Червоні водорості. Відділ червоних водоростей – rhodophyta Місце в ланцюгах харчування, вплив на природу
Біологічний сенс виникнення суцвіть – у зростаючій ймовірності запилення квіток як ентомофільних, і анемофільних рослин. Безперечно, що комаха за одиницю часу відвідає набагато більше квіток, якщо вони зібрані в суцвіття. Крім того, квітки, зібрані в суцвіття, більш помітні серед зелені листя, не жили поодинокі квіти. Багато поникають суцвіття легко розгойдуються під впливом руху повітря, сприяючи тим самим розсіювання пилку.
Суцвіття властиві величезній більшості квіток рослин. Зазвичай суцвіття групуються біля верхньої частини рослини на кінцях гілок, але іноді, особливо у тропічних дерев, виникають на стовбурах та товстих гілках. Таке явище відоме під назвою кауліфлорії. Як приклад можна навести шоколадне дерево. Вважається, що в умовах тропічного лісу кауліфлорія робить квіти доступнішими для комах – запилювачів. Інший приклад кауліфлорії – у широко культивованого в Криму та на Кавказі бобової рослини церциса ріжкового.
Суцвіття має головну вісь, або вісь суцвіття, і бічні осі, які можуть бути розгалужені різною мірою або нерозгалужені. Кінцеві їх відгалуження – квітконіжки – несуть квіти.
У простихсуцвіть бокові осі не розгалужені і є квітконіжками. Суцвіття, у яких бічні осі гілкуються, називаються складними. У складного суцвіття можуть бути бічні осі першого, другого та наступного порядків.
Головна і всі кінцеві бічні осі можуть закінчуватися верхівковими квітками, внаслідок чого їхнє зростання обмежене. Такі суцвіття отримали назву закритих ( симподіальних), чи певних. У закритих суцвіть верхівкові квітки зазвичай розкриваються раніше бічних, і тому їх називають верхоцвітими.
У відкритих суцвіть зростання головної осі необмежене (тобто відкритий), а квітки розташовуються збоку від морфологічної верхівки квітконосного втечі. Це може стосуватися і бічних осях. Такі суцвіття називають відкритими ( моноподіальними), або невизначеними. У відкритих суцвіть квітки розпускаються послідовно знизу нагору, тому їх називають бокоцвітими.
2. Монопідіальні, або ботріодні суцвіття.
Моноподіальні (відкриті, або бокоцвіті) суцвіття мають головну вісь, яка продовжує весь час зростання, а бічні осі несуть на собі квіти. У такому суцвітті спочатку розцвітають нижні квіти, а згодом – послідовно верхні. Розрізняють прості та складні ботріоїдні суцвіття
Прості ботріоїдні суцвіття
К і с т ь - на головній осі, або осі першого порядку по черзі розташовуються квітконіжки, які несуть дрібні квітки (конвалія, черемха, наперстянка);
Сережки – головна вісь суцвіття, на відміну від кисті звисає (береза);
Колос – на головній подовженій осі знаходяться сидячі квітки (подорожник);
По ч а т о к – головна вісь суцвіття відрізняється великою м'ясистістю, розташування квіток як у колосу (кукурудза);
Щиток – квітки розташовуються в одній площині, у нижніх квіток довші квітконіжки, ніж у верхніх (груша, яблуня, зливу);
З о н т і к – квітконіжки окремих квіток мають однакову довжину та відходять від верхньої частини осі, як спиці парасольки (цибуля, вишня);
Корзинка – численні сидячі квітки розташовані на укороченій і блюдцеподібно потовщеній осі. Зовні знаходиться обгортка з прикольного листя (соняшник, ромашка);
Головка – на укороченій і розширеній головній осі сидять тісно зближені квітки (конюшина, кровохлібка).
Складні ботріоїдні суцвіття
Вони характеризуються тим, що квіти розташовуються на осях 2-го, 3-го і т. д. порядків.
1. Волотко - на бічних осях знаходяться прості кисті (бузок, виноград).
2. Складний колос - на головній осі розташовуються прості колоски (пшениця, жито, ячмінь).
3. Складна парасолька – на бічних осях знаходяться прості парасольки. Прості парасольки зазвичай мають свої приквітнички (приватні обгортки), а приквітники біля основи парасольки утворюють загальну обгортку (кріп, морква, аніс).
4. Складний щиток (щитковидна волотка) - на бічних осях знаходяться маленькі суцвіття-щитки (горобина).
У деяких випадках спостерігаються змішані суцвіття. Наприклад, у вівса - суцвіття волоті, але замість квіток є маленькі колоски, так що змішані два суцвіття: волота і колос.
3. Симподіальні, або цимоїдні суцвіття. Морфологія плодів.
Симподиальними (цимоїдними) називаються такі суцвіття, у яких загальний квітконос (головна вісь) закінчується квіткою, а його зростання продовжує верхній бічний, або бічні пагони, які в свою чергу таким же чином продовжують свій зріст. До цимоїдних суцвіть відносяться такі:
1. Розвиліну, або дихазій. Головна вісь закінчується квіткою, зростання квітконоса триває супротивними бічними нижчими нирками, що утворюють квітки, і так далі (гвоздика, лялька, мильнянка).
2. Плейохазій, або хибна парасолька, - зростання квітконоса закінчується квіткою і продовжується мутовчато нижчими боковими нирками, що утворюють квітки (молочай).
3. Завиток - ріст квітконоса закінчується квіткою і триває бічною ниркою, що утворює квітку, і так неодноразово, причому всі квітки направлені в один бік (картопля, незабудка, живокіст).
Морфологія плодів.
Після запліднення стінки зав'язі маточки розвиваються в околоплодник (перикарпій). Навколоплодником називаються стінки плода, що оточують насіння, які утворюються одночасно з навколоплідником. Навколоплодник зазвичай складається з трьох шарів:
1) зовнішнього ( екзокарпія), покритого іноді різними виростами (крилатка у клена, причіпки у реп'яха);
2) середнього ( мезокарпія), що становить у соковитих плодів м'якоть, що містить багато цукру (сливу, черешня) або олію (олив);
3) внутрішнього ( ендокарпія),що перетворюється у соковитих плодів часто на шар
кам'янистих клітин (кісточка сливи, вишні) або в соковиту м'якоть (лимон).
Всі три зони добре помітні. Наприклад, у плоді вишні тонкий шкірястий зовнішній шар – екзокарпій, їстівна соковита м'якоть плода – мезокарпій, тверда кісточка з кам'янистої тканини, що оточує єдине насіння – ендокарпій.
Незрілі плоди зазвичай зеленого кольору, клітини екзокарпія та мезокарпія містять хлорофіл та фотосинтезують. У міру дозрівання плід втрачає зелений колір і часто набуває яскравого забарвлення, що обумовлюється або антоціаном у клітинному соку (у вишні, винограду, чорниці), або хромопластами (горобина, томат).
У деяких видів рослин плід утворюється не тільки із зав'язі маточки, але і з квітколожа або оцвітини. Такі плоди називаються хибними. Наприклад, ягодоподібний хибний плід суниці є соковитим забарвленим квітколожем, на якому знаходяться дрібні сухі справжні плодики (насіння), що утворилися із зав'язей пестиків. З квітлоложа, що розрісся, утворюються плоди шипшини, яблуні, груші.
У деяких видів рослин плід утворюється з кількох маточок. Такий плід називається збірним, або складним (збірний плід малини, ожини, жовтця).
4. Класифікація плодів.Вся різноманітність плодів поділяють на такі типи.
Сухі плоди, що не розкриваються.
1. Горіх, або горіх, має дерев'янистий навколоплідник, в якому знаходиться одне вільно лежаче насіння. Наприклад, у ліщини, дуба, бука, конопель.
2. Насіння має шкірястий навколоплідник, в якому знаходиться одне вільно лежаче насіння. Наприклад, у соняшника, кульбаби, ромашки та інших видів із сімейства складноцвітих.
3. Зернівка має шкірястий оплодник, в якому знаходиться одне зрощене з оплоднем насіння. Наприклад, у пшениці, ячменю та інших видів із сімейства злаків.
4. Крилатка має навколоплідник горіхоподібного плі сім'янкоподібного типу, що розрісся в один або кілька крилатих придатків. Наприклад, у клена, берези, в'яза, ясена.
5. У деяких видів сухий плід, що не розкривається, має летучку – чубчик з волосків. Наприклад, у кульбаби.
Сухі плоди, що розкриваються
1. Листівка – коробчатий одногніздний, багатонасінний плід, утворений одним плодолистком, що розкривається по черевному шву від вершини до основи.
Наприклад, у ваточника, видів сімейства лютикових та сімейства розоцвітих. У півонії утворюється багато листівок з маточок однієї квітки.
2. Боб – коробчатий одногніздний, частіше багатонасінний плід, утворений одним плодолистиком, що розкривається по черевному та спинному швам від вершини до основи. Наприклад, у гороху та інших видів сімейства бобових. У деяких видів при розкриття стулок боба вони, спірально закручуючи, розкидають насіння (наприклад, у жовтої акації). У деяких видів боби однонасінні (наприклад, у еспарцету) і тоді не розкриваються.
3. Стручок і стручочок – коробчатий двогніздний багатонасінний плід, утворений двома плодолистками. У середині плода зверху до низу проходить плівчаста перегородка, до країв якої прикріплено насіння. Розкривається плід двома стулками по двох швах від основи до вершини. Стулки відпадають, а перегородка з насінням залишається. Стручком називається довгий і вузький плід - довжина більша за ширину в 4 рази і більше (наприклад, у гірчиці), а стручочком - короткий і широкий (наприклад, у грициків).
4. Коробочка - багатогніздний (рідко одногніздний) багатонасінний плід, утворений декількома плодолистками. Розкриваються коробочки по-різному, то стулками (наприклад, у бавовнику, дурману, льону-довгунцю, чаю, лілії, рицини), то зубчиками на вершині коробочки (наприклад, у багатьох видів сімейства гвоздикових), то дірочками (наприклад, у деяких видів маку і дзвіночка), то кришечкою на верхівці коробочки (наприклад, у блекоти), то бічними щілинами (наприклад, у бамії).
У деяких видів сухі багатонасінні плоди (стручки, боби) при дозріванні розпадаються поперечно на однонасінні членики (наприклад, у дикої редьки). Такі плоди називаються членистими.
У інших видів сухі багатонасінні плоди при дозріванні розпадаються поздовжньо на окремі однонасінні членики, наприклад у клена та інших видів сімейства кленових, а також у сімейств зонтичних, губоцвітих, мальвових та деяких інших. Такі плоди називаються дрібними.
Соковиті плоди
1. Ягода – соковитий, зазвичай пофарбований, багатонасінний плід, утворений одним або декількома плодолистками. Насіння в ягоді занурене в соковитої м'якоті. Наприклад, у винограду, смородини, агрусу, беладонни, чорниці, журавлини, картоплі, томату, огірка, кавуна, дині, гарбуза, лимона, мандарина, апельсина (огірок, гарбуз, кавун, диня є хибними ягодами, так як зовнішня у них утворилася з квітколожу).
2. Костянка - зазвичай соковитий і пофарбований, одногніздний, однонасінний або багатонасінний плід, утворений одним або декількома плодолистками. Ендокарпій у кістянки має вигляд кісточки. Наприклад, у сливи, вишні, абрикоса, кизилу, маслини. Багатокісткові кістянки - у бузини, крушини проносної. Плід волоського горіха є кістянкою, але його мезокарпій не соковитий; сам так званий горіх є кісточкою плода. У костянки мигдалю також мезокарпій не соковитий, а у костянки кокосової пальми він волокнистий.
Плодючість покритонасінних рослин дуже висока. Так, одна рослина білої лободи утворює близько 100.000 насінин на рік, блекоти - ≈ 500.000, гулявника ≈ 750.000,
тополі ≈ 27. 000.000 насінин.
5. Супліддя. Поширення плодів та насіння.
У деяких видів плід утворюється із суцвіття. Така освіта називається суплоддям. Наприклад, супліддя у буряків – клубочок, супліддя у шовковиці – «ягода», супліддя у винної ягоди – «ягода». Супліддя у шовковиці є зрощеними хибними плодами, утвореними розквітлими оцвітиною.
У деяких сортів культурних рослин плоди розвиваються без запилення та запліднення і тому без насіння. Це утворення плоду без насіння називається партенокарпією. Партенокарпія спостерігається у деяких сортів яблуні, груші («безнасінка»), винограду (родзинки – сушений виноград без насіння), аґрусу, мандарину, апельсину, лимону, інжиру, японської хурми та ін. У деяких сортів можна викликати партенокарпію за допомогою роздратування (запилюючи грушу пилком яблуні, томат – пилком картоплі, баклажан – пилком томату), деякими хімічними речовинами, укусами комах, припіканням розпеченим дротом та ін. Безнасінні плоди зазвичай дрібніші за плоди з насінням і, отже, вони дають. Проте на практиці споживачем безнасінні плоди високо цінуються. Розмножуватися безнасінні сорти можуть лише вегетативно.
Поширення плодів та насіння
У насіння та плодів покритонасінних рослин спостерігається багато різноманітних пристосувань для поширення. Більшість видів рослин пристосувалася до поширення насіння і плодів у вигляді вітру (анемохорія). Насіння бавовнику, осики, тополі, верби, плоди кульбаби та ін. мають своєрідні довгі волоски та летучки, що сприяють розсіюванню їх вітром часто на далекі відстані. Плоди клена, берези, в'яза, ясена, вільхи, насіння сосни, ялини та ін. мають крилаті вирости (чому і називаються плоди крилатками), що сприяють розсіюванню їх вітром. Так само сприяє розсіюванню вітром лист, що криє, при суцвітті липи. Насіння багатьох видів із сімейств вересових, тирличових, орхідних та ін. настільки дрібні та легкі, що переносяться вітром, як пил, на дуже великі відстані. Рослини верблюжої трави, курая, клопника і ін., що мають кулясту форму, після дозрівання плодів відламуються вітром від підземної частини і котяться по степу на далекі відстані, часто утворюючи цілі вали, що рухаються, розсіюючи по шляху своє насіння. Ці рослини отримали загальну назву «перекотиполе».
Насіння та плоди водних рослин поширюються водою (гідрохорія). Вода дощова (особливо після зливи), вода струмків та річок відіграє значну роль у перенесенні насіння та плодів рослин, що виросли на суші.
Велику роль поширенні насіння грають тварини. Птахи (орнітохорія) та меншою мірою інші тварини (зоохорія) поїдають соковиті плоди. Насіння їх, проходячи через травний тракт тварини, не пошкоджується, не втрачає своєї схожості. Разом з екскрементами тварин насіння потрапляє на землю і проростає.
Таким чином, насіння проростає часто на значній відстані від материнської рослини. У деяких видів рослин (реп'ях, морква, лопух, липучка) насіння і плоди мають різні причіпки плі клейку поверхню і чіпляються за шерсть тварини, що проходить повз, або приклеюються до пір'я птахів і таким чином переносяться. Дрібне насіння поширюється мурахами, а також може прилипати з брудом до копит тварин і таким чином переноситися.
У деяких видів рослин виробилася здатність самостійно розкидати своє насіння (автохорія). Так, у жовтої акації стулки визрілих бобів розкриваються і спірально скручуються з такою силою і швидкістю, що насіння, що є в бобі, розкидається з силою на всі боки. Пояснюється таке розкриття плода у жовтої акації тим, що при дозріванні її боба зовнішні та внутрішні шари його стулок скорочуються неоднаково і між ними виникає і зростає натяг, який і обумовлює скручування та струсіння стулок при розкриванні боба.
У «шаленого» огірка, дозрілий плід якого відламується від плодоніжки, в отвір, що утворився, викидаються разом з рідким вмістом насіння. Це відбувається з такою силою, що насіння зазвичай падає на відстані кількох метрів. Пояснюється такий викидання рідини і насіння тим, що в плоді в міру його дозрівання зростає тиск і на момент дозрівання плода досягає великої напруги.
ТЕМА ЗАНЯТТЯ 27 : С-во лютикові (Ranunculaceae). С-во макові (Papaveracea)
1. Сімейство лютикові. 2. Сімейство макові. З. 379 – 383
Порядок Лютикові.Сімейство лютикові (2.000)
Близько 2000 видів. Види сімейства лютикові поширені у холодних помірних та субтропічних (гірських) районах. На Смоленщині росте 32 види. У тропіках рослини цього сімейства зустрічаються як виняток. Життєві форми – насамперед багаторічні трав'янисті рослини, рідко зустрічаються чагарники, чагарники та ліани.
Листя просте, без прилистків; Форма листової пластини різноманітна. Листя може бути як цілісним (чистяк весняний), так і розсіченими різною мірою (вітрениця дубравна, печіночниця благородна, василист водозбірний). Листорозташування чергове.
У багатьох лютикових є видозмінені підземні пагони (кореневище біля вітряниці і шишки у чистяка весняного).
Квітки можуть бути одиночними (рід вітряниця) і зібраними в суцвіття (роди жовтець, водозбір, василист). Лютикові винятково різноманітні за будовою квіток.
Різноманітність у будові квіток можна висловити, простеживши кілька еволюційних рядів.
1. Квітки з невизначеним числом членів (у вітряниці дібровний простий оцвітина може мати 5-8 листків) → квітки з певним чітко встановленим числом членів (рід борець).
2. Оцвітина проста (рід вітряниця, рід водозбір) → оцвітина подвійна (рід жовтець)
3. плодолистків від великої кількості до одного
4. гінець від апокарпного до синкарпного
5. квіти від актиноморфних квіток до зигоморфних (рід борець, рід аконіт)
6. вісь квітки подовжена, тому члени квітки розташовуються спірально → вісь квітки майже плоска, тому члени квітки розташовуються по колу.
