص البلازما هي قدرتها على تكوين المواد الهلامية أثناء المعالجة الحرارية ، وتعتمد قوتها ومستوى قدرتها على الارتباط بالماء على تركيز البروتينات في النظام ، وقيمة الأس الهيدروجيني ، ووجود الأملاح ، ودرجة الحرارة ومدة تدفئة.
يزيد إدخال البروتينات غير البلازمية (زلال البيض ، عزل الصويا ، كازينات الصوديوم) في البلازما بشكل كبير من قوة المواد الهلامية وقدرتها على امتصاص الماء والدهون بعد المعالجة الحرارية.
اعتمادًا على حالة بلازما الدم وظروف المعالجة الأولية ، قد يختلف تكوينها وخصائصها الوظيفية والتكنولوجية ، وبالتالي ، قد يختلف مجال الاستخدام.
يتيح لنا تنظيم البيانات المتوفرة حاليًا حول معالجة الكمبيوتر الشخصي تقييم الأساليب الحديثة لإدراك الإمكانات البيولوجية والوظيفية والتكنولوجية لمكون البروتين في الكمبيوتر الشخصي في إنتاج الغذاء.
يعطي المخطط فكرة عن الحالة وطرق المعالجة وتكوين وخصائص مستحضرات البروتين التي تم الحصول عليها على أساس PA ، ويحدد مجالات استخدامها العملي ، وينعكس تعدد الوظائف للغرض المقصود من PA في شكل FCS بطريقة معالجة أو بأخرى.
وتجدر الإشارة إلى أن مستوى مؤشرات FCS الفردية الواردة في الجدول 13 والمستخدمة لفك رموز الرموز المعتمدة في المخطط نسبي نظرًا لحقيقة أن القيمة الفعلية لكل خاصية تعتمد بشكل حاسم على تركيز البروتين ، وقيمة الأس الهيدروجيني في النظام ، ودرجة حرارة الوسط ، والقوة الأيونية وعدد من العوامل الأخرى.
يُظهر تحليل مخطط التصنيف أن إحدى طرق الاستخدام التكنولوجي لبلازما الدم هي استخدامه في شكل سائل مستقر (وكذلك بعد التبريد والتجميد) مع محتوى منخفض نسبيًا من البروتين والحفاظ على FCS الأصلي.
في هذه الحالة ، تتميز بروتينات PC بمستوى عالٍ من BCC والاستحلاب ، ويرجع ذلك إلى وجود بروتينات قابلة للذوبان في الماء يمكن أن تشكل مواد هلامية عند تسخينها. إن الجمع بين هذه الخصائص يجعل من الممكن استخدام البلازما على نطاق واسع ليس فقط كمكون يوازن التركيب الكيميائي العام للمنتجات النهائية ، ولكن أيضًا كإضافة وظيفية في إنتاج منتجات اللحوم المستحلب ذات المحتوى الرطوبي النهائي العالي: النقانق المسلوقة ، النقانق والنقانق والمنتجات نصف المصنعة المفرومة والأغذية المعلبة المفرومة ومنتجات لحم الخنزير. الأكثر عقلانية هو إدخال 10٪ بلازما في التركيبات بدلاً من 3٪ لحم بقري أو 2٪ لحم خنزير ؛ يوفر إدخال 20 ٪ من أجهزة الكمبيوتر الشخصية بدلاً من الماء أثناء القطع تحسينًا في المعلمات الحسية والهيكلية والميكانيكية وزيادة في إنتاجية المنتجات النهائية بنسبة 0.3-0.5 ٪. يتم الحصول على تأثير ممتاز من خلال استخدام بلازما الدم كوسيلة لترطيب مستحضرات البروتين (3-4 أجزاء من الكمبيوتر لكل جزء من تحضير البروتين).
لا غنى عن الكمبيوتر الشخصي في تصنيع مستحلبات البروتين والدهون ، والمواد الرابطة ، وأنظمة البروتين متعددة المكونات مع تركيبة معينة وخصائص وظيفية وتكنولوجية ، ومستحضرات بروتينية مهيكلة.
يؤدي تركيز PA عن طريق التجفيف ، والترشيح الفائق ، والتركيز بالتبريد ، مما يجعل من الممكن زيادة محتوى البروتين بشكل كبير ، إلى بعض التعديل في FTS للدواء.
تجفيف البلازما له تأثير كبير بشكل خاص على درجة التغيير في FCS ، في حين أن تركيز PA الجاف المعرض للترشيح الفائق له خصائص وظيفية عالية جدًا.
يتم استخدام المركزات التي تم الحصول عليها بهذه الطرق بنجاح في إنتاج منتجات اللحوم جنبًا إلى جنب مع الكمبيوتر السائل.
يعتقد الخبراء الأمريكيون أن بلازما الدم البقري ، بفضل FCS ، يمكن أن تحل محل بياض البيض بنجاح.
ترسيب التمسخ والتخثر ، الذي يوفر مزيجًا من عمليات التركيب الحراري والتلبد (الترسيب) وتركيز بروتينات الكمبيوتر ، يجعل من الممكن الحصول على مستحضرات بتركيز بروتين مرتفع نسبيًا و FCS غير عادي ، مما يسمح باستخدامها في وصفات نصف مدخن ، مدخن ، نقانق كبدة ، أغذية معلبة باتيه ومنتجات نصف نهائية ذات محتوى رطوبة نهائية محدود وقدرة عالية على امتصاص الدهون. هذه المجموعة من الأدوية تشمل: "بروتين البلازما المترسب" ، "رواسب بروتين البلازما" ، ليفيكس ، "جبن البلازما" ، حبيبات PC.
إن استخدام هذه الأنواع من مستحضرات بلازما الدم في ممارسة إنتاج اللحوم محدود للغاية.
توسع هيكلة بلازما الدم عن طريق إعادة الحساب بشكل كبير من إمكانيات استخدامها التكنولوجي. يتيح تحويل أجهزة الكمبيوتر والأنظمة متعددة المكونات المستندة إليها إلى شكل جل الحصول على مصفوفات هيكلية تحاكي الكائنات البيولوجية الطبيعية في المظهر والتكوين والخصائص ، ويخلق متطلبات أساسية لتنظيم دائرة الجمارك الفيدرالية ، ويضمن مشاركة منخفضة- تصنيف المواد الخام في عملية الإنتاج ، ويجعل من الممكن الاقتراب من الحل من مواقع جديدة.تطوير أنواع جديدة من المنتجات الغذائية. فعال بشكل خاص هو الاستخدام المعقد للكمبيوتر الشخصي ومستحضرات البروتين (عزلات الصويا ، كازينات الصوديوم ، إلخ). ، نظائر منتجات اللحوم.
نضج اللحوم
لم يتم توضيح موضوع "إنضاج اللحوم" بعد. من ملاحظات الممارسين ، من المعروف أنه بعد انتهاء حياة الحيوان ، تحدث تغيرات فيزيائية كيميائية في اللحم ، تتميز بالصلابة ، ثم استرخاء (تليين) ألياف العضلات. نتيجة لذلك ، يكتسب اللحم بعض النكهة ويسهل طهيه. يتم تحسين قيمته الغذائية. تسمى هذه التغييرات في الأنسجة الرخوة للذبيحة "النضج" ("النضج") أو "تخمر اللحم".
لشرح عملية نضج اللحوم ، فإن تعاليم Meyerhof و Embden و Palladin و Abdergalden حول ديناميكيات واستقلاب الكربوهيدرات في العضلات أثناء حياة الحيوان تستحق اهتمامًا كبيرًا.
أظهر مايرهوف أن الجليكوجين الموجود في العضلات ينفق على تكوين حمض اللاكتيك أثناء تقلص العضلات. أثناء الاسترخاء
(الراحة) العضلات ، بسبب إمداد الأكسجين ، يتم تصنيع الجليكوجين مرة أخرى من حمض اللاكتيك
أظهر Lundsgrad أن حمض الكرياتينوفوسفوريك موجود في خلايا العضلات ، وعندما يتم تقليله ، ينقسم إلى كرياتين وحمض الفوسفوريك (وفقًا لـ
بالادين) ، والذي يتحد مع الهكسوز (الجلوكوز). يتم أيضًا تكسير حمض الأدينوزينوفوسفوريك الموجود في العضلات لتشكيل الأدينوزين وحمض الفوسفوريك ، والذي ، عند دمجه مع الهكسوز (الجلوكوز) ، يعزز تكوين حمض اللاكتيك (إمبدين وزيمرمان).
لحم حيوان مذبوح حديثًا (لحم طازج) ذو قوام كثيف ، بدون رائحة محددة لطيفة واضحة ، عند طهيه يعطي مرقًا غير عطري عكرًا وليس له صفات طعم عالية. علاوة على ذلك ، في الساعات الأولى بعد ذبح الحيوان ، يتجمد اللحم ويصبح قاسيًا.
بعد 24-72 ساعة من ذبح الحيوان (اعتمادًا على درجة حرارة البيئة والتهوية وعوامل أخرى) ، يكتسب اللحم مؤشرات جودة جديدة: يختفي صلابته ، ويكتسب عصارة ورائحة لطيفة معينة ، ويتشكل فيلم كثيف على سطح الذبيحة (قشرة التجفيف) ، مع الطهي يعطي مرقًا معطرًا شفافًا ، يصبح طريًا ، إلخ.
تسمى العمليات والتغييرات التي تحدث في اللحوم ، ونتيجة لذلك تحصل على مؤشرات الجودة المرغوبة ، باسم نضج اللحوم.
نضج اللحم هو مزيج من العمليات الكيميائية الحيوية المعقدة في الأنسجة العضلية والتغيرات في التركيب الفيزيائي الغرواني للبروتين ، والتي تحدث تحت تأثير الإنزيمات الخاصة به.
يمكن تقسيم العمليات التي تحدث في الأنسجة العضلية بعد ذبح الحيوان إلى المراحل الثلاث التالية: دقة ما بعد الذبح ، والنضج ، والانحلال الذاتي.
تتطور الصلابة بعد الذبح في الذبيحة في الساعات الأولى بعد ذبح الحيوان ، وفي نفس الوقت تصبح العضلات مرنة وقصيرة قليلاً ، وهذا يزيد بشكل كبير من صلابة ومقاومة الجرح.
قدرة هذا اللحم على الانتفاخ منخفضة للغاية. عند درجة حرارة 15-20 درجة مئوية ، يحدث قسوة الموت الكامل بعد 3-5 ساعات من ذبح الحيوان ، وعند درجة حرارة 0-2 درجة مئوية - بعد 18-20 ساعة.
تترافق عملية تيبس الجثة بعد الذبح مع زيادة طفيفة في درجة حرارة الذبيحة نتيجة إطلاق الحرارة ، والتي تتكون من تفاعلات كيميائية تحدث في الأنسجة. ترجع صرامة الأنسجة العضلية ، التي لوحظت في الساعات والأيام الأولى بعد ذبح الحيوانات ، إلى تكوين مركب أكتوميوسين غير قابل للذوبان من بروتينات الأكتين والميوسين. الشرط الأساسي لتكوينه هو عدم وجود حمض الأدينوزين ثلاثي الفوسفوريك (ATP) ، والبيئة الحمضية للحوم وتراكم حمض اللاكتيك فيه. التغيرات البيوكيميائية في اللحوم تخلق هذه الظروف.
انخفاض واختفاء الـ ATP بشكل كامل مرتبط بانهياره نتيجة التأثير الأنزيمي للميوسين. يؤدي تحلل الـ ATP إلى ثنائي فسفوريك الأدينوزين (ADP ، أحادي الفوسفوريك الأدينوزين (AMP) وأحماض الفوسفوريك في حد ذاته إلى ظهور بيئة حمضية علاوة على ذلك ، بالفعل في هذه المرحلة ، يبدأ تفكك الجليكوجين العضلي ، مما يؤدي إلى تراكم حمض اللاكتيك ، مما يساهم أيضًا في تكوين بيئة حمضية فيه.
البيئة الحمضية ، وهي ظاهرة طبيعية لانهيار ATP وبداية عملية لا رجعة فيها لتحلل السكر (انهيار الجليكوجين العضلي) ، تزيد من تصلب العضلات. لوحظ أن عضلات الحيوانات التي ماتت أثناء التشنجات تصلب بشكل أسرع. تتميز الصرامة الصارمة دون تراكم حمض اللاكتيك بضعف توتر العضلات وحل سريع للعملية.
ومع ذلك ، قبل وقت طويل من نهاية مرحلة الصرامة ، يطور اللحم عمليات مرتبطة بمراحل نضجه وانحلاله الذاتي.
هناك عمليتان تؤديان إليهما - الانهيار المكثف للجليكوجين العضلي ، مما يؤدي إلى تحول حاد في قيمة الأس الهيدروجيني للحم إلى الجانب الحمضي ، بالإضافة إلى بعض التغييرات في التركيب الكيميائي والبنية الفيزيائية الغروية للبروتينات.
بسبب حقيقة أن عضلات اللحم لا تتلقى الأكسجين وتثبط عمليات الأكسدة فيها ، تتراكم فائض حمض اللاكتيك وحمض الفوسفوريك في اللحم. لذلك ، على سبيل المثال ، مع إجهاد عضلات الجسم (خلال حياته) ، يتم الوصول إلى 0.25٪ كحد أقصى من حمض اللاكتيك ، ومع صلابة ما بعد الوفاة فإنه يتراكم حتى 0.82٪. يتغير التفاعل النشط للوسط (الأس الهيدروجيني) في هذه الحالة من 7.26 إلى 6.02. من تراكم حمض اللاكتيك ، يحدث تقلص سريع (تصلب) للعضلات ، مصحوبًا بتخثر البروتين (Saxl). في هذه الحالة ، يفقد الأتوميوسين قابليته للذوبان ، وتستقر البروتينات ، ويترسب الكالسيوم من الغرويات البروتينية ويذهب إلى المحلول (عصير اللحم). بسبب المحتوى الزائد لحمض اللاكتيك ، يحدث أولاً تورم في المادة الغروية متباينة الخواص (القرص الداكن) لألياف العضلات (يترافق مع تقصير تصلب العضلات) ؛ ثم ، مع زيادة تركيز حمض اللاكتيك وتخثر البروتين ، تنعم هذه المادة. تفقد البروتينات المتخثرة خصائصها الغروية ، وتصبح غير قادرة على ربط (حبس) الماء ، وإلى حد ما تفقد وسطها المشتت (الماء): بدلاً من التورم الأولي ، تتقلص الغرويات الخلوية (تتقلص) وتصبح العضلات ناعمة (دقة الصلابة ).
نتيجة لتراكم اللاكتيك والفوسفوريك والأحماض الأخرى في اللحوم ، يزداد تركيز أيونات الهيدروجين ، ونتيجة لذلك ، في نهاية اليوم ، ينخفض الرقم الهيدروجيني إلى 5.8-5.7 (وحتى أقل).
في البيئة الحمضية ، يؤدي انهيار ATP و ADP و AMP وحمض الفوسفوريك إلى تراكم جزئي للفوسفور غير العضوي. تعتبر البيئة الحمضية الحادة ووجود الفوسفور غير العضوي سببًا في تفكك مركب الأكتوميوسين إلى الأكتين والميوسين. تفكك هذا المركب يزيل ظاهرة تصلب وصلابة اللحم. وبالتالي ، لا يمكن فصل مرحلة الصرامة عن المراحل الأخرى ويجب اعتبارها إحدى المراحل في عملية نضج اللحوم.
يمكن تمثيل مخطط التغيرات البيوكيميائية في عملية نضج اللحوم على النحو التالي.
تعمل البيئة الحمضية نفسها بشكل مضاد للجراثيم وحتى مبيد للجراثيم ، وبالتالي ، عندما يتحول الأس الهيدروجيني إلى الجانب الحمضي ، يتم إنشاء ظروف غير مواتية لتطوير الكائنات الحية الدقيقة في اللحوم.
أخيرًا ، تؤدي البيئة الحمضية إلى بعض التغييرات في التركيب الكيميائي والبنية الفيزيائية الغروانية للبروتينات. يغير نفاذية أغشية العضلات ودرجة تشتت البروتين. تتفاعل الأحماض مع بروتينات الكالسيوم وينفصل الكالسيوم عن البروتينات.
يؤدي انتقال الكالسيوم إلى المستخلص إلى انخفاض تشتت البروتينات ، مما يؤدي إلى فقدان جزء من الماء المرتبط بالهيدرات. لذلك ، يمكن فصل عصير اللحوم جزئيًا عن اللحوم الناضجة بالطرد المركزي.
الماء المرتبط بالهيدرات المنطلق ، وعمل الإنزيمات المحللة للبروتين والبيئة الحمضية تخلق ظروفًا لتخفيف غمد الليف العضلي ، وقبل كل شيء ، تخفيف وانتفاخ الكولاجين. هذا يساهم بشكل كبير في تغيير قوام اللحم وعصارة أكثر وضوحًا. من الواضح ، مع تورم الكولاجين ، ثم إطلاق جزئي للرطوبة من سطح الذبيحة إلى البيئة ، يجب أن يترافق تكوين قشرة جافة على سطحها.
تحدد مرحلة النضج الخاصة بها إلى حد كبير شدة تدفق العمليات الفيزيائية الغروانية والتغيرات الهيكلية المجهرية في ألياف العضلات ، والتي تحدث في مرحلة التحلل الذاتي. يتم تقليل التحلل الذاتي أثناء نضج اللحوم بالمعنى الواسع للكلمة ولا يرتبط فقط بتفكك البروتينات ، ولكن أيضًا بعملية تسوس أي أجزاء مكونة للخلايا. في هذا الصدد ، لا يمكن فصل العمليات التي تحدث في مرحلة النضج الخاص بها أو عزلها عن تلك التي تحدث أثناء التحلل الذاتي. ومع ذلك ، نتيجة لمجموعة من الأسباب (عمل الإنزيمات المحللة للبروتين ، وبيئة حمضية حادة ، ومنتجات التحلل الذاتي للمواد غير البروتينية ، وما إلى ذلك) ، يحدث الانهيار الذاتي لألياف العضلات إلى أجزاء منفصلة.
ينضج اللحم في غضون 24-72 ساعة عند درجة حرارة +4 درجات.