7. Квітки неспеціалізовані, ентомофільні (часто запилюються мухами) → Квітки спеціалізовані, ентомофільні (рід борець, рід живокість).
Для багатьох видів лютикових характерні стамінодії– сильно редуковані та видозмінені пелюстки (або тичинки), які перетворюються на нектарники. Нектарніков немає тільки у василистника.
Пестиков у більшості видів багато, вони вільні і розташовуються на подовженій осі (роди жовтець, печіночниця, калюжниця). Рідше кількість маточок обмежена: 3-1. Зав'язь завжди одногніздна та верхня. Насіння зачатків зазвичай багато, але є види, які мають у складі маточок 1 сім'язачаток. Плоди – листівки чи горіхи.
Тичинок може бути різною кількістю. Найчастіше зустрічаються квітки з великим і невизначеним числом тичинок, деякі види тичинок можуть бути від 3 до 1.
калюжниця болотна - * Р 5 А ¥ G ¥
Залежно від типу плодів у с-ве лютикові виділяють дві підродини:
· зимівники (пологи купальниця, борець, жвавість), що мають плід листівку
· лютикові (пологи жовтець, вітряниця, василист), що мають плід горішок
У всіх рослин цього сімейства дуже поширені алкалоїди. Лютикові отруйні, вони є кормовими рослинами, але завдяки поширеності алкалоїдів рослини використовуються як лікарських засобів. У морфологічно неспеціалізованих видів (рід василист, калюжниця, жовтець) синтезуються прості алкалоїди, а у просунутих і спеціалізованих пологів утворюються складні алкалоїди (рослини з роду аконіт і жвавість).
З різних видів лютиків у кардіологічній практиці використовують 20 алкалоїдів. Алкалоїди всіх видів лютикових отруйні, найотруйніші належать с-ву аконітові.
Горицвіт весняний, живокість сітчастоплідна, борець (або аконіт) - важливі лікарські рослини.
Алкалоїди весняного весілля роблять курареподібний ефект, використовуються для розслаблення м'язів.
Алкалоїди живокості сітчастоплідної використовується в нервовій клініці при захворюваннях, пов'язаних із розладом рухових функцій
Настоянка з трави джунгарського аконіту, що входить до складу препарату «Акофіт», рекомендується при радикулітах.
Рідкісні та охоронювані види: купальниця європейська, воронець колосистий, борець північний.
Порядок Макові.Сімейство макові (250)
Поширені рослини цього сімейства переважно у тропічних і субтропічних областях. У помірній зоні трапляються рідко. На території Смоленської області 2 види – чистотіл великий та мак-самосейка.
Макові трав'янисті багаторічні, іноді однорічні рослини. Наявність однорічників вважається еволюційно молодою ознакою. Листорозташування чергове, листя просте, без прилистків, може бути як цілісним, так і розсіченим.
Квітки маку дуже часто великі, актиноморфні:
* Са 2 З 2 +2 А ¥ G ( ¥ )
Як правило, чашолистки опадають під час розпускання квітки. Гінецей складається з декількох плодолистків, які зростаються між собою, формуючи паракарпну гінецю. Плід – суха коробочка, покрита зубчиками. Насіння зачатків багато, плоди дрібні, як запасний матеріал містять жири.
Яскравою анатомічною та біохімічною особливістю макових є наявність млечників, у яких синтезується млечний сік складної будови. До його складу входять різні алкалоїди ізохінолінового ряду. Чумацький сік та алкалоїди макових широко використовуються в медицині.
Особливого значення набув рід мак, вид мак снодійний – основна сировина для отримання опію. Опій містить понад 20 алкалоїдів, серед яких морфін, кодеїн, наркотин, папаверин. Частина алкалоїдів має потужну анестезуючу дію, але викликає звикання. Папаверин використовується як спазмолітик при стенокардії та бронхіальній астмі. У нашій країні мак снодійний не вирощується. Деяка кількість наркотичних алкалоїдів міститься також у олійних та декоративних сортах маку снодійного.
Чистотіл великий
Квітки дрібні, зібрані в зонтикоподібне суцвіття: * Са 2 З 2+2 А ¥ G (¥)
Його помаранчевий чумацький сік поряд з алкалоїдами містить флавоноїди, дубильні речовини, сапоніни, органічні кислоти, вітаміни. Трава чистотілу використовується як жовчогінний та антибактерицидний засіб при захворюваннях печінки та жовчного міхура. Чистотіл великий – отруйна рослина.
З трави чистотілу виготовляють препарат «Сангвірітрін» для полоскання рота та горла при їх запаленні, а також для промивання гнійних ран.
З лікарськими цілями використовують мачок жовтий, із нього отримують ненаркотичний протикашльовий засіб глауцину гідрохлорид. Також з лікарськими цілями використовують маклію дрібноплідну та маклію серцеподібну.
ЗАНЯТТЯ 28: Сімейство розоцвіте (Rosaceae).
1. Сімейство розоцвіте, загальна характеристика.
2. Підродина спірейні. Підродина рожеві.
3. Підродина яблоневі. Підродина сливові.
1. Сімейство розоцвіті (3500)
Представники цього сімейства поширені поза тропічними областями Північної півкулі та зустрічаються в різних фітоценозах. На Смоленщині зростає 56 видів розоцвітих. Життєва форма: дерева, чагарники, трави (однолітні та багаторічні).
Листя як просте (цілісні або розсічені) так і складне (парно і непарноперисте, трійчасте), дуже часто є прилистки. Листорозташування чергове, рідко супротивне. У деяких видів трапляються шипи (метаморфози епідермісу).
Біохімічні особливості розоцвітих:
· Відсутній неспеціалізований вторинний метаболізм. Це проявляється у повній відсутності алкалоїдів та токсинів. Ефірні олії утворюються рідко, і вони завжди мають простий склад і будову (виключення роду Роза)
Будова квіток рожевих не відрізняється високою спеціалізацією до комахозапилення. Квітки поодинокі або зібрані у суцвіття різних типів. Квітки актиноморфні, оцвітина складна - чашолистків і пелюсток зазвичай п'ять. Досить часто квітка має подвійну кількість чашолистків, у цьому випадку перше коло чашолистків утворює підчаші. Тичинок багато, у 2-4 рази більше, ніж пелюсток. Число плодолистків або невизначене, або суворо фіксоване.
Характерна риса квітки – наявність гіпантія- Особливої освіти, що сформувався з квітлоложа, що розрісся, і зрослих з ним підстав чашолистків, пелюсток і тичинок. Форма гіпантія може бути опуклою, блюдцеподібною, увігнутою. Гіпантій захищає гінець від факторів зовнішнього середовища. Водночас він відіграє важливу роль при утворенні плодів. Він може розростатися та брати участь у формуванні мезокарпію та ендокарпію. Так соковита м'якоть шипшини, яблуні, груші, сливи є гіпантієм, що розрісся. Крім того, гіпантій забезпечує зв'язок з тваринами - розповсюджувачами плодів (екзо-і-ендохорія).
Плоди розоцвітих дуже різноманітні за будовою, що забезпечує різноманітність способів розповсюдження. Удосконалення методів поширення – це з основних напрямів еволюції всередині сімейства.
Дуже часто у розоцвітих утворюються помилкові плоди (яблуко у яблуні та яблучко у горобини) та складні плоди (багатогорінок у суниці та багатокістянка у малини).
Сімейство розоцвіті включає чотири підродини: спірейні, рожеві, яблуневі, сливові.
2. Підродина спірейні. Підродина рожеві.
Підродина спірейна (180)
Це найпримітивніша підсімейство, представлене чагарниками, рідше деревами і дуже рідко багаторічними кореневищними травами. Квітки дрібні, зібрані або в мітлу, або в кистевидні суцвіття, або в щитки. Пелюстки віночка білі, рідше – світло-рожеві. Для представників цього сімейства характерно увігнуте або майже плоске квітколожі (гіпантій), а плід сухий - багатолистівка (п'ятлистівка). Гінецей апокарпний, зазвичай із 2-5 плодолистків.
До сімейства входять пологи Спірея, Таволга, Рябінник.
Центральним родом є рід Спірейні, поширений у Сибіру та Далекому Сході. Спірейні - це чагарники з простим листям, без прилистків. Квітки зазвичай білі або рожеві, в хуртових, щитковидних або зонтикоподібних суцвіттях. Плід – багатолистівка.
Рід таволга, вид таволга в'язлистий. Це велика багаторічна рослина висотою до 2м, з коротким кореневищем. Листя уривчасто-перисте, зверху темно-зелене, знизу з тонким білим опушенням. При розтиранні листя видає різкий запах. Квітки жовтувато-білі зібрані в густе суцвіття. Плоди – однонасінні листівки. Таволга в'язолиста здавна застосовується в народній медицині як в'яжучий засіб.
* Са (5) З 5 А ¥ G 6-10
Підродина Рожеві (1.700)
Трав'янисті рослини широко поширені в різних фітоценозі від тундрів до гірських тропіків. Квітки зібрані в щиткоподібні або кистевидні дихазії, рідше поодинокі.
шипшина собача - * Са (5) Co 5 А ¥ G ¥
Гінецей апокарпний, полімерний, але у зав'язі завжди одна, рідше дві сім'япочки.
Плоди різноманітні, але найчастіше це збірна кістянка та багатогорішки. Багато видів цього сімейства зустрічаються соковиті складні плоди, у формуванні яких бере участь гіпантій.
Для багатьох рослин цього сімейства характерне розмноження за допомогою кореневищ, столонів або вусів.
Речовини вторинного рожевого метаболізму використовуються як лікарські засоби.
1. малина звичайна - її плоди містять значну кількість саліцилової кислоти, що обумовлює їх потогінну і жарознижувальну дію при простудних захворюваннях.
2. кровохлібка лікарська - квітка не має віночка, філіжанка пурпурова 4хроздільна. Відвар кореневища використовується як кровоспинний засіб при маткових та гемороїдальних кровотечах.
3. перстач прямостоячий (узик, калган). відрізняється 4-членною оцвітиною і чашкою з підчахтям. Настойка калгану (25г кореневища на 500мл горілки) використовується при катарах кишківника та дизентерії.
4. шабельник болотний (народна назва – декоп) – відвар кореневища застосовується при хворобах суглобів та жовтяниці. Також застосовується як в'яжучий, кровоспинний і потогінний засіб.
5. шипшина корична – містить велику кількість вітаміну «С», а також вітаміни «В 2», «К», каротин та лимонну кислоту. Згущений водний екстракт шипшини з цукром (холосас) використовується як жовчогінний препарат.
3. Підродина яблоневі. Підродина сливові.
 ВІДДІЛ ЧЕРВОНИЙ ВОДОРОС майже виключно морські водорості. життєвий цикл – повна відсутність джгутикових стадій особлива форма оогамного статевого процесу. 2
ВІДДІЛ ЧЕРВОНИЙ ВОДОРОС майже виключно морські водорості. життєвий цикл – повна відсутність джгутикових стадій особлива форма оогамного статевого процесу. 2
 Пігменти хлорофіл «а» і «в» , каротиноїди (каротин, зеаксантин, антераксантин, криптоксантин, лютеїн, неоксантин) фікобіліни: фікоеритрини (червоний) ) 3
Пігменти хлорофіл «а» і «в» , каротиноїди (каротин, зеаксантин, антераксантин, криптоксантин, лютеїн, неоксантин) фікобіліни: фікоеритрини (червоний) ) 3
 Хлоропласти оболонка з двох мембран поодинокі тилакоїди, на поверхні фікобілісоми. Запасний продукт - полісахарид «червоний крохмаль», що від йоду набуває буро-червоного кольору. Зерна багрянкового крохмалю відкладаються в цитоплазмі завжди поза зв'язком з піреноїдами та хроматофорами. 4
Хлоропласти оболонка з двох мембран поодинокі тилакоїди, на поверхні фікобілісоми. Запасний продукт - полісахарид «червоний крохмаль», що від йоду набуває буро-червоного кольору. Зерна багрянкового крохмалю відкладаються в цитоплазмі завжди поза зв'язком з піреноїдами та хроматофорами. 4
 Будова талому одноклітинні кокоїдні форми (порфіридіум) гетеротрихальні та у вигляді розгалужених ниток, прикріплених до субстрату за допомогою різоїдів. псевдопаренхіматозні таломи, переплетення бічних гілок Пластинчасті таломи паренхіматозної будови (порфіру). 5
Будова талому одноклітинні кокоїдні форми (порфіридіум) гетеротрихальні та у вигляді розгалужених ниток, прикріплених до субстрату за допомогою різоїдів. псевдопаренхіматозні таломи, переплетення бічних гілок Пластинчасті таломи паренхіматозної будови (порфіру). 5
 Клітина одягнена оболонкою пектин – геміцелюлозні компоненти набухають у стінках відкладається вапно. Клітини одно- та багатоядерні Хроматофори парієтальні, численні, у вигляді зерен або платівок. 6
Клітина одягнена оболонкою пектин – геміцелюлозні компоненти набухають у стінках відкладається вапно. Клітини одно- та багатоядерні Хроматофори парієтальні, численні, у вигляді зерен або платівок. 6
 Безстатеве розмноження за допомогою нерухомих клітин моноспори – одна в спорангії Тетраспори – чотирьох Тетраспори – на диплоїдних безстатевих рослинах – спорофітах. У спорангіях перед освітою тетраспор – мейоз. 7
Безстатеве розмноження за допомогою нерухомих клітин моноспори – одна в спорангії Тетраспори – чотирьох Тетраспори – на диплоїдних безстатевих рослинах – спорофітах. У спорангіях перед освітою тетраспор – мейоз. 7
 Половий процес Оогамний Жіночий орган - карпогон у більшості - з розширеної базальної частини - черевця - яйцеклітина, та відростка - трихогіни. Карпогон розвивається на особливій короткій корпогоніальній гілки. 8
Половий процес Оогамний Жіночий орган - карпогон у більшості - з розширеної базальної частини - черевця - яйцеклітина, та відростка - трихогіни. Карпогон розвивається на особливій короткій корпогоніальній гілки. 8
 Антеридії - дрібні безбарвні клітини містять безжгутикові спермації пасивно переносяться струмами води і прилипають до трихогіну. У місці контакту спермація і трихогіни їх стінки розчиняються.
Антеридії - дрібні безбарвні клітини містять безжгутикові спермації пасивно переносяться струмами води і прилипають до трихогіну. У місці контакту спермація і трихогіни їх стінки розчиняються.
 освіта карпоспор Після запліднення базальна частина карпогону відокремлюється перегородкою від трихогіни – відмирає
освіта карпоспор Після запліднення базальна частина карпогону відокремлюється перегородкою від трихогіни – відмирає
 освіта карпоспор 2. з заплідненого карпогону виростають гілкуючі нитки - гонимобласти, клітини їх перетворюються на карпоспорангії, що виробляють по одній карпоспорі. 11
освіта карпоспор 2. з заплідненого карпогону виростають гілкуючі нитки - гонимобласти, клітини їх перетворюються на карпоспорангії, що виробляють по одній карпоспорі. 11
 утворення карпоспор у більшості гонимобласти розвиваються не безпосередньо з черевця заплідненого карпогону з а у к с і л я р н их клітин. можуть бути віддалені від карпогону або розташовуються на таломі в безпосередній близькості коли ауксилярні клітини віддалені від карпогону, з його черевця після запліднення виростають з'їдн е т е л н е, або про блас т е м н е, нитки. 12
утворення карпоспор у більшості гонимобласти розвиваються не безпосередньо з черевця заплідненого карпогону з а у к с і л я р н их клітин. можуть бути віддалені від карпогону або розташовуються на таломі в безпосередній близькості коли ауксилярні клітини віддалені від карпогону, з його черевця після запліднення виростають з'їдн е т е л н е, або про блас т е м н е, нитки. 12
 клітини ообластемних ниток містять диплоїдні ядра. Ообластемні нитки підростають до ауксилярних клітин, у точці контакту оболонки розчиняються і між клітиною ообластемної нитки та ауксилярною клітиною встановлюється повідомлення. Це злиття клітин не супроводжується злиттям їх ядер (диплоїдного ядра клітини ообластемної нитки та ауксилярної гаплоїдної ядра). злиття з ауксилярною клітиною стимулює поділ диплоїдного ядра клітини ообластемної нитки та розвиток гонімобластів містять диплоїдні ядра та продукують диплоїдні карпоспори. Гонімобласти - особлива генерація - 13
клітини ообластемних ниток містять диплоїдні ядра. Ообластемні нитки підростають до ауксилярних клітин, у точці контакту оболонки розчиняються і між клітиною ообластемної нитки та ауксилярною клітиною встановлюється повідомлення. Це злиття клітин не супроводжується злиттям їх ядер (диплоїдного ядра клітини ообластемної нитки та ауксилярної гаплоїдної ядра). злиття з ауксилярною клітиною стимулює поділ диплоїдного ядра клітини ообластемної нитки та розвиток гонімобластів містять диплоїдні ядра та продукують диплоїдні карпоспори. Гонімобласти - особлива генерація - 13
 У найбільш високоорганізованих червоних водоростей ауксилярні клітини розвиваються лише після запліднення карпогону і в безпосередній близькості від нього. Сукупність ауксилярної клітини (або клітин) з коропогоном носить спеціальну назву прокарпію. Тут немає потреби в утворенні довгих ообластемних ниток, що зв'язують карпогон і ауксилярні клітини, ауксилярна клітина просто зливається з черевцем заплідненого карпогону, після чого з неї розвиваються гонімобласти з карпоспорами. Карпоспорангії . 14
У найбільш високоорганізованих червоних водоростей ауксилярні клітини розвиваються лише після запліднення карпогону і в безпосередній близькості від нього. Сукупність ауксилярної клітини (або клітин) з коропогоном носить спеціальну назву прокарпію. Тут немає потреби в утворенні довгих ообластемних ниток, що зв'язують карпогон і ауксилярні клітини, ауксилярна клітина просто зливається з черевцем заплідненого карпогону, після чого з неї розвиваються гонімобласти з карпоспорами. Карпоспорангії . 14
 15
15

 КЛАС БАНГІЄВІ Клітини часто із зірчастим хроматофором та піреноїдом. Пори між клітинами, як правило, відсутні. Карпогон без трихогіни, після запліднення вміст карпогон безпосередньо ділиться з утворенням карпоспор. Безстатеве розмноження - моноспорами 17
КЛАС БАНГІЄВІ Клітини часто із зірчастим хроматофором та піреноїдом. Пори між клітинами, як правило, відсутні. Карпогон без трихогіни, після запліднення вміст карпогон безпосередньо ділиться з утворенням карпоспор. Безстатеве розмноження - моноспорами 17

 19
19
 род компсопогон Прісноводний широко поширений у тропіках занесена з акваріумними рослинами Таллом гетеротрихальний Розмноження моноспорами. Моноспорангії відчленовуються косо ідучою перегородкою від будь-якої клітини талому і мають зернистий від великої кількості багрянкового крохмалю вміст, який перетворюється на одну нерухому голу моноспору 20
род компсопогон Прісноводний широко поширений у тропіках занесена з акваріумними рослинами Таллом гетеротрихальний Розмноження моноспорами. Моноспорангії відчленовуються косо ідучою перегородкою від будь-якої клітини талому і мають зернистий від великої кількості багрянкового крохмалю вміст, який перетворюється на одну нерухому голу моноспору 20
 21
21