ومع ذلك ، ليس من الممكن دائمًا تحمل اللحوم عند درجة حرارة +4. في بعض الأحيان يكون من الضروري تخزينه في الظروف العادية (وليس في ظروف التبريد) عند درجة حرارة +6-8 درجة وما فوق ؛ في درجات الحرارة المرتفعة ، تستمر عمليات الصلابة الميتة ودقة العضلات بشكل أسرع. يعتمد معدل نضج اللحوم أيضًا على نوع وحالة الحيوان المذبوح ودهنه وعمره ؛ لكن هذه الأسئلة تتطلب مزيدًا من الملاحظة والدراسة.
أثناء نضج اللحم ، تنقسم بعض النيوكليدات
(المستخلصات النيتروجينية). تتشكل المواد المتطايرة والإسترات والألدهيدات ، مما يعطي نكهة للحوم. تظهر الأحماض الأدينية والإينوزينية ، والأدينين ، والزانثين ، وهيبوكسانثين ، والتي يعتمد عليها طعم اللحم. يتغير تفاعل بيئة اللحوم تجاه الحموضة (الرقم الهيدروجيني 6.2-
5.8). يساهم هذا في انتفاخ الغرويات البروتوبلازمي ، مما يؤدي إلى أن يصبح اللحم طريًا وطريًا ويصلح للطهي جيدًا.
يتم الحصول على اللحوم بهذه الجودة بعد 1-3 أيام من التخزين عند درجة حرارة من 4 إلى 12 درجة (حسب إمكانيات الشركات).
في المرحلة الأولى من هذه العملية ، يتم الكشف عن تجزئة ألياف العضلات الفردية مع الحفاظ على الألياف داخل الجسم. في الوقت نفسه ، يتم الحفاظ على بنية النوى والتخطيط العرضي والطولي في الأجزاء.
في المرحلة الثانية ، تخضع معظم ألياف العضلات للتجزئة.
كما هو الحال في المرحلة الأولى ، يستمر الحفاظ على بطانة الألياف ، وفي الأجزاء ، بنية النوى والتشوهات العرضية والطولية. أخيرًا ، في المرحلة الثالثة (مرحلة التحلل الذاتي العميق) ، يتم الكشف عن تفكك الأجزاء في اللييفات العضلية ، واللييفات العضلية إلى ساركوميرات.
يُنظر إلى الساركوميرات في المقاطع المجهرية المصنوعة من مثل هذه اللحوم على أنها كتلة حبيبية محاطة ببطانة داخلية.
تؤدي التغيرات المورفولوجية والبنية الدقيقة في الأنسجة أيضًا إلى تليين اللحم وتخفيفه أثناء نضجه ، مما يؤدي إلى اختراق العصارات الهضمية بحرية أكبر في الساركوبلازم ، مما يحسن قابليتها للهضم. وتجدر الإشارة إلى أن بروتينات النسيج الضام لا تخضع تقريبًا لعمليات تحلل البروتين أثناء نضوج اللحم. لذلك ، في ظل ظروف النضج المتساوية ، فإن حنان قطع اللحم المختلفة من نفس الحيوان ، وكذلك القطع المتطابقة من الحيوانات المختلفة ، ليست هي نفسها ؛ حنان اللحوم التي تحتوي على الكثير من الأنسجة الضامة صغيرة ، ولحوم الحيوانات الصغيرة أكثر رقة من اللحوم القديمة.
نتيجة لمجموعة من التحولات ذاتية التحلل لمكونات مختلفة من اللحوم ، أثناء نضوجها ، تتشكل وتتراكم المواد التي تحدد رائحة وطعم اللحم الناضج. يتم إعطاء طعم ورائحة معينة للحوم الناضجة عن طريق المواد الاستخراجية المحتوية على النيتروجين - هيبوكسانثين ، والكرياتين ، والكرياتينين ، التي تشكلت أثناء تفكك ATP ، وكذلك تراكم الأحماض الأمينية الحرة (حمض الجلوتاميك ، والأرجينين ، والثريونين ، والفينيل ألانين ، وما إلى ذلك) . على ما يبدو ، تشارك أحماض البيروفيك واللاكتيك في تكوين باقة من الذوق والرائحة.
اقترح I. A. Smorodintsev أن الطعم والرائحة يعتمدان على تراكم المواد القابلة للذوبان والمتطايرة بسهولة مثل الإسترات والألدهيدات والكيتونات في اللحوم الناضجة. بعد ذلك ، أظهر عدد من الدراسات أن الخصائص العطرية للحوم الناضجة تتحسن حيث تتراكم الكمية الإجمالية للمواد المختزلة المتطايرة فيها. في الوقت الحاضر ، باستخدام كروماتوغرافيا الغاز وتحليل طيف الكتلة ، ثبت أن المركبات التي تسبب رائحة اللحوم المسلوقة تشمل الأسيتالديهيد ، والأسيتون ، وميثيل إيثيل كيتون ، والميثانول ، وميثيل مركابتان ، وثنائي ميثيل كبريتيد ، وإيثيل مركابتان ، إلخ.
مع زيادة درجة الحرارة (حتى 30 درجة مئوية) ، وكذلك مع التعرض المطول للحوم (أكثر من 20-26 يومًا) في درجات حرارة منخفضة موجبة ، فإن عملية النضج الأنزيمي تصبح عميقة لدرجة أن كمية منتجات تكسير البروتين في الشكل من الببتيدات الصغيرة والأحماض الأمينية الحرة. في هذه المرحلة ، يكتسب اللحم اللون البني ، وتزداد كمية نيتروجين الأمونيا والأمونيا فيه ، ويحدث تحلل مائي ملحوظ للدهون ، مما يقلل بشكل كبير من صفاته التجارية والغذائية.
تختلف العمليات الكيميائية الحيوية التي تحدث أثناء النضج في لحوم الحيوانات المريضة عن العمليات الكيميائية الحيوية في لحوم الحيوانات السليمة.
مع الحمى والإرهاق ، تزداد عملية الطاقة في الجسم.
يتم تحسين عمليات الأكسدة في الأنسجة. يتميز التغيير في التمثيل الغذائي للكربوهيدرات أثناء المرض والإرهاق بانخفاض سريع في الجليكوجين في العضلات. لذلك ، تقريبًا مع أي عملية مرضية في جسم الحيوان ، يتم تقليل محتوى الجليكوجين في العضلات. نظرًا لوجود نسبة أقل من الجليكوجين في لحوم الحيوانات المريضة مقارنة بلحوم الحيوانات السليمة ، فإن كمية منتجات تكسير الجليكوجين (الجلوكوز وحمض اللاكتيك وما إلى ذلك) في لحوم الحيوانات المريضة تعد أيضًا ضئيلة.
بالإضافة إلى ذلك ، أثناء الأمراض الشديدة ، حتى أثناء حياة الحيوان ، تتراكم المنتجات الوسيطة والنهائية لعملية التمثيل الغذائي للبروتين في عضلاته. في هذه الحالات ، بالفعل في الساعات الأولى بعد ذبح الحيوان ، توجد كمية متزايدة من نيتروجين الأمونيا والأمونيا في اللحوم.
يعتبر التراكم الطفيف للأحماض وزيادة محتوى البولي ببتيدات والأحماض الأمينية والأمونيا سببًا في انخفاض تركيز أيونات الهيدروجين أثناء نضوج لحوم الحيوانات المريضة. يؤثر هذا العامل على نشاط إنزيمات اللحوم. في معظم الحالات ، يكون تركيز أيونات الهيدروجين ، الذي ينشأ نتيجة لنضوج لحوم الحيوانات المريضة ، أكثر ملاءمة لعمل الببتيدات والبروتياز.
نتيجة لذلك ، يعتبر تراكم المواد النيتروجينية المستخرجة في لحوم الحيوانات المريضة وعدم وجود تحول حاد في قيمة الأس الهيدروجيني إلى الجانب الحمضي ظروفًا مواتية لتطور الكائنات الحية الدقيقة.
تؤثر التغييرات التي تحدث في لحوم الحيوانات المريضة أيضًا على طبيعة التركيب الفيزيائي الغرواني للحوم بطريقة مختلفة. تسبب الحموضة الأقل ترسيبًا طفيفًا لأملاح الكالسيوم ، والتي بدورها تسبب تغيرًا طفيفًا في درجة تشتت البروتينات والتغيرات الأخرى المميزة لها أثناء النضج الطبيعي للحوم. إن درجة الحموضة المرتفعة نسبيًا ، وتراكم منتجات تحلل البروتين والظروف المواتية لتطوير الكائنات الحية الدقيقة ، تحدد مسبقًا المقاومة المنخفضة للحوم الحيوانات المريضة أثناء التخزين. العلامات المذكورة هي خصائص لحم كل حيوان مصاب بمرض خطير ؛ هم سبب التوحيد المعروف في التغيير في المعلمات الفيزيائية والكيميائية للحوم التي تم الحصول عليها من الحيوانات المقتولة أثناء العملية المرضية ، بغض النظر عن طبيعة المرض. لا ينفي هذا الحكم تغييرات محددة في تكوين اللحوم في بعض الأمراض ، ولكنه يعطي سببًا للحديث عن الأنماط العامة لنضج اللحوم في الأمراض في جسم الحيوان.
المركبات ذات الوزن الجزيئي العالي. في مُجَمَّعكل شيء معروف البروتينات... ثابت شكله و المواد الكيميائية مُجَمَّععلى الرغم من استمرارهم ...
المواد الكيميائية مُجَمَّعوالخصائص الفيزيائية للحيوانات المنوية
تقرير >> طب ، صحةالمواد الكيميائية مُجَمَّعوالخواص الفيزيائية للحيوانات المنوية الحيوانات المنوية هي مزيج من ... ، أ) ، الكلية والعناصر الدقيقة. المواد الكيميائية مُجَمَّعالحيوانات المنوية: 1) ماء - 75٪ 2) مادة جافة - 25٪: - السناجب- 85٪ - دهون ...
الأحماض الأمينية - المكونات الهيكلية البروتينات.أو البروتينات(البروتوس اليونانية - بارزة) ، هي بوليمرات غير متجانسة بيولوجية ، ومونومراتها أحماض أمينية.
الأحماض الأمينية عبارة عن مركبات عضوية ذات وزن جزيئي منخفض تحتوي على مجموعات كربوكسيل (-COOH) ومجموعات أمينية (-NH 2) مرتبطة بذرة الكربون نفسها. سلسلة جانبية متصلة بذرة الكربون - جذري يعطي كل حمض أميني خصائص معينة. الصيغة العامة للأحماض الأمينية هي:

تحتوي معظم الأحماض الأمينية على مجموعة كربوكسيل واحدة ومجموعة أمينية واحدة ؛ تسمى هذه الأحماض الأمينية محايد.ومع ذلك ، هناك أيضا الأحماض الأمينية الأساسية- مع أكثر من مجموعة أمينية ، وأيضًا أحماض أمينية حمضية- مع أكثر من مجموعة كربوكسيل.
من المعروف أن حوالي 200 من الأحماض الأمينية تحدث في الكائنات الحية ، ولكن 20 منها فقط هي جزء من البروتينات. هذه هي ما يسمى ب أساسي،أو تشكيل البروتين(بروتيني) ، أحماض أمينية.
اعتمادًا على نوع الجذر ، يتم تقسيم الأحماض الأمينية الأساسية إلى ثلاث مجموعات: 1) غير القطبية (ألانين ، ميثيونين ، فالين ، برو لاين ، ليسين ، إيزولوسين ، تريبتوفان ، فينيل ألانين) ؛ 2) قطبي غير مشحون (أسباراجين ، جلوتامين ، سيرين ، جلايسين ، تيروسين ، ثريونين ، سيستين) ؛ 3) الشحنة القطبية (أرجينين ، هيستيدين ، ليسين - إيجابي ؛ أحماض الأسبارتيك والغلوتاميك - سلبية).
يمكن أن تكون السلاسل الجانبية للأحماض الأمينية (الجذرية) كارهة للماء أو محبة للماء ، مما يعطي البروتينات الخصائص المقابلة التي تظهر أثناء تكوين هياكل البروتين الثانوية والثالثية والرباعية.
في النباتات الكليتم تصنيع الأحماض الأمينية الأساسية من المنتجات الأولية لعملية التمثيل الضوئي. لا يستطيع الإنسان والحيوان تخليق عدد من الأحماض الأمينية البروتينية ويجب أن يحصلوا عليها جاهزة مع الطعام. تسمى هذه الأحماض الأمينية لا يمكن الاستغناء عنه. إلىوهي تشمل ليسين ، فالين ، ليسين ، إيزولوسين ، ثريونين ، فينيل ألانين ، تريبتوفان ، ميثيونين ؛ وكذلك الأرجينين والهيستيدين - لا غنى عنه للأطفال ،
في المحلول ، يمكن أن تعمل الأحماض الأمينية كأحماض وقواعد ، أي أنها مركبات مذبذبة. مجموعة الكربوكسيل -COOH قادرة على التبرع ببروتون ، يعمل كحامض ، ومجموعة الأمين - NH2 - لتقبل البروتون ، وبالتالي تظهر خصائص القاعدة.
الببتيدات. يمكن أن تتفاعل المجموعة الأمينية لأحد الأحماض الأمينية مع مجموعة الكربوكسيل لحمض أميني آخر.
الجزيء الناتج هو ثنائي الببتيد ، وتسمى الرابطة -CO-NH- رابطة الببتيد:

في أحد طرفي جزيء ثنائي الببتيد توجد مجموعة أمينية حرة ، وفي الطرف الآخر توجد مجموعة كربوكسيل حرة. نتيجة لذلك ، يمكن أن يربط ثنائي الببتيد أحماض أمينية أخرى بنفسه ، مكونًا أوليغوبيبتيد. إذا تم توصيل العديد من الأحماض الأمينية (أكثر من عشرة) بهذه الطريقة ، فسيظهر ذلك بولي ببتيد.
تلعب الببتيدات دورًا مهمًا في الجسم. العديد من oligo- و polypeptides عبارة عن هرمونات ومضادات حيوية وسموم.
تشمل Oligopeptides الأوكسيتوسين ، والفازوبريسين ، والثيروتروبين ، وكذلك البراديكينين (ببتيد الألم) وبعض المواد الأفيونية ("الأدوية الطبيعية" للشخص) التي تؤدي وظيفة تخفيف الآلام. يؤدي تناول العقاقير إلى تدمير نظام الأفيون في الجسم ، لذا فإن مدمن المخدرات بدون جرعة من المخدرات يعاني من ألم شديد - "الانسحاب" ، والذي يتم التخلص منه عادةً بواسطة المواد الأفيونية. تحتوي Oligopeptides أيضًا على بعض المضادات الحيوية (على سبيل المثال ، gramicidin S).
العديد من الهرمونات (الأنسولين ، الهرمون الموجه لقشر الكظر ، وما إلى ذلك) ، والمضادات الحيوية (مثل الجراميسيدين أ) ، والسموم (مثل ذيفان الخناق) هي عديد الببتيدات.
البروتينات عبارة عن عديد ببتيدات ، يتكون جزيء منها من خمسين إلى عدة آلاف من الأحماض الأمينية ذات الوزن الجزيئي النسبي الذي يزيد عن 10000.
هيكل البروتينات. يتميز كل بروتين في بيئة معينة ببنية مكانية خاصة. عند توصيف الهيكل المكاني (ثلاثي الأبعاد) ، يتم تمييز أربعة مستويات من تنظيم جزيئات البروتين (الشكل 1.1).
|
تقع - glu - tre - ala - ala - ala - lis - fen - glu - arg - gln - gis - met - asp - ser - |
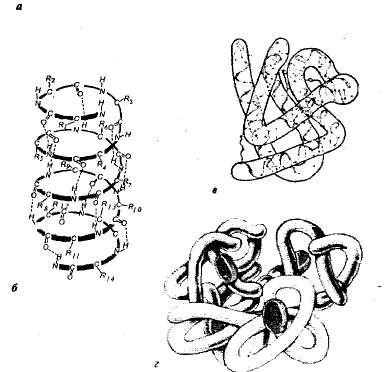
أرز. 1.1مستويات التنظيم الهيكلي للبروتين: أ — التركيب الأساسي هو تسلسل الأحماض الأمينية لبروتين الريبونوكليز (124 وحدة من الأحماض الأمينية) ؛ ب — الهيكل الثانوي — سلسلة البولي ببتيد ملتوية على شكل حلزوني ؛ في— التركيب الثالث لبروتين الميوجلوبين. جي — التركيب الرباعي للهيموجلوبين.
الهيكل الأساسي- تسلسل الأحماض الأمينية في سلسلة بولي ببتيد. هذا الهيكل خاص بكل بروتين ويتم تحديده من خلال المعلومات الجينية ، أي أنه يعتمد على تسلسل النيوكليوتيدات في منطقة جزيء الحمض النووي الذي يشفر هذا البروتين. تعتمد جميع خصائص ووظائف البروتينات على البنية الأولية. عادةً ما يستلزم استبدال حمض أميني واحد في تكوين جزيئات البروتين أو انتهاك الترتيب في ترتيبها تغييرًا في وظيفة البروتين.
بالنظر إلى أن البروتينات تحتوي على 20 نوعًا من الأحماض الأمينية ، فإن عدد المتغيرات لمجموعاتها في سلسلة البولي ببتيد غير محدود حقًا ، مما يوفر عددًا كبيرًا من أنواع البروتينات في الخلايا الحية. على سبيل المثال ، تم العثور على أكثر من 10000 بروتين مختلف في جسم الإنسان ، وكلها مبنية من نفس الأحماض الأمينية الأساسية العشرين.
في الخلايا الحية ، جزيئات البروتين أو أقسامها الفردية ليست سلسلة طويلة ، ولكنها ملتوية في دوامة تشبه الزنبرك الممتد (هذا ما يسمى الحلزون) ، أو مطوية في طبقة مطوية (طبقة p). هذه الحلزونات والطبقات p ثانوية بنية.ينتج عن تكوين روابط هيدروجينية داخل سلسلة عديد ببتيد واحدة (تكوين حلزوني) أو بين سلسلتين عديد الببتيد (طبقات مطوية).
يحتوي بروتين الكيراتين على تكوين حلزوني بالكامل. وهو بروتين بنيوي يتكون من الشعر والأظافر والمخالب والمنقار والريش والقرون. إنه جزء من الطبقة الخارجية من جلد الفقاريات.
في معظم البروتينات ، يتم طي المقاطع الحلزونية وغير الحلزونية لسلسلة البولي ببتيد في تشكيل ثلاثي الأبعاد لشكل كروي - كروي (سمة من سمات البروتينات الكروية). كرة لتكوين معين هو الهيكل الثالثسنجاب. يتم تثبيت هذا الهيكل عن طريق الروابط الأيونية والهيدروجين وثاني كبريتيد التساهمية (التي تتكون بين ذرات الكبريت التي هي جزء من السيستين والسيستين والميجيونين) ، وكذلك التفاعلات الكارهة للماء. التفاعلات الكارهة للماء هي الأهم في تكوين البنية الثلاثية ؛ في الوقت نفسه ، يتم طي البروتين بطريقة تخفي سلاسله الجانبية الكارهة للماء داخل الجزيء ، أي أنها محمية من ملامسة الماء ، والسلاسل الجانبية المحبة للماء ، على العكس من ذلك ، تتعرض للخارج.
تتكون العديد من البروتينات ذات البنية المعقدة بشكل خاص من عدة سلاسل متعددة الببتيد (وحدات فرعية) ، مكونة هيكل رباعيجزيء البروتين. مثل هذا الهيكل موجود ، على سبيل المثال ، في البروتين الكروي للهيموجلوبين. يتكون جزيءه من أربع وحدات فرعية منفصلة متعددة الببتيد (بروتومرات) تقع في البنية الثلاثية ، وجزء غير بروتيني - الهيم.
فقط في مثل هذا الهيكل يكون الهيموجلوبين قادرًا على أداء وظيفة النقل الخاصة به.
تحت تأثير العوامل الكيميائية والفيزيائية المختلفة (العلاج بالكحول والأسيتون والأحماض والقلويات ودرجة الحرارة المرتفعة والإشعاع والضغط العالي وما إلى ذلك) ، تتغير الهياكل الثانوية والثالثية والرباعية للبروتين بسبب تكسر الهيدروجين و الرابطة الأيونية. تسمى عملية تعطيل البنية الأصلية (الطبيعية) للبروتين تمسخ.في هذه الحالة ، لوحظ انخفاض في قابلية ذوبان البروتين ، وتغير في شكل وحجم الجزيئات ، وفقدان النشاط الأنزيمي ، وما إلى ذلك.يمكن أن تكون عملية التمسخ كاملة أو جزئية. في بعض الحالات ، يكون الانتقال إلى الظروف البيئية الطبيعية مصحوبًا باستعادة تلقائية للبنية الطبيعية للبروتين. تسمى هذه العملية إعادة التشبع.
بروتينات بسيطة ومعقدة. وفقًا للتركيب الكيميائي ، تنقسم البروتينات إلى بسيطة ومعقدة. اغفر ليتتكون البروتينات من الأحماض الأمينية فقط ، صعبة- البروتينات التي تحتوي على جزء بروتيني وجزء غير بروتيني (اصطناعي) ؛ يمكن تشكيل مجموعة صناعية من أيونات المعادن ، وبقايا حمض الفوسفوريك ، والكربوهيدرات ، والدهون ، وما إلى ذلك. البروتينات البسيطة هي ألبومين مصل الدم ، والفيبرين ، وبعض الإنزيمات (التربسين) ، إلخ. وتشمل البروتينات المعقدة جميع البروتينات البروتينية والبروتينات السكرية ؛ البروتينات المعقدة هي ، على سبيل المثال ، الغلوبولين المناعي (الأجسام المضادة) ، الهيموغلوبين ، معظم الإنزيمات ، إلخ.
وظائف البروتين.
- الهيكلي.البروتينات هي جزء من أغشية الخلايا ومصفوفة عضيات الخلية. تتكون جدران الأوعية الدموية والغضاريف والأوتار والشعر والأظافر والمخالب في الحيوانات العليا بشكل أساسي من البروتينات.
- حفاز (إنزيمى).تحفز إنزيمات البروتين جميع التفاعلات الكيميائية في الجسم. أنها توفر انهيار العناصر الغذائية في الجهاز الهضمي ، وتثبيت الكربون أثناء التمثيل الضوئي ، وما إلى ذلك.
- المواصلات.بعض البروتينات قادرة على الالتصاق وتحمل مواد مختلفة. ينقل الألبومينات في الدم الأحماض الدهنية ، والجلوبيولين - أيونات المعادن والهرمونات ، والهيموجلوبين - الأكسجين وثاني أكسيد الكربون. تشارك جزيئات البروتين التي يتكون منها غشاء البلازما في نقل المواد إلى الخلية.
- محمي.يتم تنفيذه بواسطة الغلوبولين المناعي (الأجسام المضادة) في الدم ، والتي توفر الدفاع المناعي للجسم. يشارك الفيبرينوجين والثرومبين في تخثر الدم ومنع النزيف.
- منقبض.بسبب انزلاق اللييفات الأولية للأكتين والميوسين بالنسبة لبعضهما البعض ، يحدث تقلص العضلات ، وكذلك تقلصات داخل الخلايا غير العضلية. ترتبط حركة الأهداب والسوط بانزلاق الأنابيب الدقيقة بالنسبة لبعضها البعض ، والتي تكون ذات طبيعة بروتينية.
- تنظيمية.العديد من الهرمونات قليلة الببتيدات أو فقيرة (على سبيل المثال ، الأنسولين ، الجلوكاجون [مضاد الأنسولين] ، الهرمون الموجه لقشر الكظر ، إلخ).
- مستقبلات.بعض البروتينات المضمنة في غشاء الخلية قادرة على تغيير هيكلها تحت تأثير البيئة الخارجية. هذه هي الطريقة التي يتم بها استقبال الإشارات من الخارج ونقل المعلومات إلى الخلية. سيكون على سبيل المثال الكروم النباتي—- بروتين حساس ينظم الاستجابة الضوئية للنباتات ، و أوبسين -مكون رودوبسين ،الصباغ الموجود في خلايا الشبكية.
- طاقة.يمكن أن تعمل البروتينات كمصدر للطاقة في الخلية (بعد تحللها المائي). عادة ، يتم استخدام البروتينات لتلبية احتياجات الطاقة في الحالات القصوى ، عندما يتم استنفاد احتياطيات الكربوهيدرات والدهون.
الانزيمات (الانزيمات). هذه بروتينات محددة موجودة في جميع الكائنات الحية وتلعب دور المحفزات البيولوجية.
تستمر التفاعلات الكيميائية في الخلية الحية عند درجة حرارة معينة وضغط طبيعي والحموضة المقابلة للوسط. في ظل هذه الظروف ، فإن تفاعلات تخليق المواد أو تحللها سوف تستمر ببطء شديد في الخلية إذا لم تتعرض للإنزيمات. تسرع الإنزيمات التفاعل دون تغيير نتيجته الإجمالية عن طريق التقليل طاقة التفعيل،أي ، في وجودها ، مطلوب طاقة أقل بكثير لنقل التفاعل إلى الجزيئات التي تتفاعل ، أو يستمر التفاعل على طول مسار مختلف بحاجز طاقة أقل.
يتم تنفيذ جميع العمليات في الكائن الحي بشكل مباشر أو غير مباشر بمشاركة الإنزيمات. على سبيل المثال ، تحت تأثيرها ، يتم تقسيم المكونات المكونة للغذاء (البروتينات ، والكربوهيدرات ، والدهون ، وما إلى ذلك) إلى مركبات أبسط ، ثم يتم تصنيع جزيئات كبيرة جديدة مميزة من هذا النوع منها. لذلك ، غالبًا ما تؤدي انتهاكات تكوين ونشاط الإنزيمات إلى حدوث أمراض خطيرة.
عن طريق التنظيم المكاني ، تتكون الإنزيمات من عدة سلاسل بولي وببتيد وعادة ما يكون لها هيكل رباعي. بالإضافة إلى ذلك ، يمكن أن تشمل الإنزيمات أيضًا هياكل غير بروتينية. يرتدي جزء البروتين اسم apoenzyme ،وغير البروتين العامل المساعد(إذا كانت هذه كاتيونات أو أنيونات من مواد غير عضوية ، على سبيل المثال ، Zn 2 Mn 2+ ، إلخ) أو أنزيم (أنزيم)(إذا كانت مادة عضوية منخفضة الوزن الجزيئي).
إن طلائع أو مكونات العديد من الإنزيمات المساعدة هي الفيتامينات. لذلك ، حمض البانتوثنيك هو جزء لا يتجزأ من أنزيم A ، وحمض النيكوتين (فيتامين PP) هو مقدمة لـ NAD و NADP ، إلخ.
يخضع التحفيز الإنزيمي لنفس قوانين التحفيز غير الإنزيمي في الصناعة الكيميائية ، ولكن على عكس ذلك ، فإنه يتميز بخاصية غير عادية درجة عالية من الخصوصية(يحفز الإنزيم تفاعل واحد فقط أو يعمل على نوع واحد فقط من الروابط). وهذا يضمن التنظيم الدقيق لجميع العمليات الحيوية (التنفس ، والهضم ، والتمثيل الضوئي ، وما إلى ذلك) التي تحدث في الخلية والجسم. على سبيل المثال ، يحفز إنزيم اليورياز تكسير مادة واحدة فقط - اليوريا (H 2 N-CO-NH 2 + H 2 O -> - »2NH 3 + CO 2) ، دون أن يكون له تأثير تحفيزي على المركبات ذات الصلة بنيوياً.
لفهم آلية عمل الإنزيمات بخصوصية عالية ، إنه أمر شديد الأهمية نظرية المركز النشط مهمة.وفقا لها ، فيمركب كل واحدإنزيم هناك واحدموقع أو أكثر يحدث فيه التحفيز بسبب التلامس الوثيق (في نقاط كثيرة) بين جزيئات الإنزيم ومادة معينة (الركيزة). المركز النشط إما مجموعة وظيفية (على سبيل المثال ، مجموعة OH من السيرين) أو حمض أميني واحد. عادة ، من أجل العمل التحفيزي ، من الضروري الجمع بين عدة (في المتوسط من 3 إلى 12) من بقايا الأحماض الأمينية المرتبة بترتيب معين. يتكون المركز النشط أيضًا من أيونات معدنية مرتبطة بالإنزيم والفيتامينات والمركبات الأخرى غير البروتينية - الإنزيمات المساعدة أو العوامل المساعدة. علاوة على ذلك ، فإن الشكل والبنية الكيميائية للمركز النشط هي كذلك معفقط ركائز معينة يمكن أن ترتبط به بسبب تطابقها المثالي (التكامل أو التكامل) مع بعضها البعض. يتمثل دور بقايا الأحماض الأمينية المتبقية في جزيء إنزيم كبير في تزويد جزيئه بالشكل الكروي المناسب ، وهو أمر ضروري للتشغيل الفعال للمركز النشط. بالإضافة إلى ذلك ، ينشأ مجال كهربائي قوي حول جزيء إنزيم كبير. في مثل هذا المجال ، يصبح اتجاه جزيئات الركيزة واكتسابها لشكل غير متماثل ممكنًا. يؤدي هذا إلى إضعاف الروابط الكيميائية ، ويحدث التفاعل المحفز مع إنفاق مبدئي أقل للطاقة ، وبالتالي بمعدل أعلى بكثير. على سبيل المثال ، يمكن لجزيء واحد من إنزيم الكاتلاز أن يكسر أكثر من 5 ملايين جزيء من بيروكسيد الهيدروجين (H2 0 2) في دقيقة واحدة ، والذي يحدث عندما تتأكسد مركبات مختلفة في الجسم.
في بعض الإنزيمات ، في وجود الركيزة ، يخضع تكوين المركز النشط للتغييرات ، أي أن الإنزيم يوجه مجموعاته الوظيفية بطريقة توفر أكبر نشاط تحفيزي.
في المرحلة النهائية من التفاعل الكيميائي ، يتم فصل مركب الركيزة الإنزيمية مع تكوين منتجات نهائية وإنزيم حر. يمكن أن يقبل المركز النشط الصادر في هذه الحالة جزيئات الركيزة الجديدة.
معدل التفاعلات الأنزيميةيعتمد على العديد من العوامل: طبيعة وتركيز الإنزيم والركيزة ، ودرجة الحرارة ، والضغط ، وحموضة الوسط ، ووجود مثبطات ، وما إلى ذلك ، على سبيل المثال ، عند درجات حرارة قريبة من الصفر ، يتباطأ معدل التفاعلات الكيميائية الحيوية إلى الحد الأدنى . تستخدم هذه الخاصية على نطاق واسع في مختلف قطاعات الاقتصاد ، على وجه الخصوص في الزراعة والطب. على وجه الخصوص ، الحفظمن الأعضاء المختلفة (الكلى والقلب والطحال والكبد) قبل زرعها للمريض يحدث أثناء التبريد من أجل تقليل شدة التفاعلات الكيميائية الحيوية وإطالة عمر الأعضاء. التجميد السريع للمواد الغذائية يمنع نمو وتكاثر الكائنات الحية الدقيقة (البكتيريا ، الفطريات ، إلخ) وكذلك يثبط نشاط إنزيماتها الهضمية بحيث لا تعود قادرة على التسبب في تحلل المواد الغذائية.
مصدر : على ال. Lemeza L.V Kamlyuk N.D. ليسوف "دليل علم الأحياء للمتقدمين للجامعات"
السناجب- مركبات عضوية جزيئية عالية ، تتكون من بقايا الأحماض الأمينية ألفا.
في تكوين البروتينيشمل الكربون والهيدروجين والنيتروجين والأكسجين والكبريت. تشكل بعض البروتينات معقدات مع جزيئات أخرى تحتوي على الفوسفور والحديد والزنك والنحاس.
البروتينات لها وزن جزيئي كبير: زلال البيض - 36000 ، الهيموجلوبين - 152000 ، الميوسين - 500000. للمقارنة: الوزن الجزيئي للكحول هو 46 ، حمض الأسيتيك - 60 ، البنزين - 78.
تكوين الأحماض الأمينية للبروتينات
السناجب- البوليمرات غير الدورية ومونومراتها الأحماض الأمينية ألفا. عادةً ما يُطلق على 20 نوعًا من الأحماض الأمينية ألفا مونومرات البروتين ، على الرغم من وجود أكثر من 170 نوعًا منها في الخلايا والأنسجة.
اعتمادًا على إمكانية تصنيع الأحماض الأمينية في جسم الإنسان والحيوانات الأخرى ، هناك: الأحماض الأمينية غير الأساسية- يمكن تصنيعه الأحماض الأمينية الأساسية- لا يمكن تصنيعه. يجب تناول الأحماض الأمينية الأساسية مع الطعام. تصنع النباتات جميع أنواع الأحماض الأمينية.
اعتمادا على تكوين الأحماض الأمينية ، البروتينات هي: كاملة- تحتوي على مجموعة كاملة من الأحماض الأمينية ؛ معيب- بعض الأحماض الأمينية غائبة في تركيبها. إذا كانت البروتينات تتكون من أحماض أمينية فقط ، فإنها تسمى بسيط. إذا كانت البروتينات تحتوي ، بالإضافة إلى الأحماض الأمينية ، على مكون غير أحماض أمينية (مجموعة اصطناعية) ، فإنها تسمى مركب. يمكن تمثيل المجموعة التعويضية بالمعادن (البروتينات المعدنية) والكربوهيدرات (البروتينات السكرية) والدهون (البروتينات الدهنية) والأحماض النووية (البروتينات النووية).
الجميع تحتوي الأحماض الأمينية: 1) مجموعة كربوكسيل (-COOH) ، 2) مجموعة أمينية (-NH 2) ، 3) مجموعة جذرية أو مجموعة R (باقي الجزيء). يختلف هيكل الراديكالي في أنواع مختلفة من الأحماض الأمينية. اعتمادًا على عدد المجموعات الأمينية ومجموعات الكربوكسيل التي تشكل الأحماض الأمينية ، هناك: أحماض أمينية محايدةوجود مجموعة كربوكسيل واحدة ومجموعة أمينية واحدة ؛ الأحماض الأمينية الأساسيةوجود أكثر من مجموعة أمينية واحدة ؛ أحماض أمينية حمضيةوجود أكثر من مجموعة كربوكسيل واحدة.
الأحماض الأمينية مركبات مذبذبة، حيث يمكن أن تعمل في المحلول كأحماض وقواعد. في المحاليل المائية ، توجد الأحماض الأمينية في أشكال أيونية مختلفة.
السندات الببتيد
الببتيدات- مواد عضوية تتكون من بقايا الأحماض الأمينية المتصلة بواسطة رابطة ببتيدية.
يحدث تكوين الببتيدات نتيجة تفاعل تكثيف الأحماض الأمينية. عندما تتفاعل المجموعة الأمينية لأحد الأحماض الأمينية مع مجموعة الكربوكسيل الأخرى ، تنشأ رابطة تساهمية بين النيتروجين والكربون ، والتي تسمى الببتيد. اعتمادًا على عدد بقايا الأحماض الأمينية التي تتكون منها الببتيد ، هناك ثنائي الببتيدات ، ثلاثي الببتيدات ، رباعي الببتيداتإلخ. يمكن تكرار تكوين رابطة الببتيد عدة مرات. هذا يؤدي إلى التكوين بولي ببتيدات. في أحد طرفي الببتيد توجد مجموعة أمينية حرة (تسمى الطرف N) ، وفي الطرف الآخر توجد مجموعة كربوكسيل حرة (تسمى الطرف C).

التنظيم المكاني لجزيئات البروتين
يعتمد أداء بعض الوظائف المحددة للبروتينات على التكوين المكاني لجزيئاتها ، بالإضافة إلى أنه من غير المواتي بقوة أن تحتفظ الخلية بالبروتينات في شكل موسع ، في شكل سلسلة ، وبالتالي ، يتم طي سلاسل البولي ببتيد ، والحصول على بنية أو شكل معين ثلاثي الأبعاد. تخصيص 4 مستويات التنظيم المكاني للبروتينات.
الهيكل الأساسي للبروتين- تسلسل بقايا الأحماض الأمينية في سلسلة البولي ببتيد التي تشكل جزيء البروتين. الرابطة بين الأحماض الأمينية هي الببتيد.

إذا كان جزيء البروتين يتكون من 10 بقايا من الأحماض الأمينية فقط ، فإن عدد المتغيرات الممكنة نظريًا لجزيئات البروتين التي تختلف في ترتيب تناوب الأحماض الأمينية هو 10 20. باستخدام 20 من الأحماض الأمينية ، يمكنك تكوين مجموعات أكثر تنوعًا منها. تم العثور على حوالي عشرة آلاف بروتين مختلف في جسم الإنسان ، والتي تختلف عن بعضها البعض وعن بروتينات الكائنات الحية الأخرى.