 КЛАС ФЛОРИДЕЇ Клітини переважно з парієтальними хроматофорами без піреноїдів Між клітинами є пори. Карпогон із трихогіною. Після запліднення розвиваються гонимобласти або безпосередньо з черевця заплідненого карпогону, або ауксилярних клітин після злиття їх з ообластемними нитками. Безстатеве розмноження більшості-тетраспорами. Гонімобласти, що несуть карпоспорангії, розглядаються як особлива генерація – карпоспорофіт. Деталі розвитку карпоспорофіту (з заплідненого карпогону або з ауксилярних клітин), а також час диференціювання ауксилярних клітин (до або після запліднення), їхнє становище на таломі тощо кладуться в основу поділу флоридів на шість порядків.
КЛАС ФЛОРИДЕЇ Клітини переважно з парієтальними хроматофорами без піреноїдів Між клітинами є пори. Карпогон із трихогіною. Після запліднення розвиваються гонимобласти або безпосередньо з черевця заплідненого карпогону, або ауксилярних клітин після злиття їх з ообластемними нитками. Безстатеве розмноження більшості-тетраспорами. Гонімобласти, що несуть карпоспорангії, розглядаються як особлива генерація – карпоспорофіт. Деталі розвитку карпоспорофіту (з заплідненого карпогону або з ауксилярних клітин), а також час диференціювання ауксилярних клітин (до або після запліднення), їхнє становище на таломі тощо кладуться в основу поділу флоридів на шість порядків.
 Порядок немаліонові Для представників характерна відсутність ауксилярних клітин гонимобласти розвиваються безпосередньо з заплідненого коропогону.
Порядок немаліонові Для представників характерна відсутність ауксилярних клітин гонимобласти розвиваються безпосередньо з заплідненого коропогону.