إنه الهيكل الأساسي لجزيء البروتين الذي يحدد خصائص جزيئات البروتين وتكوينه المكاني. يؤدي استبدال حمض أميني واحد فقط بآخر في سلسلة البولي ببتيد إلى تغيير خصائص ووظائف البروتين. على سبيل المثال ، يؤدي استبدال الحمض الأميني الجلوتامين السادس في الوحدة الفرعية β للهيموجلوبين بالفالين إلى حقيقة أن جزيء الهيموجلوبين ككل لا يمكنه أداء وظيفته الرئيسية - نقل الأكسجين ؛ في مثل هذه الحالات ، يصاب الشخص بمرض - فقر الدم المنجلي.
الهيكل الثانوي- أمر طي سلسلة البولي ببتيد في لولب (يشبه الزنبرك الممتد). يتم تقوية ملفات اللولب بواسطة روابط هيدروجينية بين مجموعات الكربوكسيل والمجموعات الأمينية. تشارك جميع مجموعات CO و NH تقريبًا في تكوين روابط هيدروجينية. إنها أضعف من الببتيد ، لكنها ، تتكرر عدة مرات ، تضفي الاستقرار والصلابة على هذا التكوين. على مستوى البنية الثانوية ، توجد بروتينات: فيبروين (حرير ، ويب) ، كيراتين (شعر ، أظافر) ، كولاجين (أوتار).



الهيكل الثالث- تعبئة سلاسل البولي ببتيد في كريات ناتجة عن حدوث روابط كيميائية (هيدروجين ، أيوني ، ثاني كبريتيد) وإنشاء تفاعلات كارهة للماء بين جذور بقايا الأحماض الأمينية. يتم لعب الدور الرئيسي في تكوين الهيكل الثالث من خلال التفاعلات المحبة للماء. في المحاليل المائية ، تميل الجذور الكارهة للماء إلى الاختباء من الماء ، وتتجمع داخل الكريات ، بينما تميل الجذور المحبة للماء إلى الظهور على سطح الجزيء نتيجة للترطيب (التفاعل مع ثنائيات أقطاب الماء). في بعض البروتينات ، يتم تثبيت البنية الثلاثية عن طريق روابط تساهمية ثنائي كبريتيد تتشكل بين ذرات الكبريت في بقايا السيستين. على مستوى البنية الثلاثية ، توجد إنزيمات وأجسام مضادة وبعض الهرمونات.

هيكل رباعيسمة من سمات البروتينات المعقدة ، والتي تتكون جزيئاتها من اثنين أو أكثر من الكريات. يتم الاحتفاظ بالوحدات الفرعية في الجزيء عن طريق التفاعلات الأيونية والطارئة للماء والكهرباء الساكنة. في بعض الأحيان ، أثناء تكوين هيكل رباعي ، تحدث روابط ثاني كبريتيد بين الوحدات الفرعية. البروتين الأكثر دراسة مع هيكل رباعي هو الهيموغلوبين. يتكون من وحدتين فرعيتين α (141 بقايا من الأحماض الأمينية) ووحدتين فرعيتين (146 من بقايا الأحماض الأمينية). ترتبط كل وحدة فرعية بجزيء هيم يحتوي على الحديد.
إذا انحرف التكوين المكاني للبروتينات عن الطبيعي لسبب ما ، فلن يتمكن البروتين من أداء وظائفه. على سبيل المثال ، سبب "مرض جنون البقر" (الاعتلال الدماغي الإسفنجي) هو التشكل غير الطبيعي للبريونات ، وهي البروتينات السطحية للخلايا العصبية.
خصائص البروتين
تكوين الأحماض الأمينية ، وهيكل جزيء البروتين تحدده الخصائص. تجمع البروتينات بين الخصائص الأساسية والحمضية التي تحددها جذور الأحماض الأمينية: كلما زادت الأحماض الأمينية الحمضية في البروتين ، زادت خصائصه الحمضية وضوحًا. القدرة على إعطاء وإرفاق H + تحديد خصائص عازلة للبروتينات؛ يعد الهيموجلوبين في كريات الدم الحمراء أحد أقوى المحاليل المنظمة ، والذي يحافظ على درجة حموضة الدم عند مستوى ثابت. توجد بروتينات قابلة للذوبان (الفيبرينوجين) ، وهناك بروتينات غير قابلة للذوبان تؤدي وظائف ميكانيكية (فيبروين ، كيراتين ، كولاجين). هناك بروتينات نشطة كيميائيًا (إنزيمات) ، وهناك بروتينات غير نشطة كيميائيًا ، ومقاومة للظروف البيئية المختلفة وغير مستقرة للغاية.
العوامل الخارجية (الحرارة ، الأشعة فوق البنفسجية ، المعادن الثقيلة وأملاحها ، تغيرات الأس الهيدروجيني ، الإشعاع ، الجفاف)
يمكن أن يسبب انتهاكًا للتنظيم الهيكلي لجزيء البروتين. تسمى عملية فقدان التشكل ثلاثي الأبعاد المتأصل في جزيء بروتين معين تمسخ. سبب التمسخ هو كسر الروابط التي تثبت بنية بروتينية معينة. في البداية ، تتمزق الروابط الأضعف ، وعندما تصبح الظروف أكثر صعوبة ، تكون أقوى. لذلك ، أولاً ، الرباعي ، ثم الهياكل الثانوية والثانوية تضيع. يؤدي التغيير في التكوين المكاني إلى تغيير في خصائص البروتين ، ونتيجة لذلك ، يجعل من المستحيل على البروتين أداء وظائفه البيولوجية. إذا لم يكن التمسخ مصحوبًا بتدمير الهيكل الأساسي ، فيمكن أن يكون كذلك تفريغ، في هذه الحالة ، يحدث الشفاء الذاتي لخاصية التشكل المميزة للبروتين. يخضع هذا التمسخ ، على سبيل المثال ، لبروتينات مستقبلات الغشاء. تسمى عملية استعادة بنية البروتين بعد التمسخ إعادة التشبع. إذا كانت استعادة التكوين المكاني للبروتين أمرًا مستحيلًا ، فسيتم استدعاء التمسخ لا رجعة فيه.
وظائف البروتينات
| دور | أمثلة وتفسيرات |
|---|---|
| بناء | تشارك البروتينات في تكوين الهياكل الخلوية وخارج الخلية: فهي جزء من أغشية الخلايا (البروتينات الدهنية والبروتينات السكرية) والشعر (الكيراتين) والأوتار (الكولاجين) ، إلخ. |
| المواصلات | يربط بروتين الهيموجلوبين في الدم الأكسجين وينقله من الرئتين إلى جميع الأنسجة والأعضاء ، وينتقل منها ثاني أكسيد الكربون إلى الرئتين ؛ يتضمن تكوين أغشية الخلايا بروتينات خاصة توفر نقلًا نشطًا وانتقائيًا صارمًا لبعض المواد والأيونات من الخلية إلى البيئة الخارجية والعكس صحيح. |
| تنظيمية | تشارك هرمونات البروتين في تنظيم عمليات التمثيل الغذائي. على سبيل المثال ، ينظم هرمون الأنسولين مستويات الجلوكوز في الدم ، ويعزز تخليق الجليكوجين ، ويزيد من تكوين الدهون من الكربوهيدرات. |
| محمي | استجابة لاختراق البروتينات الأجنبية أو الكائنات الحية الدقيقة (المستضدات) في الجسم ، يتم تكوين بروتينات خاصة - أجسام مضادة يمكنها ربطها وتحييدها. يساعد الفيبرين ، المكون من الفيبرينوجين ، على وقف النزيف. |
| محرك | توفر بروتينات الأكتين والميوسين الانقباضية تقلصًا للعضلات في الحيوانات متعددة الخلايا. |
| الإشارة | يتم تضمين جزيئات البروتينات في الغشاء السطحي للخلية ، وهي قادرة على تغيير هيكلها الثلاثي استجابة لتأثير العوامل البيئية ، وبالتالي تلقي إشارات من البيئة الخارجية ونقل الأوامر إلى الخلية. |
| احتياطي | في جسم الحيوانات ، لا يتم تخزين البروتينات ، كقاعدة عامة ، باستثناء زلال البيض وكازين الحليب. ولكن بفضل البروتينات في الجسم ، يمكن تخزين بعض المواد في احتياطي ، على سبيل المثال ، أثناء انهيار الهيموجلوبين ، لا يفرز الحديد من الجسم ، ولكن يتم تخزينه ، مكونًا مركبًا مع بروتين الفيريتين. |
| طاقة | مع تحلل 1 جرام من البروتين إلى المنتجات النهائية ، يتم تحرير 17.6 كيلو جول. أولاً ، تنقسم البروتينات إلى أحماض أمينية ، ثم إلى المنتجات النهائية - الماء وثاني أكسيد الكربون والأمونيا. ومع ذلك ، يتم استخدام البروتينات كمصدر للطاقة فقط عند استخدام مصادر أخرى (الكربوهيدرات والدهون). |
| المحفز | من أهم وظائف البروتينات. مزود بالبروتينات - الإنزيمات التي تسرع التفاعلات الكيميائية الحيوية التي تحدث في الخلايا. على سبيل المثال ، يحفز كربوكسيلاز الريبولوز ثنائي الفوسفات تثبيت ثاني أكسيد الكربون أثناء عملية التمثيل الضوئي. |
الانزيمات
الانزيمات، أو الانزيمات، هي فئة خاصة من البروتينات التي تعتبر محفزات بيولوجية. بفضل الإنزيمات ، تستمر التفاعلات الكيميائية الحيوية بسرعة هائلة. معدل التفاعلات الأنزيمية هو عشرات الآلاف من المرات (وأحيانًا الملايين) أعلى من معدل التفاعلات التي تنطوي على محفزات غير عضوية. تسمى المادة التي يعمل عليها الإنزيم المادة المتفاعلة.
الإنزيمات هي بروتينات كروية السمات الهيكليةيمكن تقسيم الإنزيمات إلى مجموعتين: بسيطة ومعقدة. إنزيمات بسيطةهي بروتينات بسيطة ، أي تتكون فقط من الأحماض الأمينية. إنزيمات معقدةهي بروتينات معقدة ، أي بالإضافة إلى الجزء البروتيني ، فهي تشمل مجموعة ذات طبيعة غير بروتينية - العامل المساعد. بالنسبة لبعض الإنزيمات ، تعمل الفيتامينات كعوامل مساعدة. في جزيء الإنزيم ، يتم عزل جزء خاص يسمى المركز النشط. مركز نشط- قسم صغير من الإنزيم (من ثلاثة إلى اثني عشر بقايا من الأحماض الأمينية) ، حيث يحدث ارتباط الركيزة أو الركائز مع تكوين مركب الركيزة الإنزيمية. عند الانتهاء من التفاعل ، يتحلل مركب الركيزة الإنزيمية إلى إنزيم ومنتج (منتجات) تفاعل. تحتوي بعض الإنزيمات (بخلاف الإنزيمات النشطة) مراكز allosteric- المواقع التي ترتبط بها منظمات معدل عمل الإنزيم ( الإنزيمات الخيفية).

تتميز تفاعلات التحفيز الإنزيمي بما يلي: 1) الكفاءة العالية ، 2) الانتقائية الصارمة واتجاه العمل ، 3) خصوصية الركيزة ، 4) التنظيم الدقيق والدقيق. يتم شرح خصوصية الركيزة والتفاعل لتفاعلات التحفيز الإنزيمي من خلال فرضيات E. Fischer (1890) و D. Koshland (1959).
إي فيشر (فرضية قفل المفاتيح)اقترح أن التكوينات المكانية للموقع النشط للإنزيم والركيزة يجب أن تتوافق تمامًا مع بعضها البعض. تتم مقارنة الركيزة بـ "المفتاح" ، والإنزيم - بـ "القفل".
كوشلاند (فرضية "قفاز اليد")اقترح أن المراسلات المكانية بين بنية الركيزة والمركز النشط للإنزيم يتم إنشاؤها فقط في لحظة تفاعلهم مع بعضهم البعض. تسمى هذه الفرضية أيضًا فرضية الملاءمة المستحثة.
يعتمد معدل التفاعلات الأنزيمية على: 1) درجة الحرارة ، 2) تركيز الإنزيم ، 3) تركيز الركيزة ، 4) درجة الحموضة. يجب التأكيد على أنه نظرًا لأن الإنزيمات عبارة عن بروتينات ، فإن نشاطها يكون أعلى في ظل الظروف الطبيعية من الناحية الفسيولوجية.
يمكن لمعظم الإنزيمات العمل فقط في درجات حرارة تتراوح بين 0 و 40 درجة مئوية. ضمن هذه الحدود ، يزداد معدل التفاعل بحوالي مرتين لكل ارتفاع في درجة الحرارة بمقدار 10 درجات مئوية. عند درجات حرارة أعلى من 40 درجة مئوية ، يخضع البروتين للتشوه ويقل نشاط الإنزيم. في درجات حرارة قريبة من التجمد ، يتم تعطيل الإنزيمات.
مع زيادة كمية الركيزة ، يزداد معدل التفاعل الإنزيمي حتى يصبح عدد جزيئات الركيزة مساوياً لعدد جزيئات الإنزيم. مع زيادة أخرى في كمية الركيزة ، لن يزداد المعدل ، لأن المواقع النشطة للإنزيم مشبعة. تؤدي الزيادة في تركيز الإنزيم إلى زيادة النشاط التحفيزي ، حيث يخضع عدد أكبر من جزيئات الركيزة للتحولات لكل وحدة زمنية.

لكل إنزيم ، توجد قيمة pH مثالية تظهر عندها أقصى نشاط (البيبسين - 2.0 ، الأميليز اللعابي - 6.8 ، الليباز البنكرياس - 9.0). عند ارتفاع أو انخفاض قيم الأس الهيدروجيني ، ينخفض نشاط الإنزيم. مع التحولات الحادة في الأس الهيدروجيني ، فإن الإنزيم يفسد.
يتم تنظيم سرعة الإنزيمات الخيفية من خلال المواد التي ترتبط بمراكز التباين. إذا كانت هذه المواد تسرع التفاعل ، يتم استدعاؤها المنشطاتإذا تباطأوا - مثبطات.
تصنيف الانزيم
وفقًا لنوع التحولات الكيميائية المحفزة ، تنقسم الإنزيمات إلى 6 فئات:
- أوكسيدوروكتاز(نقل ذرات الهيدروجين أو الأكسجين أو الإلكترون من مادة إلى أخرى - نازعة الهيدروجين) ،
- ناقل(نقل مجموعة ميثيل أو أسيل أو فوسفات أو أمينية من مادة إلى أخرى - ترانساميناز) ،
- هيدروليسات(تفاعلات التحلل المائي التي يتكون فيها منتجان من الركيزة - الأميليز والليباز) ،
- ليات(إضافة غير مائية إلى الركيزة أو إزالة مجموعة من الذرات منها ، في حين يمكن كسر روابط C-C و C-N و C-O و C-S - نزع الكربوكسيل) ،
- ايزوميراز(إعادة ترتيب الجزيئية - إيزوميراز) ،
- إنزيمات دمج الجزيئات(اتصال جزيئين نتيجة تكوين روابط C-C و C-N و C-O و C-S - synthetase).
وتنقسم الفئات بدورها إلى فئات فرعية وفئات فرعية. في التصنيف الدولي الحالي ، لكل إنزيم رمز محدد ، يتكون من أربعة أرقام مفصولة بنقاط. الرقم الأول هو الفئة ، والثاني هو الفئة الفرعية ، والثالث هو الفئة الفرعية ، والرابع هو الرقم التسلسلي للإنزيم في هذه الفئة الفرعية ، على سبيل المثال ، رمز أرجيناز هو 3.5.3.1.
اذهب إلى عدد المحاضرات 2"هيكل ووظائف الكربوهيدرات والدهون"
اذهب إلى محاضرات №4"هيكل ووظائف الأحماض النووية ATP"
إرسال عملك الجيد في قاعدة المعرفة أمر بسيط. استخدم النموذج أدناه
سيكون الطلاب وطلاب الدراسات العليا والعلماء الشباب الذين يستخدمون قاعدة المعرفة في دراساتهم وعملهم ممتنين جدًا لك.
نشر على http://www.allbest.ru
الفصل 1 مقدمة
أصبحت التقارير عن ثورة في علم الأحياء الآن مبتذلة إلى حد ما. كما أنه لا جدال في أن هذه التغييرات الثورية ارتبطت بتكوين مجموعة معقدة من العلوم عند تقاطع علم الأحياء والكيمياء ، من بينها البيولوجيا الجزيئية والكيمياء العضوية الحيوية احتلت واستمرت في احتلال موقع مركزي.
"البيولوجيا الجزيئية هي علم يهدف إلى فهم طبيعة ظواهر الحياة من خلال دراسة الكائنات والأنظمة البيولوجية على مستوى يقترب من المستوى الجزيئي ... المظاهر المميزة للحياة ... ترجع إلى بنية وخصائص وتفاعل جزيئات المواد المهمة بيولوجيا ، في المقام الأول البروتينات والأحماض النووية ”
"الكيمياء الحيوية العضوية هي علم يدرس المواد التي تكمن وراء عمليات الحياة ... الأشياء الرئيسية للكيمياء العضوية هي البوليمرات الحيوية (البروتينات والببتيدات والأحماض النووية والنيوكليوتيدات والدهون والسكريات المتعددة وما إلى ذلك).
من هذه المقارنة ، يتضح مدى أهمية دراسة البروتينات في تطوير علم الأحياء الحديث.
الكيمياء الحيوية البروتين
الفصل 2. تاريخ أبحاث البروتين
2.1 المراحل المبكرة في كيمياء البروتين
كان البروتين من بين أهداف البحث الكيميائي منذ 250 عامًا. في عام 1728 ، حصل العالم الإيطالي جاكوبو بارتولوميو بيكاري على أول تحضير بروتين ، وهو الغلوتين ، من دقيق القمح. أخضع الغلوتين للتقطير الجاف وتأكد من أن منتجات هذا التقطير قلوية. كان هذا أول دليل على وحدة طبيعة مواد الممالك النباتية والحيوانية. نشر نتائج عمله عام 1745 ، وكانت هذه أول ورقة بحثية عن بروتين.
في القرن الثامن عشر - أوائل القرن التاسع عشر ، تم وصف المواد البروتينية من أصل نباتي وحيواني بشكل متكرر. ومن سمات هذه الأوصاف تقارب هذه المواد ومقارنتها مع المواد غير العضوية.
من المهم أن نلاحظ أنه في ذلك الوقت ، حتى قبل ظهور التحليل الأولي ، كانت هناك فكرة أن البروتينات من مصادر مختلفة كانت مجموعة من المواد الفردية ذات الخصائص المتشابهة.
في عام 1810 ، حدد J. Gay-Lussac و L. Tenard لأول مرة التكوين الأولي للمواد البروتينية. في عام 1833 ، أثبت J. Gay-Lussac أن النيتروجين موجود بالضرورة في البروتينات ، وسرعان ما تبين أن محتوى النيتروجين في البروتينات المختلفة هو نفسه تقريبًا. في الوقت نفسه ، حاول الكيميائي الإنجليزي د. دالتون تصوير الصيغ الأولى للمواد البروتينية. قام بتمثيلهم كمواد بسيطة إلى حد ما ، ولكن من أجل التأكيد على اختلافاتهم الفردية مع نفس التركيب ، لجأ إلى تصوير الجزيئات التي ستسمى الآن أيزومري. ومع ذلك ، فإن مفهوم الأيزومرية لم يكن موجودًا بعد في زمن دالتون.
صيغ البروتين من قبل د دالتون
تم اشتقاق الصيغ التجريبية الأولى للبروتينات وطُرحت الفرضيات الأولى فيما يتعلق بانتظام تكوينها. لذلك ، اعتقد ن. ليبيركون أن الألبومين موصوف بالصيغة C 72 H 112 N 18 SO 22 ، ويعتقد A. Danilevsky أن جزيء هذا البروتين أكبر على الأقل من حيث الحجم: C 726 H 1171 N 194 S 3 O 214.
اقترح الكيميائي الألماني J. Liebig في عام 1841 أن البروتينات الحيوانية لها نظائر بين البروتينات النباتية: أدى استيعاب بروتين البقول في جسم الحيوان ، وفقًا لـ Liebig ، إلى تراكم بروتين مشابه - الكازين. واحدة من أكثر النظريات انتشارًا للكيمياء العضوية السابقة للهيكل كانت نظرية الجذور ، وهي المكونات الثابتة للمواد ذات الصلة. في عام 1836 ، اقترح الهولندي جي مولدر أن جميع البروتينات تحتوي على نفس الجذر ، وهو ما أسماه بروتين (من الكلمة اليونانية "أنا أتولى القيادة" ، "أنا آخذ المركز الأول"). البروتين ، وفقًا لمولدر ، كان له تكوين Pr = C 40 H 62 N 10 O 12. في عام 1838 ، نشر G.Mulder صيغ البروتين على أساس نظرية البروتين. كانت هذه ما يسمى ب. الصيغ الثنائية ، حيث يعمل الجذر البروتيني كتجمع إيجابي ، وذرات الكبريت أو الفوسفور كتجمع سالب. قاموا معًا بتكوين جزيء محايد كهربائيًا: بروتين مصل الدم Pr 10 S 2 P ، fibrin Pr 10 SP. ومع ذلك ، أظهر التحقق التحليلي لبيانات G.Mulder ، الذي أجراه الكيميائي الروسي Lyaskovskii ، وكذلك Yu. Liebig ، أن "جذور البروتين" غير موجودة.
في عام 1833 ، اكتشف العالم الألماني ف. روز تفاعل البيوريت للبروتينات - أحد التفاعلات اللونية الرئيسية للمواد البروتينية ومشتقاتها في الوقت الحاضر (المزيد عن التفاعلات اللونية في الصفحة 53). وخلص أيضًا إلى أن هذا كان أكثر تفاعلات البروتين حساسية ، لذلك جذب انتباه الكيميائيين في ذلك الوقت.
في منتصف القرن التاسع عشر ، تم تطوير طرق عديدة لاستخراج البروتينات وتنقيتها وعزلها في محاليل الأملاح المحايدة. في عام 1847 ، اكتشف K. Reichert قدرة البروتينات على تكوين البلورات. في عام 1836 ، اكتشف T. Schwann البيبسين ، وهو إنزيم يكسر البروتينات. في عام 1856 ، اكتشف L. Corvisar إنزيمًا مشابهًا آخر - التربسين. من خلال دراسة تأثير هذه الإنزيمات على البروتينات ، حاول علماء الكيمياء الحيوية كشف لغز الهضم. ومع ذلك ، فإن المواد الناتجة عن عمل الإنزيمات البروتينية (البروتياز ، وتشمل هذه الإنزيمات المذكورة أعلاه) على البروتينات جذبت أكبر قدر من الاهتمام: بعضها عبارة عن شظايا من جزيئات البروتين الأصلية (كانت تسمى ببتون ) ، بينما لم يتعرض البعض الآخر لمزيد من الانقسام بواسطة البروتياز وينتمون إلى فئة المركبات المعروفة منذ بداية القرن - الأحماض الأمينية (أول مشتق من الأحماض الأمينية ، الأسباراجين أميد ، تم اكتشافه في عام 1806 ، وأول حمض أميني ، سيستين ، في عام 1810). تم اكتشاف الأحماض الأمينية في تكوين البروتينات لأول مرة في عام 1820 من قبل الكيميائي الفرنسي A. Braconno. قام بتطبيق التحلل الحمضي للبروتين ووجد مادة حلوة في التحلل المائي ، والتي أطلق عليها اسم الجلايسين. في عام 1839 ، تم إثبات وجود الليوسين في البروتينات ، وفي عام 1849 ، عزل F. Bopp حمضًا أمينيًا آخر من البروتين - التيروزين (انظر الملحق الثاني للحصول على قائمة كاملة بتواريخ اكتشافات الأحماض الأمينية في البروتينات).
بحلول نهاية الثمانينيات. في القرن التاسع عشر ، تم بالفعل عزل 19 حمضًا أمينيًا من تحلل البروتين ، وبدأ الرأي يزداد ببطء حيث أن المعلومات حول منتجات التحلل المائي للبروتين تحمل معلومات مهمة حول بنية جزيء البروتين. ومع ذلك ، اعتبرت الأحماض الأمينية ضرورية ، ولكنها ليست المكون الرئيسي للبروتين.
فيما يتعلق باكتشافات الأحماض الأمينية في تكوين البروتينات ، قام العالم الفرنسي P. Schutzenberger في السبعينيات. اقترح القرن التاسع عشر ما يسمى ب. نظرية أوريد هياكل البروتين. وفقًا لذلك ، يتكون جزيء البروتين من لب مركزي ، يلعب دوره جزيء التيروزين ، ومجموعات معقدة مرتبطة به (مع استبدال 4 ذرات هيدروجين) ، تسمى شوتزنبرجر ليسين . ومع ذلك ، تم دعم الفرضية بشكل ضعيف جدًا تجريبيًا ، وأثبتت الأبحاث الإضافية أنها غير متسقة.
2.2 نظرية "مجمعات الكربون والنيتروجين" A.Ya. دانيلفسكي
تم التعبير عن النظرية الأصلية حول بنية البروتين في الثمانينيات. عالم الكيمياء الحيوية الروسي في القرن التاسع عشر A. Ya. Danilevsky. كان أول كيميائي يلفت الانتباه إلى الطبيعة البوليمرية المحتملة لهيكل جزيئات البروتين. في أوائل السبعينيات. كتب إلى أ.م. باتلروف أن "جسيمات الألبومين عبارة عن بوليمر مختلط" ، ولم يجد تعريف البروتين "مصطلحًا أكثر ملاءمة من كلمة بوليمر بالمعنى الواسع". عند دراسة تفاعل البيوريت ، اقترح أن هذا التفاعل مرتبط ببنية ذرات الكربون والنيتروجين المتقطعة - N - C - N - C - N - ، والتي يتم تضمينها في ما يسمى. كاربونازو ر مركب R "- NH - CO - NH - CO - R". بناءً على هذه الصيغة ، يعتقد Danilevsky أن جزيء البروتين يحتوي على 40 من هذه المركبات من الكربون والنيتروجين. بدت مجمعات الأحماض الأمينية الكربونية والنيتروجينية ، وفقًا لـ Danilevsky ، كما يلي:
وفقًا لـ Danilevsky ، يمكن ربط مجمعات الكربون النيتروجين عن طريق رابطة إيثر أو أميد لتشكيل بنية جزيئية عالية.
2.3 نظرية "kirins" أ. كوسيل
اكتشف عالم الفيزياء والكيمياء الحيوية الألماني أ. كوسيل ، الذي درس البروتامين والهيستونات ، وهي بروتينات بسيطة نسبيًا ، أن كمية كبيرة من الأرجينين تتشكل أثناء التحلل المائي. بالإضافة إلى ذلك ، اكتشف في تكوين التحلل المائي الحمض الأميني غير المعروف آنذاك - الهيستيدين. بناءً على ذلك ، اقترح كوسيل أن هذه المواد البروتينية يمكن اعتبارها بعضًا من أبسط النماذج للبروتينات الأكثر تعقيدًا ، والتي تم بناؤها ، في رأيه ، وفقًا للمبدأ التالي: الأرجينين والهيستيدين يشكلان لبًا مركزيًا ("نواة البروتامين") ، وهو محاط بمجمعات من الأحماض الأمينية الأخرى.
كانت نظرية كوسيل المثال الأكثر كمالًا لتطور فرضية البنية المجزأة للبروتينات (اقترحها ج. مولدر لأول مرة ، كما ذكر أعلاه). استخدم الكيميائي الألماني م. سيجفريد هذه الفرضية في بداية القرن العشرين. كان يعتقد أن البروتينات تُبنى من مجمعات الأحماض الأمينية (أرجينين + ليسين + حمض الجلوتامين) ، وهو ما أسماه كيرينامي (من الكلمة اليونانية "kyrios" الأساسية). ومع ذلك ، تم طرح هذه الفرضية في عام 1903 ، عندما كان إي فيشر يعمل بنشاط على تطوير نظريته نظرية الببتيد الذي أعطى مفتاح سر بنية البروتينات.
2.4 نظرية الببتيد E. فيشر
قام الكيميائي الألماني إميل فيشر ، المشهور بالفعل في جميع أنحاء العالم بدراساته لمركبات البيورين (قلويدات مجموعة الكافيين) وفك تشفير بنية السكريات ، بإنشاء نظرية الببتيد ، والتي تم تأكيدها إلى حد كبير في الممارسة وحصلت على اعتراف عالمي خلال حياته ، التي نال عنها جائزة نوبل الثانية في تاريخ الكيمياء. الجوائز (الأولى حصل عليها Ya.G. Van't Hoff).
من المهم أن يقوم فيشر ببناء خطة بحث تختلف بشكل حاد عما تم القيام به من قبل ، ولكنها تأخذ في الاعتبار جميع الحقائق المعروفة في ذلك الوقت. بادئ ذي بدء ، وافق على أنها الفرضية الأكثر ترجيحًا القائلة بأن البروتينات تُبنى من الأحماض الأمينية المتصلة برابطة أميد:
دعا فيشر هذا النوع من السندات (عن طريق القياس مع بيبتونيس) الببتيد . اقترح أن البروتينات بوليمرات الأحماض الأمينية المرتبطة بروابط الببتيد . تم التعبير عن فكرة الطبيعة البوليمرية لبنية البروتينات ، كما هو معروف جيدًا ، من قبل Danilevsky و Hert ، لكنهم اعتقدوا أن "المونومرات" عبارة عن تكوينات معقدة للغاية - بيبتون أو "معقدات كربون-نيتروجين".
إثبات نوع مركب الببتيد من بقايا الأحماض الأمينية. E. انطلق فيشر من الملاحظات التالية. أولاً ، أثناء التحلل المائي للبروتينات وأثناء تحللها الأنزيمي ، تم تكوين أحماض أمينية مختلفة. كان من الصعب للغاية وصف المركبات الأخرى بل وكان الحصول عليها أكثر صعوبة. بالإضافة إلى ذلك ، عرف فيشر أن البروتينات ليس لها غلبة للخصائص الحمضية أو الأساسية ، مما يعني ، كما جادل ، أن المجموعات الأمينية والكربوكسيلية في تكوين الأحماض الأمينية في جزيئات البروتين مغلقة ، كما هي ، تخفي بعضها البعض ( تذبذب البروتينات ، كما يقولون الآن).
قسم فيشر الحل لمشكلة بنية البروتين ، واختصره إلى البنود التالية:
التحديد النوعي والكمي لمنتجات التحلل المائي الكامل للبروتينات.
إنشاء هيكل هذه المنتجات النهائية.
تخليق بوليمرات الأحماض الأمينية مع مركبات من نوع الأميد (الببتيد).
مقارنة المركبات التي تم الحصول عليها بهذه الطريقة مع البروتينات الطبيعية.
من هذه الخطة ، يمكن ملاحظة أن فيشر استخدم لأول مرة نهجًا منهجيًا جديدًا - توليف مركبات النموذج ، كطريقة للإثبات عن طريق القياس.
2.5 تطوير طرق لتخليق الأحماض الأمينية
من أجل المضي قدمًا في تصنيع مشتقات الأحماض الأمينية المرتبطة برابطة الببتيد ، قام فيشر بعمل كبير في دراسة بنية الأحماض الأمينية وتركيبها.
قبل فيشر ، كانت الطريقة العامة لتركيب الأحماض الأمينية هي توليف A. Strecker cyanohydrin:
وفقًا لتفاعل Strecker ، كان من الممكن تصنيع الألانين والسيرين وبعض الأحماض الأمينية الأخرى ، ووفقًا لتعديله (تفاعل Zelinsky-Stadnikov) ، كل من الأحماض الأمينية والأحماض N- المستبدلة.
ومع ذلك ، سعى فيشر نفسه إلى تطوير طرق لتخليق جميع الأحماض الأمينية المعروفة آنذاك. لقد اعتبر طريقة ستريكر ليست عالمية بما فيه الكفاية. لذلك ، كان على E. Fischer البحث عن طريقة عامة لتخليق الأحماض الأمينية ، بما في ذلك الأحماض الأمينية مع الجذور الجانبية المعقدة.
واقترح تحسين الأحماض الكربوكسيلية المستبدلة بالبرومو في الوضع. للحصول على مشتقات البرومو ، استخدم ، على سبيل المثال ، في تركيب حمض مالونيك الليوسين أو الأريللات أو الألكيلات:
لكن إي فيشر فشل في إنشاء طريقة عالمية تمامًا. كما تم تطوير تفاعلات أكثر موثوقية. على سبيل المثال ، اقترح طالب فيشر جي.لايكس التعديل التالي للحصول على سيرين:
أثبت فيشر أيضًا أن البروتينات تتكون من بقايا أحماض أمينية نشطة بصريًا (انظر ص 11). أجبره ذلك على تطوير تسمية جديدة للمركبات النشطة بصريًا ، وطرق لفصل وتوليف الأيزومرات الضوئية للأحماض الأمينية. توصل فيشر أيضًا إلى استنتاج مفاده أن البروتينات تحتوي على بقايا أشكال L للأحماض الأمينية النشطة بصريًا ، وقد أثبت ذلك باستخدام مبدأ التشاؤم الدياستيري. كان هذا المبدأ على النحو التالي: تمت إضافة قلويد نشط بصريًا (بروسين ، ستريكنين ، سينكونين ، كينيدين ، كينين) إلى مشتق N-acyl من حمض أميني راسيمي. نتيجة لذلك ، تم تشكيل شكلين من الأيزومرات الفراغية للأملاح ذات قابلية ذوبان مختلفة. بعد فصل هذه الأيزومرات ، تمت استعادة القلويد وإزالة مجموعة الأسيل عن طريق التحلل المائي.
تمكن فيشر من تطوير طريقة للتقدير الكامل للأحماض الأمينية في منتجات التحلل المائي للبروتين: قام بتحويل إسترات هيدروكلوريد الأحماض الأمينية عن طريق المعالجة بالقلويات المركزة في البرد إلى إسترات حرة ، والتي لم يتم تصبنها بشكل ملحوظ. ثم تعرض خليط هذه الإيثرات للتقطير الجزئي وعزل الأحماض الأمينية الفردية من الكسور الناتجة عن طريق التبلور الجزئي.
لم تؤكد طريقة التحليل الجديدة أخيرًا أن البروتينات تتكون من بقايا الأحماض الأمينية فحسب ، بل جعلت من الممكن صقل قائمة الأحماض الأمينية الموجودة في البروتينات وتكميلها. لكن مع ذلك ، لم تستطع التحليلات الكمية الإجابة على السؤال الرئيسي: ما هي مبادئ بنية جزيء البروتين. وصاغ إي فيشر إحدى المهام الرئيسية في دراسة بنية وخصائص البروتينات: التطور م التجريبيةهطرق لتخليق المركبات التي تكون مكوناتها الرئيسية هي الأحماض الأمينيةحولقمت بتوصيله برباط ببتيد.
وهكذا ، وضع فيشر مهمة غير تافهة - لتجميع فئة جديدة من المركبات من أجل إنشاء مبادئ هيكلها.
قام فيشر بحل هذه المشكلة ، وتلقى الكيميائيون دليلًا مقنعًا على أن البروتينات عبارة عن بوليمرات من الأحماض الأمينية المرتبطة برابطة الببتيد:
CO - CHR "- NH - CO - CHR" "- NH - CO CHR" "" - NH -
كان هذا الموقف مدعومًا بالأدلة البيوكيميائية. على طول الطريق ، اتضح أن البروتياز لا يحلل جميع الروابط بين الأحماض الأمينية بنفس المعدل. تأثرت قدرتها على شق رابطة الببتيد بالتكوين البصري للأحماض الأمينية ، والبدائل في نيتروجين المجموعة الأمينية ، وطول سلسلة الببتيد ، ومجموعة البقايا المتضمنة فيها.
كان الدليل الرئيسي لنظرية الببتيد هو تخليق الببتيدات النموذجية ومقارنتها مع بيبتون للبروتين المائي. وأظهرت النتائج أن الببتيدات المماثلة لتلك المصنعة معزولة من تحلل البروتين.
في سياق هذه الدراسات ، طور إي فيشر وتلميذه إي أبديرغالدن طريقة لتحديد تسلسل الأحماض الأمينية في البروتين لأول مرة. كان جوهره هو تحديد طبيعة بقايا الأحماض الأمينية لبولي ببتيد الذي يحتوي على مجموعة أمينية حرة (حمض أميني N- طرفي). للقيام بذلك ، اقترحوا حجب النهاية الأمينية في الببتيد بمجموعة النفثالين-سلفونيل ، والتي لا تنفصل أثناء التحلل المائي. من خلال عزل الحمض الأميني المسمى بمثل هذه المجموعة من التحلل المائي ، كان من الممكن تحديد أي من الأحماض الأمينية كان N- طرفي.