 Рід Лемане зустрічається в швидко поточних річках з холодною водою. Таллом, що має вигляд щетини, що не гілкується, 10-15 см завдовжки і 1 мм товщиною, темно-фіолетового або оливково бурого кольору з вузлуватими здуттями. Прикріплюється до субстрату за допомогою підошви з ниток, що стелиться. По осі талому проходить одна нитка з витягнутих безбарвних клітин. Від верхньої частини кожної клітини центральної осі відходить мутовка здебільшого з чотирьох гілок, що розходяться по радіусах. Базальні клітини кожної такої гілки великі, витягнуті. Від їх дистального (морфологічно верх нього) кінця відходять гілки другого порядку, які у свою чергу багаторазово гілкуються. Кінцеві розгалуження зростаються в багатошарову кору. Її зовнішні клітини дрібні та заповнені хроматофорами.
Рід Лемане зустрічається в швидко поточних річках з холодною водою. Таллом, що має вигляд щетини, що не гілкується, 10-15 см завдовжки і 1 мм товщиною, темно-фіолетового або оливково бурого кольору з вузлуватими здуттями. Прикріплюється до субстрату за допомогою підошви з ниток, що стелиться. По осі талому проходить одна нитка з витягнутих безбарвних клітин. Від верхньої частини кожної клітини центральної осі відходить мутовка здебільшого з чотирьох гілок, що розходяться по радіусах. Базальні клітини кожної такої гілки великі, витягнуті. Від їх дистального (морфологічно верх нього) кінця відходять гілки другого порядку, які у свою чергу багаторазово гілкуються. Кінцеві розгалуження зростаються в багатошарову кору. Її зовнішні клітини дрібні та заповнені хроматофорами.
 Порядок криптонемії є ауксилярні клітини, які розвиваються до запліднення карпогону і розсіяні по талому на відомій відстані від коропогону. З карпогону після запліднення до ауксилярних клітин ростуть більш менш довгі багатоклітинні сполучні, або ообластемні, нитки. Після злиття ауксилярної клітини з клітиною ообластемної нитки і переходу диплоїдного ядра клітини нитки в ауксилярну клітину виникають гонімобласти. При утворенні тетраспор відбувається мейоз і гаплоїдні тетраспори проростають у гаплоїдні рослини-гаметофіти, що несуть статеві органи. Гаметофіт та тетраспорофіт зовні (морфологічно) не розрізняються. ізоморфна зміна генерацій, ускладнена диплоїдним карпоспорофптом. 29
Порядок криптонемії є ауксилярні клітини, які розвиваються до запліднення карпогону і розсіяні по талому на відомій відстані від коропогону. З карпогону після запліднення до ауксилярних клітин ростуть більш менш довгі багатоклітинні сполучні, або ообластемні, нитки. Після злиття ауксилярної клітини з клітиною ообластемної нитки і переходу диплоїдного ядра клітини нитки в ауксилярну клітину виникають гонімобласти. При утворенні тетраспор відбувається мейоз і гаплоїдні тетраспори проростають у гаплоїдні рослини-гаметофіти, що несуть статеві органи. Гаметофіт та тетраспорофіт зовні (морфологічно) не розрізняються. ізоморфна зміна генерацій, ускладнена диплоїдним карпоспорофптом. 29
 рід дюренеа поширений у південних морях. Таллом має вигляд розгалужених слизових кущів рожевого кольору. 30
рід дюренеа поширений у південних морях. Таллом має вигляд розгалужених слизових кущів рожевого кольору. 30
 31
31
 32
32
 33
33


 Порядок церамієві високо організовані багато видів ауксилярні клітини диференціюються тільки після того, як сталося запліднення карпогону Довгих ообластемних ниток не утворюється: черевце карпогону або безпосередньо зливається з розташованою ауксилярною клітиною (клітинами), або через короткі вирости, формуючи
Порядок церамієві високо організовані багато видів ауксилярні клітини диференціюються тільки після того, як сталося запліднення карпогону Довгих ообластемних ниток не утворюється: черевце карпогону або безпосередньо зливається з розташованою ауксилярною клітиною (клітинами), або через короткі вирости, формуючи


 еволюція у межах відділу Бангієві більш примітивні. у карпогон ще не виробилася типова форма, мало відрізняється від звичайних вегетативних клітин. Після запліднення вміст карпогон безпосередньо ділиться на карпоспори. Флорідеї - група, що більш еволюціонувала Карпогоном з трихогіною. З класу флоридей найпростіший порядок - немалионовые, які мають ауксилярних клітин, і гонимобласты, у яких розвиваються карпоспорангії, утворюються безпосередньо з черевної частини заплідненого карпогону. Криптонемієві – наступний крок прогресивної еволюції: мають ауксилярні клітини, що сприяють збільшенню продукції карпоспор, оскільки утворюється не один цистокарпій, а багато – за кількістю ауксилярних клітин. При розкиданих безладно по талому ауксилярних клітинах необхідні більш менш довгі ообластемні нитки. Найвищого ступеня еволюції досягли церамієві, у яких є прокарпій та ауксилярні клітини диференціюються лише після того, як сталося запліднення. Безпосереднє сусідство карпогону та ауксилярної клітини (клітин) у прокарпії полегшує утворення цпстокарпіїв. Цей порядок найбільш багатий на види. 39
еволюція у межах відділу Бангієві більш примітивні. у карпогон ще не виробилася типова форма, мало відрізняється від звичайних вегетативних клітин. Після запліднення вміст карпогон безпосередньо ділиться на карпоспори. Флорідеї - група, що більш еволюціонувала Карпогоном з трихогіною. З класу флоридей найпростіший порядок - немалионовые, які мають ауксилярних клітин, і гонимобласты, у яких розвиваються карпоспорангії, утворюються безпосередньо з черевної частини заплідненого карпогону. Криптонемієві – наступний крок прогресивної еволюції: мають ауксилярні клітини, що сприяють збільшенню продукції карпоспор, оскільки утворюється не один цистокарпій, а багато – за кількістю ауксилярних клітин. При розкиданих безладно по талому ауксилярних клітинах необхідні більш менш довгі ообластемні нитки. Найвищого ступеня еволюції досягли церамієві, у яких є прокарпій та ауксилярні клітини диференціюються лише після того, як сталося запліднення. Безпосереднє сусідство карпогону та ауксилярної клітини (клітин) у прокарпії полегшує утворення цпстокарпіїв. Цей порядок найбільш багатий на види. 39
 Найдавніша скам'янілість, що ідентифікується як червона водорість, є також найдавнішою скам'янілістю-еукаріотом, що належить до конкретного сучасного таксону. Bangiomorpha pubescens, скам'янілість багатоклітинного організму знайдена в арктичній Канаді, лише трохи відрізняється від сучасної червоної водорості роду Бангія (Bangia), незважаючи на те, що вона була відкладена в шарах 1, 2 мільярдної давності. 41
Найдавніша скам'янілість, що ідентифікується як червона водорість, є також найдавнішою скам'янілістю-еукаріотом, що належить до конкретного сучасного таксону. Bangiomorpha pubescens, скам'янілість багатоклітинного організму знайдена в арктичній Канаді, лише трохи відрізняється від сучасної червоної водорості роду Бангія (Bangia), незважаючи на те, що вона була відкладена в шарах 1, 2 мільярдної давності. 41
 ВІДДІЛ БУРІ ВОДОРОСЛІ РНАЄОРНУТА морські, особливо багато - в холодних водах північної та південної півкулі. По морфологічному та анатомічному диференціюванню талому - на вищому рівні, ніж раніше розглянуті групи водоростей. невідомі ні одноклітинні, ні колоніальні форми, ні таломи у вигляді простої нерозгалуженої нитки. найпростіші - гетеротрихальні таломи великі, хибно-або істиннотканинної будови.
ВІДДІЛ БУРІ ВОДОРОСЛІ РНАЄОРНУТА морські, особливо багато - в холодних водах північної та південної півкулі. По морфологічному та анатомічному диференціюванню талому - на вищому рівні, ніж раніше розглянуті групи водоростей. невідомі ні одноклітинні, ні колоніальні форми, ні таломи у вигляді простої нерозгалуженої нитки. найпростіші - гетеротрихальні таломи великі, хибно-або істиннотканинної будови.
 Кліткові стінки, що сильно ослизняються, одне ядро, одну або багато вакуолей зазвичай постенні хроматофори різної форми. хроматофори оточені складною системою мембран - у прямому зв'язку з оболонкою ядра - «хлоропластною ендоплазматичною мережею» Матрікс хлоропласту перетинається паралельними тритилакоїдними ламеллами
Кліткові стінки, що сильно ослизняються, одне ядро, одну або багато вакуолей зазвичай постенні хроматофори різної форми. хроматофори оточені складною системою мембран - у прямому зв'язку з оболонкою ядра - «хлоропластною ендоплазматичною мережею» Матрікс хлоропласту перетинається паралельними тритилакоїдними ламеллами
 Пігменти хроматофори пофарбовані в бурий колір хлорофілі «а» і «с» (хлорофіл «в» відсутня) в-каротин Багато бурих ксантофілів, особливо фукоксантину. 44
Пігменти хроматофори пофарбовані в бурий колір хлорофілі «а» і «с» (хлорофіл «в» відсутня) в-каротин Багато бурих ксантофілів, особливо фукоксантину. 44
 Запасний полісахарид - ламінарин спирт маніт жири відкладається поза хлоропласту в цитоплазмі. Монадні клітини (зооспори та гамети) мають вічко та джгутики. Око - частина пластиди, пов'язана зі джгутиковим апаратом. Джгутики – гетероморфні. 45
Запасний полісахарид - ламінарин спирт маніт жири відкладається поза хлоропласту в цитоплазмі. Монадні клітини (зооспори та гамети) мають вічко та джгутики. Око - частина пластиди, пов'язана зі джгутиковим апаратом. Джгутики – гетероморфні. 45


 Розмноження вегетативне, безстатеве та статеве. Вегетативне розмноження - ділянками талому. У деяких - спеціалізовані гілочки (висновкові бруньки), відламуються в нові таломи. Безстатеве розмноження - зооспори, що утворюються в одногніздних або однокамерних спорангіях на диплоїдних рослинах (спорофітах) перед формуванням зооспор. У найпростіших - статевий процес ізогамний, гамети розвиваються в багатогніздних або багатокамерних спорангіях У найбільш високоорганізованих бурих водоростей статевий процес оагамний. В оогоніях та антеридіях, як правило, утворюється по одній гаметі (яйцеклітині та сперматозоїду відповідно). Яйцеклітина запліднюється завжди поза оогонієм. Зигота без періоду спокою проростає у диплоїдну рослину.
Розмноження вегетативне, безстатеве та статеве. Вегетативне розмноження - ділянками талому. У деяких - спеціалізовані гілочки (висновкові бруньки), відламуються в нові таломи. Безстатеве розмноження - зооспори, що утворюються в одногніздних або однокамерних спорангіях на диплоїдних рослинах (спорофітах) перед формуванням зооспор. У найпростіших - статевий процес ізогамний, гамети розвиваються в багатогніздних або багатокамерних спорангіях У найбільш високоорганізованих бурих водоростей статевий процес оагамний. В оогоніях та антеридіях, як правило, утворюється по одній гаметі (яйцеклітині та сперматозоїду відповідно). Яйцеклітина запліднюється завжди поза оогонієм. Зигота без періоду спокою проростає у диплоїдну рослину.

 КЛАС ІЗОГЕНЕРАТНІ Порядок ектокарповий рід ектокарпус - найпримітивніші бурі водорості. Поширені у всіх морях, особливо холодних, і ростуть на підводних предметах та інших більших водоростях. 50
КЛАС ІЗОГЕНЕРАТНІ Порядок ектокарповий рід ектокарпус - найпримітивніші бурі водорості. Поширені у всіх морях, особливо холодних, і ростуть на підводних предметах та інших більших водоростях. 50
 51
51
 52
52

 54
54
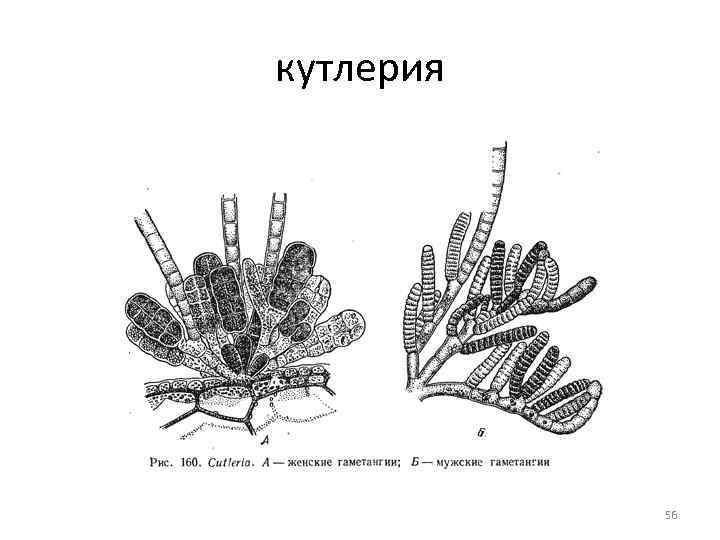
 Порядок кутлерієвий Порядок включає тільки два роди: кутлерія та занардинія Кутлерія поширена по всьому узбережжю Європи, занардинія – переважно у Середземному морі, а також у Чорному. Обидва роди виявляють чергування поколінь: у кутлерії зміна генерацій гетероморфна, у занардинії - ізоморфна 55
Порядок кутлерієвий Порядок включає тільки два роди: кутлерія та занардинія Кутлерія поширена по всьому узбережжю Європи, занардинія – переважно у Середземному морі, а також у Чорному. Обидва роди виявляють чергування поколінь: у кутлерії зміна генерацій гетероморфна, у занардинії - ізоморфна 55
 57
57
 58
58

 60
60

 62
62
 КЛАС ГЕТЕРОГЕНЕРАТНІ Порядок ламінарієвих Гаметофіти мало різняться між собою і представлені мікроскопічними, часто редукованими до кількох клітин нитчастими рослинками-заростками, які несуть статеві органи. На чоловічих гаметофітах утворюються антеридії у вигляді дрібних клітин, що розвиваються по одному сперматозоїду, на жіночих - оогоніях, у яких формується по одній яйцеклітині. Спорофіти різних пологів різко розрізняються і є найбільшими нижчими рослинами, що досягають у довжину 60- 100 м, зі значним морфологічним розчленуванням і складною анатомічною будовою. У більшості випадків спорофіти ламінарієвих розчленовуються на листоподібну пластинку, «ствол» та різозиди, за допомогою яких вся рослина прикріплюється до підводного каміння та скель. У місці переходу листової пластинки в стволик знаходиться інтеркалярна меристема, за рахунок діяльності якої наростає як листова пластинка, так і ствол. рід ламінарію, види якого широко поширені в північних морях. 63
КЛАС ГЕТЕРОГЕНЕРАТНІ Порядок ламінарієвих Гаметофіти мало різняться між собою і представлені мікроскопічними, часто редукованими до кількох клітин нитчастими рослинками-заростками, які несуть статеві органи. На чоловічих гаметофітах утворюються антеридії у вигляді дрібних клітин, що розвиваються по одному сперматозоїду, на жіночих - оогоніях, у яких формується по одній яйцеклітині. Спорофіти різних пологів різко розрізняються і є найбільшими нижчими рослинами, що досягають у довжину 60- 100 м, зі значним морфологічним розчленуванням і складною анатомічною будовою. У більшості випадків спорофіти ламінарієвих розчленовуються на листоподібну пластинку, «ствол» та різозиди, за допомогою яких вся рослина прикріплюється до підводного каміння та скель. У місці переходу листової пластинки в стволик знаходиться інтеркалярна меристема, за рахунок діяльності якої наростає як листова пластинка, так і ствол. рід ламінарію, види якого широко поширені в північних морях. 63
 КЛАС ЦИКЛОСПОРОВІ Порядок фукусові характеризується - верхівковим зростанням талому Відсутністю безстатевого розмноження оогамним статевим процесом статеві органи розташовуються в поглибленнях талому - концептакулах, або скафідіях.
КЛАС ЦИКЛОСПОРОВІ Порядок фукусові характеризується - верхівковим зростанням талому Відсутністю безстатевого розмноження оогамним статевим процесом статеві органи розташовуються в поглибленнях талому - концептакулах, або скафідіях.
 рід саргассум складне морфологічне розчленування талому широко поширений у південній півкулі, особливо у теплих морях. Стволик, прикріплений до субстрату підошвою гілки несе сплощені листоподібні утворення, кулясті повітряні міхури на спеціальних стеблинках розгалужені плодючі гілочки 71
рід саргассум складне морфологічне розчленування талому широко поширений у південній півкулі, особливо у теплих морях. Стволик, прикріплений до субстрату підошвою гілки несе сплощені листоподібні утворення, кулясті повітряні міхури на спеціальних стеблинках розгалужені плодючі гілочки 71