بعد بحث E. فيشر ، أصبح من الواضح أن البروتينات هي عديد الببتيدات. كان هذا إنجازًا مهمًا ، بما في ذلك مهام تخليق البروتين: أصبح من الواضح ما هو بالضبط المطلوب تصنيعه.فقط بعد هذه الأعمال اكتسبت مشكلة تخليق البروتين اتجاهًا معينًا والصرامة اللازمة.
عند الحديث عن عمل فيشر ككل ، تجدر الإشارة إلى أن نهج البحث نفسه كان نموذجيًا إلى حد ما في القرن العشرين القادم - فقد عمل مع مجموعة واسعة من المواقف النظرية والتقنيات المنهجية ؛ بدت تركيباته أقل وأقل شبهاً بالفن القائم على الحدس أكثر من كونها على المعرفة الدقيقة ، واقتربت من إنشاء سلسلة من الأجهزة الدقيقة شبه التكنولوجية.
2. 6 أزمة نظرية الببتيد
فيما يتعلق باستخدام طرق البحث الفيزيائية والكيميائية الجديدة في أوائل العشرينات. القرن ال 20 كانت هناك شكوك في أن جزيء البروتين عبارة عن سلسلة طويلة من عديد الببتيد. تم التعامل مع الفرضية حول إمكانية التعبئة المدمجة لسلاسل الببتيد بشك. كل هذا يتطلب مراجعة نظرية الببتيد لإي فيشر.
في 20-30s. تم تبني نظرية ديكيتوبيبرازين على نطاق واسع. وفقًا لذلك ، تلعب حلقات diketopiperase ، التي تتشكل أثناء تحلل اثنين من بقايا الأحماض الأمينية ، دورًا رئيسيًا في بناء بنية البروتين. وافترض أيضًا أن هذه الهياكل تشكل اللب المركزي للجزيء ، الذي ترتبط به الببتيدات القصيرة أو الأحماض الأمينية ("حشوات" الهيكل الدوري للهيكل الرئيسي). تم تقديم المخططات الأكثر إقناعًا لمشاركة diketopiperazines في بناء بنية البروتين من قبل طلاب ND Zelinsky و E. Fisher.
ومع ذلك ، فإن محاولات تصنيع مركبات نموذجية تحتوي على ديكيتوبيبرازين لم تحقق سوى القليل في كيمياء البروتين ؛ وبالتالي انتصرت نظرية الببتيد ، ولكن كان لهذه الأعمال تأثير محفز على كيمياء البيبرازينات بشكل عام.
بعد نظريات الببتيد و diketopiperase ، استمرت المحاولات لإثبات وجود هياكل الببتيد فقط في جزيء البروتين. في الوقت نفسه ، حاولوا تخيل ليس فقط نوع الجزيء ، ولكن أيضًا الخطوط العريضة العامة له.
تم التعبير عن الفرضية الأصلية من قبل الكيميائي السوفيتي DL التلمود. اقترح أن سلاسل الببتيد في تكوين جزيئات البروتين مطوية في حلقات كبيرة ، والتي بدورها كانت خطوة نحو تكوين فكرته عن الكريات البروتينية.
في الوقت نفسه ، ظهرت بيانات تشير إلى مجموعة مختلفة من الأحماض الأمينية في بروتينات مختلفة. لكن الأنماط التي تحكم تسلسل الأحماض الأمينية في بنية البروتين لم تكن واضحة.
كان M. Bergman و K. Niemann أول من حاول الإجابة على هذا السؤال في فرضيتهما حول "الترددات المتقطعة". وفقًا لذلك ، فإن تسلسل بقايا الأحماض الأمينية في جزيء البروتين يخضع للأنماط العددية ، والتي تم اشتقاق أسسها من مبادئ بنية جزيء بروتين الفيبروين الحريري. لكن هذا الاختيار لم ينجح ، لأنه. هذا البروتين هو ليفي ، بينما يتبع هيكل البروتينات الكروية أنماطًا مختلفة تمامًا.
وفقًا لـ M. Bergman و K. Nieman ، يحدث كل حمض أميني في سلسلة polypeptide في فترة زمنية معينة أو ، كما قال M. Bergman ، له "دورية" معينة. يتم تحديد هذه الدورية من خلال طبيعة مخلفات الأحماض الأمينية.
لقد تخيلوا جزيء فيبروين الحرير على النحو التالي:
جليالجليتير جليالجليارج جليالجليكس جليالجليكس
(GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyx) 12
GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyArg
(GlyAlaGlyTyr GlyAlaGlyx GlyAlaGlyx GlyAlaGlyx) 13
كان لفرضية Bergman-Niemann تأثير كبير على تطوير كيمياء الأحماض الأمينية ، حيث تم تخصيص عدد كبير من الأعمال للتحقق منها.
في ختام هذا الفصل ، تجدر الإشارة إلى أنه بحلول منتصف القرن العشرين. تم تجميع أدلة كافية على صحة نظرية الببتيد ، وتم استكمال وتنقيح أحكامها الرئيسية. لذلك كان مركز أبحاث البروتين في القرن العشرين. وضع بالفعل مجال البحث والبحث عن طرق تخليق البروتين بالوسائل الاصطناعية. تم حل هذه المشكلة بنجاح ، وتم تطوير طرق موثوقة لتحديد الهيكل الأساسي للبروتين - تسلسل الأحماض الأمينية في سلسلة الببتيد ، وتم تطوير طرق التخليق الكيميائي (غير الحيوي) لعديد الببتيدات غير النظامية (تمت مناقشة هذه الطرق بمزيد من التفصيل في الفصل 8 ، ص 36) ، بما في ذلك طرق التوليف التلقائي لعديد الببتيدات. هذا جعل من الممكن بالفعل في عام 1962 لأكبر كيميائي إنكليزي F. Senger فك شفرة الهيكل وتوليف هرمون الأنسولين بشكل مصطنع ، والذي يمثل حقبة جديدة في تخليق البروتين الوظيفي متعدد الببتيدات.
الفصل 3. التركيب الكيميائي للبروتينات
3.1 رابطة الببتيد
البروتينات عبارة عن بوليمرات غير منتظمة مبنية من بقايا حمض أميني ألفا ، ويمكن كتابة الصيغة العامة لها في محلول مائي عند قيم الأس الهيدروجيني القريبة من المحايد على أنها NH 3 + CHRCOO -. ترتبط بقايا الأحماض الأمينية في البروتينات ببعضها البعض عن طريق رابطة أميد بين مجموعات α-amino و β-carboxyl. رابطة الببتيد بين اثنين- بقايا الأحماض الأمينية يشار إليها عادة باسم السندات الببتيد ، والبوليمرات المبنية من بقايا الأحماض الأمينية ألفا المتصلة بواسطة روابط الببتيد تسمى بولي ببتيدات. يمكن أن يكون البروتين كهيكل مهم بيولوجيًا إما عديد ببتيد واحد أو عدة ببتيدات متعددة تشكل معقدًا واحدًا نتيجة للتفاعلات غير التساهمية.
3.2 التكوين الأولي للبروتينات
عند دراسة التركيب الكيميائي للبروتينات ، من الضروري معرفة ، أولاً ، ما هي العناصر الكيميائية التي تتكون منها ، وثانيًا ، بنية مونومراتها. للإجابة على السؤال الأول ، يتم تحديد التركيب الكمي والنوعي للعناصر الكيميائية للبروتين. أظهر التحليل الكيميائي موجود في جميع البروتينات كربون (50-55٪) ، أكسجين (21-23٪) ، نيتروجين (15-17٪) ، هيدروجين (6-7٪) ، كبريت (0.3-2.5٪). تم العثور أيضًا على الفوسفور واليود والحديد والنحاس وبعض العناصر الكبيرة والصغرى الأخرى في تكوين البروتينات الفردية ، بكميات مختلفة ، وغالبًا ما تكون صغيرة جدًا.
يمكن أن يختلف محتوى العناصر الكيميائية الرئيسية في البروتينات ، باستثناء النيتروجين الذي يتميز تركيزه بأكبر ثبات ويبلغ متوسطه 16٪. بالإضافة إلى ذلك ، فإن محتوى النيتروجين في المواد العضوية الأخرى منخفض. وفقًا لهذا ، تم اقتراح تحديد كمية البروتين بواسطة النيتروجين المكون له. مع العلم أن 1 جرام من النيتروجين موجود في 6.25 جرام من البروتين ، يتم ضرب الكمية الموجودة من النيتروجين في معامل 6.25 ويتم الحصول على كمية البروتين.
لتحديد الطبيعة الكيميائية لمونومرات البروتين ، من الضروري حل مشكلتين: فصل البروتين إلى مونومرات ومعرفة تركيبها الكيميائي. يتم تحقيق تفكك البروتين إلى الأجزاء المكونة له عن طريق التحلل المائي - غليان البروتين لفترة طويلة مع الأحماض المعدنية القوية. (التحلل الحمضي)أو أسباب (تحلل قلوي). الغليان عند 110 درجة مئوية مع حمض الهيدروكلوريك لمدة 24 ساعة هو الأكثر شيوعًا ، وفي المرحلة التالية ، يتم فصل المواد التي يتكون منها التحلل المائي. لهذا الغرض ، يتم استخدام طرق مختلفة ، في أغلب الأحيان - الكروماتوغرافيا (لمزيد من التفاصيل ، انظر فصل "طرق البحث ..."). الأحماض الأمينية هي الجزء الرئيسي من التحلل المائي المنفصل.
3.3 أحماض أمينية
حاليًا ، تم العثور على ما يصل إلى 200 من الأحماض الأمينية المختلفة في كائنات مختلفة من الحياة البرية. في جسم الإنسان ، على سبيل المثال ، هناك حوالي 60 نوعًا ، لكن البروتينات تحتوي فقط على 20 حمضًا أمينيًا ، وأحيانًا تسمى الأحماض الأمينية الطبيعية.
الأحماض الأمينية هي أحماض عضوية يتم فيها استبدال ذرة الهيدروجين - ذرة الكربون بمجموعة أمينية - NH 2. لذلك ، بطبيعتها الكيميائية ، هذه أحماض أمينية لها الصيغة العامة:
من هذه الصيغة يمكن ملاحظة أن تكوين جميع الأحماض الأمينية يشمل المجموعات العامة التالية: - CH 2 - NH 2 - COOH. السلاسل الجانبية (الجذور - ص) الأحماض الأمينية تختلف. كما يتضح من الملحق الأول ، فإن الطبيعة الكيميائية للجذور متنوعة: من ذرة الهيدروجين إلى المركبات الحلقية. الجذور هي التي تحدد السمات الهيكلية والوظيفية للأحماض الأمينية.
جميع الأحماض الأمينية ، باستثناء أبسط الأحماض الأمينية بالنسبة لك الجلايسين (NH 3 + CH 2 COO) لها ذرة مراوان C ويمكن أن توجد في شكل اثنين من المتشاهدين (أيزومرات بصرية):
تحتوي جميع البروتينات المدروسة حاليًا على أحماض أمينية فقط من السلسلة L ، والتي ، إذا أخذنا في الاعتبار الذرة اللولبية من جانب ذرة H ، فإن مجموعات NH 3 + و COO وجذر R تقع في اتجاه عقارب الساعة. إن الحاجة إلى بناء جزيء بوليمر مهم بيولوجيًا من متماثل محدد بدقة أمر واضح - من خليط راسمي من اثنين من المتضادمات ، يمكن الحصول على خليط معقد بشكل لا يمكن تصوره من الأيزومرات ثنائية الاتجاه. لا يزال السؤال عن سبب استناد الحياة على الأرض إلى البروتينات المبنية بدقة من L- وليس الأحماض الأمينية D- لغزًا مثيرًا للاهتمام. تجدر الإشارة إلى أن الأحماض الأمينية D منتشرة إلى حد ما بطبيعتها ، علاوة على ذلك ، فهي جزء من oligopeptides ذات أهمية بيولوجية.
تُبنى البروتينات من عشرين حمضًا أمينيًا أساسيًا ، لكن البقية ، وهي أحماض أمينية متنوعة تمامًا ، تتشكل من 20 بقايا من الأحماض الأمينية الموجودة بالفعل في تكوين جزيء البروتين. من بين هذه التحولات ، ينبغي للمرء أولاً وقبل كل شيء أن يلاحظ التكوين جسور ثاني كبريتيد أثناء أكسدة اثنين من بقايا السيستين في تكوين سلاسل الببتيد المشكلة بالفعل. نتيجة لذلك ، يتم تكوين بقايا حمض ديامينوديكاربوكسيليك من اثنين من بقايا السيستين سيستين (انظر الملحق الأول). في هذه الحالة ، يحدث الارتباط المتقاطع إما داخل سلسلة بولي ببتيد واحدة أو بين سلسلتين مختلفتين. كبروتين صغير يحتوي على سلسلتين متعدد الببتيد متصلتين بجسور ثاني كبريتيد ، بالإضافة إلى روابط متقاطعة داخل إحدى سلاسل البولي ببتيد:
مثال مهم لتعديل بقايا الأحماض الأمينية هو تحويل بقايا البرولين إلى بقايا هيدروكسي برولين :
يحدث هذا التحول ، وعلى نطاق واسع ، أثناء تكوين مكون بروتيني مهم للنسيج الضام - الكولاجين .
نوع آخر مهم جدًا من تعديل البروتين هو فسفرة مجموعات الهيدروكسو من بقايا السيرين والثريونين والتيروزين ، على سبيل المثال:
الأحماض الأمينية في محلول مائي هي في حالة متأينة بسبب تفكك مجموعات الأمينو والكربوكسيل التي تشكل الجذور. بمعنى آخر ، فهي مركبات مذبذبة ويمكن أن توجد إما كأحماض (مانحون بروتون) أو كقاعدة (متقبلات مانحة).
جميع الأحماض الأمينية ، حسب التركيب ، تنقسم إلى عدة مجموعات:
لا دوري. أحماض أمينية أحادية أمينومونوكربوكسيليةتحتوي في تركيبتها على أمين واحد ومجموعة كربوكسيل واحدة ، في محلول مائي تكون محايدة. يحتوي بعضها على ميزات هيكلية مشتركة ، مما يسمح بالنظر فيها معًا:
جلايسين وألانين.الجلايسين (الجليكوكول أو الأحماض الأمينية) غير نشط بصريًا - إنه الحمض الأميني الوحيد الذي لا يحتوي على مادة متناهية الصغر. يشارك الجليسين في تكوين المادة الصفراوية والنووية إلى الهيم ، وهو ضروري لتحييد المنتجات السامة في الكبد. يستخدم الجسم ألانين في عمليات التمثيل الغذائي للكربوهيدرات والطاقة المختلفة. الأيزومير - ألانين هو جزء لا يتجزأ من فيتامين البانتوثينيك - أنت ، الإنزيم المساعد A (CoA) ، والمواد المستخرجة للعضلات.
سيرين وثريونين.إنهم ينتمون إلى مجموعة أحماض هيدروكسي ، لأن. لديك مجموعة الهيدروكسيل. السرين هو جزء من الإنزيمات المختلفة ، البروتين الرئيسي للحليب - الكازين ، وهو أيضًا جزء من العديد من البروتينات الدهنية. يشارك ثريونين في التخليق الحيوي للبروتين ، كونه حمض أميني أساسي.
السيستين والميثيونين.أحماض أمينية تحتوي على ذرة كبريت. يتم تحديد قيمة السيستين من خلال وجود مجموعة السلفهيدريل (-SH) في تركيبته ، مما يمنحها القدرة على أكسدة الجسم بسهولة وحماية الجسم من المواد ذات القدرة العالية على الأكسدة (في حالة الإصابة الإشعاعية ، الفوسفور تسمم). يتميز الميثيونين بوجود مجموعة ميثيل سهلة الحركة ، والتي تستخدم لتخليق مركبات مهمة في الجسم (الكولين ، الكرياتين ، الثايمين ، الأدرينالين ، إلخ).
فالين وليوسين وآيزولوسين.وهي عبارة عن أحماض أمينية متفرعة تشارك بنشاط في عملية التمثيل الغذائي ولا يتم تصنيعها في الجسم.
أحماض أمينية أحادية الكربوكسيلتحتوي على مجموعة أمينية واثنين من مجموعات الكربوكسيل وتعطي تفاعلًا حمضيًا في محلول مائي. وتشمل هذه الأسبارتيك والجلوتامين بالنسبة لك ، والأسباراجين والجلوتامين. هم جزء من الوسطاء المثبطين للجهاز العصبي.
الأحماض الأمينية Diaminomonocarboxylicفي محلول مائي لها تفاعل قلوي بسبب وجود مجموعتين أمين. بالنسبة لهم ، يعتبر اللايسين ضروريًا لتخليق الهستونات وأيضًا في عدد من الإنزيمات. يشارك أرجينين في تركيب اليوريا والكرياتين.
دوري. تحتوي هذه الأحماض الأمينية على نواة عطرية أو حلقية غير متجانسة في تكوينها ، وكقاعدة عامة ، لا يتم تصنيعها في جسم الإنسان ويجب تزويدها بالطعام. يشاركون بنشاط في مجموعة متنوعة من عمليات التمثيل الغذائي. لذا فإن فينيل ألانين يعمل كمصدر رئيسي لتخليق التيروزين - مقدمة لعدد من المواد المهمة بيولوجيًا: الهرمونات (هرمون الغدة الدرقية ، الأدرينالين) ، وبعض الأصباغ. التربتوفان ، بالإضافة إلى مشاركته في تخليق البروتين ، هو أحد مكونات فيتامين PP ، السيروتونين ، التربتامين ، وعدد من الأصباغ. الهيستيدين ضروري لتخليق البروتينات ، وهو مقدمة للهيستامين ، الذي يؤثر على ضغط الدم وإفراز العصارة المعدية.
الفصل 4. الهيكل
عند دراسة تكوين البروتينات ، وجد أنها كلها مبنية وفقًا لمبدأ واحد ولها أربعة مستويات من التنظيم: الابتدائي والثانوي والجامعي ،وبعضهم رباعيالهياكل.
4.1 الهيكل الأساسي
إنها سلسلة خطية من الأحماض الأمينية مرتبة في تسلسل معين ومترابطة بواسطة روابط الببتيد. السندات الببتيد تتكون من مجموعة-carboxyl لحمض أميني ومجموعة -amine أخرى:
لا يمكن اعتبار الرابطة الببتيدية الناتجة عن p ، - الاقتران - مجموعة كربونيل والمدار p للذرة N ، حيث يوجد زوج الإلكترونات غير المشترك ، كواحد واحد ولا يوجد عمليًا دوران حوله هو - هي. للسبب نفسه ، تكون الذرة اللولبية C وذرة الكربونيل Ck من أي بقايا حمض أميني i من سلسلة الببتيد وذرات N و C من البقايا (i + 1) في نفس المستوى. تقع ذرة الكربونيل O وذرة الأميد H في نفس المستوى (ومع ذلك ، فإن المادة المتراكمة في دراسة بنية البروتينات تظهر أن هذا البيان ليس صارمًا تمامًا: الذرات المرتبطة بذرة النيتروجين الببتيدية ليست في نفس المستوى معها ، ولكن تشكل هرمًا ثلاثي السطوح بزوايا بين روابط قريبة جدًا من 120. لذلك ، بين المستويات التي تكونت من الذرات C i و C i k و O i و N i +1 ، H i +1 ، C i + 1 ، هناك بعض الزاوية التي تختلف عن 0. ولكن كقاعدة عامة ، لا تتجاوز 1 ولا تلعب دورًا خاصًا). لذلك ، من الناحية الهندسية ، يمكن اعتبار سلسلة البولي ببتيد متكونة من مثل هذه الأجزاء المسطحة التي تحتوي كل منها على ست ذرات. يجب تحديد الترتيب المتبادل لهذه الأجزاء ، مثل أي ترتيب متبادل لطائرتين ، من خلال زاويتين. على هذا النحو ، من المعتاد أخذ زوايا الالتواء التي تميز الدوران حول N C و C C k -bonds.
يتم تحديد هندسة أي جزيء من خلال ثلاث مجموعات من الخصائص الهندسية لروابطه الكيميائية - أطوال الرابطة وزوايا الرابطة وزوايا الالتواءبين الروابط المتاخمة للذرات المجاورة. يتم تحديد أول مجموعتين إلى حد حاسم من خلال طبيعة الذرات المشاركة والروابط المتكونة. لذلك ، يتم تحديد التركيب المكاني للبوليمرات بشكل أساسي من خلال زوايا الالتواء بين روابط العمود الفقري للبوليمر للجزيئات ، أي تشكيل سلسلة البوليمر. الذي - التي ص زاوية سيون ، بمعنى آخر. زاوية دوران اتصال A-B حول اتصال B-C بالنسبة لاتصال C-د، هي الزاوية بين المستويات التي تحتوي على ذرات A و B و C والذراتب, ج, د.
في مثل هذا النظام ، من الممكن أن تكون السندات A-B و C-D موجودة على التوازي وعلى نفس الجانب من الرابطة B-C. إذا اعتبرنا هذا النظام على طولأناzi B-C ، ثم الاتصال A-B ، كما كان ، يحجب الاتصالج- د، لذلك يسمى هذا التشكلسفيتسيامشوشة. وفقًا لتوصيات الاتحادات الدولية للكيمياء IUPAC (الاتحاد الدولي للكيمياء البحتة والتطبيقية) و IUB (الاتحاد الدولي للكيمياء الحيوية) ، تعتبر الزاوية بين طائرات ABC و BCD إيجابية إذا ، من أجل إحضار التشكل إلى حالة الكسوف عن طريق الدوران بزاوية لا تزيد عن 180 ، يجب تدوير الأقرب إلى اتصال المراقب في اتجاه عقارب الساعة. إذا كان لا بد من تدوير هذه الرابطة عكس اتجاه عقارب الساعة للحصول على شكل مكسوف ، فإن الزاوية تعتبر سالبة. يمكن ملاحظة أن هذا التعريف لا يعتمد على أي الروابط أقرب إلى المراقب.
في هذه الحالة ، كما يتضح من الشكل ، اتجاه الجزء الذي يحتوي على الذرات C i -1 و C i [(i-1) -th part] ، والجزء الذي يحتوي على الذرات C i و C i + 1 (الجزء الأول) ، يتم تحديده بواسطة زوايا الالتواء المقابلة للدوران حول الرابطة N i C i والرابطة C i C i k. عادةً ما يتم الإشارة إلى هذه الزوايا كـ i و i على التوالي في هذه الحالة. تحدد قيمها لجميع وحدات المونومر لسلسلة البولي ببتيد بشكل أساسي هندسة هذه السلسلة. لا توجد قيم لا لبس فيها لقيمة كل من هذه الزوايا أو لمجموعاتها ، على الرغم من القيود المفروضة على كل منهما ، والتي تحددها كل من خصائص شظايا الببتيد نفسها وطبيعة الجذور الجانبية ، أي. طبيعة بقايا الأحماض الأمينية.
حتى الآن ، تم إنشاء تسلسل الأحماض الأمينية لعدة آلاف من البروتينات المختلفة. يعد تسجيل بنية البروتينات في شكل صيغ هيكلية مفصلة أمرًا مرهقًا وليس مرئيًا. لذلك ، يتم استخدام شكل مختصر من الكتابة - ثلاثة أحرف أو حرف واحد (جزيء فاسوبريسين):
عند كتابة تسلسل حمض أميني في سلاسل polypeptide أو oligopeptide باستخدام رموز مختصرة ، يُفترض ، ما لم يُذكر خلاف ذلك ، أن مجموعة α-amino على اليسار ومجموعة α-carboxyl على اليمين. تسمى الأقسام المقابلة من سلسلة البولي ببتيد بالنهاية N (نهاية الأمين) والطرف C (نهاية الكربوكسيل) ، وتسمى بقايا الأحماض الأمينية بمخلفات N-terminal و C- الطرفية ، على التوالي.
4.2 الهيكل الثانوي
تعتبر شظايا التركيب المكاني للبوليمر الحيوي الذي له هيكل دوري للعمود الفقري للبوليمر عناصر من البنية الثانوية.
إذا كانت الزوايا من نفس النوع ، والتي تم ذكرها في الصفحة 15 ، متماثلة تقريبًا فوق قسم معين من السلسلة ، فإن هيكل سلسلة البولي ببتيد يكتسب طابعًا دوريًا. هناك فئتان من هذه الهياكل - حلزوني وممتد (مسطح أو مطوي).
حلزونييعتبر الهيكل الذي تقع فيه جميع الذرات من نفس النوع على نفس اللولب. في هذه الحالة ، يعتبر اللولب صحيحًا إذا تحرك بعيدًا عن المراقب في اتجاه عقارب الساعة ، عند ملاحظته على طول محور اللولب ، ويسارًا - إذا تحرك بعيدًا عكس اتجاه عقارب الساعة. تحتوي سلسلة البولي ببتيد على شكل حلزوني إذا كانت جميع ذرات C على حلزون واحد ، وجميع ذرات الكربونيل C k - من جهة أخرى ، جميع ذرات N - في المجموعة الثالثة ، ويجب أن تكون درجة اللولب لجميع المجموعات الثلاث من الذرات متماثلة. يجب أن يكون عدد الذرات في كل دورة واحدة من اللولب هو نفسه أيضًا ، بغض النظر عما إذا كنا نتحدث عن الذرات C k أو C أو N. تختلف المسافة إلى اللولب المشترك لكل نوع من هذه الأنواع الثلاثة من الذرات.
العناصر الرئيسية للتركيب الثانوي للبروتينات هي -الصفائح و- الطيات.
هياكل البروتين الحلزونية. تُعرف عدة أنواع مختلفة من الحلزونات بسلاسل البولي ببتيد. من بينها ، الحلزون الأيمن هو الأكثر شيوعًا. الحلزون المثالي له درجة 0.54 نانومتر وعدد الذرات من نفس النوع في كل دورة من اللولب هو 3.6 ، مما يعني دورية كاملة على خمس لفات من اللولب كل 18 بقايا من الأحماض الأمينية. قيم زوايا الالتواء للحلزون المثالي α = - 57 = - 47 والمسافات من الذرات التي تشكل سلسلة البولي ببتيد إلى محور اللولب هي 0.15 نانومتر لـ N و 0.23 نانومتر لـ C و 0.17 نانومتر لـ C k. يوجد أي تشوه بشرط أن تكون هناك عوامل تعمل على استقراره. في حالة اللولب ، تكون هذه العوامل هي الروابط الهيدروجينية التي تتكون من كل ذرة كربونيل من الجزء (i + 4). عامل مهم في استقرار الحلزون α هو أيضًا الاتجاه الموازي للحظات ثنائية القطب للروابط الببتيدية.
هياكل البروتين المطوية. أحد الأمثلة الشائعة على التركيب الدوري المطوي للبروتين هو ما يسمى. طيات، تتكون من جزأين ، كل منهما يمثله بولي ببتيد.
يتم تثبيت الطيات أيضًا بواسطة روابط هيدروجينية بين ذرة الهيدروجين لمجموعة الأمين لجزء واحد وذرة الأكسجين لمجموعة الكربوكسيل لجزء آخر. في هذه الحالة ، يمكن أن يكون للأجزاء اتجاه متوازي ومضاد للتوازي بالنسبة لبعضها البعض.
الهيكل الناتج عن مثل هذه التفاعلات هو هيكل مموج. هذا يؤثر على قيم زوايا الالتواء و. إذا كانت في هيكل مسطح وممتد بالكامل ، فيجب أن تكون 180 ، ثم في طبقات الحقيقية يكون لها القيم = - 119 و = +113. من أجل وضع قسمين من سلسلة البولي ببتيد في اتجاه يفضل تشكيل طيات α ، قسم له هيكل يختلف بشكل حاد عن الهيكل الدوري.
4.2.1 العوامل التي تؤثر على تكوين الهيكل الثانوي
يعتمد هيكل قسم معين من سلسلة البولي ببتيد بشكل أساسي على بنية الجزيء ككل. تتنوع العوامل التي تؤثر على تكوين المناطق ذات البنية الثانوية بشكل كبير ولم يتم تحديدها بالكامل بأي حال من الأحوال. من المعروف أن عددًا من بقايا الأحماض الأمينية تحدث بشكل تفضيلي في شظايا حلزونية ألفا ، وعدد آخر - في طيات ألفا ، وبعض الأحماض الأمينية - بشكل أساسي في مناطق خالية من بنية دورية. يتم تحديد الهيكل الثانوي إلى حد كبير من خلال الهيكل الأساسي. في بعض الحالات ، يمكن فهم المعنى المادي لمثل هذا الاعتماد من التحليل الفراغي الكيميائي للبنية المكانية. على سبيل المثال ، كما يتضح من الشكل ، لا يتم فقط تجميع الجذور الجانبية لبقايا الأحماض الأمينية المجاورة على طول السلسلة معًا في الحلزون ، ولكن أيضًا بعض أزواج البقايا الموجودة على المنعطفات المجاورة للحلزون ، أولاً وقبل كل شيء ، كل (i + 1) المتبقي مع (i + 4) -th ومع (i + 5) -th. لذلك ، في المواضع (i + 1) و (i + 2) و (i + 1) و (i + 4) و (i + 1) و (i + 5) - نادرًا ما يحدث جذرين ضخمين في وقت واحد ، مثل مثل ، على سبيل المثال ، الجذور الجانبية للتيروزين ، التربتوفان ، آيسولوسين. حتى أقل توافقًا مع هيكل اللولب هو التواجد المتزامن لثلاثة مخلفات ضخمة في المواضع (i + 1) و (i + 2) و (i + 5) أو (i + 1) و (i + 4) و (i + 5). لذلك ، فإن مثل هذه التوليفات من الأحماض الأمينية في شظايا حلزونية ألفا هي استثناءات نادرة.
4.3 الهيكل الثالث
يشير هذا المصطلح إلى الطي الكامل في الفضاء لسلسلة البولي ببتيد بأكملها ، بما في ذلك طي الجذور الجانبية. يتم إعطاء صورة كاملة للبنية الثلاثية من خلال إحداثيات جميع ذرات البروتين. بفضل النجاح الهائل لتحليل حيود الأشعة السينية ، تم الحصول على هذه البيانات ، باستثناء إحداثيات ذرات الهيدروجين ، لعدد كبير من البروتينات. هذه كميات هائلة من المعلومات المخزنة في بنوك بيانات خاصة على وسائط يمكن للآلة قراءتها ، ومعالجتها لا يمكن تصورها دون استخدام أجهزة كمبيوتر عالية السرعة. توفر الإحداثيات الذرية التي تم الحصول عليها على أجهزة الكمبيوتر معلومات كاملة حول هندسة سلسلة البولي ببتيد ، بما في ذلك قيم زوايا الالتواء ، مما يجعل من الممكن الكشف عن بنية حلزونية أو طيات أو أجزاء غير منتظمة. مثال على مثل هذا النهج البحثي هو النموذج المكاني التالي لهيكل إنزيم فوسفوجليسيرات كيناز:
المخطط العام لهيكل كيناز الفوسفوجليسيرات. من أجل الوضوح ، يتم تقديم المقاطع الحلزونية α على شكل أسطوانات ، ويتم تقديم طيات α كشرائط مع سهم يشير إلى اتجاه السلسلة من الطرف N إلى الطرف C. الخطوط هي أقسام غير منتظمة تربط الأجزاء الهيكلية.
إن صورة التركيب الكامل حتى لجزيء بروتين صغير على مستوى ما ، سواء كانت صفحة من كتاب أو شاشة عرض ، ليست مفيدة للغاية بسبب التركيب المعقد للغاية للكائن. لكي يتمكن الباحث من تصور التركيب المكاني لجزيئات المواد المعقدة ، فإنه يستخدم طرق رسومات الكمبيوتر ثلاثية الأبعاد ، والتي تسمح بعرض الأجزاء الفردية للجزيئات والتلاعب بها ، على وجه الخصوص ، تحويلها في الزوايا الصحيحة.
يتكون الهيكل الثالث نتيجة للتفاعلات غير التساهمية (قوى الكهروستاتيكية ، الأيونية ، قوى فان دير فال ، وما إلى ذلك) للجذور الجانبية التي تؤطر حلزونات ألفا والطيات والأجزاء غير الدورية من سلسلة البولي ببتيد. من بين السندات التي تحمل الهيكل الثالث ، تجدر الإشارة إلى:
أ) جسر ثاني كبريتيد (- S - S -)
ب) جسر استر (بين مجموعة الكربوكسيل ومجموعة الهيدروكسيل)
ج) جسر الملح (بين مجموعة الكربوكسيل والمجموعة الأمينية)
د) روابط الهيدروجين.
وفقًا لشكل جزيء البروتين بسبب البنية الثلاثية ، يتم تمييز مجموعات البروتينات التالية:
بروتينات كروية. يمكن تمثيل التركيب المكاني لهذه البروتينات بتقريب تقريبي على شكل كرة أو شكل إهليلجي غير ممدود للغاية - الكرة الأرضيةفيلاي. كقاعدة عامة ، يشكل جزء كبير من سلسلة البولي ببتيد لهذه البروتينات β- حلزونات وطيات β. يمكن أن تكون النسبة بينهما مختلفة جدًا. على سبيل المثال ، في الميوغلوبين(المزيد حول هذا الموضوع في الصفحة 28) هناك 5 مقاطع حلزونية وليست طية واحدة. في الجلوبولينات المناعية (مزيد من التفاصيل في الصفحة 42) ، على العكس من ذلك ، فإن العناصر الرئيسية للبنية الثانوية هي - طيات ، و - القفزات غائبة تمامًا. في الهيكل أعلاه من فوسفوجليسيرات كيناز ، يتم تمثيل كلا النوعين من الهياكل متماثلًا تقريبًا. في بعض الحالات ، كما يمكن رؤيته في مثال فوسفوجليسيرات كيناز ، يمكن رؤية جزأين أو أكثر بشكل واضح في الفضاء (ولكن مع ذلك ، بالطبع ، متصلة بواسطة جسور الببتيد) - المجالات.في كثير من الأحيان ، يتم فصل المناطق الوظيفية المختلفة للبروتين إلى مجالات مختلفة.
بروتينات ليفية. هذه البروتينات لها شكل خيطي ممدود ؛ تؤدي وظيفة هيكلية في الجسم. في الهيكل الأساسي ، لديهم أقسام متكررة ويشكلون بنية ثانوية موحدة إلى حد ما لسلسلة البولي ببتيد بأكملها. وهكذا ، فإن البروتين - الكرياتين (المكون الرئيسي للبروتين في الأظافر والشعر والجلد) يُبنى من اللوالب الممتدة. يتكون الحرير الليفي من شظايا متكررة بشكل دوري Gly - Ala - Gly - Ser ، مكونة طيات. هناك عناصر أقل شيوعًا في البنية الثانوية ، على سبيل المثال ، سلاسل بولي ببتيد الكولاجين التي تتشكل اليسار اللوالبمع معلمات مختلفة بشكل حاد عن تلك الموجودة في القفزات. في ألياف الكولاجين ، يتم لف ثلاث سلاسل حلزونية متعددة الببتيد في ملف سوبير أيمن واحد:
4.4 هيكل رباعي
في معظم الحالات ، من أجل عمل البروتينات ، من الضروري دمج العديد من سلاسل البوليمر في مجمع واحد. يعتبر هذا المركب أيضًا بروتينًا يتكون من عدة الوحدات الفرعية. غالبًا ما يظهر هيكل الوحدة الفرعية في الأدبيات العلمية كهيكل رباعي.
يتم توزيع البروتينات المكونة من عدة وحدات فرعية على نطاق واسع في الطبيعة. المثال الكلاسيكي هو التركيب الرباعي للهيموجلوبين (مزيد من التفاصيل - ص 26). عادة ما يتم الإشارة إلى الوحدات الفرعية بأحرف يونانية. يحتوي الهيموغلوبين على وحدتين فرعيتين. إن وجود العديد من الوحدات الفرعية مهم وظيفيًا - فهو يزيد من درجة تشبع الأكسجين. تم تحديد التركيب الرباعي للهيموجلوبين على أنه 2 2.
تتميز بنية الوحدة الفرعية بالعديد من الإنزيمات ، وخاصة تلك التي تؤدي وظائف معقدة. على سبيل المثال ، RNA polymerase من ه. القولونيةله بنية وحدة فرعية 2 "، أي أنه مبني من أربعة أنواع مختلفة من الوحدات الفرعية ، ويتم تكرار الوحدة الفرعية. يؤدي هذا البروتين وظائف معقدة ومتنوعة - يبدأ الحمض النووي ، ويربط الركائز - ثلاثي فوسفات الريبونوكليوزيد ، وينقل أيضًا بقايا النوكليوتيدات إلى نمو سلسلة polyribonucleotide وبعض الوظائف الأخرى.