 Бурі водорості - природна група, яка за своєю морфологією добре відрізняється від інших відділів водоростей. Однак, як уже говорилося, за низкою ознак (наявністю хлорофілів «а» і «с», відсутністю хлорофілу «Ь», розташування тилакоїдів у хлоропластах - тритилакоїдні ламели, за подібними продуктами запасу - ламінарії, хризоламінарин, за будовою гетероморфними джгутиками) бурі водорості виявляють подібність із золотистими, жовтозеленими, діатомовими та пірофітовими водоростями. На цій підставі деякі автори зводять бурі та інші перераховані групи водоростей до рангу класів у межах великого відділу СЬгошор. Ьу1 а. Існування монадних клітин у життєвому циклі бурих водоростей дозволяє виводити їх, як і інші перелічені групи, з якихось первинних фотосинтезирующих жгутиконосцев з величезним переважанням бурих пігментів. Ці джгутикові еволюціонували кількома шляхами, одна з яких і привела до виникнення бурих водоростей. Однак у бурих водоростей зв'язок із джгутиковими більш віддалений, ніж у подібних з ними по ряду ознак золотистих, різножгутикових, діатомових, пірофітових, а також у зелених водоростей, тому що прямих перехідних форм тут немає. Серед бурих водоростей не простежується послідовний розвиток типів організації талому від монадного до нитчастого та пластинчастого, як це легко спостерігається у зелених, жовтозелених, золотистих та пірофйтбових водоростей. Як зазначалося, серед бурих водоростей представлені лише вищі щаблі морфологічної диференціювання талому - гетеротрихальная і пластинчаста. Можливо, простіші форми (монадні, кокоїдні, прості нитчасті) повністю втрачені протягом тривалої еволюції, яку проробили бурі водорості - давня група, відома вже; з відкладень силуру та девону. Щодо споріднених зв'язків порядків у межах відділу бурих водоростей та розташування їх у системі єдиного погляду немає. Відповідно до однієї зі схем філогенезу бурих водоростей, побудованої з урахуванням подібності будови талому на ранніх стадіях розвитку, бурі водорості - група дифілетична: одна лінія еволюції об'єднує (із згаданих у викладі) сфацелярієві, диктіотові та фукусові, інша - фукусові. Обидві 219" 75
Бурі водорості - природна група, яка за своєю морфологією добре відрізняється від інших відділів водоростей. Однак, як уже говорилося, за низкою ознак (наявністю хлорофілів «а» і «с», відсутністю хлорофілу «Ь», розташування тилакоїдів у хлоропластах - тритилакоїдні ламели, за подібними продуктами запасу - ламінарії, хризоламінарин, за будовою гетероморфними джгутиками) бурі водорості виявляють подібність із золотистими, жовтозеленими, діатомовими та пірофітовими водоростями. На цій підставі деякі автори зводять бурі та інші перераховані групи водоростей до рангу класів у межах великого відділу СЬгошор. Ьу1 а. Існування монадних клітин у життєвому циклі бурих водоростей дозволяє виводити їх, як і інші перелічені групи, з якихось первинних фотосинтезирующих жгутиконосцев з величезним переважанням бурих пігментів. Ці джгутикові еволюціонували кількома шляхами, одна з яких і привела до виникнення бурих водоростей. Однак у бурих водоростей зв'язок із джгутиковими більш віддалений, ніж у подібних з ними по ряду ознак золотистих, різножгутикових, діатомових, пірофітових, а також у зелених водоростей, тому що прямих перехідних форм тут немає. Серед бурих водоростей не простежується послідовний розвиток типів організації талому від монадного до нитчастого та пластинчастого, як це легко спостерігається у зелених, жовтозелених, золотистих та пірофйтбових водоростей. Як зазначалося, серед бурих водоростей представлені лише вищі щаблі морфологічної диференціювання талому - гетеротрихальная і пластинчаста. Можливо, простіші форми (монадні, кокоїдні, прості нитчасті) повністю втрачені протягом тривалої еволюції, яку проробили бурі водорості - давня група, відома вже; з відкладень силуру та девону. Щодо споріднених зв'язків порядків у межах відділу бурих водоростей та розташування їх у системі єдиного погляду немає. Відповідно до однієї зі схем філогенезу бурих водоростей, побудованої з урахуванням подібності будови талому на ранніх стадіях розвитку, бурі водорості - група дифілетична: одна лінія еволюції об'єднує (із згаданих у викладі) сфацелярієві, диктіотові та фукусові, інша - фукусові. Обидві 219" 75
 Групи порядків розрізняються і за способом зростання таломів: перша характеризується верхівковим зростанням, друга - інтеркалярним. Кілін (Н. Ку 1 ш), система якого найпоширеніша в даний час, від загального предка бурих водоростей виводить три еволюційні лінії, які представлені ним як класи ізогенератні, гетерогенератні та циклоспорові; в основу їх покладені відмінності у життєвих циклах та морфологічній будові водоростей. Великі бурі водорості знаходять велике застосування у господарстві людини. З них добувають альгін - клеючу речовину, що використовується в текстильній, харчовій та інших галузях промисловості. У приморських країнах викиди морських водоростей, багаті калієм і азотом, використовують як добрива, і навіть використовують для корм худобі. Деякі, наприклад, ламінарії («морська капуста»), їстівні. 76
Групи порядків розрізняються і за способом зростання таломів: перша характеризується верхівковим зростанням, друга - інтеркалярним. Кілін (Н. Ку 1 ш), система якого найпоширеніша в даний час, від загального предка бурих водоростей виводить три еволюційні лінії, які представлені ним як класи ізогенератні, гетерогенератні та циклоспорові; в основу їх покладені відмінності у життєвих циклах та морфологічній будові водоростей. Великі бурі водорості знаходять велике застосування у господарстві людини. З них добувають альгін - клеючу речовину, що використовується в текстильній, харчовій та інших галузях промисловості. У приморських країнах викиди морських водоростей, багаті калієм і азотом, використовують як добрива, і навіть використовують для корм худобі. Деякі, наприклад, ламінарії («морська капуста»), їстівні. 76
На великих глибинах, до 250 метрів, зростають червоні водорості, інакше звані багрянками. У поєднанні з коралами та яскравими рибами різноманітно забарвлені багрянки створюють неповторну красу підводного світу. Це переважно великі водорості, але, наприклад, до класу бангієвих належать і мікроскопічні червоні водорості.
Чому червоні водорості можуть зростати на такій значній глибині? Це питання було поставлено в ЄДІ з біології. Водорості дозволяє рости на великій глибині червоний пігмент фікоеритрин. Завдяки йому при фотосинтезі червоні водорості поглинають зелені, блакитні, синьо-фіолетові промені спектра. Саме це проміння, на відміну червоних, здатні проникнути глибоко в товщу води.
Для червоних водоростей характерно спорове безстатеве розмноження, а також статеве розмноження (оогамія), іноді зустрічається і вегетативне розмноження частинами талому.
До червоних водоростей відносяться філофора, порфіра, грацилярія, птилота, хондрус, всього їх налічується близько п'яти тисяч видів.
Порфіра- Плоска і тонка овальна пластинка до півметра в діаметрі. Для неї характерне лише статеве розмноження. Чоловічі статеві клітини не мають джгутиків (спермії). Це і зрозуміло, тому що на великій глибині під товщею води складно пересуватися за допомогою джгутика.
Значення водоростей
Місце в ланцюгах харчування, вплив на природу
1. Величезна маса водоростей створює фітопланктон, причому навіть у арктичних морях налічується 20-30 млн особин на 1 кубічний метр води. Це первинна продукція, що є основою харчового ланцюжка.
2. Фітопланктон є їжею для зоопланктону (вторинна продукція), який поїдається великими морськими мешканцями, наприклад, китами. Цікаво, що Тур Хейєрдал під час експедиції на плоту «Кон-Тікі» спробував «суп» із планктону і знайшов його досить смачним та поживним.
3. Донні водорості дають притулок рибам і різноманітним морським тваринам і є їжею їм. Ламінарію, наприклад, із задоволенням поїдає морський їжак.
4. Водорості насичують океани та атмосферу киснем.
5. Однак при масовому розмноженні водоростей (наприклад, хламідомонади) у пору так званого «цвітіння» води вміст кисню в ній падає і вода насичується токсинами. Від нестачі кисню гинуть річкові жителі.
Значення для людини
1. Водорості у багатьох регіонах тисячі років вживаються для харчування. Особливо широкою популярністю користуються такі бурі та червоні водорості, як ламінарія, ундарія, порфіра, хідзики (і взагалі саргасові).
2. Червоні водорості - джерело йоду, особливо деякі багаті на нього види.
3. Також червоні водорості є джерелом агар-агару - гелеутворюючої речовини, яка використовується в кондитерській промисловості, при культивуванні бактерій та ін.
4. За допомогою водоростей проводиться очищення стічних вод від фтору, азоту та ін., а також повітря від вуглекислого газу (у цьому досягли успіху хламідомонаду, хлорела, евглену).
5. Водорості є харчовими добавками: спіруліна, ламінарія, фукус, ульва, хлорела та інші.
Назва відділу походить від грецького слова rhodon("Родон") -рожевий. Забарвлення червоних водоростей обумовлене різним поєднанням пігментів. Вона буває від сірого та фіолетового до майже чорного кольору, а також включає всі відтінки червоного та рожевого. Червоні водорості, що мешкають у сильно освітлених місцях, пофарбовані в жовтий, коричневий або чорний колір через присутність у клітинах великої кількості каротиноїдів. Поєднує одноклітинні, колоніальні та багатоклітинні організми з кокоїдним, нитчастим, псевдопаренхіматозним та паренхіматозним типами структури тіла. Для всіх представників характерна повна відсутність джгутикових стадій у життєвому циклі. Форма талому різноманітна: нитчаста, кущиста, кіркоподібна, пластинчаста, міхурова, мішковидна та ін. Мешкають головним чином у морях і океанах (зазвичай прикріплені форми), рідше зустрічаються в прісних водах. Відомо близько чотирьох тисяч видів.
Будова клітини. Клітина у червоних водоростей еукаріотна: має оформлені органели з власними мембранами: ядро, мітохондрії, пластиди, апарат Гольджі та інші. У клітинах червоних водоростей зустрічається від однієї до багатьох пластид. Між клітинами є порові сполуки. Крім зовнішньої клітинної мембрани, клітини родофіт мають особливу освіту – клітинну стінку.
Ядро найчастіше одне, але є багатоядерні представники. Ядра червоних водоростей дрібні. Для деяких видів відома ендоредуплікація, або реплікація ядерного геному без мітозу. Вона призводить до поліплоїдії, або кратного збільшення набору хромосом у клітинах. Мітохондрії із сплощеними христами.
Хлоропласти червоних водоростей різної форми, вони зазвичай розташовані вздовж клітинних стінок, або парієтально. Ці пластиди в основному дископодібні та стрічкоподібні. Форма хлоропластів може змінюватись залежно від віку водорості. Кожен хлоропласт оточений власною двомембранною оболонкою, хлоропластна ендоплазматична мережа відсутня. Тилакоїди в хлоропластах поодинокі, не зібрані в групи та лежать на однаковій відстані один від одного. Один або два тилакоїди розташовані зазвичай по периферії хлоропласту паралельно його оболонці. Хлоропластна ДНК є у вигляді дрібних нуклеоїдів, розкиданих по всій стромі хлоропласту. Кожен нуклеоїд містить кілька кільцевих молекул хлоропластної ДНК.
З хлорофілів у пластидах червоних водоростей присутній тільки хлорофіл а, який маскується додатковими пігментами – фікобілінами: червоним фікоеритрином, синіми фікоціаніном та аллофікоціаніном. Ці фікобіліни локалізовані на поверхні тилакоїдів у особливих утвореннях – фікобілісомах напівсферичної та напівдископодібної форми.
Клітинна стінка складається із структурної фібрилярної фракції та аморфного матриксу. Міцність клітинної стінки надають фібрили з целюлози, які у червоних водоростей утворюють неправильну мережу. Найбільш важливими речовинами полісахаридного аморфного матриксу є агар, агароїди, карагінан. Ці речовини синтезуються у цистернах апарату Гольджі, потім транспортуються до клітинної поверхні та вбудовуються у стінку. Їх частку припадає до 70 % сухої ваги клітинної стінки. У ряду червоних водоростей поверх клітинної стінки може розташовуватися кутикула, що складається з білка. Серед багрянок є види з обвапненими оболонками: у них відкладається кальцит або арагоніт. Інкрустуватися може частина талому або весь талом.
Загальна характеристика.
Майже всі червоні водорості є фототрофами та будують своє тіло за допомогою фотосинтезу. Продукти фотосинтезу – особливий багрянцевий крохмаль, який відкладається у цитоплазмі, а не в хлоропласті, як у зелених водоростей. Багрянцевий крохмаль дає з йодом яскраво-червоне фарбування. Важливий запасний продукт - низькомолекулярний вуглеводень флоридозід. Його вміст у таломах деяких представників може перевищувати 10% від величини сухої ваги. Він виконує осморегуляторну функцію. Крім червоних водоростей, флоридозид зустрічається у ціанобактерій та криптомонад. Його концентрація у клітинах збільшується зі збільшенням солоності середовища. Деякі багрянки можуть запасати багатоатомні спирти.
Більшість видів червоних водоростей - багатоклітинні складовлаштовані організми, розміри яких можуть досягати 1-2 метрів і лише примітивні представники мають одноклітинну або колоніальну будову (рис. 17). Зустрічаються як однорічні, і багаторічні види, вік яких зазвичай становить 3–6 років. Форма тіла багрянок дуже
Рис. 17 Зовнішній вигляд червоних водоростей: А – одноклітинна водорість Порфірідіум; Б – багатоклітинна водорість Делессерія
різноманітна. Вона буває: ниткоподібна (волосоподібна або груба), пластинчаста цільна або складно розсічена з виростами по краю, циліндрична, кіркоподібна (кірки, плівки, притиснуті до субстрату), коралоподібна. Різноманітність зовнішніх форм червоних водоростей зводиться до кількох типів диференціації талому: кокоїдний, нитчастий, різнонітчастий, ложнотканинний і тканинний. Прикріплюються таломи родофіт різозій або підошвою.
Найбільш складно влаштовані таломи Флорідеєвих водоростей. Їхні слані мають ознаки тканинного диференціювання зі спеціалізацією клітин. У їхньому таломі можна розрізнити: кору, що складається з декількох шарів інтенсивно забарвлених клітин; серцевину, що складається з безбарвних клітин, часто зібраних у нитки. Серцевина виконує як транспортну функцію, а й механічну, оскільки у ній перебувають нитки з товстими поздовжніми стінками. Між корою та серцевиною у багатьох червоних водоростей може бути проміжний шар із великих безбарвних клітин. Зростання слані найчастіше інтеркалярний (вставковий) та апікальний (верхівковий), рідше базальний.
Розмноження.
У червоних водоростей зустрічається три способи розмноження: вегетативне, безстатеве і статеве.
Безстатеве розмноженняздійснюється за допомогою різних суперечок. Спори можуть розташовуватися в спорангії по одній, дві та чотири; вони називаються відповідно моноспори, біспори та тетраспори. Тетраспори можуть розташовуватися в тетраспорангіях по-різному: один над одним – лінійно, або зонально, хрестоподібно та в кутах тетраедра (рис. 18).

Рис. 18. Типи розташування тетраспор у тетраспорангіях (за: С. Ноєк van den et al., 1995): A– хрестоподібне; Б- хрестоподібне з поворотом; В- Лінійне; Г – тетраедральне
Статеве відтворенняу червоних водоростей оогамне, воно має низку особливостей, що не зустрічаються в інших груп водоростей. Чоловічі статеві клітини спермації, позбавлені джгутиків і пасивно зі струмом води переносяться до жіночих статевих органів. коропогонам. Карпогон червоних водоростей має вигляд колби і складається з розширеної нижньої частини (черевця) та витягнутої верхньої – трихогіни, яка служить для уловлювання спермацій. Більшість червоних водоростей карпогон формується на кінці короткої гілки, званої карпогонной гілкою. Клітина, що дає початок карпогонної гілки, називається підтримуючою.
Спермації формуються по одному в сперматангії, які у свою чергу утворюються на материнських клітинах. Зрілий спермацій одноядерний і позбавлений жорсткої клітинної стінки, оточений слизом, може містити хлоропласти. Спермацій пасивно переноситься струмами води, що контактує з трихогіною, яка розташована над поверхнею жіночого гаметофіту. Стінки сперматіїв та трихогінів у місці контакту розчиняються, чоловіче ядро проходить через центральний канал у трихогіну і зливається з гаплоїдним ядром карпогону. Подальший розвиток видів різних порядків має свої особливості.
Життєвий цикл.
У більшості Rhodophyta після запліднення жіночих гамет – карпогонів чоловічими гаметами – спермаціями із зиготи після багаторазових клітинних злиттів виникає багатоклітинне нитчасто-паренхімне утворення – карпоспорофіт (рис. 19). Він представляє

Рис. 19. Життєвий цикл Батрахоспермуму(по: R. E. Lee, 1999)
собою гонимобласт,у якому розвиваються диплоїдні клітини – суперечки статевого розмноження, або карпоспори, що проростають у нову диплоїдну рослину – спорофіт. Гонімобласт,або цистокарп – це комплекс плаценти, від якої відходять нитки гонімобласту з карпоспорангіями на закінченнях та покривів гонимобласту – обгортки. У гонимобластах всі клітини мають диплоїдний набір хромосом.
У ряду Флорідеєвих гонімобластів оточує оболонка. У такому разі ця структура називається цистокарп (рис. 20). У карпоспорангіях

Рис. 20. Життєвий цикл Полісифонії(по: R. E. Lee, 1999)
утворюються карпоспори, які проростають у диплоїдний тетраспорофіт. На тетраспорофіті в тетраспорангіях відбувається редукційний поділ. Гаплоїдні тетраспори проростають у гаплоїдний гаметофіт. Цей життєвий цикл із чергуванням поколінь: одного гаплоїдного – гаметофіт та двох диплоїдних – карпоспорофіт та тетраспорофіт – зустрічається у більшості червоних водоростей, але існує ряд відхилень від нього, залежно від варіабельності умов у навколишньому середовищі.
Життєвий цикл який завжди жорстко закріплений. Якщо немає умов для розвитку однієї з фаз, щоразу, поки не зміняться фактори середовища, відновлюється та сама фаза – або гаметофіт, або спорофіт. Такий цикл називають гетероморфним із нерегулярною зміною форм розвитку. Він наголошується у Мастокарпових. Існують ізоморфні цикли з регулярною зміною форм розвитку, коли статева і безстатева форми представлені зовні однаковими самостійними вільноживучими генераціями. Такий цикл розвитку характерний для Грацилярії, Хондруса, Мазели.
Систематика
Відділ Rhodophyta традиційно ділять на два класи: Бангієві – Bangiophyceae та Флорідеєві – Florideophyceae. До останнього класу належить більшість пологів та видів багрянок.
 Дві голови та шість ніг; чотири ходять, а дві смиренно лежать
Дві голови та шість ніг; чотири ходять, а дві смиренно лежать Самооцінка – що це таке: поняття, структура, види та рівні
Самооцінка – що це таке: поняття, структура, види та рівні Шлях касандри, або пригоди з макаронами Війна на землі та під землею
Шлях касандри, або пригоди з макаронами Війна на землі та під землею