عمل العديد من البروتينات يخضع لما يسمى. تفارغية- المركبات الخاصة (المؤثرات) "إيقاف" أو "تشغيل" عمل المركز النشط للإنزيم. هذه الإنزيمات لها مواقع خاصة للتعرف على المستجيب. وهناك حتى خاص الوحدات الفرعية التنظيمية، والتي تشمل ، من بين أشياء أخرى ، الأقسام المشار إليها. المثال الكلاسيكي هو إنزيمات بروتين كيناز التي تحفز نقل بقايا حمض الفوسفوريك من جزيء ATP إلى بروتينات الركيزة.
الفصل 5. الخصائص
البروتينات لها وزن جزيئي مرتفع ، بعضها قابل للذوبان في الماء ، وقادر على الانتفاخ ، ويتميز بالنشاط البصري ، والتنقل في مجال كهربائي ، وبعض الخصائص الأخرى.
تشارك البروتينات بنشاط في التفاعلات الكيميائية. ترجع هذه الخاصية إلى حقيقة أن الأحماض الأمينية التي تتكون منها البروتينات تحتوي على مجموعات وظيفية مختلفة يمكن أن تتفاعل مع مواد أخرى. من المهم أن تحدث مثل هذه التفاعلات أيضًا داخل جزيء البروتين ، مما يؤدي إلى تكوين الببتيد وثاني كبريتيد الهيدروجين وأنواع أخرى من الروابط. يمكن أن ترتبط المركبات والأيونات المختلفة بجذور الأحماض الأمينية ، وبالتالي البروتينات ، مما يضمن نقلها عبر الدم.
البروتينات هي مركبات جزيئية كبيرة. هذه عبارة عن بوليمرات تتكون من مئات وآلاف من بقايا الأحماض الأمينية - مونومرات. تبعا لذلك و الكتلة الجزيئيةتتراوح البروتينات بين 10000 و 1000000. لذلك ، يحتوي الريبونوكلياز (إنزيم يفكك الحمض النووي الريبي) على 124 بقايا من الأحماض الأمينية ويبلغ وزنها الجزيئي 14000 تقريبًا. يحتوي الميوغلوبين (بروتين عضلي) ، الذي يتكون من 153 من بقايا الأحماض الأمينية ، على جزيئات الوزن 17000 ، والهيموغلوبين - 64500 (574 من مخلفات الأحماض الأمينية). الأوزان الجزيئية للبروتينات الأخرى أعلى: - يتكون الجلوبيولين (يشكل أجسامًا مضادة) من 1250 من الأحماض الأمينية ويبلغ وزنه الجزيئي حوالي 150.000 ، ويزيد الوزن الجزيئي لإنزيم نازعة هيدروجين الجلوتامات عن 1.000.000.
يتم تحديد الوزن الجزيئي من خلال طرق مختلفة: مقياس التناضح ، الترشيح الهلامي ، البصري ، إلخ. ومع ذلك ، فإن الطريقة الأكثر دقة هي طريقة الترسيب التي اقترحها T. Svedberg. يعتمد على حقيقة أنه أثناء التنبيذ الفائق مع تسارع يصل إلى 900000 جم ، يعتمد معدل ترسيب البروتين على وزنه الجزيئي.
أهم خاصية للبروتينات هي قدرتها على إظهار كل من الحمضية والقاعدية ، أي أنها تعمل مذبذبالشوارد. يتم ضمان ذلك من خلال مجموعات منفصلة مختلفة تشكل جذور الأحماض الأمينية. على سبيل المثال ، يتم نقل الخصائص الحمضية للبروتين بواسطة مجموعات الكربوكسيل من حمض الجلوتاميك الأميني الأسبارتي ، ويتم نقل الخصائص القلوية بواسطة جذور الأرجينين والليسين والهيستيدين. كلما زاد عدد الأحماض الأمينية ثنائية الكربوكسيل التي يحتوي عليها البروتين ، زادت خصائصه الحمضية والعكس صحيح.
تحتوي هذه المجموعات أيضًا على شحنات كهربائية تشكل الشحنة الكلية لجزيء البروتين. في البروتينات التي تسود فيها الأحماض الأمينية الأسبارتيك والجلوتامين ، تكون شحنة البروتين سالبة ؛ ففائض الأحماض الأمينية الأساسية يعطي شحنة موجبة لجزيء البروتين. نتيجة لذلك ، في مجال كهربائي ، تتحرك البروتينات نحو القطب السالب أو الأنود ، اعتمادًا على حجم شحنتها الإجمالية. لذلك ، في البيئة القلوية (الرقم الهيدروجيني 7-14) ، يتبرع البروتين بالبروتون ويصبح سالبًا ، بينما في البيئة الحمضية (الرقم الهيدروجيني 1-7) ، يتم قمع تفكك المجموعات الحمضية ويصبح البروتين كاتيونًا.
وبالتالي ، فإن العامل الذي يحدد سلوك البروتين ككاتيون أو أنيون هو تفاعل الوسط ، والذي يتم تحديده بواسطة تركيز أيونات الهيدروجين ويتم التعبير عنه بقيمة الأس الهيدروجيني. ومع ذلك ، عند بعض قيم الأس الهيدروجيني ، يتساوى عدد الشحنات الموجبة والسالبة ويصبح الجزيء محايدًا كهربائيًا ، أي لن تتحرك في مجال كهربائي. تُعرَّف قيمة الأس الهيدروجيني للوسط بأنها نقطة تساوي الكهرباء للبروتينات. في هذه الحالة ، يكون البروتين في أقل حالة استقرارًا ، ومع تغيرات طفيفة في درجة الحموضة إلى الجانب الحمضي أو القلوي ، فإنه يترسب بسهولة. بالنسبة لمعظم البروتينات الطبيعية ، تكون النقطة الكهربية في بيئة حمضية قليلاً (الرقم الهيدروجيني 4.8 - 5.4) ، مما يشير إلى غلبة الأحماض الأمينية ثنائية الكربوكسيل في تركيبها.
تكمن الخاصية المتذبذبة في خواص التخزين المؤقت للبروتينات ومشاركتها في تنظيم درجة الحموضة في الدم. قيمة الرقم الهيدروجيني لدم الإنسان ثابتة وتتراوح بين 7.36 - 7.4 ، على الرغم من المواد المختلفة ذات الطبيعة الحمضية أو الأساسية ، والتي يتم تزويدها بانتظام بالطعام أو تتشكل في عمليات التمثيل الغذائي - لذلك ، هناك آليات خاصة لتنظيم القاعدة الحمضية توازن البيئة الداخلية للجسم. وتشمل هذه الأنظمة التي تم النظر فيها في الفصل. نظام عازلة الهيموجلوبين "التصنيف" (الصفحة 28). يشير التغيير في درجة الحموضة في الدم بأكثر من 0.07 إلى تطور عملية مرضية. يسمى التحول في درجة الحموضة إلى الجانب الحمضي بالحماض ، ويسمى الجانب القلوي القلاء.
من الأهمية بمكان بالنسبة للجسم قدرة البروتينات على امتصاص بعض المواد والأيونات على سطحها (الهرمونات والفيتامينات والحديد والنحاس) ، والتي تكون إما ضعيفة الذوبان في الماء أو سامة (البيليروبين والأحماض الدهنية الحرة). تنقلها البروتينات عبر الدم إلى أماكن أخرى من التحولات أو التحييد.
المحاليل المائية للبروتينات لها خصائصها الخاصة. أولاً ، البروتينات لها انجذاب كبير للماء ، أي أنهم محبة للماء.هذا يعني أن جزيئات البروتين ، مثل الجسيمات المشحونة ، تجذب ثنائيات أقطاب الماء ، والتي توجد حول جزيء البروتين وتشكل قشرة مائية أو قشرة هيدراتية. تحمي هذه القشرة جزيئات البروتين من الالتصاق ببعضها البعض والترسيب. يعتمد حجم قشرة الماء على بنية البروتين. على سبيل المثال ، ترتبط الألبومات بسهولة بجزيئات الماء ولها قشرة مائية كبيرة نسبيًا ، في حين أن الجلوبيولين والفيبرينوجين يعلقان الماء بشكل أسوأ ، وتكون قشرة الماء أصغر. وبالتالي ، فإن استقرار المحلول المائي للبروتين يتحدد بعاملين: وجود شحنة على جزيء البروتين وقشرة الماء المحيطة به. عندما تتم إزالة هذه العوامل ، يترسب البروتين. يمكن أن تكون هذه العملية قابلة للعكس ولا رجعة فيها.
...وثائق مماثلة
البروتينات (البروتينات) هي مواد عضوية طبيعية عالية الجزيئات تحتوي على النيتروجين ، جزيئاتها مبنية من الأحماض الأمينية. هيكل البروتينات. تصنيف البروتينات. الخصائص الفيزيائية والكيميائية للبروتينات. الوظائف البيولوجية للبروتينات. إنزيم.
الملخص ، تمت الإضافة 05/15/2007
الملامح الرئيسية لعمليات التمثيل الغذائي. التمثيل الغذائي والطاقة. الخصائص العامة والتصنيف والوظائف والتركيب الكيميائي وخصائص البروتينات ودورها البيولوجي في تكوين المادة الحية. البروتينات الهيكلية والمعقدة. طرق هطول الأمطار.
عرض تقديمي ، تمت الإضافة بتاريخ 04/24/2013
الخصائص الفيزيائية والكيميائية ، التفاعلات اللونية للبروتينات. تكوين وهيكل وظائف البروتينات في الخلية. مستويات بنية البروتين. التحلل المائي للبروتينات ودورها في النقل والحماية. البروتين كمادة بناء للخلية ، قيمته الطاقية.
الملخص ، تمت الإضافة 06/18/2010
الخصائص الفيزيائية والبيولوجية والكيميائية للبروتينات. توليف وتحليل البروتينات. تحديد البنية الأولية والثانوية والثالثية والرباعية للبروتينات. تمسخ وعزل وتنقية البروتينات. استخدام البروتينات في الصناعة والطب.
الملخص ، تمت إضافته في 06/10/2015
البروتينات - المركبات العضوية ذات الوزن الجزيئي العالي ، تكوينها من الأحماض الأمينية. تحديد خصائص البروتينات من خلال تكوينها وهيكل جزيء البروتين. توصيف الوظائف الرئيسية للبروتينات. عضيات الخلية ووظائفها. التنفس الخلوي وهيكله.
الاختبار ، تمت إضافة 2012/06/24
مفهوم وهيكل البروتينات والأحماض الأمينية كمونومراتها. تصنيف وأنواع الأحماض الأمينية ، طبيعة الرابطة الببتيدية. مستويات تنظيم جزيء البروتين. الخصائص الكيميائية والفيزيائية للبروتينات وطرق تحليلها ووظائفها.
عرض تقديمي ، تمت إضافة 2014/04/14
الدور البيولوجي للمياه. وظائف الأملاح المعدنية. دهون بسيطة ومعقدة. مستويات تنظيم البروتينات. وظائف البناء والطاقة والتخزين والتنظيم للدهون. الوظائف الهيكلية ، التحفيزية ، الحركية ، النقل للبروتينات.
عرض تقديمي ، تمت الإضافة 21/05/2015
تكوين الأحماض الأمينية للبروتينات في الكائنات الحية ، دور الكود الجيني. مجموعات من 20 حمض أميني قياسي. فصل البروتينات إلى فئة منفصلة من الجزيئات البيولوجية. البروتينات المحبة للماء والطارئة للماء. مبدأ بناء البروتينات ، مستوى تنظيمها.
عمل إبداعي تمت الإضافة في 11/08/2009
العناصر الأساسية والتركيب الكيميائي للأنسجة العضلية. أنواع بروتينات الساركوبلازم واللييفات العضلية ، ومحتواها من العدد الإجمالي للبروتينات ، والوزن الجزيئي ، والتوزيع في العناصر الهيكلية للعضلة. وظائفهم ودورهم في الجسم. هيكل جزيء الميوسين.
عرض تقديمي ، تمت إضافة 12/14/2014
البروتينات كمصادر للغذاء ، وظائفها الرئيسية. تشارك الأحماض الأمينية في صنع البروتينات. هيكل سلسلة البولي ببتيد. تحول البروتينات في الجسم. بروتينات كاملة وغير مكتملة. تركيب البروتين ، الخواص الكيميائية ، التفاعلات النوعية.
من المعروف أن المادة الحية تعتمد على المواد العضوية - البروتينات والدهون والكربوهيدرات والأحماض النووية. لكن أهم هذه المواد هو البروتين.
تتغير معظم المواد المعروفة للعلم من صلبة إلى سائلة عند تسخينها. ولكن هناك مواد ، على العكس من ذلك ، تتحول إلى حالة صلبة عند تسخينها. تم دمج هذه المواد في فئة منفصلة بواسطة الكيميائي الفرنسي بيير جوزيف ماكي في عام 1777. وبالمقارنة مع بياض البيض ، الذي يتخثر عند تسخينه ، سميت هذه المواد بالبروتينات. تُعرف البروتينات أيضًا بالبروتينات. البروتين اليوناني يعني "في المقام الأول". أُطلق هذا الاسم على البروتين في عام 1838 ، عندما كتب عالم الكيمياء الحيوية الهولندي جيرارد مولدر أن الحياة على هذا الكوكب ستكون مستحيلة بدون مادة معينة ، وهي أهم المواد المعروفة للعلم والتي توجد بالضرورة في جميع النباتات تمامًا. والحيوانات. دعا مولدر هذه المادة بروتين.
البروتين هو المادة الأكثر تعقيدًا بين جميع العناصر الغذائية. في كل خلية من خلايا جسم الإنسان ، تحدث تفاعلات كيميائية يلعب فيها البروتين دورًا مهمًا للغاية.
مما يتكون البروتين

تشمل البروتينات: النيتروجين والأكسجين والهيدروجين والكربون. لكن العناصر الغذائية الأخرى لا تحتوي على النيتروجين.
البروتين هو بوليمر طبيعي. البوليمرات هي مواد تحتوي جزيئاتها على عدد كبير جدًا من الذرات. في القرن التاسع عشر ، أثبت الكيميائي الروسي ألكسندر ميخائيلوفيتش بتلروف أنه إذا تغيرت بنية الجزيء ، فإن خصائص المادة تتغير أيضًا. اللبنات الأساسية للبروتينات هي الأحماض الأمينية. وفي البروتينات توجد تركيبات مختلفة من الأحماض الأمينية. لذلك ، يوجد في الطبيعة مجموعة متنوعة من البروتينات ذات الخصائص المختلفة. بمساعدة البحث ، تم اكتشاف ما يقرب من 20 من الأحماض الأمينية التي تشارك في تكوين البروتينات.
كيف هي عملية تكوين جزيء البروتين

ترتبط الأحماض الأمينية ببعضها البعض بالتتابع. نتيجة لهذه العملية ، يتم تشكيل سلسلة تسمى بولي ببتيد. بعد ذلك ، قد تلتف البولي ببتيدات أو تأخذ شكلًا آخر. تعتمد خصائص البروتين على تكوين الأحماض الأمينية ، وعلى عدد الأحماض الأمينية المشاركة في التركيب ، وبأي ترتيب ترتبط هذه الأحماض الأمينية ببعضها البعض. على سبيل المثال ، يتضمن تركيب بروتينين نفس عدد الأحماض الأمينية ، والتي لها أيضًا نفس التركيب. ولكن إذا كانت هذه الأحماض الأمينية موجودة في تسلسل مختلف ، فسنحصل على بروتينين مختلفين تمامًا.
إذا كانت الببتيدات لا تحتوي على أكثر من 15 من بقايا الأحماض الأمينية ، فيُطلق عليها اسم oligopeptides. وتسمى الببتيدات التي تحتوي على ما يصل إلى عدة عشرات الآلاف أو حتى مئات الآلاف من بقايا الأحماض الأمينية بالبروتينات. جزيء البروتين له هيكل مكاني مضغوط. قد يكون هذا الهيكل في شكل ألياف. تسمى هذه البروتينات الليفية. إنهم يبنون البروتينات. إذا كان جزيء البروتين له بنية على شكل كرة ، فإن البروتينات تسمى كروية. تشمل هذه البروتينات الإنزيمات والأجسام المضادة وبعض الهرمونات.
اعتمادًا على الأحماض الأمينية المتضمنة في تكوين البروتينات ، تكون البروتينات كاملة وغير كاملة. تحتوي البروتينات الكاملة على مجموعة كاملة من الأحماض الأمينية. بروتينات غير مكتملة ، بعض الأحماض الأمينية مفقودة.
تنقسم البروتينات أيضًا إلى بسيطة ومعقدة. تحتوي البروتينات البسيطة على أحماض أمينية فقط. يشمل تكوين البروتينات المعقدة ، بالإضافة إلى الأحماض الأمينية ، المعادن والكربوهيدرات والدهون والأحماض النووية.
دور البروتينات في جسم الإنسان

تؤدي البروتينات وظائف مختلفة في جسم الإنسان.
1.الهيكلي. البروتينات هي جزء من خلايا جميع الأنسجة والأعضاء.
2. محمي. يتم تصنيع بروتين الإنترفيرون في الجسم للحماية من الفيروسات.
3. Dvigatelnaأنا. يشارك بروتين الميوسين في عملية تقلص العضلات.
4. المواصلات.الهيموغلوبين ، وهو بروتين في تكوين خلايا الدم الحمراء ، يشارك في نقل الأكسجين وثاني أكسيد الكربون.
5. طاقةأنا. نتيجة لأكسدة جزيئات البروتين ، يتم إطلاق الطاقة اللازمة لحياة الجسم.
6. المحفزأنا. تعمل إنزيمات البروتين كمحفزات بيولوجية تزيد من معدل التفاعلات الكيميائية في الخلايا.
7. تنظيميةأنا. تنظم الهرمونات وظائف الجسم المختلفة. على سبيل المثال ، الأنسولين ينظم مستويات السكر في الدم.
يوجد في الطبيعة عدد كبير من البروتينات التي يمكنها أداء مجموعة متنوعة من الوظائف. لكن أهم وظيفة للبروتينات هي الحفاظ على الحياة على الأرض مع الجزيئات الحيوية الأخرى.
 الخطابة كنموذج أولي للصحافة
الخطابة كنموذج أولي للصحافة اقتباسات عن Napoleon - dslinkov - LiveJournal
اقتباسات عن Napoleon - dslinkov - LiveJournal لي الانتقام دمر الرجل على الجرافة المدينة
لي الانتقام دمر الرجل على الجرافة المدينة